
Fe3O4 Nanoparticles to Optimize the Co-Digestion of Vinasse, Filter Cake, and Deacetylation Liquor: Operational Aspects and Microbiological Routes


Abstract
1. Introduction
2. Materials and Methods
2.1. Residues and Inoculum
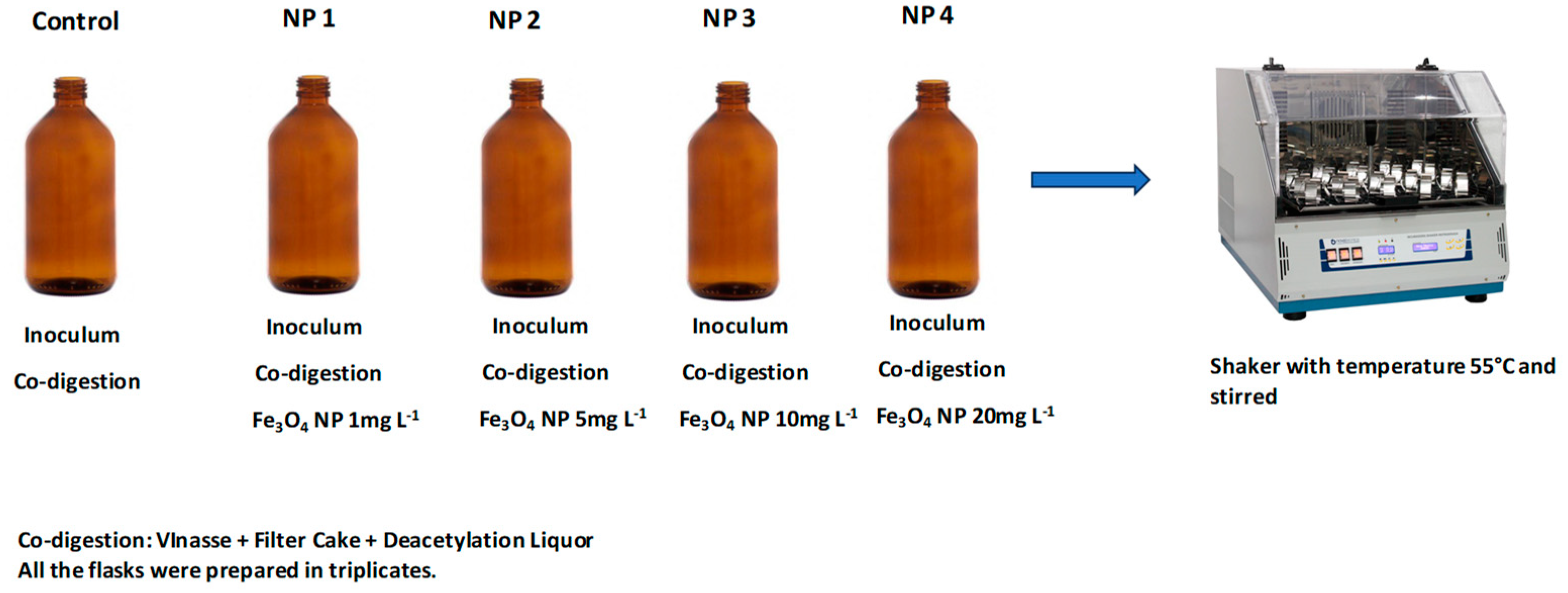
2.2. Batch Tests
2.3. Operation of Semi-Continuous Stirred Tank Reactor (s-CSTR)
s-CSTR Monitoring Analyses
2.4. Molecular Analysis in Biological Studies
2.5. NP Preparations and Characterization
3. Results
4. Discussion
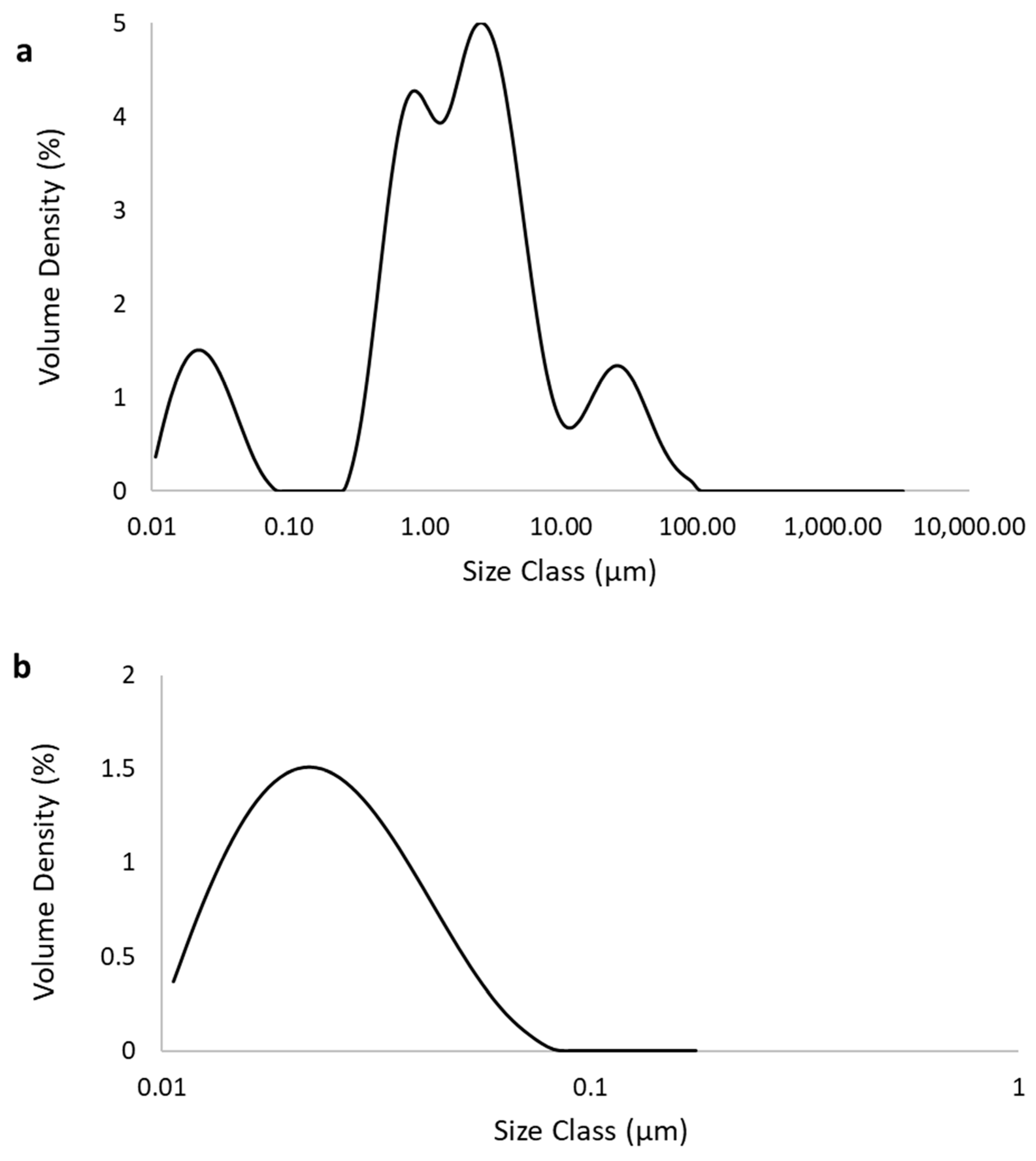
4.1. Characterization of Fe3O4 NP
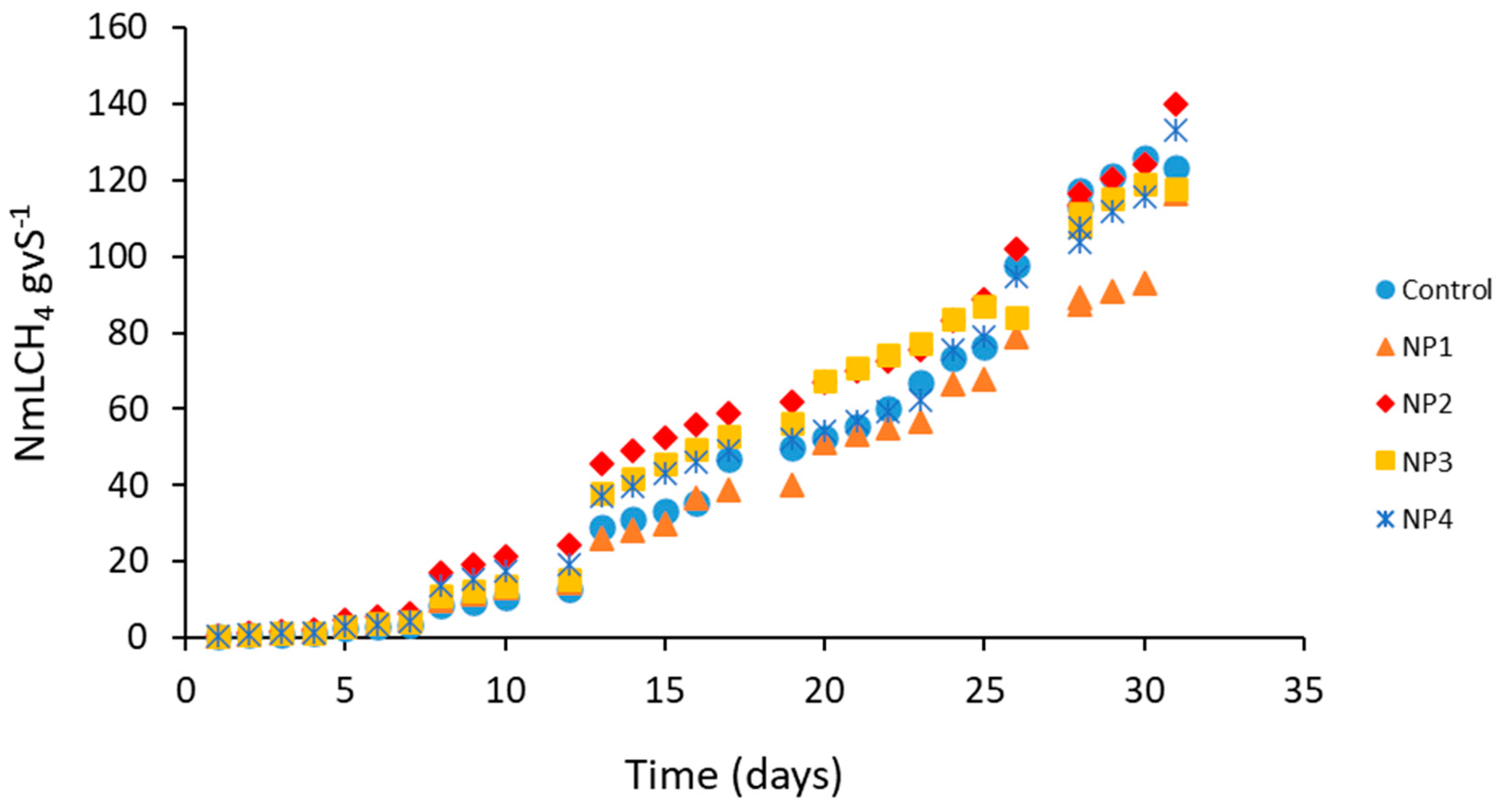
4.2. Batch Preliminary Assays
4.3. Analysis of s-CSTR Operational Efficiency
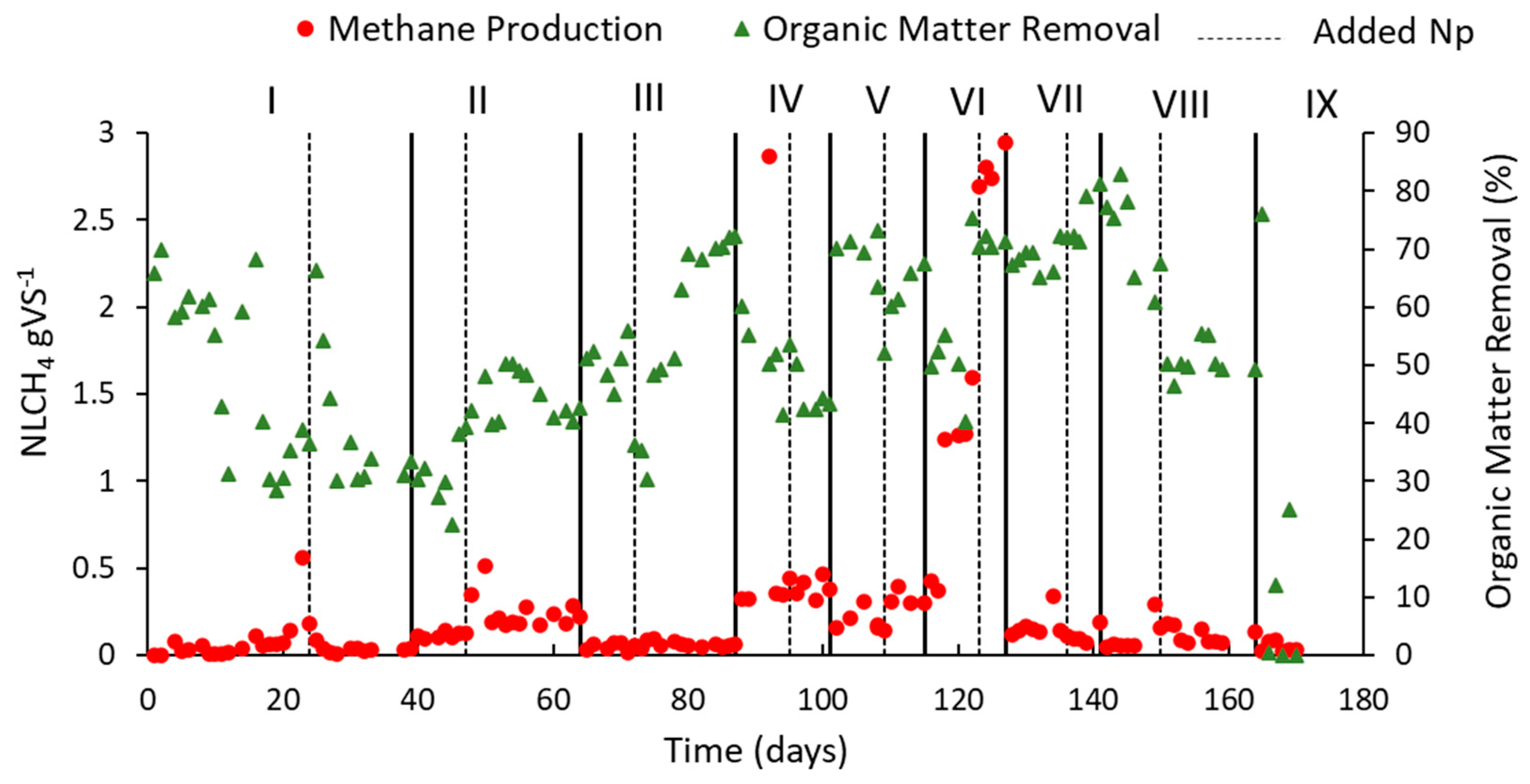
4.3.1. Biogas Generations and Reactor Performance
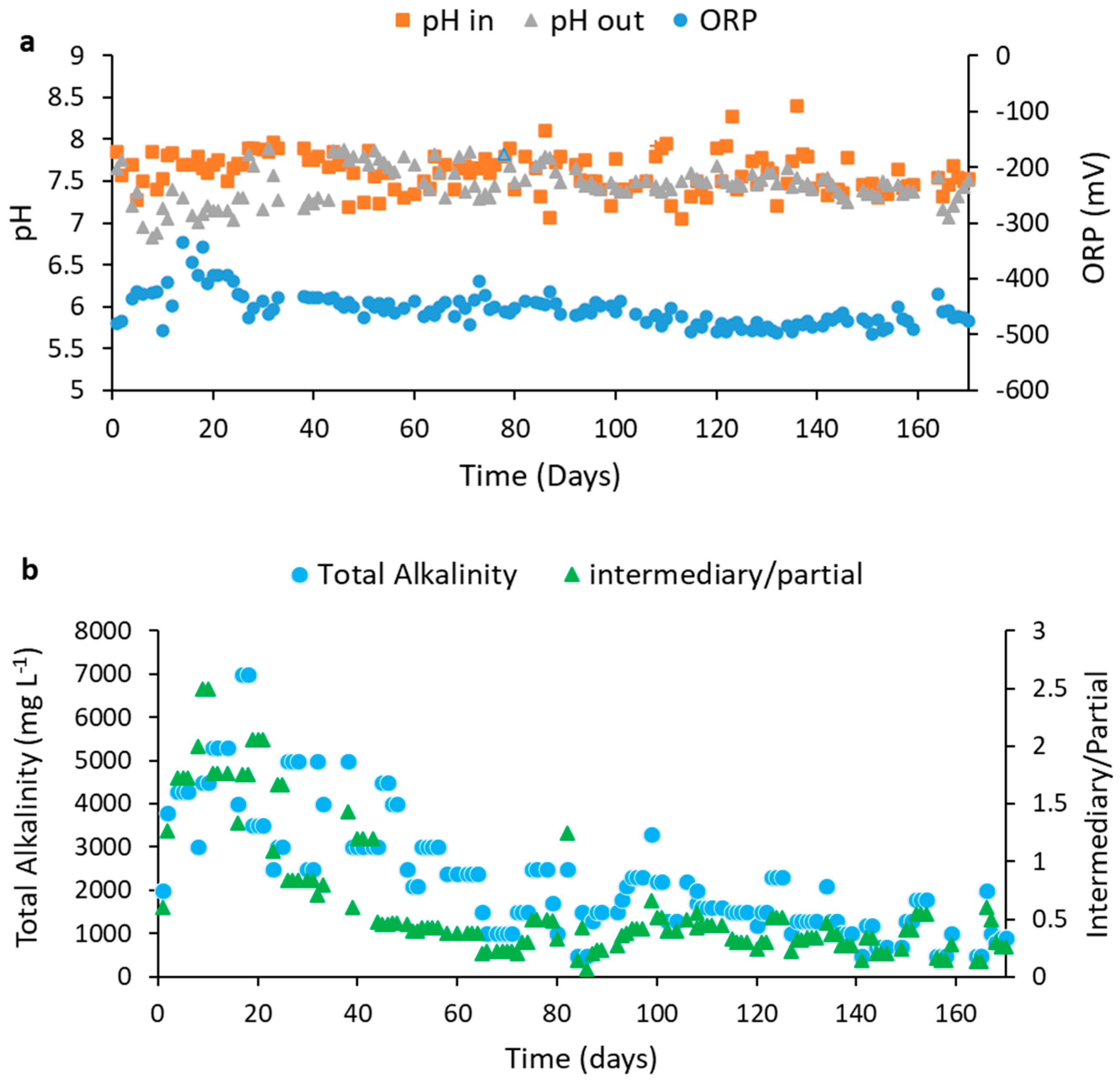
4.3.2. Evaluation of pH, ORP, and Alkalinity Readings
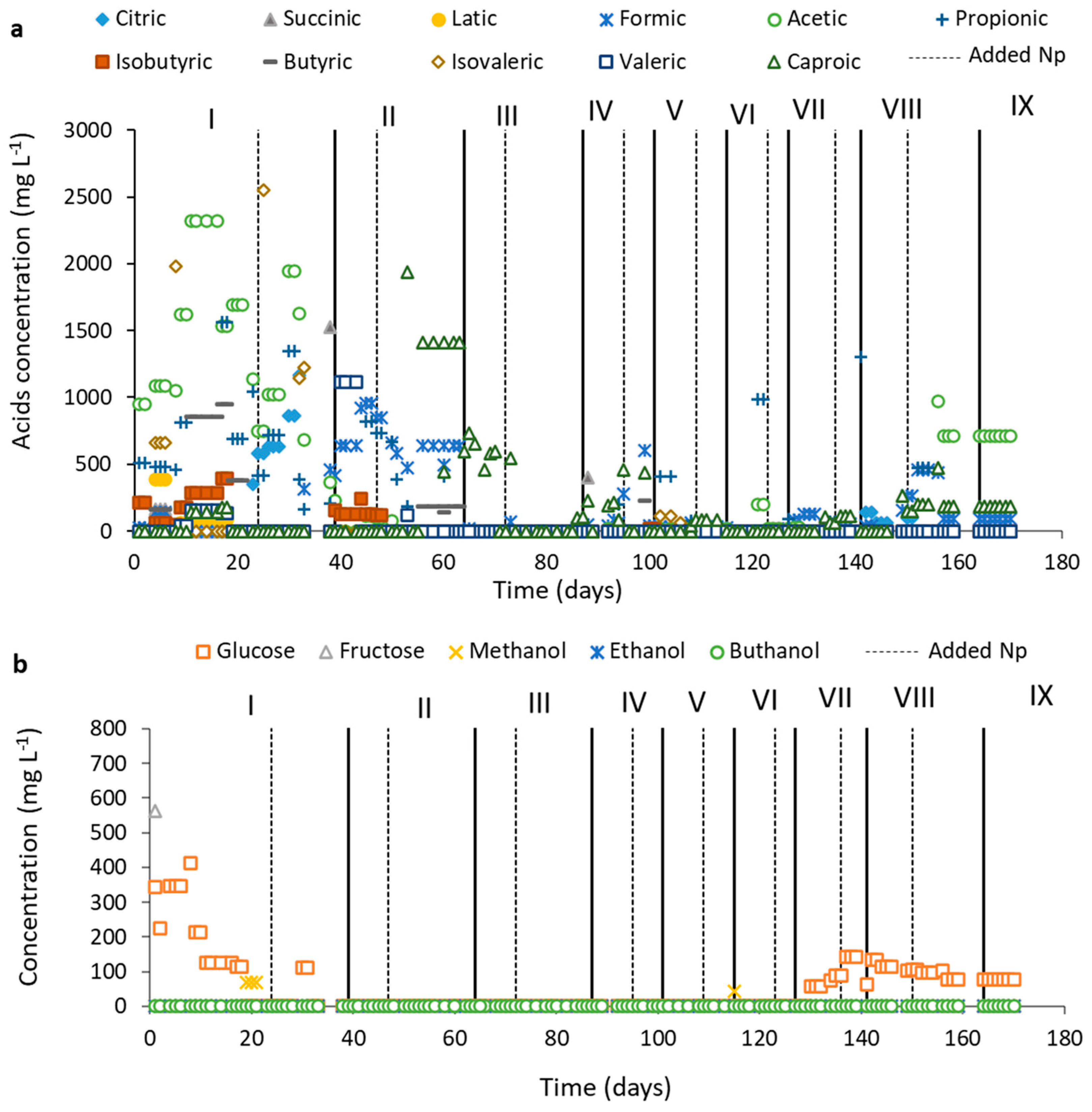
4.3.3. Assessment of Degradation Pathways: OA, Carbohydrate, and Alcohol Indications
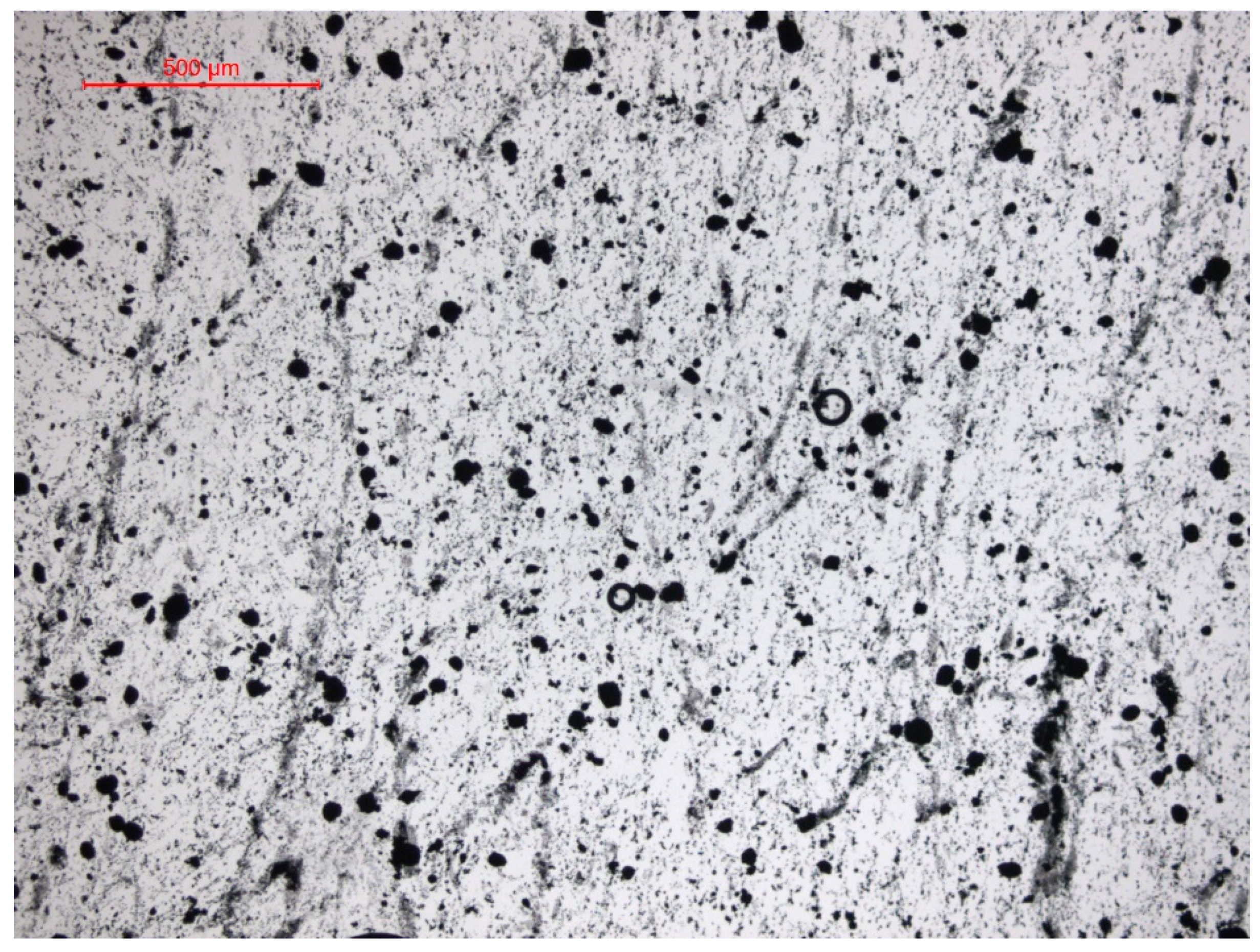
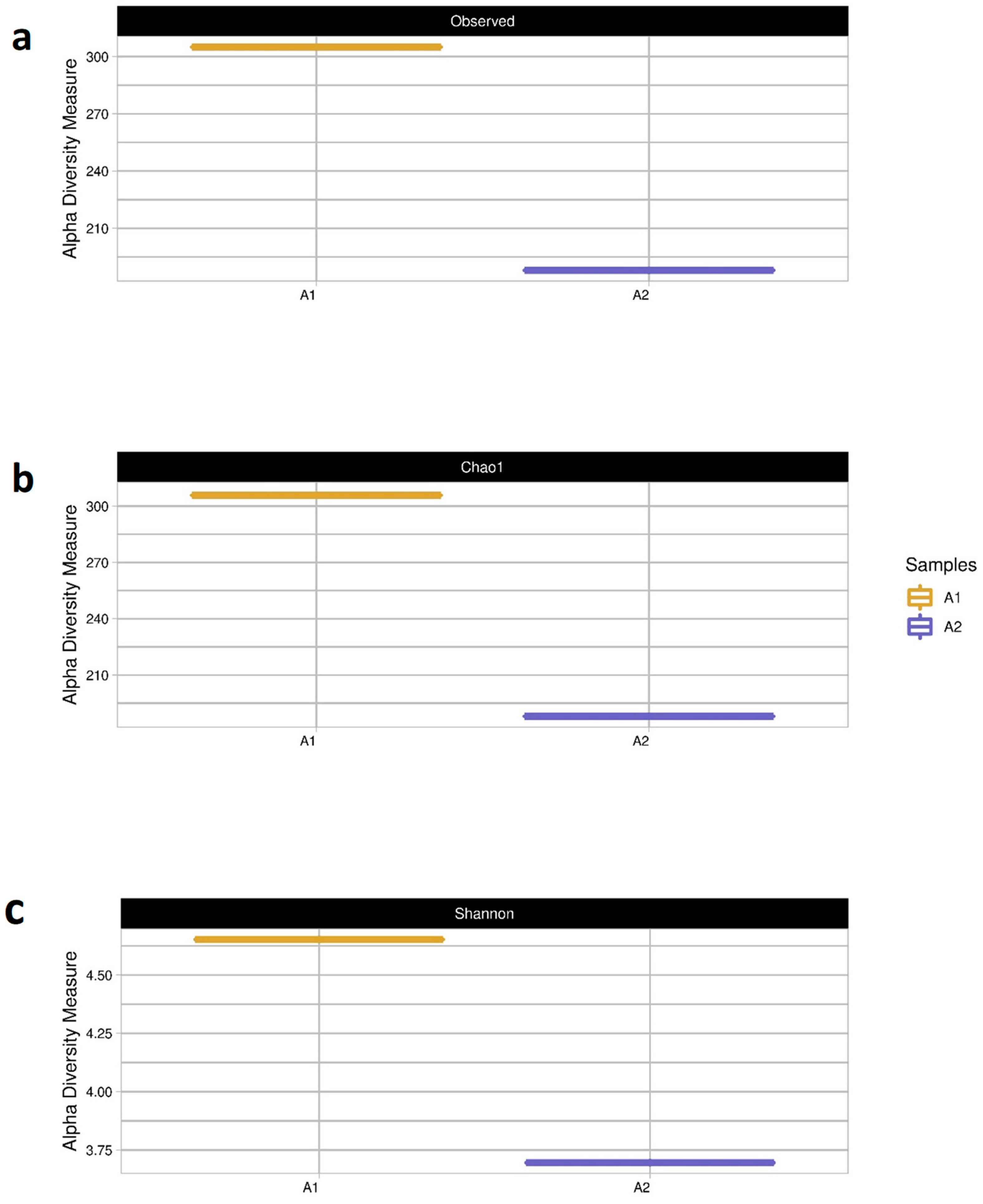
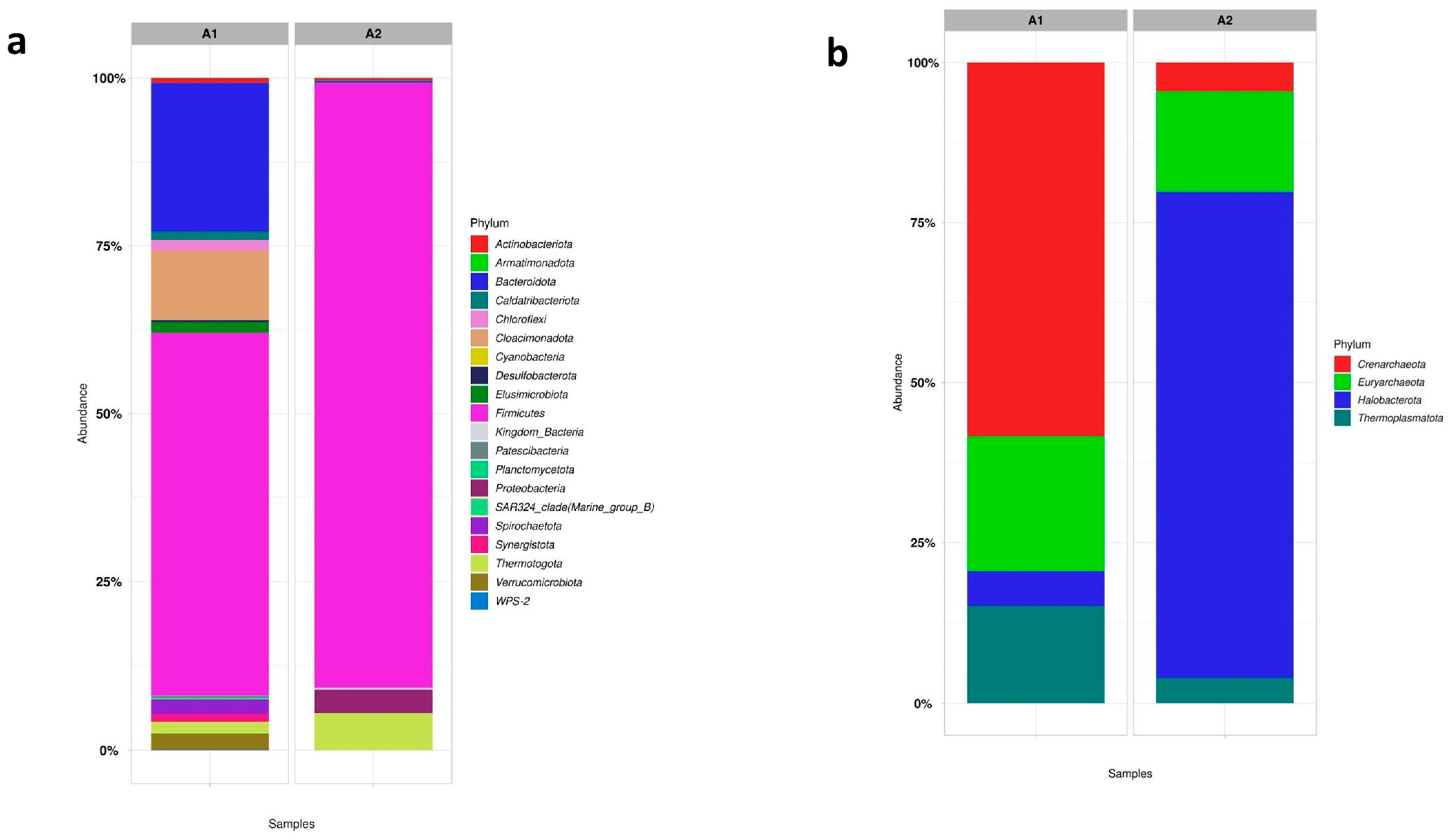
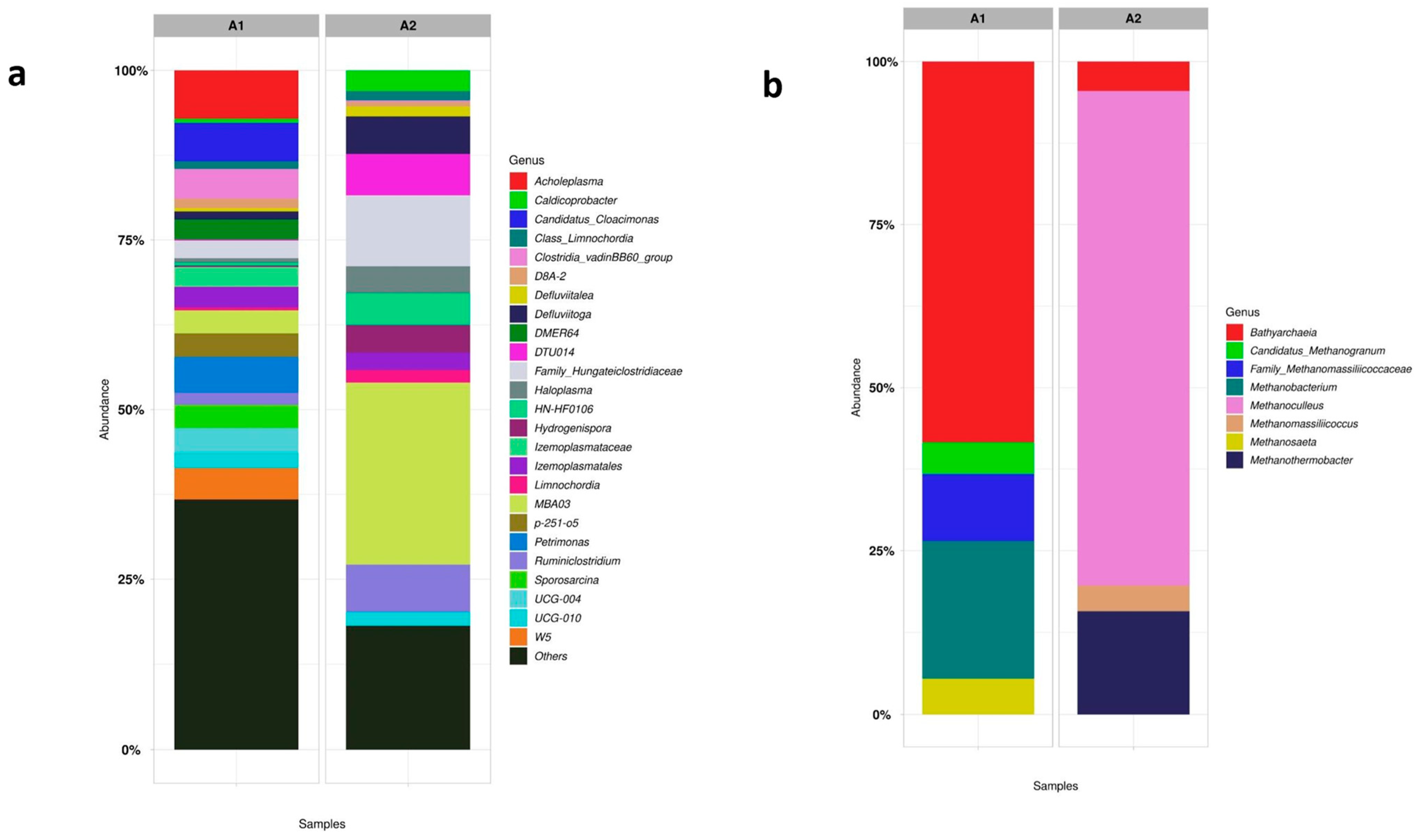
