




Using Stable Isotope Analyses to Assess the Trophic Ecology of Scleractinian Corals


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
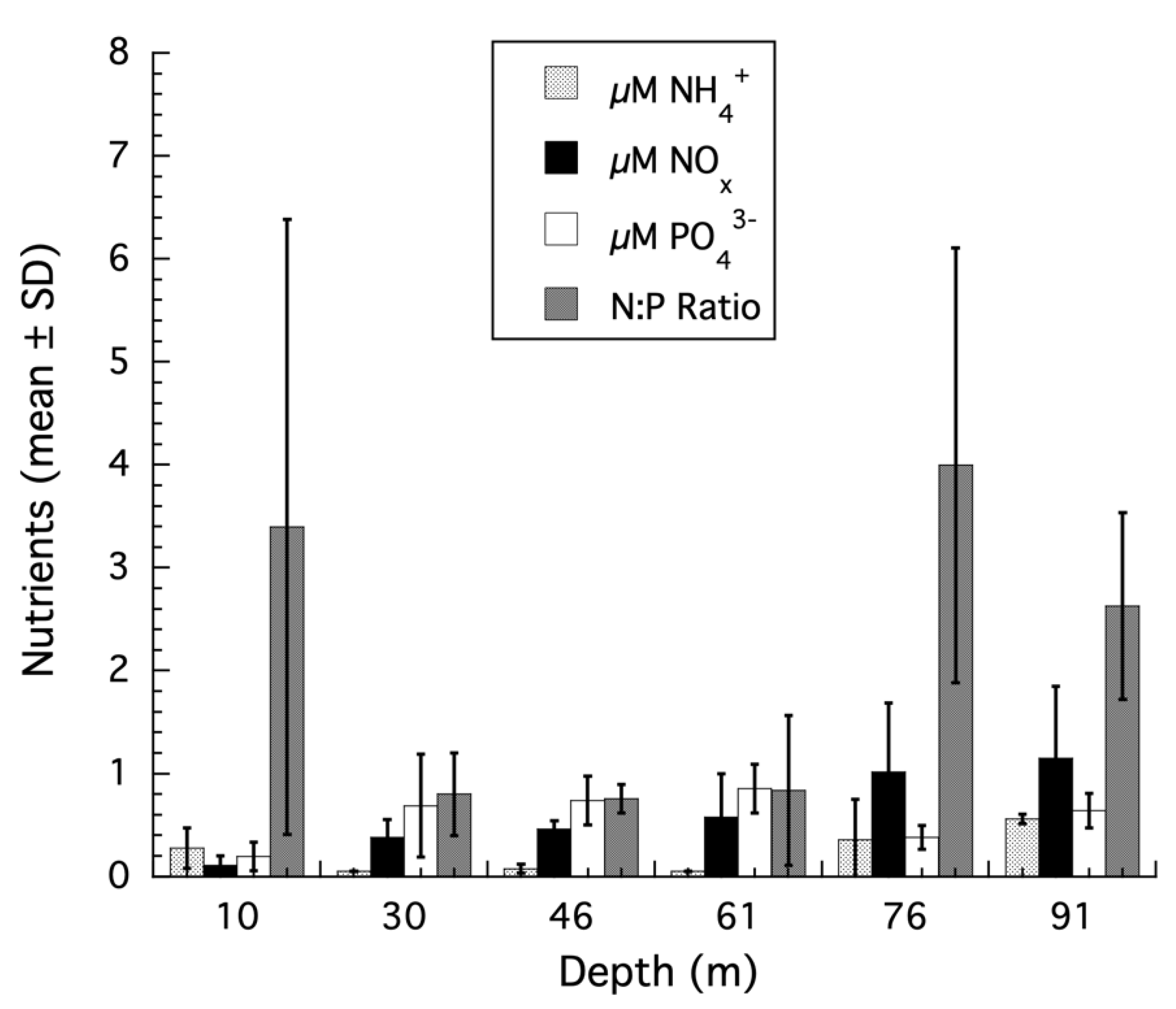
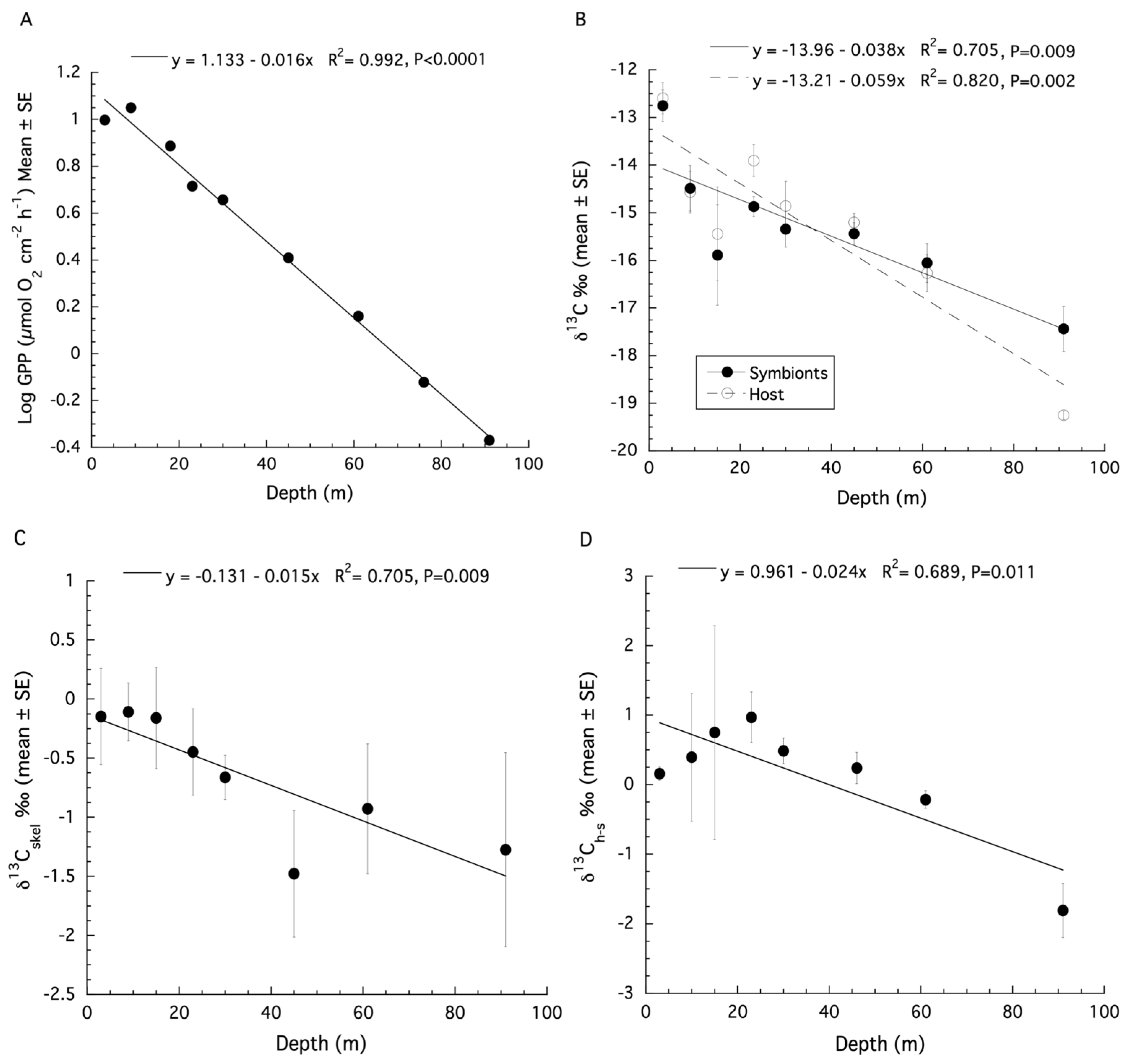
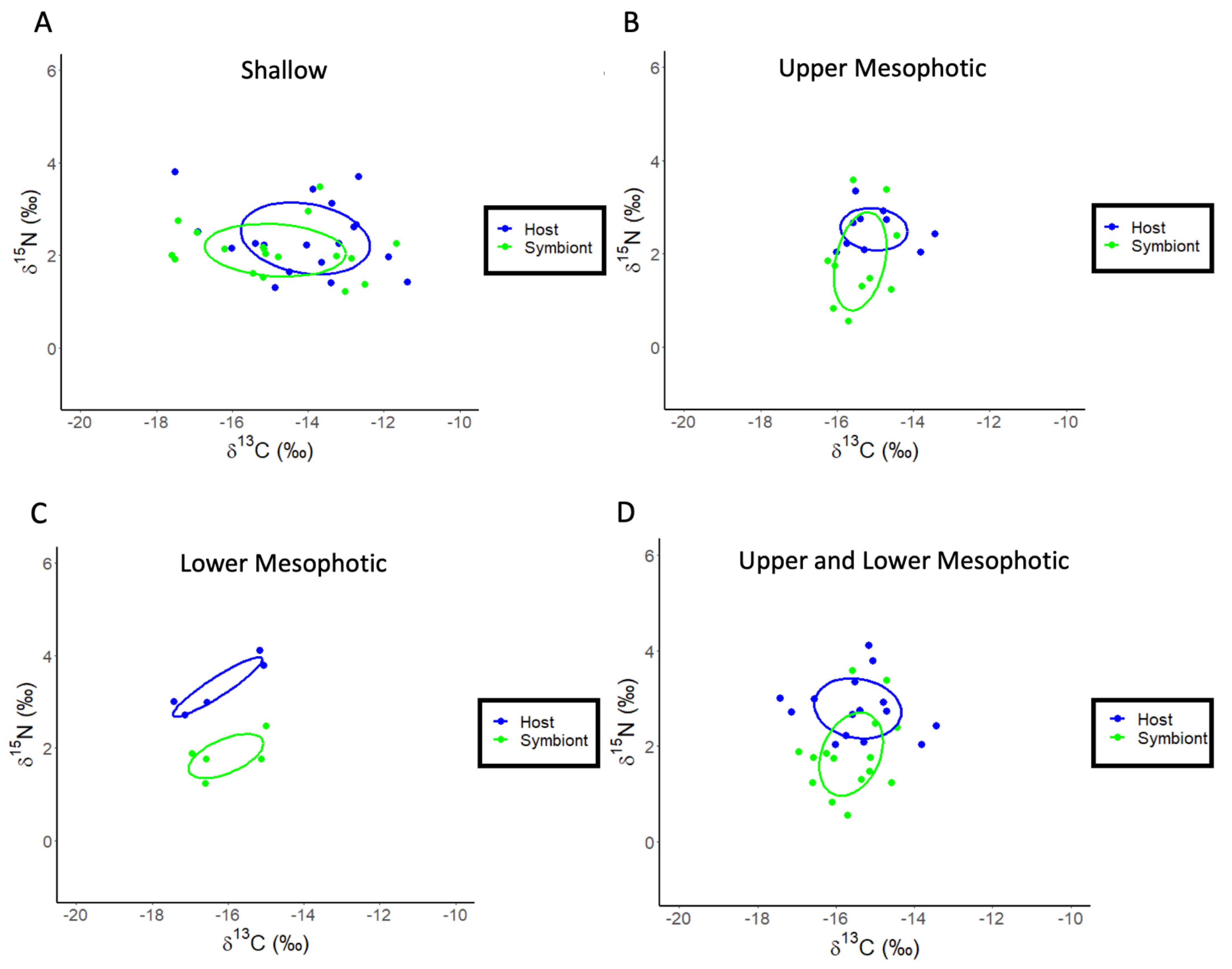
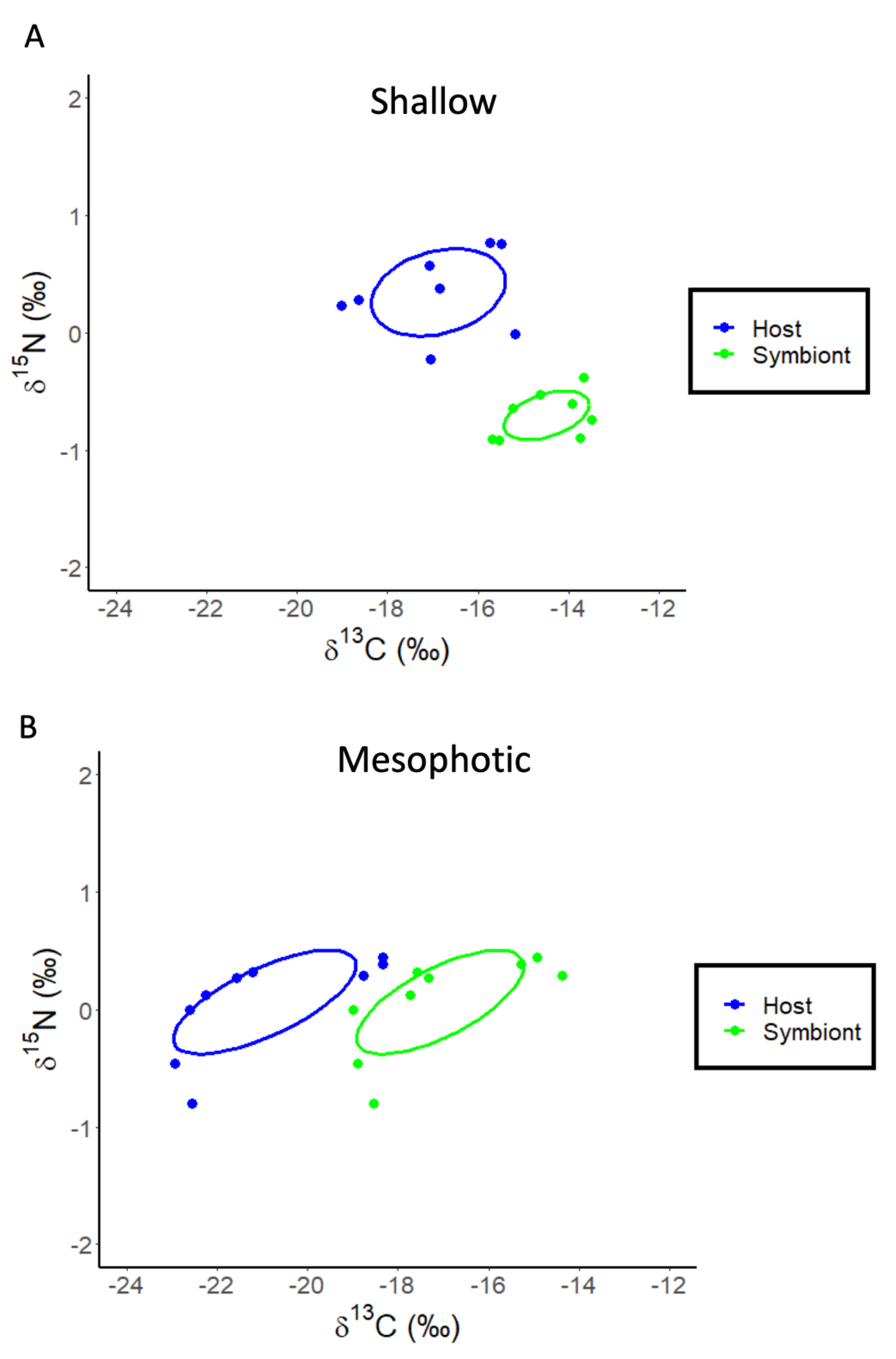
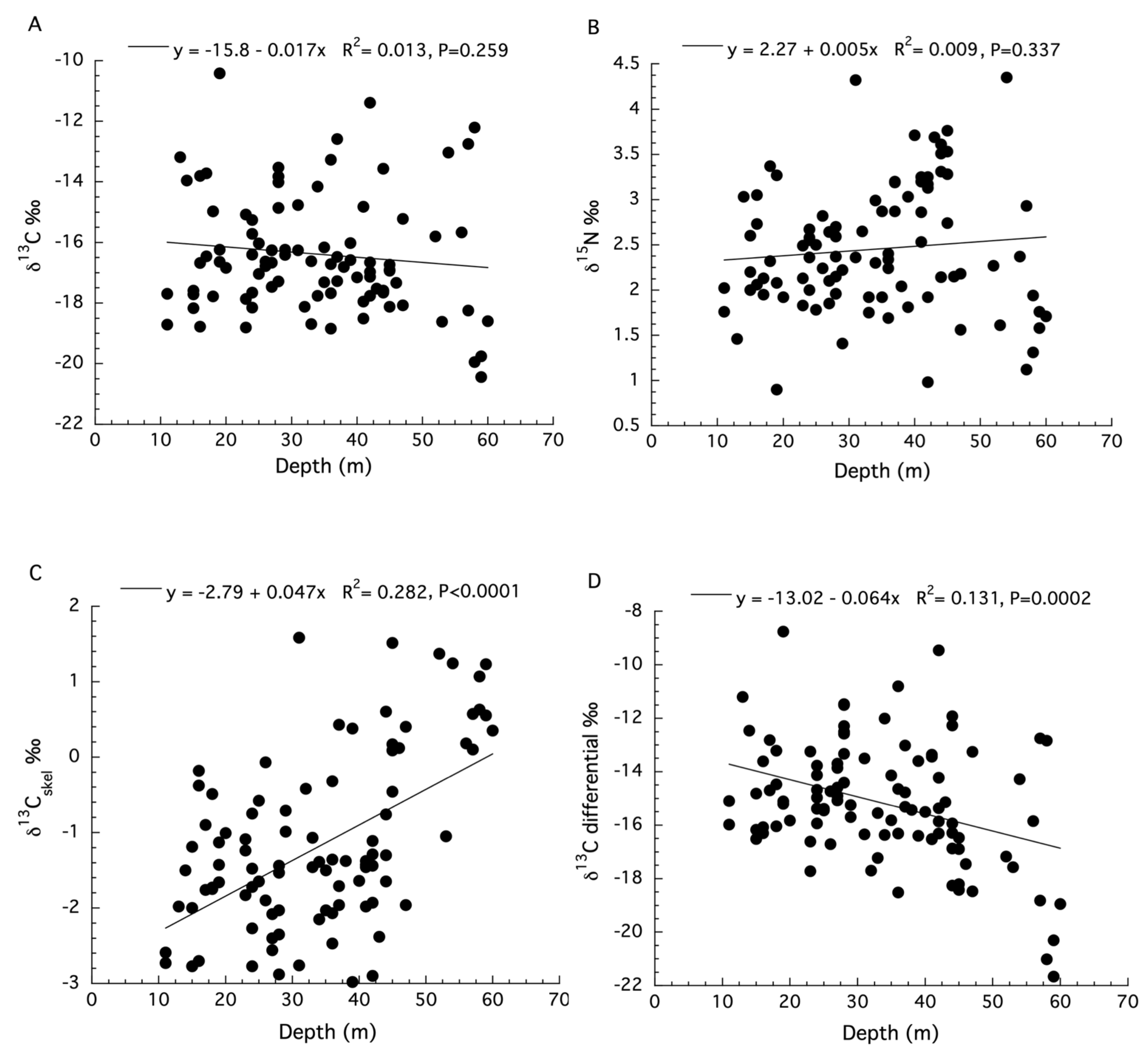
3. Results
3.1. Montastraea cavernosa
3.2. Stylophora pistillata
3.3. Agaricia lamarki
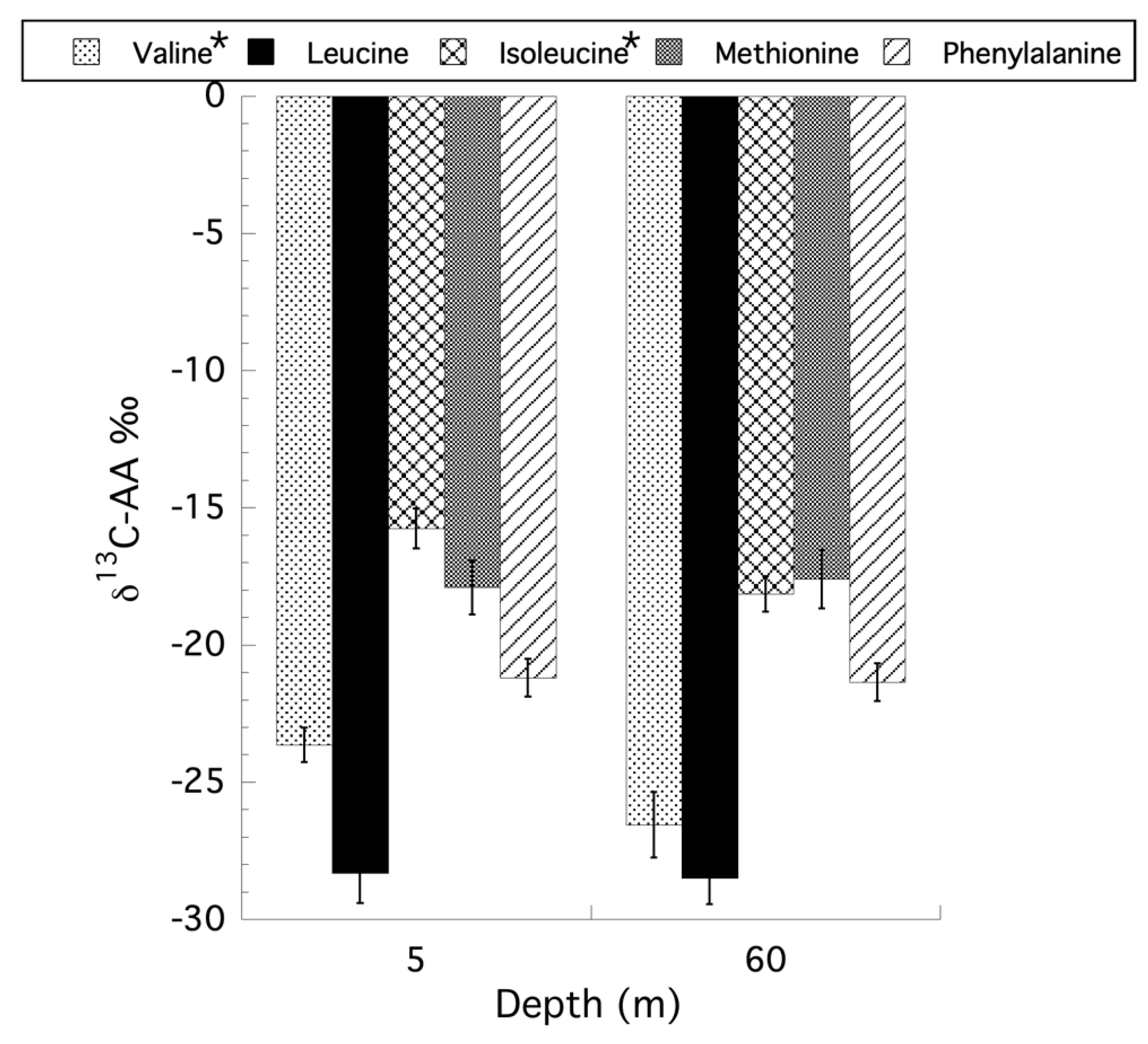
3.4. Compound-Specific Isotopic Analysis of Amino Acids (CSIA-AA) for Stylophora pistillata
4. Discussion
4.1. The Case for Heterotrophy in Mesophotic Corals
4.2. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lesser, M.P.; Slattery, M.; Leichter, J.J. Ecology of mesophotic coral reefs. J. Exp. Mar. Biol. Ecol. 2009, 375, 1–8. [Google Scholar] [CrossRef]
- Lesser, M.P.; Slattery, M.; Mobley, C.D. Biodiversity and functional ecology of mesophotic coral reefs. Ann. Rev. Ecol. Syst. 2018, 49, 49–71. [Google Scholar] [CrossRef]
- Loya, Y.; Eyal, G.; Treibitz, T.; Lesser, M.P.; Appeldoorn, R. Theme section on mesophotic coral ecosystems: Advances in knowledge and future perspectives. Coral Reefs 2016, 35, 1–9. [Google Scholar] [CrossRef]
- Laverick, J.H.; Andradi-Brown, D.A.; Rogers, A.D. Using light-dependent scleractinia to define the upper boundary of mesophotic coral ecosystems on the reefs of Utila, Honduras. PLoS ONE 2017, 12, e0183075. [Google Scholar] [CrossRef]
- Laverick, J.H.; Tamir, R.; Eyal, G.; Loya, Y. A generalized light-driven model of community transitions along coral reef depth gradients. Glob. Ecol. Biogeogr. 2020, 29, 1554–1564. [Google Scholar] [CrossRef]
- Lesser, M.P.; Slattery, M. Will Coral Reef Sponges Be Winners in the Anthropocene? Glob. Chang. Biol. 2020, 26, 3202–3211. [Google Scholar] [CrossRef] [PubMed]
- Tamir, R.; Eyal, G.; Kramer, N.; Laverick, J.H.; Loya, Y. Light environment drives the shallow-to-mesophotic coral community transition. Ecosphere 2019, 10, e02839. [Google Scholar] [CrossRef]
- Gattuso, J.-P.; Gentilli, B.; Duarte, C.M.; Kleypas, J.A.; Middleburg, J.J.; Antoine, D. Light availability in the coastal ocean: Impact on the distribution of benthic photosynthetic organisms and their contribution to primary production. Biogeosciences 2006, 3, 489–513. [Google Scholar] [CrossRef]
- Dustan, P. Depth-dependent photoadaptation by zooxanthellae of the reef coral Montastrea annularis. Mar. Biol. 1982, 68, 253–264. [Google Scholar] [CrossRef]
