





The Final Pliocene and Early Pleistocene Faunal Dispersals from East to Europe and Correlation of the Villafranchian Biochronology between Eastern and Western Europe

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. The Faunal Dispersals as Key Bioevents for the Formation and the Evolution of the Villafranchian Faunas of Europe
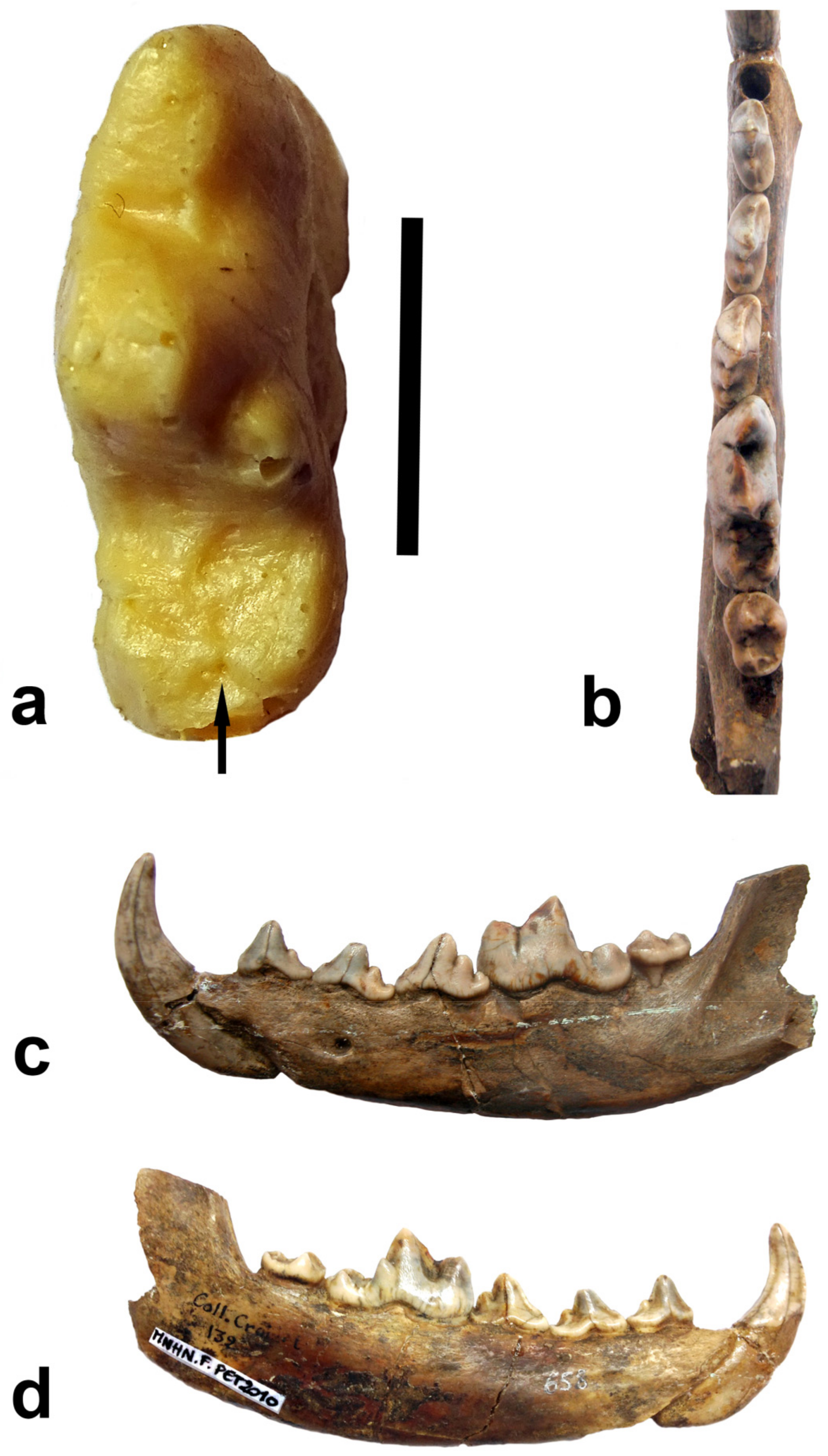
- The earliest appearance of the genus Canis in Europe: Lacombat et al. [61] mention several fragmentary remains of mandibles from Vialette, whose age is estimated (see there) to be 3.14 Ma and refer them to Canis sp. (Canis has been mentioned in this Late Pliocene locality since Heintz et al. [62]). Following the publication of Lacombat et al. [61], the first occurrence of the genus Canis in Europe is generally accepted to be associated with this Late Pliocene locality (Sotnikova, Rook [7]; Rook and Martínez-Navarro, [4]; Palombo [27]). Iannucci et al. [31], however, note the existence of chronological heterogeneity of the fauna preserved in the old collections (Crozatier Museum) labeled as Vialette. At the same time, we [1] (Spassov in Böhme et al.) expressed the opinion that the canid from Vialette most likely represents the genus Eucyon, known in Europe from the latest Miocene until the Pleistocene beginning. The most informative is the mandibular fragment (2003-5-401-VIA) with m1-m2. The analysis of a cast, kindly provided to me (2006) by A. Monguillon ( Université de Lyon), enabled me to draw some conclusions. The canid is really large, and its dental dimensions are similar to those of a coyote (Lm1 = 21 mm; p4 = 11.7 × 5 mm; p3 = 10.2 × 4 mm, after cast). This is also reminiscent of the possible Eucyon—“Canis” michauxi (Martin [63]). The lower carnassial tooth of this enigmatic canid from the Pliocene of Perpignan is not known, but the preserved p3-p4 (incorrectly represented in Martin’s [63] figure as p2-p3) are even larger. The talonid of m1 of the specimen 2003-5-401-VIA is heavily eroded and of incomplete relief, but as seen in Figure 1, the hypoconid and the entoconid do not contact at their bases. This morphology should correspond to the state of the talonid in Eucyon; in Canis well-developed cristids that contact and fuse at their bases descend from the hypoconid and the entoconid towards each other (Tedford and Qiu [64]).
- 2.