4.3.4. Assessing the Diversity of Microbial Communities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karrabi, M.; Ranjbar, F.M.; Shahnavaz, B.; Seyedi, S. A Comprehensive Review on Biogas Production from Lignocellulosic Wastes through Anaerobic Digestion: An Insight into Performance Improvement Strategies. Fuel 2023, 340, 127239. [Google Scholar] [CrossRef]
- Nunes Ferraz, D.A., Jr.; Koyama, M.H.; de Araújo, M.M., Jr.; Zaiat, M. Thermophilic Anaerobic Digestion of Raw Sugarcane Vinasse. Renew. Energy 2016, 89, 245–252. [Google Scholar] [CrossRef]
- Fuess, L.T.; Kiyuna, L.S.M.; Ferraz, A.D.N.; Persinoti, G.F.; Squina, F.M.; Garcia, M.L.; Zaiat, M. Thermophilic Two-Phase Anaerobic Digestion Using an Innovative Fixed-Bed Reactor for Enhanced Organic Matter Removal and Bioenergy Recovery from Sugarcane Vinasse. Appl. Energy 2017, 189, 480–491. [Google Scholar] [CrossRef]
- Moraes, B.S.; Zaiat, M.; Bonomi, A. Anaerobic Digestion of Vinasse from Sugarcane Ethanol Production in Brazil: Challenges and Perspectives. Renew. Sustain. Energy Rev. 2015, 44, 888–903. [Google Scholar] [CrossRef]
- Almomani, F.; Bhosale, R.R. Enhancing the Production of Biogas through Anaerobic Co-Digestion of Agricultural Waste and Chemical Pre-Treatments. Chemosphere 2020, 255, 126805. [Google Scholar] [CrossRef]
