
Early to Mid-Holocene Tree Immigration and Spread in the Isle of Man: The Roles of Climate and Other Factors

 ,
,
Abstract
1. Introduction
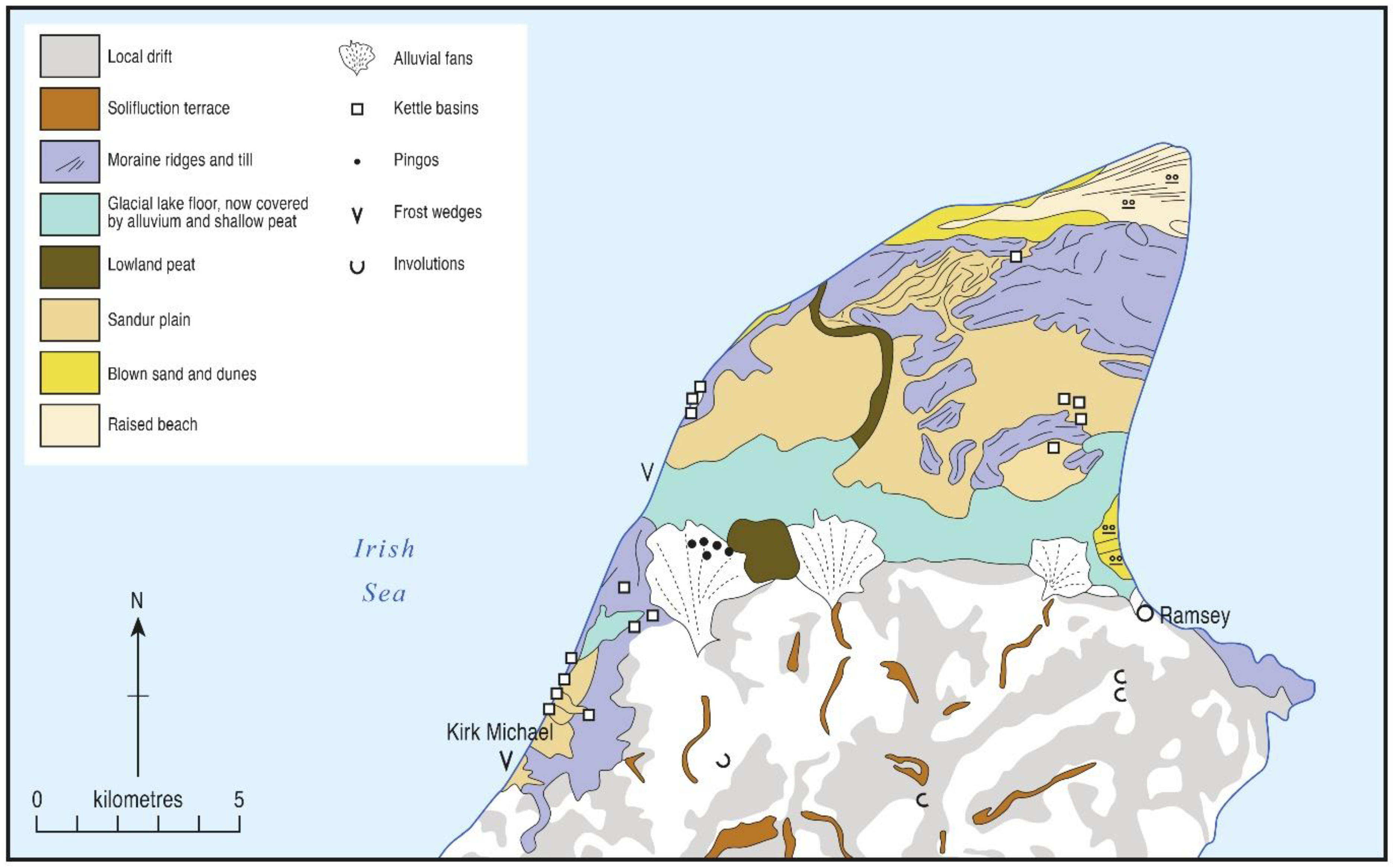
2. Environmental Background

3. Materials and Methods
3.1. Methodology
3.2. Radiocarbon Dating
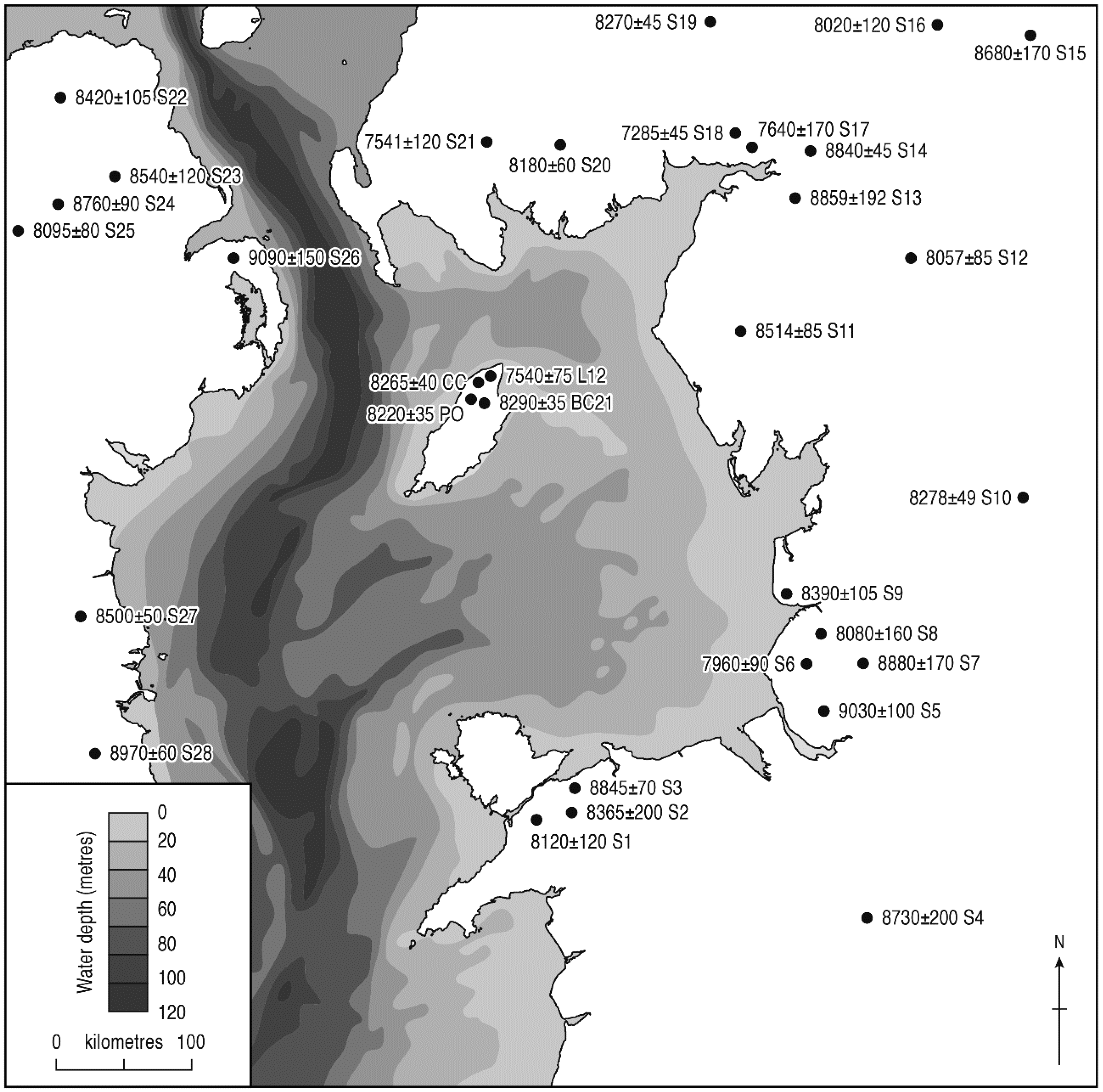
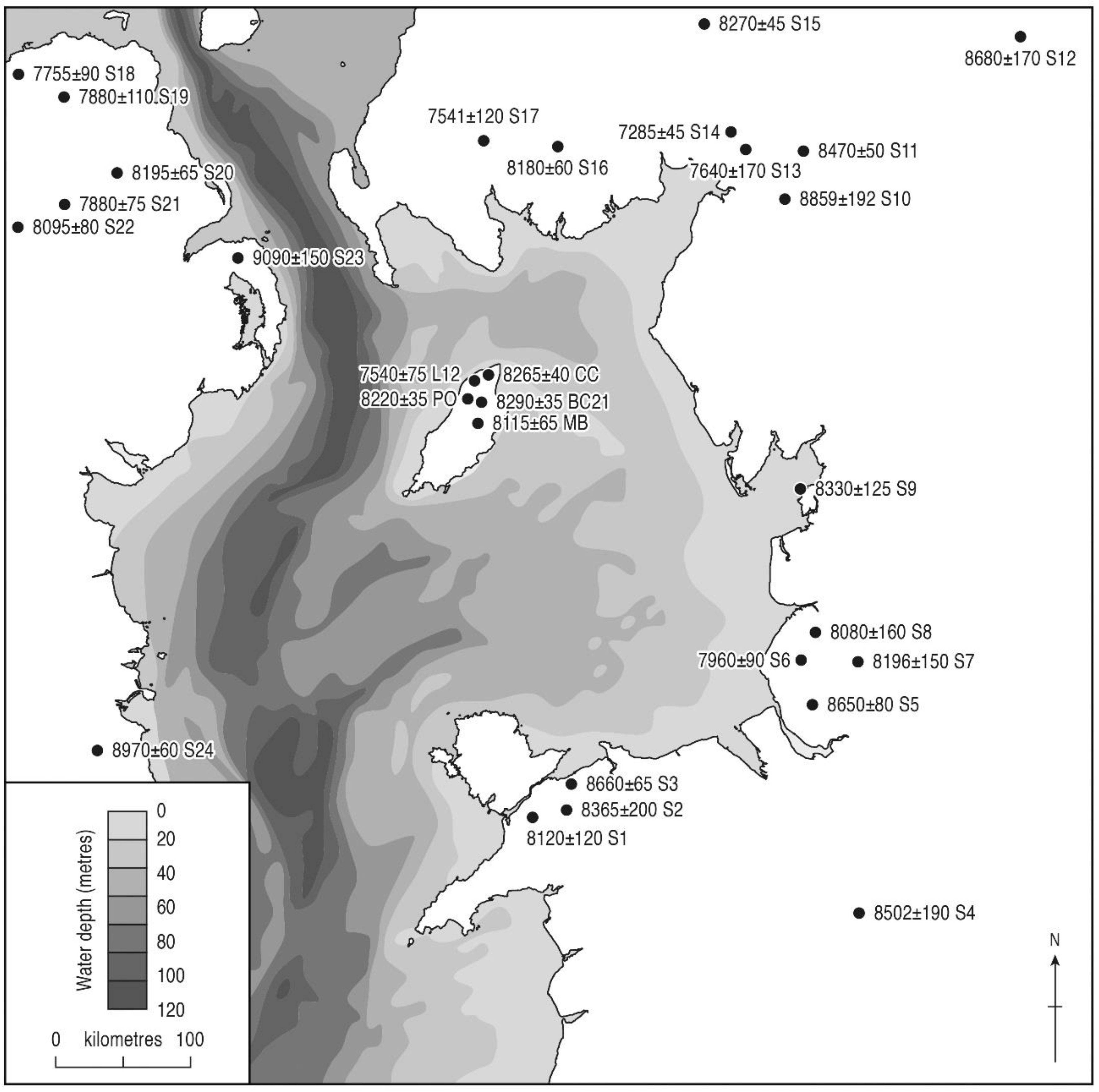
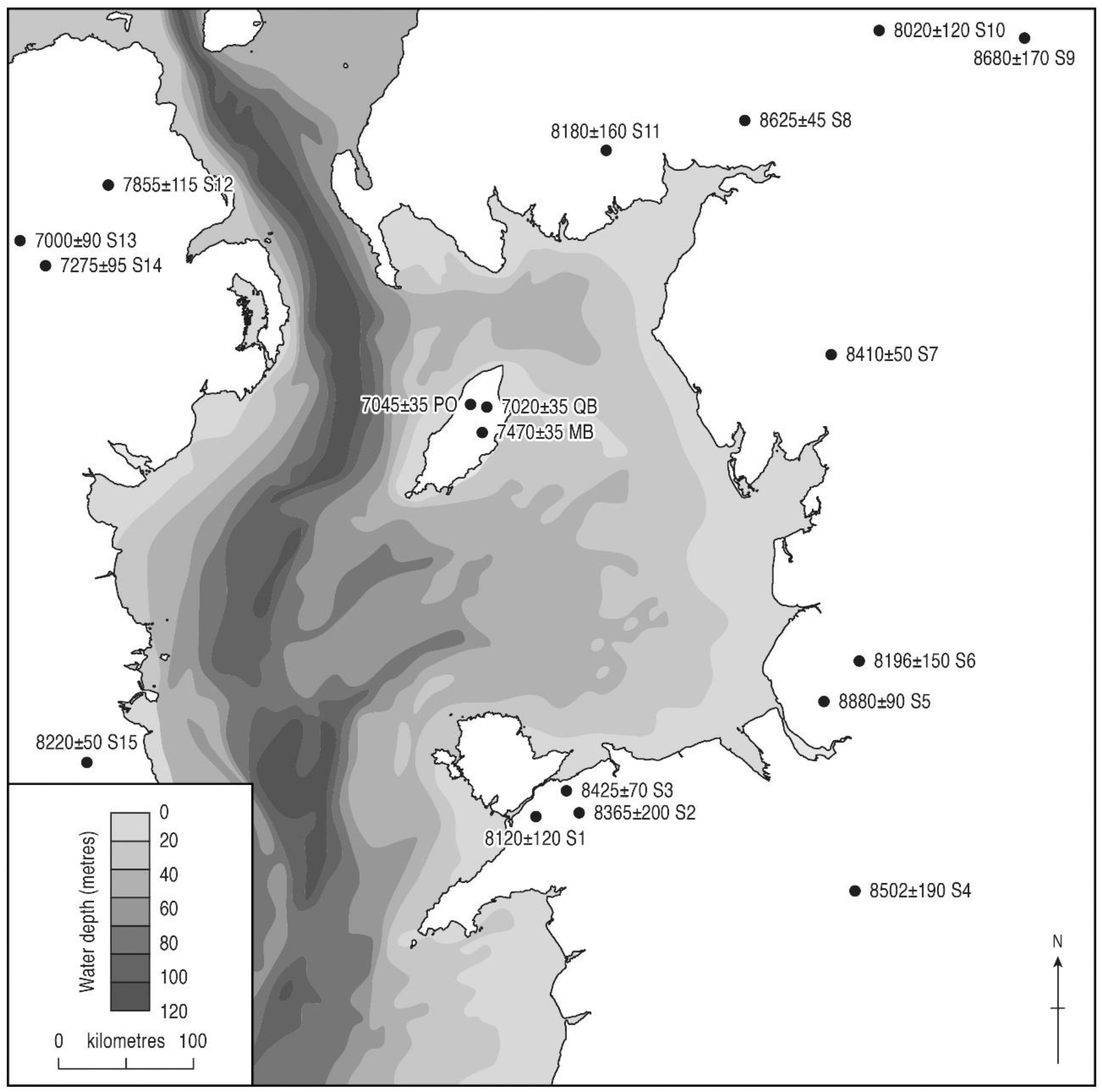
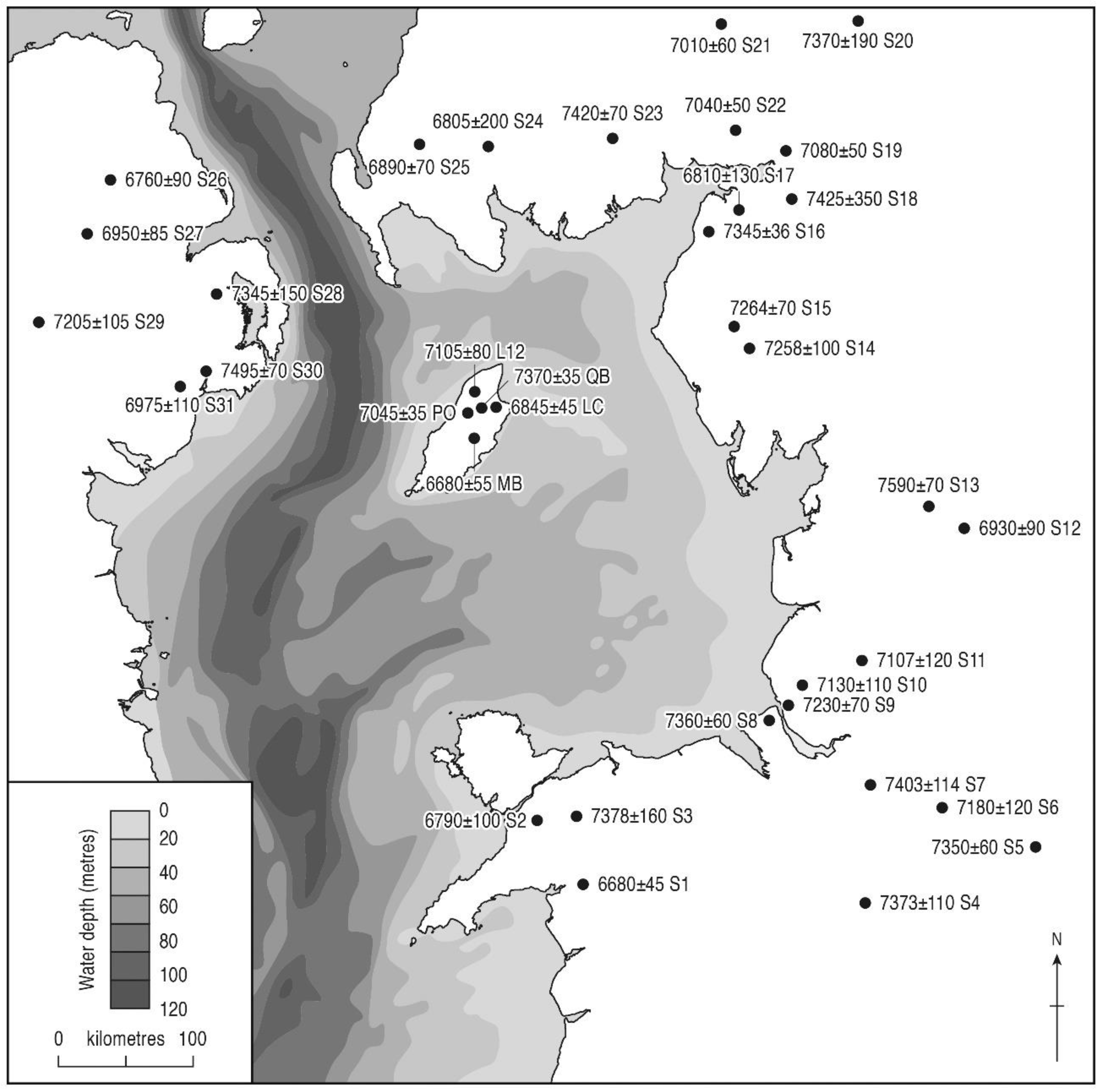
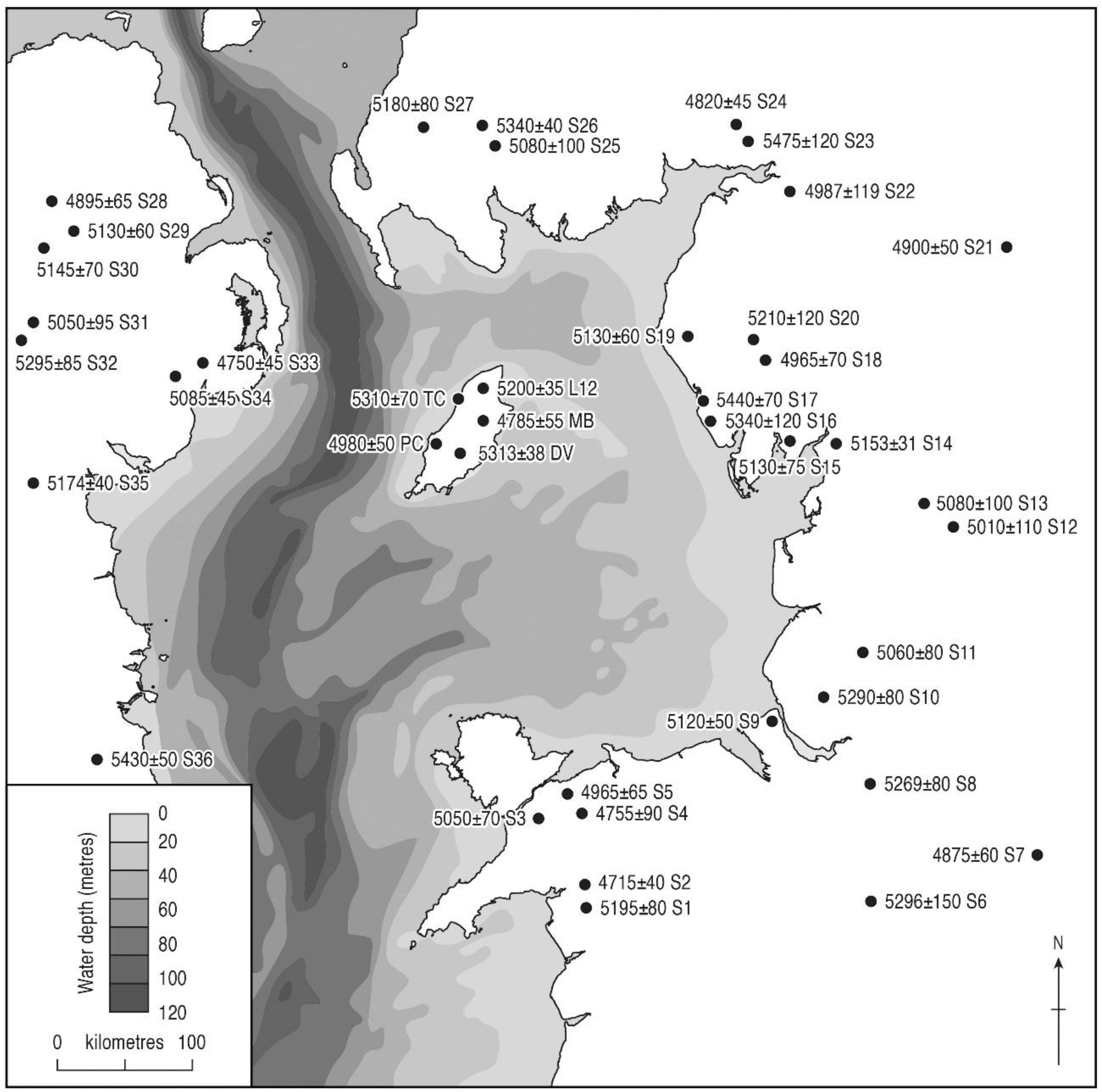
3.3. Tree Immigration Maps
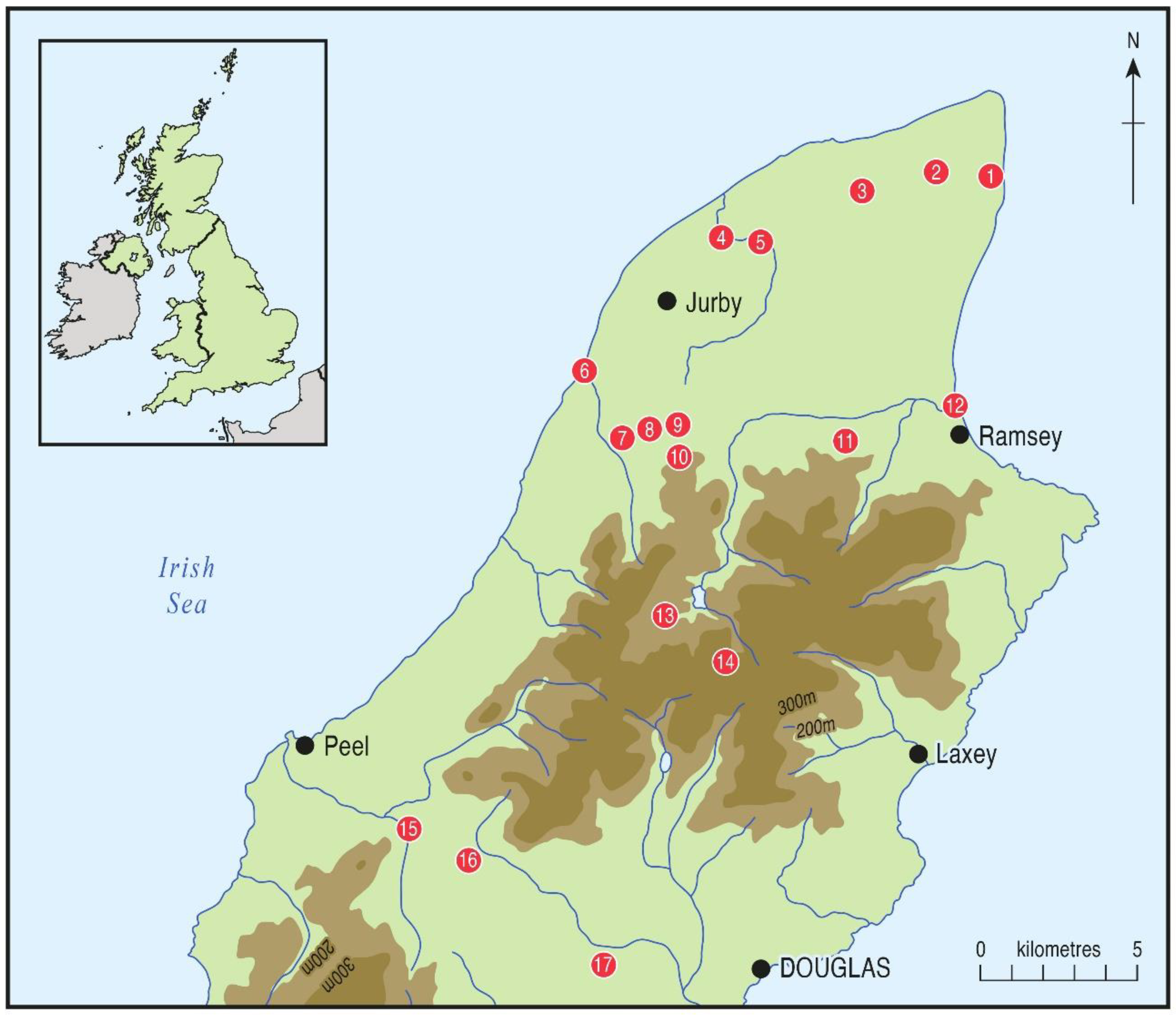
4. Palaeoecological Sites and Data
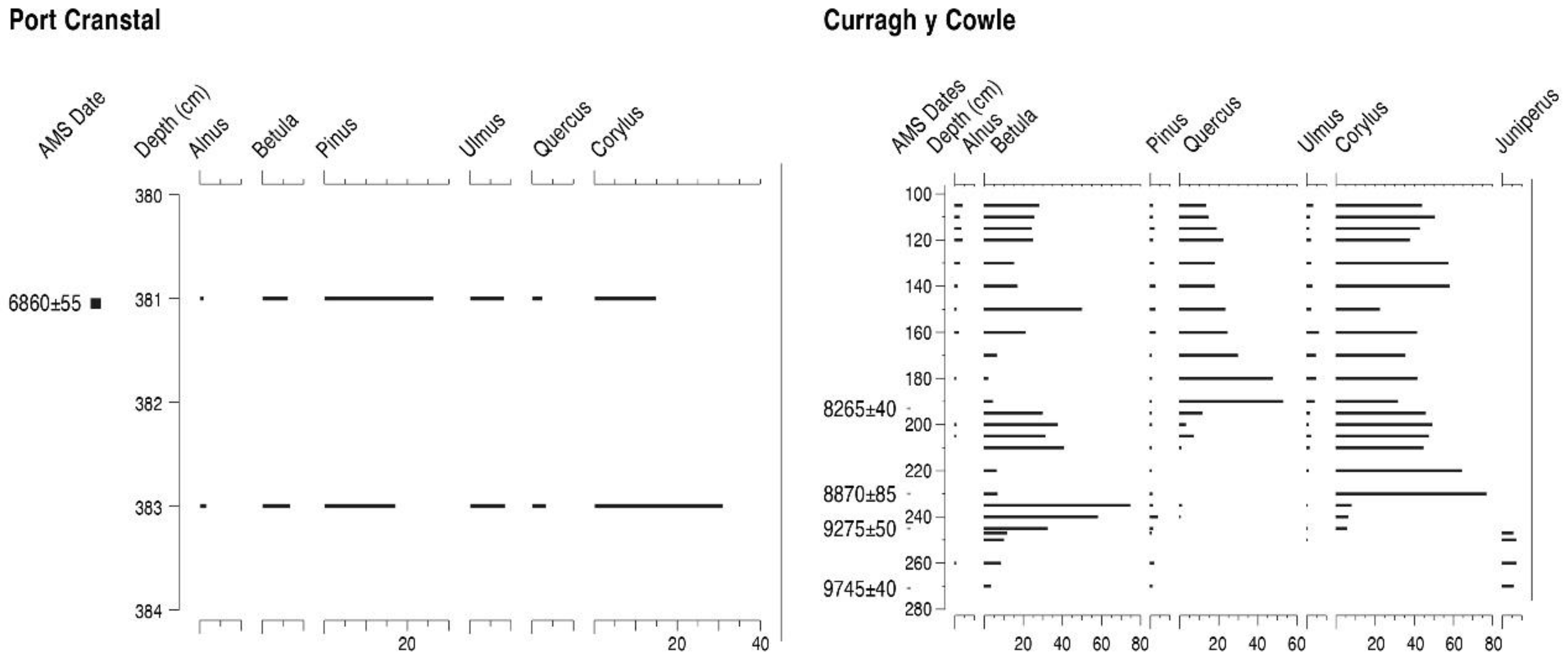
4.1. Cranstal (C)
4.2. Curragh y Cowle (CC)
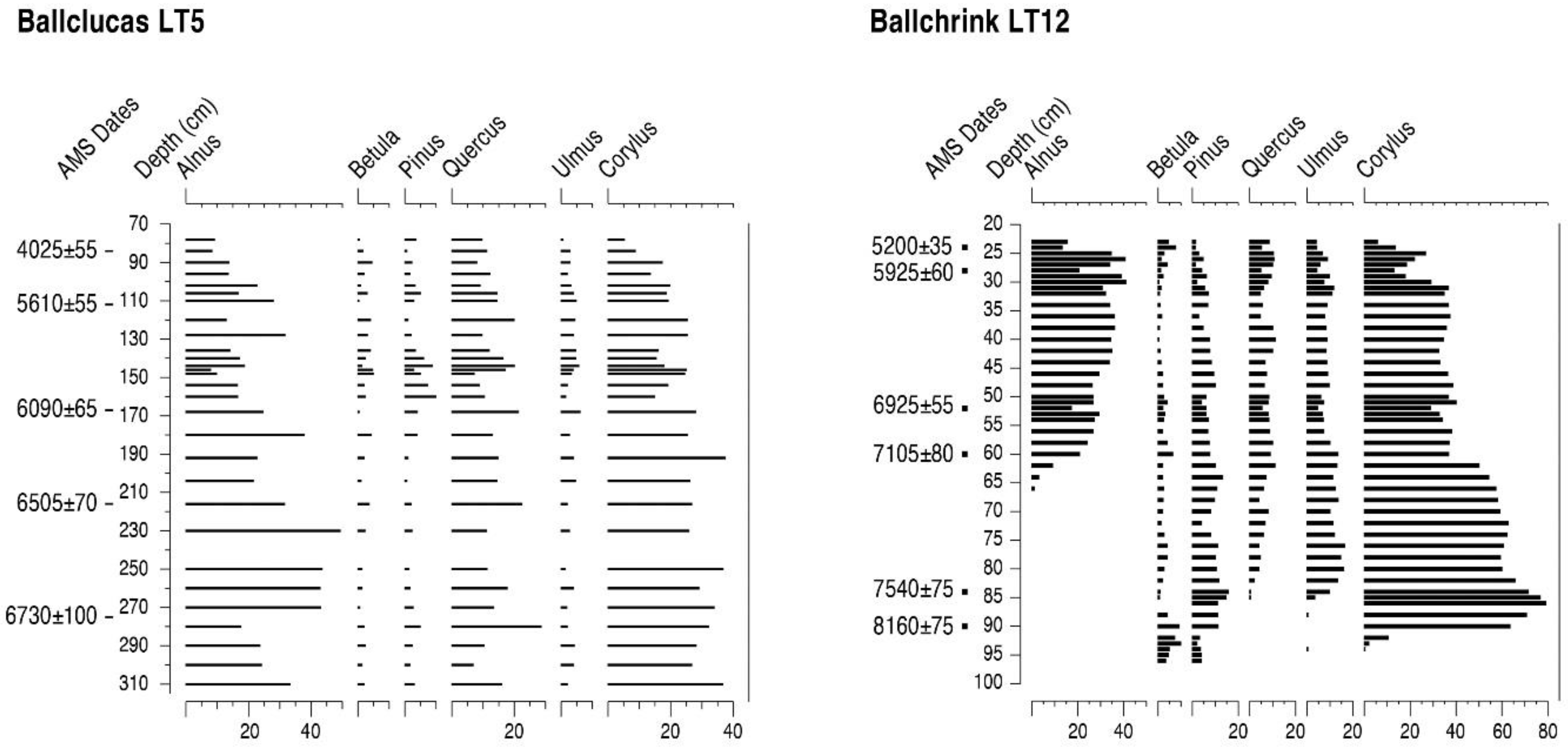
4.3. The Lhen Trench (LT5 and LT12)
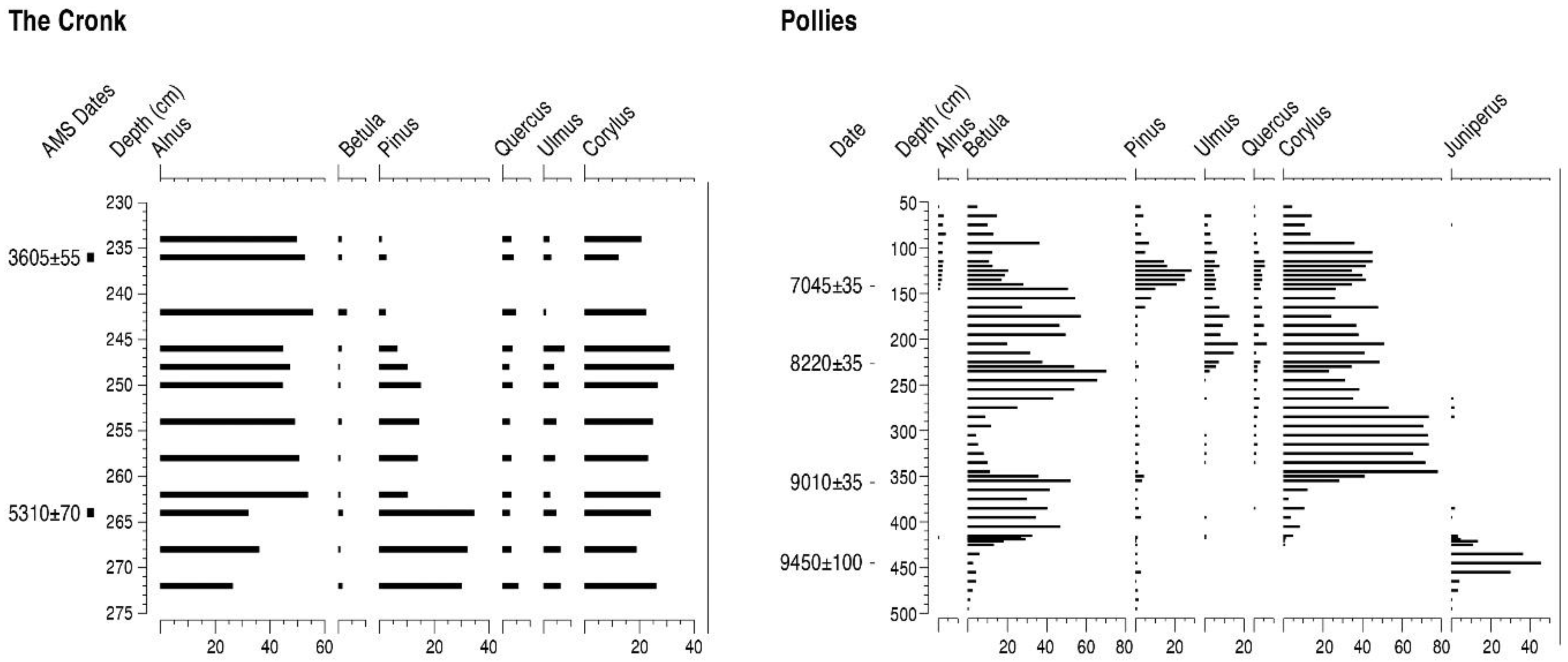
4.4. The Cronk (TC)
4.5. Pollies (PO)
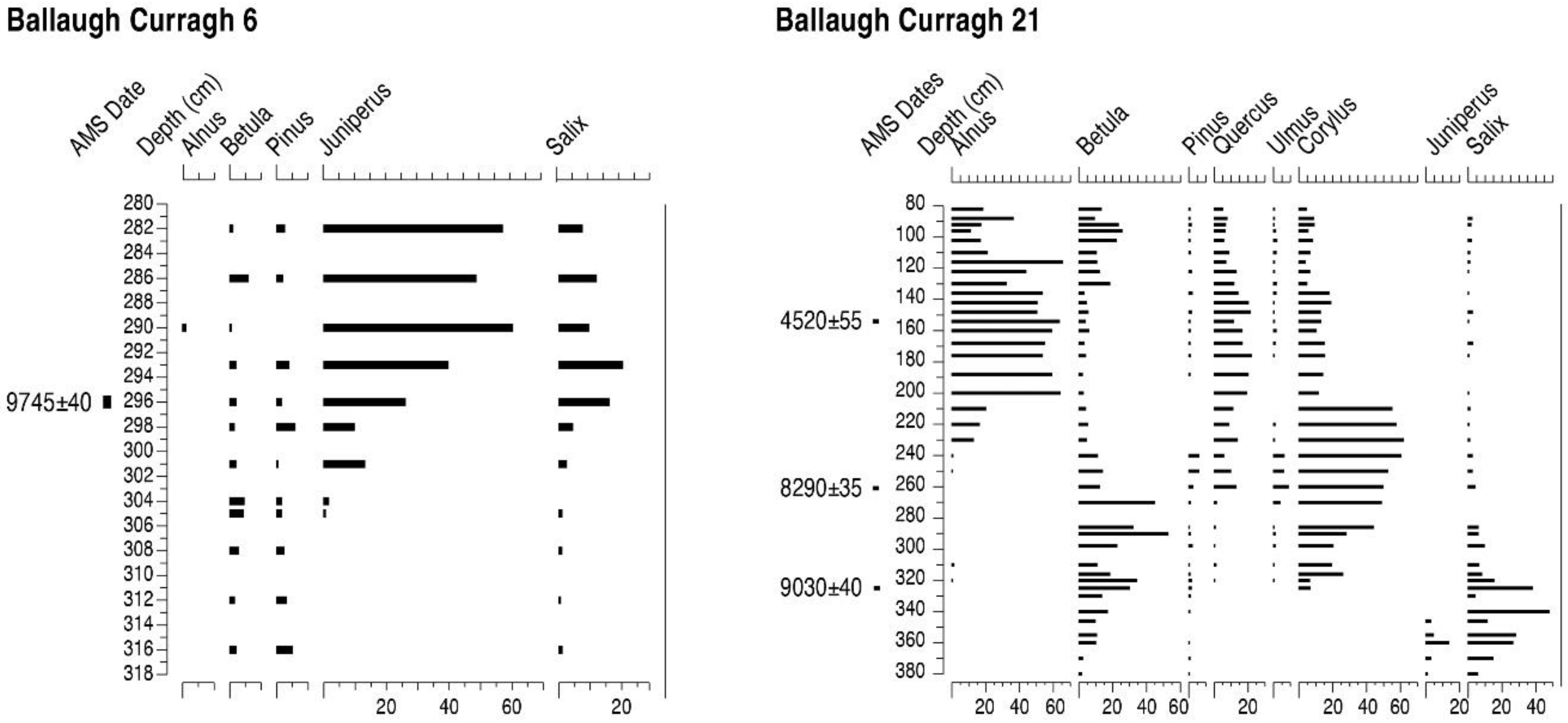
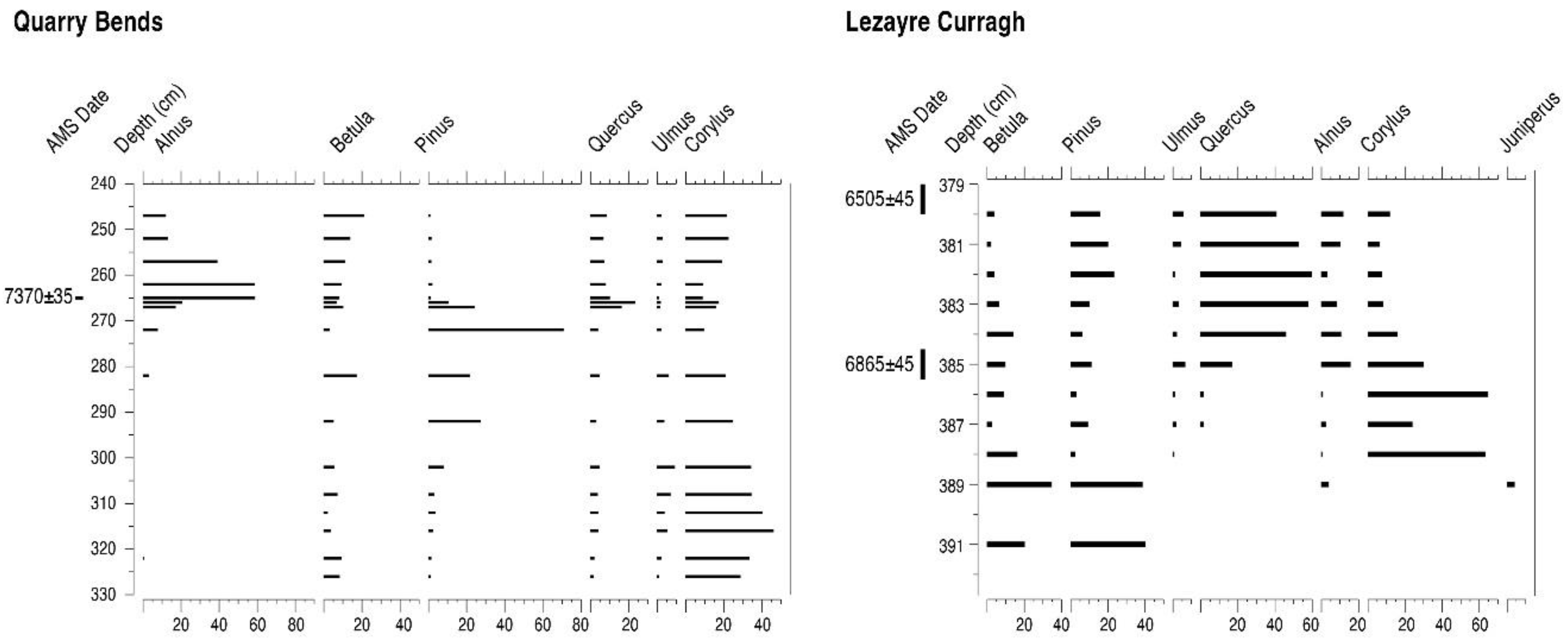
4.6. Ballaugh Curragh and Quarry Bends (BC6, BC21 and QB)
4.7. Lezayre Curragh (LC)
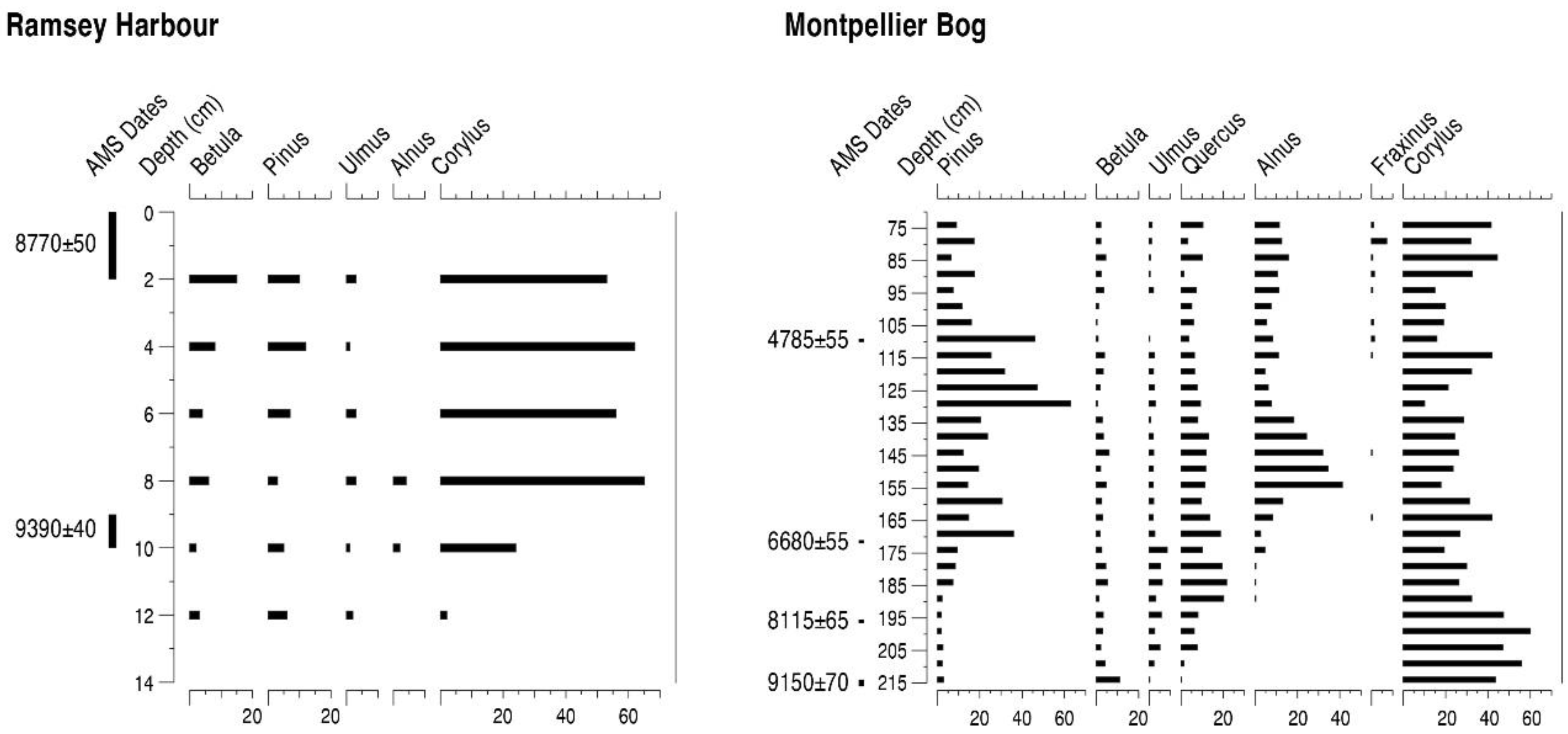
4.8. Ramsey Harbour (RH)
4.9. Montpellier Bog (MB)
4.10. Beinn y Phott
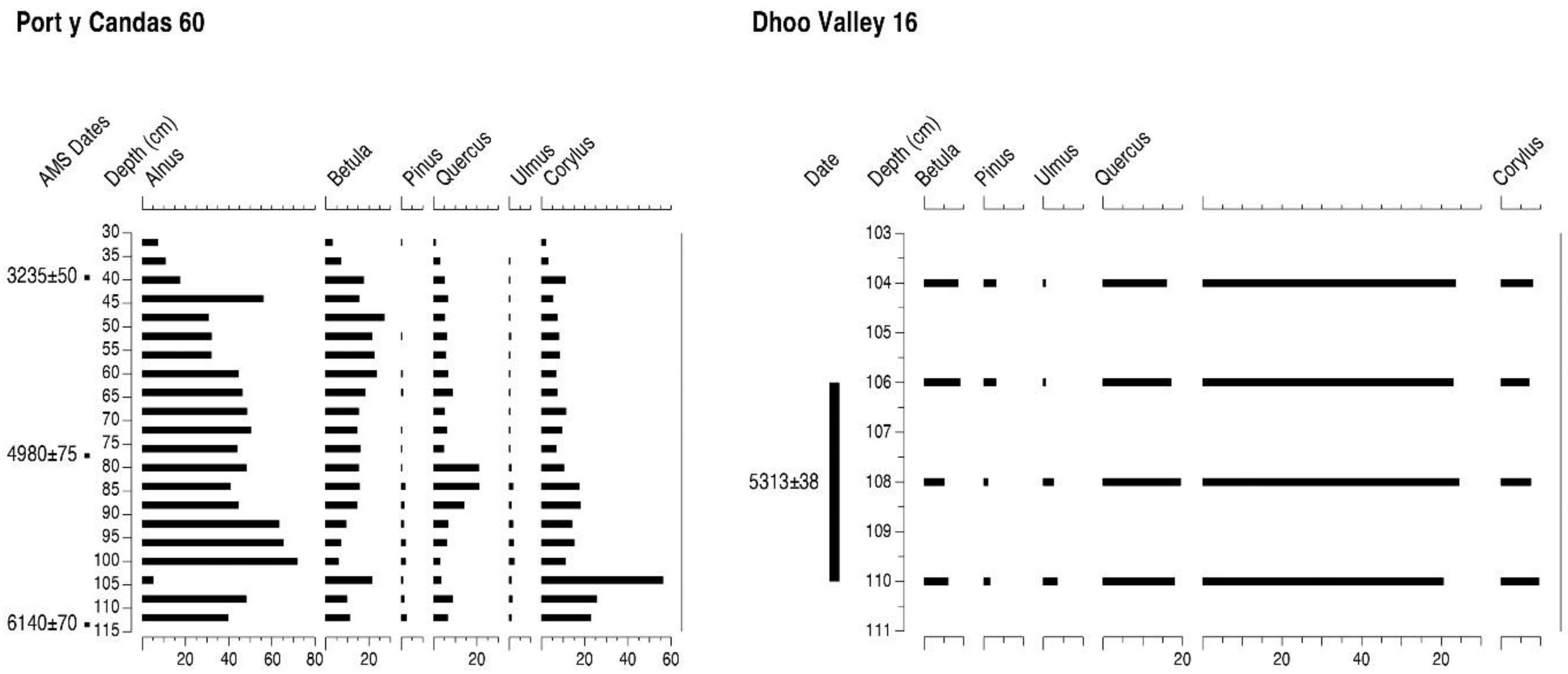
4.11. Port-y-Candas (PC)
4.12. Greeba Curragh
4.13. Dhoo Valley (DV)