- The mass appearance of the genus: In Western Europe (France, Italy, Spain), the Canis mass appearance occurred in the period of about 2.2–1.98 Ma (Palombo [27]), but this can be said for all of Europe in general. We can call this dispersal, using Azzaroli’s concept [16], Canis event s. str. or mass Canis appearance in Europe. The earliest Canis mass arrival on the continent must be related to the Coste San Giacomo unit. We can claim that the following localities (starting from east to west) record practically the same wave of dispersal of the “wolf-like” Canis to Europe: Slivnitsa, Bulgaria (see below: Spassov [11,72]), Coste San Giacomo (2.2 Ma: Florindo et al. [73]) and Quercia (2.2–2.1 Ma: Iannucci [74], Italy), and Senèze, France (its base age is of ca. 2.2 and the youngest fossils ranged in age between 2.10 and 2.08 Ma after a new investigation in. prep.: Eric Delson, pers. comm.). It seems that these localities, which record the first secure mass invasion of the genus (Spassov [11,72,75]; Rook, Martἱnez-Navarro [4]; Iannucci [74]) have a very similar/close age and should be placed in the C. San Giacomo Unit. As it seems, two species entered practically simultaneously from the East to Europe (C. etruscus and C. arnensis are apparently present simultaneously in the locality of Slivnitsa; Spassov [72]), and two species of Canis are detected in Senèze after a new investigation (chapter in press on Senèze carnivores: A. Argant, pers. comm.). Slivnitsa and Senèze have very similar fauna and should be of the same/very close age (Spassov [75]). They show other mass dispersals, as well (see below). The Slivnitsa faunal event must, therefore, correlate (Spassov [34]) with the climatochronologic zone SCT10 of Zubakov and Borzenkova [76]. This zone, documented in Georgia and the Azov region, has an estimated age predating the Olduvai warming (between the Reunion subchron [2.193 Ma] and the beginning of the Olduvai event [ca. 1.95 Ma]).
2.2. Major Villafranchian Events. A Summary
2.3. Correlation of the Villafranchian Biochronology between Eastern and Western Europe (Correlation of Selected Eastern European Localities)
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Böhme, M.; Spassov, N.; Majidifard, M.; Gärtner, A.; Kirscher, U.; Marks, M.; Dietzel, C.; Uhlig, G.; El Atfy, K.; Begun, D.B.; et al. Neogene hyperaridity in Arabia drove the directions of mammalian dispersal between Africa and Eurasia. Commun. Earth Environ. 2021, 2, 85. [Google Scholar] [CrossRef]
- Van Asperen, E.; Kahlke, R.-D. Dietary traits of the late Early Pleistocene Bison menneri (Bovidae, Mammalia) from its type site Untermassfeld (Central Germany) and the problem of Pleistocene ‘wood bison’. Quat. Sci. Rev. 2017, 177, 299–313. [Google Scholar] [CrossRef]
- Spassov, N. The Mammalian Megafauna from the Late Villafranchian Localities Varshets and Sivnitsa (Bulgaria) and the Biochronology of the Villafranchian in S.-E. Europe. Ph.D. Thesis, National Museum Natural History, Bulgarian Academy of Sciences, Sofia, Bulgaria, 1999; p. 284. (In Bulgarian). [Google Scholar]
- Rook, L.; Martínez-Navarro, B. Villafranchian: The long story of a Plio-Pleistocene European large mammal biochronologic unit. Quat. Int. 2010, 219, 134–144. [Google Scholar] [CrossRef]
- Vangengeim, E.; Sotnikova, M. Paleozoogeographic characteristics of the fauna. In Biostratigraphy of the Late Pliocene–the Early Pleistocene of Tajikistan; Nikiforova, K., Vangengeim, E., Eds.; Nauka: Moscow, Russia, 1988; pp. 103–114. [Google Scholar]
- Sotnikova, M. Carnivore Mammals of the Pliocene and the Early Pleistocene; Nauka: Moskow, Russia, 1989; p. 122. [Google Scholar]
- Sotnikova, M.; Rook, L. Dispersal of the Canini (Mammalia, Canidae: Caninae) across Eurasia during the Late Miocene to Early Pleistocene. Quat. Int. 2010, 212, 86–97. [Google Scholar] [CrossRef]
- Azzaroli, A. Villafranchian correlations based on large mammals. G. Geol. 1970, 35, 111–131. [Google Scholar]
- Azzaroli, A. The villafranchian stage in Italy and the Plio–Pleistocene boundary. G. Geol. 1977, 41, 61–79. [Google Scholar]
- Spassov, N. The Villafranchian mammalian fauna and its investigation in Europe, on the Balkans and in Bulgaria. Geol. Balc. 1997, 27, 83–94. [Google Scholar]
- Spassov, N. The Plio-Pleistocene vertebrate fauna in South-Eastern Europe and the megafaunal migratory waves from the east to Europe. Rev. Paleobiol. 2003, 22, 197–229. [Google Scholar]
- Lindsay, E. Chronostratigraphy, Biochronology, Datum Events, Land Mammal Ages, Stage of Evolution, and Appearance Event Ordination. Bull. Am. Mus. Nat. Hist. 2003, 212–230. [Google Scholar] [CrossRef]
- Mein, P. Résultats du groupe de travail des Vertébrés. In Report on Activity on the RCMNS Working Group (1971–1975); VIth Congress of the Regional Committee on Mediterranean Neogene Stratigraphy; Bratislava, Czechoslovakia, 1975; pp. 78–81. [Google Scholar]