- Wyman, K.D.; Dubinsky, Z.; Porter, J.W.; Falkowski, P.G. Light absorption and utilization among hermatypic corals: A study in Jamaica, West Indies. Mar. Biol. 1987, 96, 283–292. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Jokiel, P.L.; Kinzie, R.A., III. Irradiance and Corals. In Coral Reefs. Ecosystems of the World; Dubinsky, Z., Ed.; Elsevier: Amsterdam, The Netherlands, 1990; Volume 25, pp. 89–107. [Google Scholar]
- Lesser, M.P.; Slattery, M.; Stat, M.; Ojimi, M.; Gates, R.D.; Grottoli, A. Photoacclimatization by the coral Montastraea cavernosa in the mesophotic zone: Light, food, and genetics. Ecology 2010, 91, 990–1003. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.; Mobley, C.D.; Hedley, J.D.; Slattery, M. Incident light on mesophotic corals is constrained by reef topography and colony morphology. Mar. Ecol. Prog. Ser. 2021, 670, 49–60. [Google Scholar] [CrossRef]
- Lesser, M.; Slattery, M.; Mobley, C.D. Incident light and morphology determine coral productivity along a shallow to mesophotic depth gradient. Ecol. Evol. 2021, 11, 13445–13454. [Google Scholar] [CrossRef] [PubMed]
- Enríques, S.; Méndez, E.R.; Iglesias-Prieto, R. Multiple scattering on coral skeletons enhances light absorption by symbiotic algae. Limnol. Oceanogr. 2005, 50, 1025–1032. [Google Scholar] [CrossRef]
- Enríques, S.; Méndez, E.R.; Hoegh-Guldberg, O.; Iglesias-Prieto, R. Key functional role of the optical properties of coral skeletons in coral ecology and evolution. Proc. Roy. Soc. B. 2017, 284, 20161667. [Google Scholar] [CrossRef]
- Kühl, M.; Cohen, Y.; Dalsgaard, T.; Jørgensen, B.B.; Revsbech, N.P. Microenvironment and photosynthesis of zooxanthellae in scleractinian corals studied with microsensors for O2, pH and light. Mar. Ecol. Prog. Ser. 1995, 117, 159–172. [Google Scholar] [CrossRef]
- Broderson, K.E.; Lichtenberg, M.; Ralph, P.J.; Kühl, M.; Wangpraseurt, D. Radiative energy budgets reveals high photosynthetic efficiency in symbiont-bearing coral. J. R. Soc. Interface 2014, 11, 20130997. [Google Scholar] [CrossRef]
- Wangpraseurt, D.; Larkum, A.W.D.; Ralph, P.J.; Kühl, M. Light gradients and optical microniches in coral tissues. Front. Microbiol. 2012, 3, 316. [Google Scholar] [CrossRef] [PubMed]
- Mass, T.; Einbinder, S.; Brokovich, E.; Shashar, N.; Vago, R.; Erez, J.; Dubinsky, Z. Photoacclimation of Stylophora pistillata to light extremes: Metabolism and calcification. Mar. Ecol. Prog. Ser. 2007, 334, 93–102. [Google Scholar] [CrossRef]