5. Discussion
5.1. Early to Mid-Holocene Tree Spreading in the Isle of Man
5.1.1. Pre-Woodland Succession and the Juniperus Maximum
5.1.2. The Betula Rise
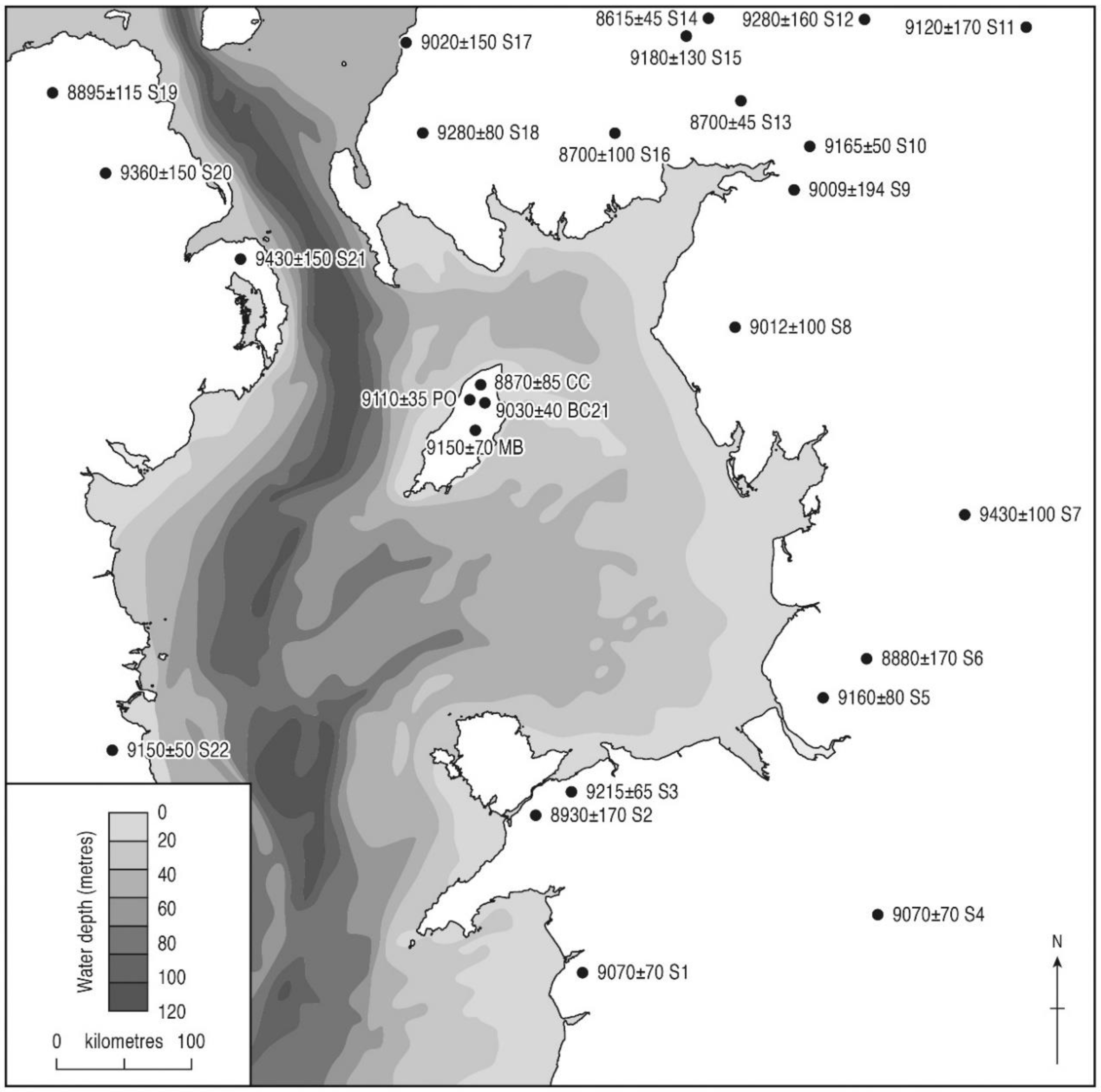
5.1.3. The Corylus Rise
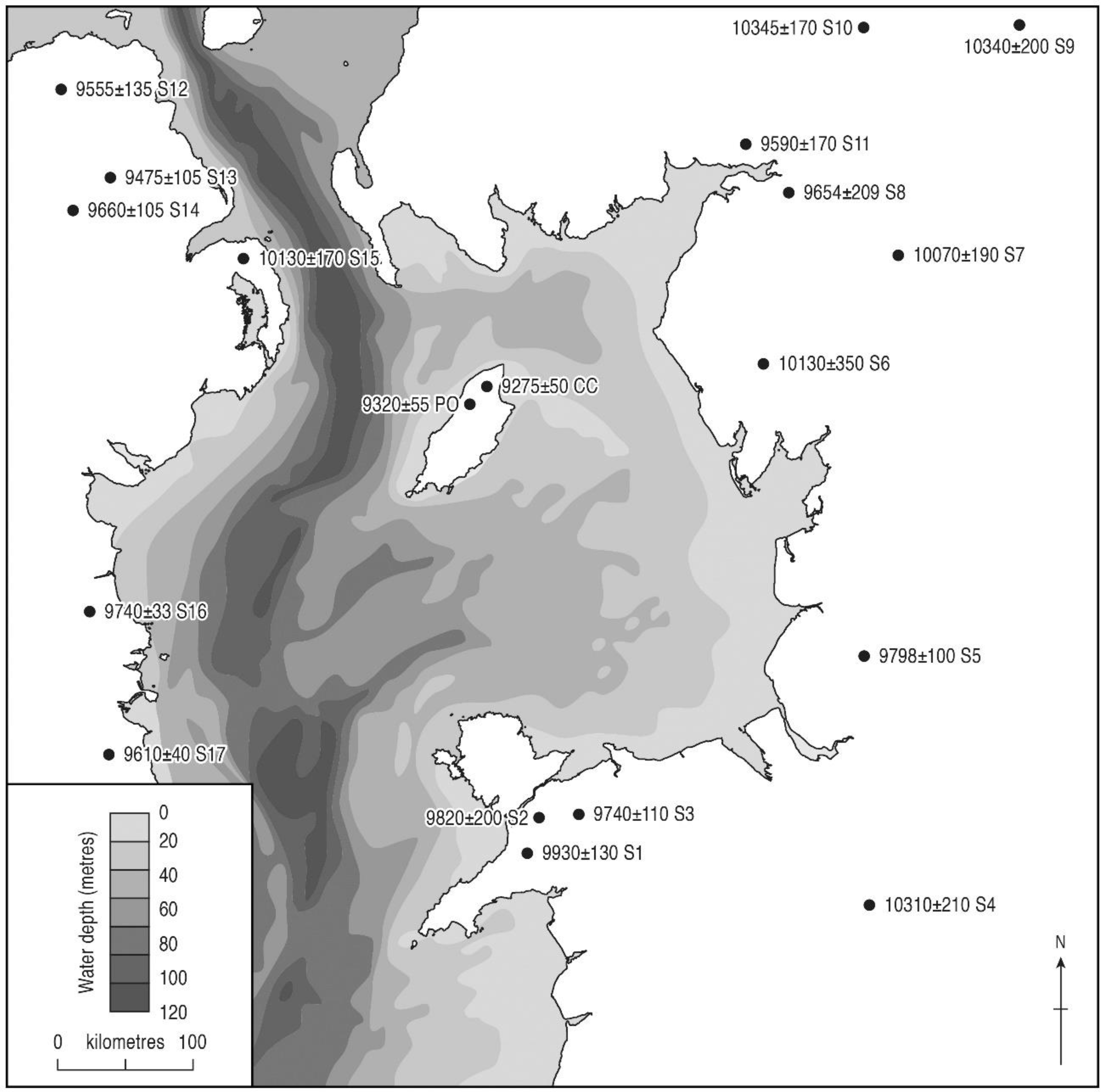
5.1.4. The Ulmus and Quercus Rises
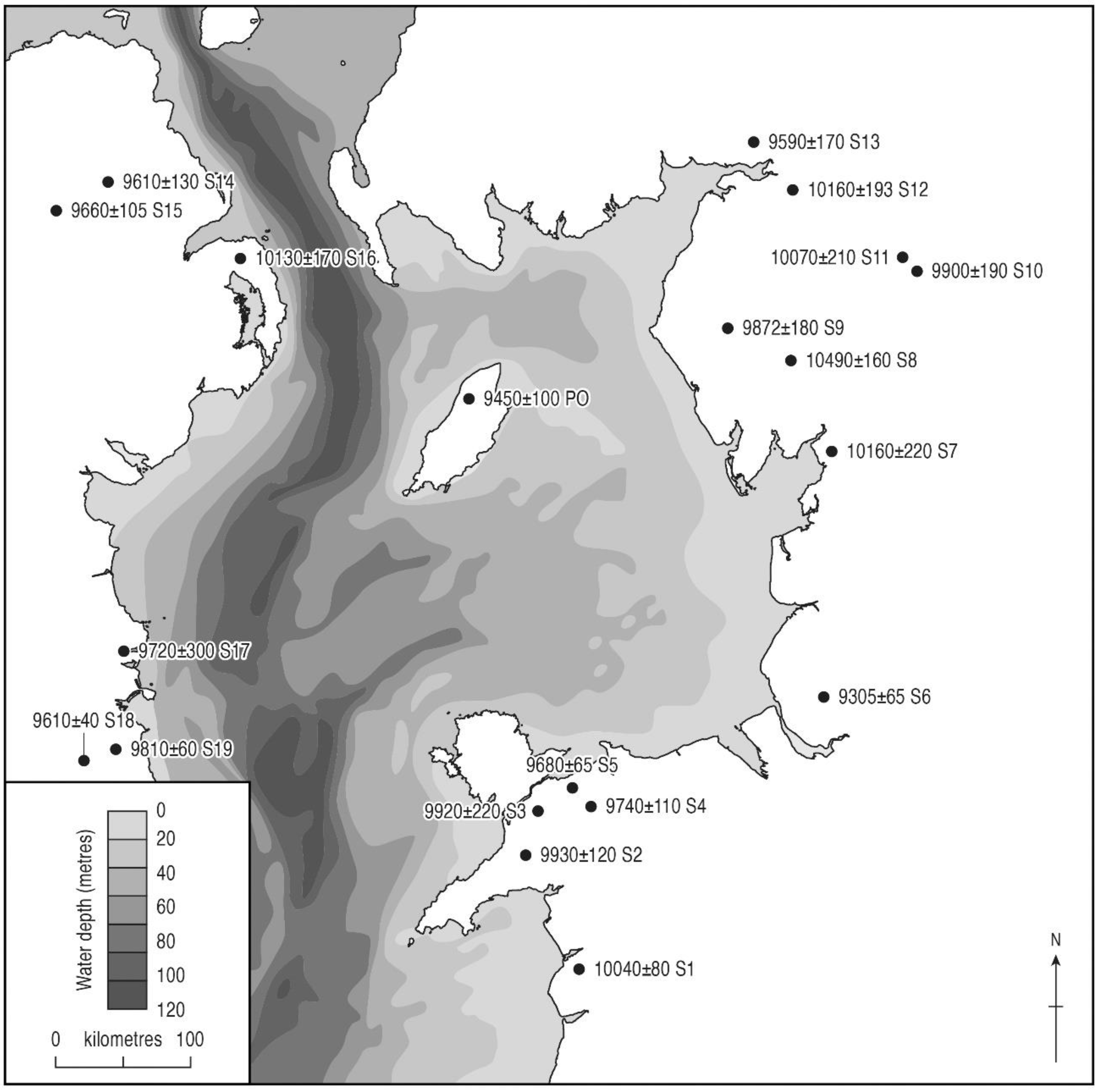
5.1.5. The Pinus Rise
5.1.6. The Alnus Rise
5.1.7. Later Tree Arrivals and the Ulmus Decline
5.2. Insularity
5.3. Climate
5.4. Geology and Soils
5.5. Competition and Tree Spreading
5.6. Disturbance and Human Activity
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huntley, B.; Birks, H.J.B. An Atlas of Past and Present Pollen Maps for Europe 0–13,000 Years Ago; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Birks, H.J.B. Holocene isochrone maps and patterns of tree-spreading in the British Isles. J. Biogeogr. 1989, 16, 503–540. [Google Scholar] [CrossRef]
- Smith, A.G.; Pilcher, J.R. Radiocarbon dates and vegetational history of the British Isles. New Phytol. 1973, 72, 903–914. [Google Scholar] [CrossRef]
- Lang, G. Quartäre Vegetationsgeschichte Europas. Methoden und Ergebnisse; Gustav Fischer Verlag: Jena, Germany, 1994. [Google Scholar]