- Guérin, C. Premiere Biozonation du Pleistocene Europeen, principal resultat biostratigraphique de l’etude des Rhinocerotidae du miocene terminal au pleistocene superieur d’Europe Occidentale. Geobios 1982, 15, 593–598. [Google Scholar] [CrossRef]
- Guérin, C. Biozones or Mammal Units? Methods and limits in Biochronology. In European Neogene Mammal Chronologt; Lindsay, E.H., Fahlbusch, V., Mein, P., Eds.; Plenum Press: New York, NY, USA, 1990; pp. 119–130. [Google Scholar]
- Azzaroli, A. Quaternary mammals and the end-villafranchian dispersal Event—A turning point in the history of Eurasia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1983, 44, 117–139. [Google Scholar] [CrossRef]
- Torre, D.; Ficcarelli, G.; Masini, F.; Rook, L.; Sala, B. Mammal dispersal events in the early Pleistocene of Western Europe. Cour. Forschungsinstitut Senckenberg 1992, 153, 51–58. [Google Scholar]
- Gliozzi, E.; Abbazzi, L.; Argenti, P.; Azzaroli, A.; Caloi, L.; Capasso Barbato, L.; Di Stefano, G.; Esu, D.; Ficcarelli, G.; Girotti, O.; et al. Biochronology of selected mammals, molluscs and ostracods from the Middle Pliocene to the Late Pleistocene in Italy. The state of the art. Riv. Ital. Paleontol. Stratigr. 1997, 103, 369–388. [Google Scholar] [CrossRef]
- Nomade, S.; Pastre, J.; Guillou, H.; Faure, M.; Guérin, C.; Delson, E.; Debard, E.; Voinchet, P.; Messager, E. 40Ar/39Ar constraints on some French Landmark Late Pliocene to Early Pleistocene large mammalian paleofaunas, paleoenvironmental and paleoecological implications. Quat. Geochronol. 2014, 421, 2–15. [Google Scholar] [CrossRef]
- Lindsay, E.; Opdyke, N.; Johnson, N. Pliocene dispersal of the horse Equus and late Cenozoic mammalian dispersal events. Nature 1980, 287, 135–138. [Google Scholar] [CrossRef]
- Spassov, N. The Late Villafranchian and biochronology of South-East Europe: Faunal migrations, environmental changes and possible first appearance of the genus Homo on the continent. In On Late Plio/Pleistocene Extinctions and Evolution in the Palearctic: The Vatera Site, Proceedings of the Ist International Workshop, Vatera, Lesvos, Greece, 29 September–2 October 1999; 2002; Annales Geologiques des Pays Hellenique, 1 Série, A; Volume 39, pp. 221–243. [Google Scholar]
- Cregut-Bonnoure, É. Apport des Caprinae et Antilopinae (Mammalia, Bovidae) à la biostratigraphie du Pliocène terminal et du Pléistocène d’Europe. Quaternaire 2007, 18, 73–97. [Google Scholar] [CrossRef]
- Sardella, R.; Palombo, M. The Pliocene-Pleistocene boundary: Which significance for the so called “Wolf event”? evidences from Western Europe. Quaternaire 2007, 18, 65–71. [Google Scholar] [CrossRef]
- Palombo, M.; Sardella, R.; Novelli, M. Carnivora dispersal in Western Mediterranean during the last 2.6Ma. Quat. Int. 2008, 179, 176–189. [Google Scholar] [CrossRef]
- Croitor, R.; Brugal, J.-P. Ecological and evolutionary dynamics of the carnivore community in Europe during the last 3 million years. Quat. Int. 2010, 212, 98–108. [Google Scholar] [CrossRef]
- O’Regan, H.; Turner, A.; Bishop, L.; Elton, S.; Lamb, A. Hominins without fellow travellers? First appearances and inferred dispersals of Afro-Eurasian large-mammals in the Plio-Pleistocene. Quat. Sci. Rev. 2011, 30, 1343–1352. [Google Scholar] [CrossRef]
- Palombo, M. Deconstructing mammal dispersals and faunal dynamics in SW Europe during the Quaternary. Quat. Sci. Rev. 2014, 96, 50–71. [Google Scholar] [CrossRef]
- Palombo, M. Discrete dispersal bioevents of large mammals in Southern Europe in the post-Olduvai Early Pleistocene: A critical overview. Quat. Int. 2017, 431, 3–19. [Google Scholar] [CrossRef]
- Koufos, G.; Kostopoulos, D. The Plio-Pleistocene Large Mammal Record of Greece: Implications for Early Human Dispersals into Europe. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Vertebrate Paleobiology and Paleoanthropology Series; Springer Science+Business Media: Dordrecht, The Netherlands, 2016; pp. 269–281. [Google Scholar]
- Iannucci, A.; Mecozzi, B.; Sardella, R.; Iurino, D. The extinction of the giant hyena Pachycrocuta brevirostris and a reappraisal of the Epivillafranchian and Galerian Hyaenidae in Europe: Faunal turnover during the Early–Middle Pleistocene Transition. Quat. Sci. Rev. 2021, 272, 107240. [Google Scholar] [CrossRef]
- Iannucci, A.; Mecozzi, B.; Sardella, R. Beware of the “Wolf event”—Remarks on large mammal dispersals in Europe and the late Villafranchian faunal turnover. Alp. Mediterr. Quat. 2023, 36, 75–90. [Google Scholar]
- Samson, P. Les Equidés fossils de Roumanie (Pliocène moyen—Pléistocène supérieur). Geol. Romana 1975, 14, 165–352. [Google Scholar]
- Radulescu, C.; Samson, P. Biochronology and evolution of the Early Pliocene to the Early Pleistocene mammalian faunas of Romania. Boll. Della Soc. Paleontol. Ital. 2001, 40, 285–291. [Google Scholar]
- Spassov, N. Southeastern Europe as a Route for the Earliest Dispersal of Homo Toward Europe: Ecological Conditions and the Timing of the First Human Occupation of Europe. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Vertebrate Paleobiology and Paleoanthropology Series; Springer Science+Business Media: Dordrecht, The Netherlands, 2016; pp. 281–290. [Google Scholar] [CrossRef]