- Abbas, Y.; Yun, S.; Mehmood, A.; Shah, F.A.; Wang, K.; Eldin, E.T.; Al-Qahtani, W.H.; Ali, S.; Bocchetta, P. Co-Digestion of Cow Manure and Food Waste for Biogas Enhancement and Nutrients Revival in Bio-Circular Economy. Chemosphere 2023, 311, 137018. [Google Scholar] [CrossRef]
- Volpi, M.P.C.; Brenelli, L.B.; Mockaitis, G.; Rabelo, S.C.; Franco, T.T.; Moraes, B.S. Use of Lignocellulosic Residue from Second-Generation Ethanol Production to Enhance Methane Production through Co-Digestion. Bioenergy Res. 2021, 15, 602–616. [Google Scholar] [CrossRef]
- Volpi, M.P.C.; Junior, A.D.N.F.; Franco, T.T.; Moraes, B.S. Operational and Biochemical Aspects of Co-Digestion (Co-AD) from Sugarcane Vinasse, Filter Cake, and Deacetylation Liquor. Appl. Microbiol. Biotechnol. 2021, 105, 8969–8987. [Google Scholar] [CrossRef]
- Longati, A.A.; Lino, A.R.A.; Giordano, R.C.; Furlan, F.F.; Cruz, A.J.G. Biogas Production from Anaerobic Digestion of Vinasse in Sugarcane Biorefinery: A Techno-Economic and Environmental Analysis. Waste Biomass Valori. 2020, 11, 4573–4591. [Google Scholar] [CrossRef]
- Brenelli, L.B.; Figueiredo, F.L.; Damasio, A.; Franco, T.T.; Rabelo, S.C. An Integrated Approach to Obtain Xylo-Oligosaccharides from Sugarcane Straw: From Lab to Pilot Scale. Bioresour. Technol. 2020, 313, 123637. [Google Scholar] [CrossRef]