- Brubaker, L.B. Responses of tree populations to climatic change. Vegetatio 1986, 67, 119–130. [Google Scholar] [CrossRef]
- Ritchie, J.C. Climate change and vegetation response. Vegetatio 1986, 67, 65–74. [Google Scholar] [CrossRef]
- Moore, P.D. Tree boundaries on the move. Nature 1987, 326, 545. [Google Scholar] [CrossRef]
- Huntley, B. European vegetation history: Palaeovegetation maps from pollen data—13,000 yr BP to present. J. Quat. Sci. 1990, 5, 103–122. [Google Scholar] [CrossRef]
- Huntley, B. Rates of change in the European palynological record of the last 13,000 years and their climatic interpretation. Clim. Dyn. 1992, 6, 185–191. [Google Scholar] [CrossRef]
- Huntley, B. The post-glacial history of British woodlands. In Woodland in the Landscape: Past and Future Perspectives; Atherden, M., Butlin, R.A., Eds.; PLACE Research Centre: York, UK, 1998; pp. 9–25. [Google Scholar]
- Mitchell, F.J.G. Natural invaders: The Postglacial tree colonisation of Ireland. In Biology Invaders: The Impact of Exotic Species; Moriarty, C., Murray, D.A., Eds.; Royal Irish Academy: Dublin, Republic of Ireland, 2002; pp. 2–12. [Google Scholar]