- Palombo, M.; Valli, A.; Kostopoulos, D.; Alberdi, M.; Spassov, N.; Vislobokova, I. Similarity Relationships between the Pliocene to Middle Pleistocene Large Mammal Faunas of Southern Europe from Spain to the Balkans and the North Pontic Region. Cour. Forsch. Inst. Senckenberg 2006, 256, 329–347. [Google Scholar]
- Kostopoulos, D.; Palombo, M.; Alberdi, M.; Valli, A. Pliocene to Pleistocene large mammal diversity and turnover in North Mediterranean region: The Greek Peninsula with respect to the Iberian and Italian ones. Geodiversitas 2007, 29, 401–419. [Google Scholar]
- Kahlke, R.-D.; García, N.; Kostopoulos, D.; Lacombat, F.; Lister, A.M.; Mazza, P.; Spassov, N.; Titov, V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Andreescu, I.; Codrea, V.; Lubenescu, V.; Munteanu, T.; Petculescu, A.; Stiucǎ, E.; Terzea, E. New developments in the upper pliocene–pleistocene stratigraphic units of the dacian basin (eastern paratethys), Romania. Quat. Int. 2013, 284, 15–29. [Google Scholar] [CrossRef]
- Terhune, C.; Curran, S.; Croitor, R.; Dragușin, V.; Gaudin, T.; Petculescu, A.; Robinson, C.; Robu, M.; Werdelin, L. Early Pleistocene fauna of the Oltet¸ River Valley of Romania: Biochronological and biogeographic implications. Quat. Int. 2020, 533, 14–33. [Google Scholar] [CrossRef]
- Palombo, M.R. Large mammal faunal dynamics in southwestern Europe during the late Early Pleistocene: Implications for the biochronological assessment and correlation of mammalian faunas. Alp. Mediterr. Quat. 2016, 29, 143–168. [Google Scholar]
- Konidaris, G.E.; Kostopoulos, D.S. The Late Pliocene–Middle Pleistocene Large Mammal Faunal Units of Greece. Quaternary 2024, 7, 27. [Google Scholar] [CrossRef]
- Iannucci, A.; Sardella, R. What Does the “Elephant-Equus” Event Mean Today? Reflections on Mammal Dispersal Events around the Pliocene-Pleistocene Boundary and the Flexible Ambiguity of Biochronology. Quaternary 2023, 6, 16. [Google Scholar] [CrossRef]
- Radulesco, C.; Samson, P. Review of the Villafranchian s.s. faunas of Romania. Alp. Mediterr. Quat. 1995, 8, 377–382. [Google Scholar]
- Alexeeva, L. On the new theriocomplex in Northern Black Sea zone. Bull. Aacademy Sci. Georgian SSR 1977, 86, 485–488. (In Russian) [Google Scholar]
- Nikiforova, K.; Alexandrova, L.; Troubikhin, V.; Tchepalyga, A. Correlation of Pliocene and Eopleistocene deposits of the European part of the and Romania. The continental Upper Pliocene of the Black-Caspian region. Izv. Akad. Nauk. USSR Ser. Geol. 1986, 5–17. (In Russian) [Google Scholar]
- Markov, G.; Spassov, N. Primitive mammoths from Northeast Bulgaria in the context of the earliest mammoth migrations in Europe. In Advances in Vertebrate Paleontology “Hent to Pantha”; Petculescu, A., Stiuca, E., Eds.; Institute of Speleology of the Romanian Academy: Bucharest, Romania, 2003; pp. 53–58. [Google Scholar]
- Lister, A.M.; van Essen, H.E. Mammuthus rumanus the earliest mammoth in Europe. In Advances in Vertebrate Paleontology “Hent to Pantha”; Petculescu, A., Stiuca, E., Eds.; Institute of Speleology of the Romanian Academy: Bucharest, Romania, 2003; pp. 47–52. [Google Scholar]
- Markov, G. Mammuthus rumanus, early mammoths, and migration out of Africa: Some interrelated problems. Quat. Int. 2012, 276, 23–26. [Google Scholar] [CrossRef]
- Kostopoulos, D.; Koulidou, I. An early mammoth maxilla from north-western Greece. Quat. Int. 2015, 379, 155–163. [Google Scholar] [CrossRef]
- Rook, L.; Bernor, R.; Avilla, L.S.; Cirilli, O.; Flynn, L.; Jukar, A.; Sanders, W.; Scott, E.; Wang, X. Mammal Biochronology (Land Mammal Ages) Around the World from Late Miocene to Middle Pleistocene and Major Events in Horse Evolutionary History. Front. Ecol. Evol. 2019, 7, 278. [Google Scholar] [CrossRef]
- Sun, B.; Deng, T. The Equus Datum and the Early Radiation of Equus in China. Front. Ecol. Evol. 2019, 7, 429. [Google Scholar] [CrossRef]
- Bernor, R.; Cirilli, O.; Jukar, A.M.; Potts, R.; Buskianidze, M.; Rook, L. Evolution of Early Equus in Italy, Georgia, the Indian Subcontinent, East Africa, and the Origins of African Zebras. Front. Ecol. Evol. 2019, 7, 166. [Google Scholar] [CrossRef]
- Eisenmann, V. The equids from Liventsovka and other localities of the Khaprovskii Faunal Complex, Russia: A revision. Geobios 2022, 70, 17–33. [Google Scholar] [CrossRef]
- Spassov, N. Brief Review of the Pliocene Ungulate Fauna of Bulgaria. Quat. Hors-Sér. 2005, 2, 201–212. [Google Scholar]
- Radulescu, C.; Samson, P.; Petculescu, A.; Stiucã, E. Pliocene Large Mammals of Romania. Coloq. Paleontol. 2003, 1, 549–558. [Google Scholar]
- Forstén, A. A review of Equus stenonis Cocchi (Perissodactyla, Equidae) and related forms. Quat. Sci. Rev. 1999, 18, 1373–1408. [Google Scholar] [CrossRef]
- Forstén, A. Mitochondrial-DNA time-table and the evolution of Equus: Comparison of molecular and paleontological evidence. Ann. Zool. Fenn. 1991, 28, 301–309. [Google Scholar]