- Lesser, M.P.; Mazel, C.; Phinney, D.; Yentsch, C.S. Light absorption and utilization by colonies of the congeneric hermatypic corals Montastraea faveolata and Montastraea cavernosa. Limnol. Oceanogr. 2000, 45, 76–86. [Google Scholar] [CrossRef]
- Einbinder, S.; Gruber, D.F.; Salomon, E.; Liran, O.; Keren, N.; Tchernov, D. Novel adaptive photosynthetic characteristics of mesophotic symbiotic microalgae within the reef-building coral, Stylophota pistillata. Front. Mar. Sci. 2016, 3, 195. [Google Scholar] [CrossRef]
- Padilla-Gamiño, J.L.; Roth, M.S.; Rodrigues, L.J.; Bradley, C.J.; Bidigare, R.R.; Gates, R.D.; Smith, C.M.; Spalding, H.L. Ecophysiology of mesophotic reef-building corals in Hawai’i is influenced by symbiont-host associations, photoacclimatization, trophic plasticity, and adaptation. Limnol. Oceanogr. 2019, 64, 1980–1995. [Google Scholar] [CrossRef]
- Ben-Zvi, O.; Wangpraseurt, D.; Bronstein, O.; Eyal, G.; Loya, Y. Photosynthesis and bio-optical properties of fluorescent mesophotic corals. Front. Mar. Sci. 2021, 8, 651601. [Google Scholar] [CrossRef]
- Kramer, N.; Tamir, R.; Ben-Zvi, O.; Jacques, S.L.; Loya, Y.; Wangpraseurt, D. Efficient light-harvesting of mesophotic corals is facilitated by coral optical traits. Funct. Ecol. 2022, 36, 406–418. [Google Scholar] [CrossRef]
- Ferrier-Pagés, C.; Leal, M.G. Stable isotopes as tracers of trophic interactions in mutualistic symbioses. Ecol. Evol. 2019, 9, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Houlbrèque, F.; Ferrier-Pagès, C. Heterotrophy in tropical scleractinian corals. Biol. Rev. 2009, 84, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Conti-Jerpe, I.E.; Thompson, P.D.; Wong, C.W.M.; Oliveira, N.L.; Duprey, N.N.; Moynihan, M.A.; Baker, D.M. Trophic strategy and bleaching resistance in reef-building corals. Sci. Adv. 2020, 6, eaaz5443. [Google Scholar] [CrossRef]
- Kahng, S.E.; Garcia-Sais, J.R.; Spalding, H.L.; Brokovich, E.; Wagner, D.; Weil, E.; Hinderstein, L.; Toonen, R.J. Community ecology of mesophotic coral reef ecosystems. Coral Reefs. 2010, 29, 255–275. [Google Scholar] [CrossRef]
- Kahng, S.E.; Copus, J.M.; Wagner, D. Recent advances in the ecology of mesophotic coral ecosystems (MCEs). Curr. Opin. Environ. Sustain. 2014, 7, 72–81. [Google Scholar] [CrossRef]
- Kahng, S.E.; Akkaynak, D.; Shlesinger, T.; Hochberg, E.J.; Wiedenmann, J.; Tamir, R.; Tchernov, D. Light, temperature, photosynthesis, heterotrophy, and the lower depth limits of mesophotic coral ecosystems. In Mesophotic Coral Ecosystems. Coral Reefs of the World; Loya, Y., Puglise, K.A., Bridge, T., Eds.; Springer International: Cham, Switzerland, 2019; pp. 801–828. [Google Scholar]