- Juntupally, S.; Begum, S.; Anupoju, G.R. Impact of Additives (Macronutrient and Nanoparticles of Micronutrients) on the Anaerobic Digestion of Food Waste: Focus on Feeding Strategy for Improved Performance. Biomass Bioenergy 2023, 172, 106751. [Google Scholar] [CrossRef]
- Scherer, P.; Lippert, H.; Wolff, G. Composition of the Major Elements and Trace Elements of 10 Methanogenic Bacteria Determined by Inductively Coupled Plasma Emission Spectrometry. Biol. Trace Elem. Res. 1983, 5, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Z.; Suzuki, K.; Maekawa, T. Uptake and Mass Balance of Trace Metals for Methane Producing Bacteria. Biomass Bioenergy 2003, 25, 427–433. [Google Scholar] [CrossRef]
- Cui, Z.; Liu, Z.; Fan, Y.; He, Z.W.; Liu, W.; Yue, X.; Zhou, A. Improving Methane Production from Waste Activated Sludge Assisted by Fe(II)-Activated Peroxydisulfate Pretreatment via Anaerobic Digestion: Role of Interspecific Syntrophism Mediated by Sulfate-Reducing Bacteria. ACS Sustain. Chem. Eng. 2023, 11, 3012–3022. [Google Scholar] [CrossRef]
- Yu, B.; Lou, Z.; Zhang, D.; Shan, A.; Yuan, H.; Zhu, N.; Zhang, K. Variations of Organic Matters and Microbial Community in Thermophilic Anaerobic Digestion of Waste Activated Sludge with the Addition of Ferric Salts. Bioresour. Technol. 2015, 179, 291–298. [Google Scholar] [CrossRef]