- Crespo, V.; Vasile, S.; Petculescu, A.; Rătoi, B.; Haiduc, B. The Early Pliocene small mammals (Eulipotyphla, Rodentia, Lagomorpha) from Berești and Mălușteni (eastern Romania): A fresh look at old collections. Earth Environ. Sci. Trans. R. Soc. Edinb. 2023, 114, 49–63. [Google Scholar] [CrossRef]
- Turner, A. Villafranchian-Galerian larger carnivores of Europe dispersions and extinctions. Cour. Forschungsinst. Senckenberg 1992, 153, 153–160. [Google Scholar]
- Bartolini-Lucenti, S.; Spassov, N. Cave canem! The earliest Canis (Xenocyon) (Canidae, Mammalia) of Europe: Taxonomic affinities and paleoecology of the fossil wild dogs. Quat. Sci. Rev. 2022, 276, 107315. [Google Scholar] [CrossRef]
- Lacombat, F.; Abbazzi, L.; Ferretti, M.; Martínez-Navarro, B.; Moullé, P.-E.; Palombo, M.; Rook, L.; Turner, A.; Valli, A.-M. New data on the Early Villafranchian fauna from Vialette (Haute-Loire, France) based on the collection of the Crozatier Museum (Le Puy-en-Velay, Haute-Loire, France). Quat. Int. 2008, 179, 64–71. [Google Scholar] [CrossRef]
- Heintz, E.; Guérin, C.; Martin, R.; Prat, A. Principaux gisements villafranchiens de France: Listes fauniques et biostratigraphie. Mémoires Bur. Rech. Géol. Min. 1974, 78, 169–182. [Google Scholar]
- Martin, R. Trois nouvelles espèces de Caninae (Canidae, Carnivora) des gisements plio-villafranchiens d’Europe. Doc. Lab. Géol. La Fac. Sci. Lyon. Notes Mém. 1973, 57, 87–96. [Google Scholar]
- Tedford, R.; Qiu, Z. A new canid genus from the Pliocene of Yushe, Shanxi Province. Vertebr. PalAsiatica 1996, 34, 27–40. [Google Scholar]
- Heintz, E. Les Cervides Villafranchiens de France et d’Espagne; Voll. I; Nouv. Ser., Ser. C; Muséum National d’Histoire Naturelle: Paris, France, 1970; Volume 34, p. 303. [Google Scholar]
- Spassov, N. A New Late Villafranchian Locality of vertebrate Fauna—Slivnitsa (Bulgaria) and the Carnivore Dispersal Events in Europe on the Pliocene/Pleistocene Boundary. Hist. Nat. Bulg. 1998, 9, 101–113. [Google Scholar]
- Torre, D. The Ruscinian and the Villafranchian dogs of Europe. Boll. Della Soc. Paleontol. Ital. 1979, 18, 162–165. [Google Scholar]
- Fosse, P.; Madelaine, S.; Oberlin, C.; Cretin, C.; Bonnet-Jacquement, P. Un lion des cavernes (Panthera (Leo) spelaea) exploité au Dryas récent: Les données du gisement du Peyrat (Saint-Rabier, Dordogne, France). Paleo 2017, 28, 91–115. [Google Scholar] [CrossRef]
- Marciszak, A.; Kropczyk, A.; Gornig, W.; Kot, M.; Nadachowski, A.; Lipecki, G. History of Polish Canidae (Carnivora, Mammalia) and Their Biochronological Implications on the Eurasian Background. Genes 2023, 14, 539. [Google Scholar] [CrossRef]
- Argant, A. Les carnivores du gisement Pliocène final de Saint-Vallier (Drôme, France). Geobios 2004, 37, 133–182. [Google Scholar] [CrossRef]
- Monguillon-Douillet, A. Les carnivores du site plio-pleistocène de Chilhac (Haute-Loire, France). In Etude Paléontologique, Biochronologique, Paléoécologique et Taphonomique; Unpublished Mémoire de D.E.A. Quaternaire et Géologie; Muséum National d’Histoire Naturelle: Paris, France, 2000. [Google Scholar]
- Spassov, N. The position of jackals in the Canis Genus and life-history of the golden jackal (Canis aureus L.) in Bulgaria and the Balkans. Hist. Nat. Bulg. 1989, 1, 44–56. [Google Scholar]
- Florindo, F.; Marra1, F.; Angelucci, D.; Biddittu, I.; Bruni, L.; Florindo, F.; Gaeta, M.; Guillou, H.; Jicha, L.; Macrì, P.; et al. Environmental evolution, faunal and human occupation since 2 Ma in the Anagni basin, central Italy. Sci. Rep. 2021, 11, 7056. [Google Scholar] [CrossRef] [PubMed]
- Iannucci, A. Sus strozzii (Suidae, Mammalia) from the historical locality of Quercia (Early Pleistocene, Italy). Geobios 2023, 77, 27–36. [Google Scholar] [CrossRef]
- Spassov, N. Biochronology and zoogeographic affinities of the Villafranchian faunas of South Europe. Historia naturalis bulgarica. 2000, 12, 89–128. [Google Scholar]
- Zubakov, V.; Borzenkova, I. Global Palaeoclimate of the Late Cenozoic. Developments in palaeontology and stratigraphy. Amst. Elsevier 1990, 12, 456. [Google Scholar]
- Titov, V. Late Pliocene Large Mammals from Northeastern Sea of Azov Region; Matishov, G.G., Ed.; SSC RAS Publishing: Rostov-on-Don, Russia, 2008; p. 264. (In Russian) [Google Scholar]
- Sotnikova, M.; Bajgusheva, V.; Titov, V. Carnivores of the Khapry faunal assemblage and their stratigraphic implications. Stratigr. Geol. Correl. 2002, 10, 375–390. [Google Scholar]
- Werdelin, L.; Dehghani, R. Carnivora. In Paleontology and Geology of Laetoli: Human Evolution in Context; Harrison, T., Ed.; Vertebrate Paleobiology and Paleoanthropology Series; Springer: Dordrecht, The Netherlands, 2011; pp. 189–232. [Google Scholar] [CrossRef]
- Hemmer, H.; Kahlke, R.-D.; Vekua, A.K. The cheetah Acinonyx pardinensis (Croizet et Jobert, 1828) s.l. at the hominin site of Dmanisi (Georgia)—A potential prime meat supplier in Early Pleistocene ecosystems. Quat. Sci. Rev. 2011, 30, 2703–2714. [Google Scholar] [CrossRef]