- Wall, C.B.; Ritson-Williams, R.; Popp, B.N.; Gates, R.D. Spatial variation in the biochemical and isotopic composition of corals during bleaching and recovery. Limnol. Oceanogr. 2019, 64, 2011–2028. [Google Scholar] [CrossRef]
- Swart, P.K.; Szmant, A.; Porter, J.W.; Dodge, R.E.; Tougas, J.I.; Southam, J.R. Isotopic composition of respired carbon dioxide in scleractinian corals: Implications for cycling of organic carbon in corals. Geochim. Cosmochim. Acta 2005, 69, 1495–1509. [Google Scholar] [CrossRef]
- Alamaru, A.; Loya, Y.; Brokovich, E.; Yam, R.; Shemesh, A. Carbon and nitrogen utilization in two species of Red Sea corals along a depth gradient: Insights from stable isotope analysis of total organic material and lipids. Geochim. Cosmochim. Acta 2009, 73, 5333–5342. [Google Scholar] [CrossRef]
- Tremblay, P.; Maguer, J.F.; Grover, R.; Ferrier-Pagès, C. Trophic dynamics of scleractinian corals: A stable isotope approach. J. Exp. Biol. 2015, 218, 1223–1234. [Google Scholar] [PubMed]
- Maier, C.; Weinbauer, M.G.; Pätzold, J. Stable isotopes reveal limitations in C and N assimilation in the Caribbean reef corals Madracis auretenra, M. carmabi and M. formosa. Mar. Ecol. Prog. Ser. 2010, 412, 103–112. [Google Scholar] [CrossRef]
- Tanaka, Y.; Grottoli, A.G.; Matsui, Y.; Suzuki, A.; Sakai, K. Effects of nitrate and phosphate availability on the tissues and carbonate skeleton of scleractinian corals. Mar. Ecol. Prog. Ser. 2017, 570, 101–112. [Google Scholar] [CrossRef]
- Wall, C.B.; Kaluhiokalani, M.; Popp, B.N.; Donahue, M.J.; Gates, R.D. Divergent symbiont communities determine the physiology and nutrition of a reef coral across a light-availability gradient. ISME J. 2020, 14, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Muscatine, L.; Porter, J.W.; Kaplan, I.R. Resource partitioning by reef corals as determined from stable isotope composition I. δ13C of zooxanthellae and animal tissue versus depth. Mar. Biol. 1989, 100, 185–193. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef]
- Lesser, M.P. Coral bleaching: Causes and mechanisms. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 405–419. [Google Scholar]
- Edmunds, P.J.; Spencer-Davies, P. Post-illumination stimulation of respiration rate in the coral Porites porites. Coral Reefs 1988, 7, 7–9. [Google Scholar] [CrossRef]