- Deacon, J. The location of refugia of Corylus avellana L. during the Weichselian glaciation. New Phytol. 1974, 73, 1055–1073. [Google Scholar] [CrossRef]
- Bennett, K.D.; Tzedakis, P.C.; Willis, K.J. Quaternary refugia of north European trees. J. Biogeogr. 1991, 18, 103–115. [Google Scholar] [CrossRef]
- Tzedakis, P.C.; Emerson, B.C.; Hewitt, G.M. Cryptic or mystic? Glacial tree refugia in northern Europe. TREE 2013, 28, 696–704. [Google Scholar] [CrossRef]
- Kelly, A.; Charman, D.J.; Newnham, R.M. A last glacial maximum pollen record from Bodmin Moor showing a possible cryptic northern refugium in southwest England. J. Quat. Sci. 2010, 25, 296–308. [Google Scholar] [CrossRef]
- Young, D.S.; Green, C.P.; Batchelor, C.R.; Austin, P.; Elias, S.A.; Athersuch, J.; Lincoln, P. Macrofossil evidence of alder (Alnus sp.) in Britain early in the Late Glacial Interstadial: Implications for the northern cryptic refugia debate. J. Quat. Sci. 2021, 36, 40–55. [Google Scholar] [CrossRef]
- Mitchell, F.J.G. Where did Ireland’s trees come from? Biol. Env. Proc. R. Irish Acad. 2006, 106B, 251–259. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Deacon, J.; Peglar, S.M. Pollen maps for the British Isles 5000 years ago. Proc. Roy. Soc. B 1975, 189, 87–105. [Google Scholar]