- Cherin, M.; Iurino, D.; Sardella, R.; Rook, L. Acinonyx pardinensis (Carnivora, Felidae) from the Early Pleistocene of Pantalla (Italy): Predatory behavior and ecological role of the giant PlioePleistocene cheetah. Quat. Sci. Rev. 2014, 87, 82–97. [Google Scholar] [CrossRef]
- Hugueney, M.; Guérin, C.; Poidevin, J.L. Découverte de Trogontherium minus Newton 1890 (Rodentia: Castoridae) dans le Villafranchien inférieur de Perrier- Étouaires (Puy-de-Dôme, France): Implications phylogénétiques. C. R. Acad. Sci. Ser. II 1989, 309, 763–768. [Google Scholar]
- Hemmer, H.; Kahlke, R.-D.; Vekua, A. The Old World puma—Puma pardoides (Owen, 1846) (Carnivora: Felidae)—In the Lower Villafranchian (Upper Pliocene) of Kvabebi (East Georgia, Transcaucasia) and its evolutionary and biogeographical significance. Neues Jahrb. Geol. Paläontol. Abh. 2004, 233, 197–231. [Google Scholar] [CrossRef]
- Hemmer, H.; Kahlke, R.-D. New results on felids from the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld near Meiningen; Kahlke, R.-D., Ed.; Part 5; Verlag des Römisch-Germanischen Zentralmuseums: Weimar, Germany, 2022; pp. 1465–1566. [Google Scholar]
- Cherin, M.; Iurino, D.; Sardella, R.; Rook, L. Earliest occurrence of Puma pardoides (Owen, 1846) (Carnivora, Felidae) at the Plio/Pleistocene transition in western Europe: New evidence from the Middle Villafranchian assemblage of Montopoli, Italy. C. R. Palevol 2013, 12, 165–171. [Google Scholar] [CrossRef]
- Sotnikova, M. Verhnepliotsenovye Carnivora Tsentralnoy Azii (The Late Pliocene carnivores of Central Asia). Izv. Akad. Nauk. SSSR. Seriya Geol. 1976, 11, 133–137. (In Russian) [Google Scholar]
- Hemmer, H. Zur Charakterisierung und stratigraphischen Bedeutung von Panthera gombaszoegensis (Kretzoi, 1938). Neues Jahrb. Geol. Paläontol. Monatshefte 1971, 12, 701–711. [Google Scholar]
- Hemmer, H. Die Feliden aus dem Epivillafranchium von Untermaßfeld. In Das Pleistozän von Untermaßfeld bei Meiningen (Thüringen). Teil 3; Kahlke, R.-D., Ed.; Des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2001; Volume 40, pp. 699–782. [Google Scholar]
- Jiangzuo, Q.; Liu, J. First record of the Eurasian jaguar in southern Asia and a review of dental differences between pantherine cats. J. Quat. Sci. 2020, 35, 817–830. [Google Scholar] [CrossRef]
- Wagner, J. A short nomenclatural note to correct spelling of the species-group name “gombaszögensis”. Quat. Hors-Ser. 2011, 4, 245. [Google Scholar]
- van den Hoek Ostende, L.; de Vos, J. A century of research on the classical locality of Tegelen (province of Limburg, The Netherlands). Cour. Forsch.-Inst. Senckenberg 2006, 256, 291–304. [Google Scholar]
- Koufos, G. The Pleistocene carnivores of the Mygdonia basin (Macedonia, Greece). Ann. Paléontol. 1992, 78, 205–257. [Google Scholar]
- Martínez-Navarro, B. Early Pleistocene faunas of Eurasia and hominid dispersals. In Out of Africa I: The First Hominin Colonization of Eurasia; Fleagle, J., Shea, J., Grine, F., Baden, A., Leakey, R., Eds.; Vertebrate Paleobiology and Paleoanthropology Series; Springer: Dordrecht, The Netherlands, 2010; pp. 207–224. [Google Scholar]
- Croitor, R.; Robinson, C.; Curran, S.; Terhune, C.; Drăgușin, V.; Pobiner, B.; Popescu, D.; Petculescu, A. Early pleistocene ruminants (Artiodactyla, Mammalia) from the Dacian Basin (South Romania) before and after the Pachycrocuta event: Implications for hominin dispersals in Western Eurasia. Hist. Biol. 2023, 36, 485–533. [Google Scholar] [CrossRef]
- Sianis, P.; Athanassiou, A.; Roussiakis, S.; Iliopoulos, G. Carnivora from the Early Pleistocene locality of Karnezeika (Southern Greece). Geobios 2023, 79, 43–59. [Google Scholar] [CrossRef]
- Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.; Alba, D.; Aurell-Garrido, J. Villafranchian large mammals from the Iberian Peninsula: Paleobiogeography, paleoecology and dispersal events. J. Iber. Geol. 2014, 40, 167–178. [Google Scholar] [CrossRef]
- Koufos, G. The Villafranchian carnivoran guild of Greece: Implications for the fauna, biochronology and paleoecology. Integr. Zool. 2014, 9, 444–460. [Google Scholar] [CrossRef]
- Napoleone, G.; Albianelli, A.; Azzaroli, A.; Bertini, A.; Magi, M.; Mazzini, M. Calibration of the Upper Valdarno basin to the Plio-Pleistocene for correlating the Apennine continental Sequences. Alp. Mediterr. Quat. 2003, 16, 131–166. [Google Scholar]
- Bellucci, L.; Mazzini, I.; Scardia, G.; Bruni, L.; Parenti, F.; Segre, A.G.; Segre Naldini, E.; Sardella, R. The site of Coste San Giacomo (Early Pleistocene, Central Italy): Palaeoenvironmental analysis and biochronological overview. Quat. Int. 2012, 267, 30–39. [Google Scholar] [CrossRef]
- Fidalgo, D.; Galli, E.; Madurell-Malapeira, J.; Rosas, A. Earliest Pleistocene European hippos: A review. Comun. Geol. 2021, 108, 65–69. [Google Scholar]
- Reimann, C.; Strauch, F. Ein Hippopotamus-Schädel aus dem Pliozän von Elis (Peloponnes, Griechenland). Neues Jahrb. Geol. Paläontol. Abh. 2008, 249, 203–222. [Google Scholar] [CrossRef]