- Demirel, B.; Scherer, P. Trace Element Requirements of Agricultural Biogas Digesters during Biological Conversion of Renewable Biomass to Methane. Biomass Bioenergy 2011, 35, 992–998. [Google Scholar] [CrossRef]
- Zhang, Y.; Jing, Y.; Zhang, J.; Sun, L.; Quan, X. Performance of a ZVI-UASB Reactor for Azo Dye Wastewater Treatment. J. Chem. Technol. Biotechnol. 2011, 86, 199–204. [Google Scholar] [CrossRef]
- Abdelsalam, E.; Samer, M.; Attia, Y.A.; Abdel-Hadi, M.A.; Hassan, H.E.; Badr, Y. Comparison of Nanoparticles Effects on Biogas and Methane Production from Anaerobic Digestion of Cattle Dung Slurry. Renew. Energy 2016, 87, 592–598. [Google Scholar] [CrossRef]
- Abdelsalam, E.; Samer, M.; Attia, Y.A.; Abdel-Hadi, M.A.; Hassan, H.E.; Badr, Y. Effects of Co and Ni Nanoparticles on Biogas and Methane Production from Anaerobic Digestion of Slurry. Energy Convers. Manag. 2017, 141, 108–119. [Google Scholar] [CrossRef]
- Abdelsalam, E.; Samer, M.; Attia, Y.A.; Abdel-Hadi, M.A.; Hassan, H.E.; Badr, Y. Influence of Zero Valent Iron Nanoparticles and Magnetic Iron Oxide Nanoparticles on Biogas and Methane Production from Anaerobic Digestion of Manure. Energy 2017, 120, 842–853. [Google Scholar] [CrossRef]
- Hassanein, A.; Lansing, S.; Tikekar, R. Bioresource Technology Impact of Metal Nanoparticles on Biogas Production from Poultry Litter. Bioresour. Technol. 2019, 275, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Amo-Duodu, G.; Rathilal, S.; Chollom, M.N.; Tetteh, E.K. Effects of Synthesized AlFe2O4 and MgFe2O4 Nanoparticles on Biogas Production from Anaerobically Digested Sugar Refinery Wastewater. Environ. Sci. Pollut. Res. 2023, 30, 25613–25619. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.W.; Baird, R.B.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater; American Public Health Association, American Water Works Association, Water Environment Federation: Washington, DC, USA, 2012. [Google Scholar]
- Triolo, J.M.; Pedersen, L.; Qu, H.; Sommer, S.G. Biochemical Methane Potential and Anaerobic Biodegradability of Non-Herbaceous and Herbaceous Phytomass in Biogas Production. Bioresour. Technol. 2012, 125, 226–232. [Google Scholar] [CrossRef] [PubMed]
- VDI 4630; Fermentation of Organic Materials. Characterization of the Substrate, Sampling, Collection of Material Data, Fermentation Tests. Verein Deutscher Ingenieure: Düsseldorf, Germany, 2006.