- Schrameyer, V.; Wangpraseurt, D.; Hill, R.; Kühl, M.; Larkum, A.W.D.; Ralph, P.J. Light respiratory processes and gross photosynthesis in two scleractinian corals. PLoS ONE 2014, 9, e110814. [Google Scholar] [CrossRef]
- Einbinder, S.; Mass, T.; Brokovich, E.; Dubinsky, Z.; Erez, J.; Tchernov, D. Changes in morphology and diet of the coral Stylophora pistillata along a depth gradient. Mar. Ecol. Prog. Ser. 2009, 381, 167–174. [Google Scholar] [CrossRef]
- Morrow, K.M.; Fiore, C.L.; Lesser, M.P. Environmental drivers of microbial community shifts in the giant barrel sponge, Xestospongia muta, over a shallow to mesophotic depth gradient. Environ. Microbiol. 2016, 18, 2025–2038. [Google Scholar] [CrossRef]
- Crandall, J.B.; Teece, M.A.; Estes, B.A.; Manfrino, C.; Ciesla, J.H. Nutrient acquisition strategies in mesophotic hard corals using compound specific stable isotope analysis of sterols. J. Exp. Mar. Biol. Ecol. 2016, 474, 133–141. [Google Scholar] [CrossRef]
- Price, J.T.; McLachlan, R.H.; Jury, C.P.; Toonen, R.J.; Grottoli, A.G. Isotopic approaches to estimating the contribution of heterotrophic sources to Hawaiian corals. Limnol. Oceanogr. 2021, 66, 2393–2407. [Google Scholar] [CrossRef]
- Laverick, J.H.; Green, T.K.; Burdett, H.L.; Newton, J.; Rogers, A.D. Depth alone is an inappropriate proxy for physiological change in the mesophotic coral Agaricia larmarki. J. Mar. Biol. Assoc. UK 2019, 99, 1535–1546. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER-Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Martinez, S.; Kolodny, Y.; Shemesh, E.; Scucchia, F.; Nevo, R.; Levin-Zaidman, S.; Paltiel, Y.; Keren, N.; Tchernov, D.; Mass, T. Energy sources of the depth-generalist mixotrophic coral Stylophora pistillata. Front. Mar. Sci. 2020, 7, 566663. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, E.J.; Atkinson, M.J. Coral reef benthic productivity based on optical absorptance and light-use efficiency. Coral Reefs 2008, 27, 49–59. [Google Scholar] [CrossRef]
- Fujii, T.; Tanaka, Y.; Maki, K.; Saotome, N.; Morimoto, N.; Watanabe, A.; Miyajima, T. Organic carbon and nitrogen isoscapes of reef corals and algal symbionts: Relative influences of environmental gradients and heterotrophy. Microorganisms 2020, 8, 1221. [Google Scholar] [CrossRef] [PubMed]
- Wall, C.B.; Wallsgrove, N.J.; Gates, R.D.; Popp, B.N. Amino acid δ13C and δ15N analyses reveal distinct species-specific patterns of trophic plasticity in a marine symbiosis. Limnol. Oceanogr. 2021, 66, 2033–2050. [Google Scholar] [CrossRef]