- Bennett, K.D. A provisional map of forest types for the British Isles 5000 years ago. J. Quat. Sci. 1989, 4, 141–144. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Saarnisto, M. Isopollen maps and principal components analysis of Finnish pollen data for 4000, 6000 and 8000 years ago. Boreas 1975, 5, 77–96. [Google Scholar] [CrossRef]
- Jacobson, G.L., Jr.; Webb, T., III; Grimm, E.C. Patterns and rates of vegetation change during the deglaciation of eastern North America. In North America and Adjacent Oceans during the Last Deglaciation; Ruddiman, W.F., Wright, H.E., Jr., Eds.; Geological Society of America, The Geology of North America, v. K-3: Boulder, CO, USA, 1987; pp. 277–288. [Google Scholar]
- Clark, J.S. Why trees migrate so fast: Confronting theory with dispersal biology and the paleorecord. Am. Nat. 1998, 152, 204–224. [Google Scholar] [CrossRef]
- Gieseke, T.; Brewer, S.; Finsinger, W.; Leydet, M.; Bradshaw, R.H.W. Patterns and dynamics of European vegetation change over the last 15,000 years. J. Biogeogr. 2017, 44, 1441–1456. [Google Scholar] [CrossRef]
- Gieseke, T.; Brewer, S. Notes on the postglacial spread of abundant European tree taxa. Veget. Hist. Archaeobot. 2018, 27, 337–349. [Google Scholar] [CrossRef]