- Athanassiou, A. The fossil record of continental hippopotamids (Mammalia: Artiodactyla: Hippopotamidae) in Greece. In Fossil Vertebrates of Greece, 2; Vlachos, E., Ed.; Springer Nature: Cham, Switzerland, 2022; pp. 281–289. [Google Scholar]
- Marra, F.; Pandolfi, L.; Petronio, C.; Di Stefano, G.; Gaeta, M.; Salari, L. Reassessing the sedimentary deposits and vertebrate assemblages from Ponte Galeria area (Rome, central Italy): An archive for the Middle Pleistocene faunas of Europe. Earth-Sci. Rev. 2018, 139, 104–122. [Google Scholar] [CrossRef]
- Mecozzi, B.; Iannucci, A.; Mancini, M.; Tentori, D.; Cavasinni, C.; Conti, J.; Messina, M.Y.; Sarra, A.; Sardella, R. Reinforcing the idea of an early dispersal of Hippopotamus amphibius in Europe: Restoration and multidisciplinary study of the skull from the Middle Pleistocene of Cava Montanari (Rome, central Italy). PLoS ONE 2023, 18, e0293405. [Google Scholar] [CrossRef]
- Mazza, P.; Rustioni, M. The fossil bear from Senèze (Southern France). Rend. Lincei 1994, 5, 17–26. [Google Scholar] [CrossRef]
- Mazza, P. If Hippopotamuses cannot swim, how did they colonize islands? Lethaia 2014, 47, 494–499. [Google Scholar] [CrossRef]
- Made, J.; Sahnouni, M.; Boulaghraief, K. Hippopotamus gorgops from El Kherba (Algeria) and the context of its biogeography. In Proceedings of the II Meeting of African Prehistory, Burgos, Spain, 15–16 April 2015; Sahnouni, M., Semaw, S., Rios Garaizar, J., Eds.; Consorcio CENIEH: Burgos, Spain, 2017; pp. 135–169. [Google Scholar]
- Parker, J. What Biome Do Hippos Live In? Build a Stash Newsletter. 16 March 2023. Available online: https://www.buildastash.com/post/what-biome-do-hippos-live-in (accessed on 10 February 2024).
- Koufos, G. The Villafranchian mammalian faunas and biochronology of Greece. Boll. Della Soc. Paleontol. Ital. 2001, 40, 217–223. [Google Scholar]
- Kahlke, R.-D. Late Early Pleistocene European large mammals and the concept of an Epivillafranchian biochron. In Late Neogene and Quaternary Biodiversity and Evolution: Regional Developments and Interregional Correlations, Proceedings of the 18th International Senckenberg Conference (VI International Palaeontological Colloquium in Weimar, 2004); Volume II; Kahlke, R.-D., Maul, L.C., Mazza, P., Eds.; 2007; Volume 259, pp. 265–278, Courier Forschungsinstitut Senckenberg. [Google Scholar]
- Bellucci, L.; Sardella, R.; Rook, L. Large mammal biochronology framework in Europe at Jaramillo: The Epivillafranchian as a formal biochron. Quat. Int. 2014, 389, 84–89. [Google Scholar] [CrossRef]
- Cherin, M.; D’Allestro, V.; Masini, F. New Bovid Remains from the Early Pleistocene of Umbria (Italy) and a Reappraisal of Leptobos merlai. J. Mamm. Evol. 2019, 26, 201–224. [Google Scholar] [CrossRef]
- Akbar Khan, M.; Kostopoulos, D.; Akhtar, M.; Nazir, M. Bison remains from the Upper Siwaliks of Pakistan. Neues Jahrb. Geol. Paläontologie Abh. 2010, 258, 121–128. [Google Scholar] [CrossRef]
- Masini, F.; Palombo, M.R.; Rozzi, R. A reappraisal of the Early to Middle Pleistocene Italian Bovidae. Quat. Int. 2013, 288, 45–62. [Google Scholar] [CrossRef]
- Mead, J.; Jin, C.; Wei, G.; Sun, C.; Wang, Y.; Swift, S.; Zheng, L. New data on Leptobos crassus (Artiodactyla, Bovidae) from Renzidong Cave, Early Pleistocene (Nihewanian) of Anhui, China, and an overview of the genus. Quat. Int. 2014, 354, 139–146. [Google Scholar] [CrossRef]
- Burchak-Abramovich, N.I.; Vekua, A.K. On a new pleistocene bovine from Eastern Georgia. In Paleotheriology; Tatarinov, L., Ed.; Nauka: Moscow, Russia, 1994; pp. 253–261. (In Russian) [Google Scholar]
- Sorbelli, L.; Cherin, M.; Kostopoulos, D.; Sardella, R.; Mecozzi, B.; Plotnikov, V.; Prat-Vericat, M.; Azzara, B.; Bartolini-Lucenti, S.; Madurell-Malapeira, J. Earliest bison dispersal in Western Palearctic: Insights from the Eobison record from Pietrafitta (Early Pleistocene, central Italy). Quat. Sci. Rev. 2023, 301, 107923. [Google Scholar] [CrossRef]
- Gromova, V. Hippariony (Rod Hipparion) po Materialam Taraklii, Pavlodara Idrugim; Trudy Paleontologicheskogo Instituta. Publ. House of the Academy of Sciences: Moscow, Russia, 1952; Volume 36, 480p. (In Russian) [Google Scholar]
- Eisenmann, V. What metapodial morphometry has to say about some Miocene hipparions. In Paleoclimate and Evolution, with Emphasis on Human Origins; Vrba, E.S., Denton, G.H., Partridge, T.C., Burckle, L.H., Eds.; Yale University Press: New Haven, CT, USA, 1995; pp. 148–163. [Google Scholar]
- Scott, R. The Comparative Paleoecology of Late Miocene Eurasian Hominoids. Ph.D. Thesis, The University of Texas at Austin, Austin, TX, USA, 2004; p. 458. [Google Scholar]
- Gambaryan, P. The Running of Mammals; Publishing House Nauka: Leningrad, Russia, 1972; p. 334. [Google Scholar]
- Gentry, A. An Ovibovine (Mammalia, Bovidae) from the Neogene of Stratzing, Austria. Ann. Naturhistorische MuseumWien 2001, 102, 189–199. [Google Scholar]