- Land, L.S.; Lang, J.C. On the stable carbon and oxygen isotopic composition of some shallow water, ahermatypic, scleractinian coral skeletons. Geochim. Cosmochim. Acta 1977, 41, 169–172. [Google Scholar] [CrossRef]
- Van Duyl, F.C.; Mueller, B.; Meesters, E.H. Spatio–temporal variation in stable isotope signatures (δ13C and δ15N) of sponges on the Saba Bank. Peer J. 2018, 6, e5460. [Google Scholar] [CrossRef]
- McConnaughey, T. 13C and 18O isotopic disequilibrium in biological carbonates: I. Patterns. Geochim. Cosmochim. Acta 1989, 53, 151–162. [Google Scholar] [CrossRef]
- Heikoop, J.M.; Dunn, J.J.; Risk, M.J.; Sandeman, I.M.; Schwarcz, H.P.; Waltho, N. Relationship between light and δ15N of coral tissue: Examples from Jamaica and Zanzibar. Limnol. Oceanogr. 1998, 43, 909–920. [Google Scholar] [CrossRef]
- Muscatine, L.; Kaplan, I.R. Resource partitioning by reef corals as determined from stable isotope composition II. δ15N of zooxanthellae and animal tissue versus depth. Pac. Sci. 1994, 48, 304–312. [Google Scholar]
- Swart, P.K.; Saied, A.; Lamb, K. Temporal and spatial variation in the δ15N and δ13C of coral tissue and zooxanthellae in Montastraea faveolata collected from the Florida reef tract. Limnol. Oceanogr. 2005, 50, 1049–1058. [Google Scholar] [CrossRef]
- Reynaud, S.; Martinez, P.; Houlbrèque, F.; Billy, I.; Allemand, D.; Ferrier-Pagès, C. Effect of light and feeding on the nitrogen isotopic composition of a zooxanthellate coral: Role of nitrogen cycling. Mar. Ecol. Prog. Ser. 2009, 393, 103–110. [Google Scholar] [CrossRef]
- Glaze, T.D.; Erler, D.V.; Siljanen, M.P. Microbially facilitated nitrogen cycling in tropical corals. ISME J. 2021, 16, 68–77. [Google Scholar] [CrossRef]
- Tanaka, Y.; Suzuki, A.; Sakai, K. The stoichiometry of coral-dinoflagellate symbiosis: Carbon and nitrogen cycles are balanced in the recycling and double translocation system. ISME J. 2018, 12, 860–868. [Google Scholar] [CrossRef]
- Grottoli, A.G.; Wellington, G.M. Effect of light and zooplankton on skeletal δ13C values in the eastern Pacific corals Pavona clavus and Pavona gigantea. Coral Reefs 1999, 18, 29–41. [Google Scholar] [CrossRef]
- Heikoop, J.M.; Dunn, J.J.; Risk, M.J.; Schwarz, H.P.; McConnaughey, T.A.; Sandeman, I.M. Separation of kinetic and metabolic isotope effects in carbo-13 records preserved in reef coral skeletons. Geochim. Cosmochim. Acta 2000, 64, 975–987. [Google Scholar] [CrossRef]
- Grottoli, A.G. Effect of light and brine shrimp levels on skeletal δ13C values in the Hawaiian coral Porites compressa: A tank experiment. Geochim. Cosmochim. Acta 2002, 66, 1955–1967. [Google Scholar] [CrossRef]