- Bennett, K.D. The rate of spread and population increase of forest trees during the postglacial. Philos. Trans. R. Soc. Lon. B 1986, 314, 523–531. [Google Scholar]
- Huntley, B. Glacial and Holocene vegetation history: Europe. In Vegetation History; Huntley, B., Webb, T., III, Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 341–383. [Google Scholar]
- Prentice, I.C. Vegetation responses to past climatic variation. Vegetatio 1986, 67, 131–141. [Google Scholar] [CrossRef]
- Giesecke, T. Changing plant distributions. In Encyclopaedia of Quaternary Science; Elias, S.C., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 2544–2551. [Google Scholar]
- McCarroll, D.; Garrad, L.; Dackombe, R. Lateglacial and postglacial environmental history. In The Isle of Man: Celebrating a Sense of Place; Robinson, V., McCarroll, D., Eds.; Liverpool University Press: Liverpool, UK, 1990; pp. 55–76. [Google Scholar]
- Chiverrell, R.C. Changing Landscapes of the Isle of Man. In Mannin Revisited: Twelve Essays on Manx Culture and Environment; Davey, P., Finlayson, D., Eds.; Scottish Society for Northern Studies: Edinburgh, UK, 2002; pp. 1–9. [Google Scholar]
- Chiverrell, R.C.; Thomas, G.S.P. A New History of the Isle of Man. Vol. I. The Evolution of the Natural Landscape; Liverpool University Press: Liverpool, UK, 2006. [Google Scholar]
- Godwin, H. The History of the British Flora; Cambridge University Press: Cambridge, UK, 1975. [Google Scholar]
- Birks, H.J.B.; Deacon, J. A numerical analysis of the past and present flora of the British Isles. New Phytol. 1973, 72, 877–902. [Google Scholar] [CrossRef]
- Allen, D.E. The Flora of the Isle of Man; The Manx Museum and National Trust: Douglas, Isle of Man, 1984. [Google Scholar]
- Tomlinson, P.R.; Charter, E.; Garrad, L. Biodiversity: Vascular plants. In A New History of the Isle of Man. Vol. I. The Evolution of the Natural Landscape; Chiverrell, R.C., Thomas, G.S.P., Eds.; Liverpool University Press: Liverpool, UK, 2006; pp. 406–408. [Google Scholar]
- Allen, D.E. The present-day fauna and flora of the Isle of Man as indicators of the date of Flandrian severance. In Man and Environment in the Isle of Man; Davey, P.J., Ed.; British Archaeological Reports, British Series 54; BAR Publishing: Oxford, UK, 1978; pp. 9–14. [Google Scholar]