- Wang, T.; Zhang, D.; Dai, L.; Chen, Y.; Dai, X. Effects of Metal Nanoparticles on Methane Production from Waste-Activated Sludge and Microorganism Community Shift in Anaerobic Granular Sludge. Sci. Rep. 2016, 6, 25857. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Mahar, R.B.; Soomro, R.A.; Sherazi, S.T.H. Fe3O4 Nanoparticles Facilitated Anaerobic Digestion of Organic Fraction of Municipal Solid Waste for Enhancement of Methane Production. Energy Sources Part A Recovery Util. Environ. Eff. 2017, 39, 1815–1822. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, L.; Wang, Y.; Zhao, Y.; She, Z.; Gao, M.; Guo, Y. Application of Iron Oxide (Fe3O4) Nanoparticles during the Two-Stage Anaerobic Digestion with Waste Sludge: Impact on the Biogas Production and the Substrate Metabolism. Renew. Energy 2020, 146, 2724–2735. [Google Scholar] [CrossRef]
- Vongvichiankul, C.; Deebao, J.; Khongnakorn, W. Relationship between PH, Oxidation Reduction Potential (ORP) and Biogas Production in Mesophilic Screw Anaerobic Digester. Energy Procedia 2017, 138, 877–882. [Google Scholar] [CrossRef]
- Xie, S.; Li, X.; Wang, C.; Kulandaivelu, J.; Jiang, G. Enhanced Anaerobic Digestion of Primary Sludge with Additives: Performance and Mechanisms. Bioresour. Technol. 2020, 316, 123a970. [Google Scholar] [CrossRef]
- Zhang, L.; Gong, X.; Wang, L.; Guo, K.; Cao, S.; Zhou, Y. Metagenomic Insights into the Effect of Thermal Hydrolysis Pre-Treatment on Microbial Community of an Anaerobic Digestion System. Sci. Total Environ. 2021, 791, 148096. [Google Scholar] [CrossRef]
- Jiang, W.E.N.; Kim, B.Y.S.; Rutka, J.T.; Chan, W.C.W. Nanoparticle-Mediated Cellular Response Is Size-Dependent. Nat. Nanotechnol. 2008, 3, 145–150. [Google Scholar] [CrossRef]
- Dompara, I.; Maragkaki, A.; Papastefanakis, N.; Floraki, C.; Vernardou, D.; Manios, T. Effects of Different Materials on Biogas Production during Anaerobic Digestion of Food Waste. Sustainability 2023, 15, 5698. [Google Scholar] [CrossRef]
- Gonzalez-estrella, J.; Sierra-alvarez, R.; Field, J.A. Toxicity Assessment of Inorganic Nanoparticles to Acetoclastic and Hydrogenotrophic Methanogenic Activity in Anaerobic Granular Sludge. J. Hazard. Mater. 2013, 260, 278–285. [Google Scholar] [CrossRef]
- Krongthamchat, K.; Riffat, R.; Dararat, S. Effect of Trace Metals on Halophilic and Mixed Cultures in Anaerobic Treatment. Int. J. Environ. Sci. Technol. 2006, 3, 103–112. [Google Scholar] [CrossRef][Green Version]
- Zhu, X.; Blanco, E.; Bhatti, M.; Borrion, A. Impact of Metallic Nanoparticles on Anaerobic Digestion: A Systematic Review. Sci. Total Environ. 2021, 757, 143747. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.Q.; Ni, J.; Yang, N.; Wang, J. Effect of Magnetic Nanoparticles on the Performance of Activated Sludge Treatment System. Bioresour. Technol. 2013, 143, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Ameen, F.; Ranjitha, J.; Ahsan, N.; Shankar, V. Co-Digestion of Microbial Biomass with Animal Manure in Three-Stage Anaerobic Digestion. Fuel 2021, 306, 121746. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Y.; Chen, M.; Xie, X.; Li, C.; Wang, J.; Yuan, L. Dry Reforming of Methane for Syngas Production over Attapulgite-Derived MFI Zeolite Encapsulated Bimetallic Ni-Co Catalysts. Appl. Catal. B 2023, 322, 122088. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, Y.; Quan, X.; Chen, S. Enhanced Anaerobic Digestion of Waste Activated Sludge Digestion by the Addition of Zero Valent Iron. Water Res. 2014, 52, 242–250. [Google Scholar] [CrossRef]
- Ajayi-Banji, A.; Rahman, S. A Review of Process Parameters Influence in Solid-State Anaerobic Digestion: Focus on Performance Stability Thresholds. Renew. Sustain. Energy Rev. 2022, 167, 112756. [Google Scholar] [CrossRef]
- Lee, Y.J.; Lee, D.J. Impact of Adding Metal Nanoparticles on Anaerobic Digestion Performance—A Review. Bioresour. Technol. 2019, 292, 121926. [Google Scholar] [CrossRef]
- Ripley, L.E.; Boyle, W.C.; Converse, J.C. Improved Alkalimetric Monitoring for Anaerobic Digestion of High-Strength Waste. J. Water Pollut. Control. Fed. 1986, 58, 406–411. [Google Scholar]
- Wiegant, W.M.; Hennink, M.; Lettinga, G. Separation of the Propionate Degradation to Improve the Efficiency of Thermophilic Anaerobic Treatment of Acidified Wastewaters. Water Res. 1986, 20, 517–524. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Wang, J.; Meng, L. Effects of Volatile Fatty Acid Concentrations on Methane Yield and Methanogenic Bacteria. Biomass Bioenergy 2009, 33, 848–853. [Google Scholar] [CrossRef]