- Omata, T.; Suzuki, A.; Sato, T.; Minoshima, K.; Nomaru, E.; Murakami, A.; Muruyama, S.; Kawahata, H.; Maruyama, T. Effect of photosynthetic light dosage on carbon isotope composition in the coral skeleton: Long-term culture of Porites spp. J. Geophys. Res. 2008, 113, G02014. [Google Scholar]
- Linsley, B.K.; Dunbar, R.B.; Dassié, E.P.; Tangri, N.; Wu, H.C.; Brenner, L.D.; Wellington, G.M. Coral carbon isotope sensitivity to growth rate and water depth with paleo-sea level implications. Nat. Commun. 2019, 10, 2056. [Google Scholar] [CrossRef] [PubMed]
- McConnaughey, T.A.; Burdett, J.; Whelan, J.F.; Paull, C.K. Carbon isotopes in biological carbonates: Respiration and photosynthesis. Geochim. Cosmochim. Acta 1997, 61, 611–622. [Google Scholar] [CrossRef]
- Schoepf, V.; Levas, S.J.; Rodrigues, L.J.; McBride, M.O.; Aschaffenburg, M.D.; Matsui, Y.; Warner, M.E.; Hughes, A.D.; Grottoli, A.G. Kinetic and metabolic isotope effects in coral skeletal carbon isotopes: A re-evaluation using experimental coral bleaching as a case study. Geochim. Cosmochim. Acta 2014, 146, 164–178. [Google Scholar] [CrossRef]
- Andradi-Brown, D.A.; Head, C.E.I.; Exton, D.A.; Hunt, C.L.; Hendrix, A.; Gress, E.; Rogers, A.D. Identifying zooplankton community changes between shallow and upper-mesophotic reefs on Mesoamerican Barrier, Caribbean. Peer J. 2017, 5, e2853. [Google Scholar] [CrossRef] [PubMed]
- Sponaugle, S.; Goldstein, E.; Doering, K.; D’Alessandro, E.; Guigand, C.; Cowen, R.K. Near-reef zooplankton differences across depths in a subtropical zone. J. Plankton Res. 2021, 43, 586–597. [Google Scholar] [CrossRef]
- Ezzat, L.; Maoz, F.; Maguer, J.-F.; Grover, R.; Ferrier-Pagès, C. Carbon and nitrogen acquisition in shallow and deep holobionts of the scleractinian coral, Stylophora pistillata. Front. Mar. Sci. 2017, 4, 102. [Google Scholar] [CrossRef]
- Fox, M.D.; Smith, E.A.E.; Smith, J.E.; Newsome, S.D. Trophic plasticity in a common reef-building coral: Insights from δ13C analysis of essential amino acids. Funct. Ecol. 2019, 33, 2203–2214. [Google Scholar] [CrossRef]
- Ferrier-Pagès, C.; Martinez, S.; Grover, R.; Cybulski, J.; Shemesh, E.; Tchernov, D. Tracing the trophic plasticity of the coral-dinoflagellate symbiosis using amino acid compound-specific stable isotope analysis. Microorganisms 2021, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.; Taylor, D.L.; Leigh, M.B.; O’Brien, D.M. Stable isotope fingerprinting: A novel method for identifying plant, fungal, or bacterial origins of amino acids. Ecology 2009, 90, 3526–3535. [Google Scholar] [CrossRef]