- Cherin, M.; Sorbelli, L.; Crotti, M.; Iurino, D.; Sardella, R.; Souron, A. New material of Sus strozzii (Suidae, Mammalia) from the Early Pleistocene of Italy and a phylogenetic analysis of suines. Quat. Sci. Rev. 2018, 194, 94–115. [Google Scholar] [CrossRef]
- Brugal, J.-P.; Boudadi-Maligne, M. Quaternary small to large canids in Europe: Taxonomic status and biochronological contribution. Quat. Int. 2011, 243, 171–182. [Google Scholar] [CrossRef]
- Dermitzakis, M. The evolution of the Aegeis during the Late Cenozoic. Geol. Balc. 1990, 20, 3–16. [Google Scholar] [CrossRef]
- Spassov, N. Villafranchian Succession of Mammalian Megafaunas from Bulgaria and the Biozonation of South-East Europe. In Actes du Congres Biochrom ’97; Aguilar, J.-P., Legendre, S., Michaux, J., Eds.; Mémoires et travaux de l’Institut de Montpellier de l’École Pratique des Hautes Études; Institut de Montpellier: Montpellier, France, 1997; Volume 21, pp. 669–676. [Google Scholar]
- Strait, D.; Orr, C.; Hodgkins, J.; Spassov, N.; Gurova, M.; Miller, C.; Tzankov, T. The Human Fossil Record of Bulgaria and the Formulation of Biogeographic Hypotheses. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Vertebrate Paleobiology and Paleoanthropology Series; Springer Science+Business Media: Dordrecht, The Netherlands, 2016; pp. 187–211. [Google Scholar]
- Shchelinsky, V.; Gurova, M.; Tesakov, A.; Titov, V.; Frolov, P.; Simakova, A. The early Pleistocene site of Kermek in western Ciscaucasia (Southern Russia): Stratigraphy, biotic record and lithic industry (preliminary results). Quat. Int. 2016, 393, 51–69. [Google Scholar] [CrossRef]
- Vangengeim, E. Paleontological Justification of the Quaternaqr Stratigraphy of North Asia; Nauka: Moscow, Russia, 1977; p. 171. (In Russian) [Google Scholar]
- Tamvakis, A.; Savvidou, A.; Spassov, N.; Youlatos, D.; Merceron, G.; Kostopoulos, D. New insights on Early Pleistocene Nyctereutes from the Balkans based on material from Dafnero-3 (Greece) and Varshets (Bulgaria). Palaeoworld 2023, 32, 555–572. [Google Scholar] [CrossRef]
- Marciszak, A.; Lipecki, G.; Spassov, N. Martes wenzensis Stach, 1959 within the early history of the genus Martes Pinel, 1792. Acta Geol. Pol. 2024, 74, e4. [Google Scholar] [CrossRef]
- Koufos, G.; Kostopoulos, D. New Carnivore Material from the Plio-Pleistocene of Macedonia (Greece) with the description of a new Canid. Münch. Geowiss. Abh. 1997, 34, 33–63. [Google Scholar]
- Benammi, M.; Aidona, E.; Merceron, G.; Koufos, G.; Kostopoulos, D. Magnetostratigraphy and Chronology of the Lower Pleistocene Primate Bearing Dafnero Fossil Site, N. Greece. Quaternary 2020, 3, 22. [Google Scholar] [CrossRef]
- Konidaris, G.; Kostopoulos, D. The Early Pleistocene large mammal succession of Greece: Implications for pan-European biogeographic correlations. In Proceedings of the XXI INQUA Congress, Rome, Italy, 13–20 July 2023; Session 135. Mammals, Biochronology and Palaeoecology of the Euro-Mediterrane an Quaternary; Abstracts of Papers. Sapienza University of Rome: Rome, Italy, 2023. Available online: https://inquaroma2023.exordo.com/programme/presentations (accessed on 2 January 2024).
- Konidaris, G.; Amanatidou, M.; Chitoglou, K.; Fragkioudakis, E.; Gerakakis, N.; Giannakou, V.; Gkeme, A.; Kalaitzi, C.; Ladas, A.; Tsakalidis, C.; et al. Krimni-3: A Νew Lower Pleistocene Vertebrate Site in Mygdonia Basin. In Proceedings of the 16th International Congress of the Geological Society of Greece, Patras, Greece, 17–19 October 2022. Bulletin of the Geological Society of Greece, Sp. Publ. 10, Ext. Abs. GSG 2022, 138. [Google Scholar]
- Pandolfi, L.; Erten, H. Stephanorhinus hundsheimensis (Mammalia, Rhinocerotidae) from the late early Pleistocene deposits of the Denizli Basin (Anatolia, Turkey). Geobios 2017, 50, 65–73. [Google Scholar] [CrossRef]
- Vislobokova, I.; Agadzhanyan, A.; Lopatin, A. The case of Trlica TRL11–10 (Montenegro): Implications for possible early hominin dispersals into the Balkans in the middle of the Early Pleistocene. Quat. Int. 2020, 554, 15–35. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spassov, N. The Final Pliocene and Early Pleistocene Faunal Dispersals from East to Europe and Correlation of the Villafranchian Biochronology between Eastern and Western Europe. Quaternary 2024, 7, 43. https://doi.org/10.3390/quat7040043
Spassov N. The Final Pliocene and Early Pleistocene Faunal Dispersals from East to Europe and Correlation of the Villafranchian Biochronology between Eastern and Western Europe. Quaternary. 2024; 7(4):43. https://doi.org/10.3390/quat7040043
Chicago/Turabian StyleSpassov, Nikolai. 2024. "The Final Pliocene and Early Pleistocene Faunal Dispersals from East to Europe and Correlation of the Villafranchian Biochronology between Eastern and Western Europe" Quaternary 7, no. 4: 43. https://doi.org/10.3390/quat7040043
APA StyleSpassov, N. (2024). The Final Pliocene and Early Pleistocene Faunal Dispersals from East to Europe and Correlation of the Villafranchian Biochronology between Eastern and Western Europe. Quaternary, 7(4), 43. https://doi.org/10.3390/quat7040043