- Romero-Güiza, M.S.; Vila, J.; Mata-Alvarez, J.; Chimenos, J.M.; Astals, S. The Role of Additives on Anaerobic Digestion: A Review. Renew. Sustain. Energy Rev. 2016, 58, 1486–1499. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, S.; Guo, J.; Zhou, J.; Dong, R. Performance and Kinetic Evaluation of Semi-Continuously Fed Anaerobic Digesters Treating Food Waste: Role of Trace Elements. Bioresour. Technol. 2015, 178, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Agyeman, I.; Plaza, E.; Cetecioglu, Z. Production of Volatile Fatty Acids through Co-Digestion of Sewage Sludge and External Organic Waste: Effect of Substrate Proportions and Long-Term Operation. Waste Manag. 2020, 112, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Angenent, L.T.; Richter, H.; Buckel, W.; Spirito, C.M.; Steinbusch, K.J.J.; Plugge, C.M.; Strik, D.P.B.T.B.; Grootscholten, T.I.M.; Buisman, C.J.N.; Hamelers, H.V.M. Chain Elongation with Reactor Microbiomes: Open-Culture Biotechnology to Produce Biochemicals. Env. Sci. Technol. 2016, 50, 2796–2810. [Google Scholar] [CrossRef] [PubMed]
- de Souza Moraes, B.; Mary dos Santos, G.; Palladino Delforno, T.; Tadeu Fuess, L.; José da Silva, A. Enriched Microbial Consortia for Dark Fermentation of Sugarcane Vinasse towards Value-Added Short-Chain Organic Acids and Alcohol Production. J. Biosci. Bioeng. 2019, 127, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Tian, Z.; Lv, Z.; Chen, Z.; Liu, Y.; Yong, X.; Zhou, J.; Xie, X.; Jia, H.; Wei, P. Effects of Copper Salts on Performance, Antibiotic Resistance Genes, and Microbial Community during Thermophilic Anaerobic Digestion of Swine Manure. Bioresour. Technol. 2020, 300, 122728. [Google Scholar] [CrossRef]
- Wang, P.; Wang, H.; Qiu, Y.; Ren, L.; Jiang, B. Microbial Characteristics in Anaerobic Digestion Process of Food Waste for Methane Production–A Review. Bioresour. Technol. 2018, 248, 29–36. [Google Scholar] [CrossRef]
- Guo, X.; Wang, C.; Sun, F.; Zhu, W.; Wu, W. A Comparison of Microbial Characteristics between the Thermophilic and Mesophilic Anaerobic Digesters Exposed to Elevated Food Waste Loadings. Bioresour. Technol. 2014, 152, 420–428. [Google Scholar] [CrossRef]
- Peng, X.; Börner, R.A.; Nges, I.A.; Liu, J. Impact of Bioaugmentation on Biochemical Methane Potential for Wheat Straw with Addition of Clostridium Cellulolyticum. Bioresour. Technol. 2014, 152, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.R.; Su, Y.; Wang, J.; Chu, Y.X.; Tian, G.; He, R. Effects of Different Pretreatment Methods on Biogas Production and Microbial Community in Anaerobic Digestion of Wheat Straw. Environ. Sci. Pollut. Res. 2021, 28, 51772–51785. [Google Scholar] [CrossRef]
- Schnürer, A.; Zellner, G.; Svensson, B.H. Mesophilic Syntrophic Acetate Oxidation during Methane Formation in Biogas Reactors. FEMS Microbiol. Ecol. 1999, 29, 249–261. [Google Scholar] [CrossRef]
- Li, Z.; Wachemo, A.C.; Yuan, H.; Korai, R.M.; Li, X. Improving Methane Content and Yield from Rice Straw by Adding Extra Hydrogen into a Two-Stage Anaerobic Digestion System. Int. J. Hydrogen Energy 2020, 45, 3739–3749. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Vinasse (mg L−1) | Deacetylation Liquor (mg L−1) |
|---|---|---|
| Acetate | 1268.41 | 3250.00 |
| Formate | - | 650.00 |
| Lactate | 3706.94 | 423.18 |
| Propionate | 634.85 | 368.29 |
| Butyrate | - | 250.02 |
| Isovalerate | 931.63 | 269.03 |
| Glucose | 809.05 | 546.23 |
| Methanol | 8674.83 | - |
| Flasks | Name in Graph | Fe3O4 NP Concentration (mg L−1) |
|---|---|---|
| Control | 0 |
| NP 1 | 1 |
| NP 2 | 5 |
| NP 3 | 10 |
| NP 4 | 20 |
| Phase in Graph | OLR (gVS L−1 Day−1) | Feeding Rate (L Day−1) | HRT (Days) | NP Addition Day |
|---|---|---|---|---|
| I | 2 | 0.250 | 16 | 24 |
| II | 2.35 | 0.285 | 14 | 47 |
| III | 3 | 0.363 | 11 | 72 |
| IV | 4 | 0.500 | 8 | 95 |
| V | 4.70 | 0.571 | 7 | 109 |
| VI | 5.5 | 0.666 | 6 | 123 |
| VII | 6.6 | 0.800 | 5 | 136 |
| VIII | 8 | 1.000 | 4 | 150 |
| IX | 9 | 1.140 | 3.5 | - |
| Assay | Cumulative CH4 (NmLVS−1) a |
|---|---|
| Control | 123.24 ± 9.60 |
| NP 1 | 116.49 ± 17.45 |
| NP 2 | 140.13 ± 95.60 |
| NP 3 | 117.90 ± 10.68 |
| NP 4 | 133.02 ± 106.29 |
| A1 | A2 | |
|---|---|---|
| Species Abundance | 308 | 189 |
| Estimated species richness | 308 | 189 |
| Computed diversity values | 4.60 | 3.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volpi, M.P.C.; Mockaitis, G.; Moraes, B.d.S. Fe3O4 Nanoparticles to Optimize the Co-Digestion of Vinasse, Filter Cake, and Deacetylation Liquor: Operational Aspects and Microbiological Routes. Appl. Nano 2023, 4, 240-259. https://doi.org/10.3390/applnano4030014
Volpi MPC, Mockaitis G, Moraes BdS. Fe3O4 Nanoparticles to Optimize the Co-Digestion of Vinasse, Filter Cake, and Deacetylation Liquor: Operational Aspects and Microbiological Routes. Applied Nano. 2023; 4(3):240-259. https://doi.org/10.3390/applnano4030014
Chicago/Turabian StyleVolpi, Maria Paula Cardeal, Gustavo Mockaitis, and Bruna de Souza Moraes. 2023. "Fe3O4 Nanoparticles to Optimize the Co-Digestion of Vinasse, Filter Cake, and Deacetylation Liquor: Operational Aspects and Microbiological Routes" Applied Nano 4, no. 3: 240-259. https://doi.org/10.3390/applnano4030014
APA StyleVolpi, M. P. C., Mockaitis, G., & Moraes, B. d. S. (2023). Fe3O4 Nanoparticles to Optimize the Co-Digestion of Vinasse, Filter Cake, and Deacetylation Liquor: Operational Aspects and Microbiological Routes. Applied Nano, 4(3), 240-259. https://doi.org/10.3390/applnano4030014