- Larsen, T.; Ventura, M.; Andersen, N.; O’Brien, D.M.; Piatkowski, U.; McCarthy, M.D. Tracing carbon sources through aquatic and terrestrial food webs using amino acid stable isotope fingerprinting. PLoS ONE 2013, 8, e73441. [Google Scholar] [CrossRef]
- McMahon, K.W.; Thorrold, S.R.; Houghton, L.A.; Berumen, M.L. Tracing carbon flow through coral reef food webs using a compound-specific stable isotope approach. Oecologia 2016, 180, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.; Hansen, T.; Dierking, J. Characterizing niche differentiation among marine consumers with amino acid δ13C fingerprinting. Ecol. Evol. 2020, 10, 7768–7782. [Google Scholar] [CrossRef]
- Macartney, K.J.; Slattery, M.; Lesser, M.P. Trophic ecology of Caribbean sponges in the mesophotic zone. Limnol. Oceanogr. 2021, 66, 1113–1124. [Google Scholar] [CrossRef]
- McClelland, J.W.; Montoya, J.P. Trophic relationships and the nitrogen isotopic composition of amino acids in plankton. Ecology 2002, 83, 2173–2180. [Google Scholar] [CrossRef]
- Chikaraishi, Y.; Ogawa, N.O.; Kashiyama, Y.; Takano, Y.; Suga, H.; Tomitani, A.; Miyashita, H.; Kitazato, H.; Ohkouchi, N. Determination of aquatic food-web structure based on compound-specific nitrogen isotopic composition of amino acids. Limnol. Oceanogr. Methods 2009, 7, 740–750. [Google Scholar] [CrossRef]
- Farstey, V.; Lazar, B.; Genin, A. Expansion and homogeneity of the vertical distribution of zooplankton in a very deep mixed layer. Mar. Ecol. Prog. Ser. 2002, 238, 91–100. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lesser, M.P.; Slattery, M.; Macartney, K.J. Using Stable Isotope Analyses to Assess the Trophic Ecology of Scleractinian Corals. Oceans 2022, 3, 527-546. https://doi.org/10.3390/oceans3040035
Lesser MP, Slattery M, Macartney KJ. Using Stable Isotope Analyses to Assess the Trophic Ecology of Scleractinian Corals. Oceans. 2022; 3(4):527-546. https://doi.org/10.3390/oceans3040035
Chicago/Turabian StyleLesser, Michael P., Marc Slattery, and Keir J. Macartney. 2022. "Using Stable Isotope Analyses to Assess the Trophic Ecology of Scleractinian Corals" Oceans 3, no. 4: 527-546. https://doi.org/10.3390/oceans3040035
APA StyleLesser, M. P., Slattery, M., & Macartney, K. J. (2022). Using Stable Isotope Analyses to Assess the Trophic Ecology of Scleractinian Corals. Oceans, 3(4), 527-546. https://doi.org/10.3390/oceans3040035