- Tomlinson, P.R.; Pooley, E. History of the native vertebrate fauna. In A New History of the Isle of Man. Vol. I. The Evolution of the Natural Landscape; Chiverrell, R.C., Thomas, G.S.P., Eds.; Liverpool University Press: Liverpool, UK, 2006; pp. 297–300. [Google Scholar]
- Dackombe, R.; McCarroll, D. The Manx Landscape. In The Isle of Man: Celebrating a Sense of Place; Robinson, V., McCarroll, D., Eds.; Liverpool University Press: Liverpool, UK, 1990; pp. 10–17. [Google Scholar]
- Chiverrell, R.C.; Thomas, G.S.P.; Long, D.; Roberts, D.H. Quaternary. In Geology of the Isle of Man and Its Offshore Area; Chadwick, R.A., Jackson, D.I., Barnes, Kimbell, G.S., Johnson, H., Chiverrell, R.C., Thomas, G.S.P., Jones, N.S., Riley, N.J., Pickett, E.A., et al., Eds.; British Geological Survey Research Report RR01/06; British Geological Survey: Nottingham, UK, 2001; pp. 107–116. [Google Scholar]
- Fullen, M.; Harris, J.; Hallett, M. Soils of the Isle of Man. In A New History of the Isle of Man. Vol. I. The Evolution of the Natural Landscape; Chiverrell, R.C., Thomas, G.S.P., Eds.; Liverpool University Press: Liverpool, UK, 2006; pp. 340–350. [Google Scholar]
- Russell, G. The structure and vegetation history of the Manx hill peats. In Man and Environment in the Isle of Man; Davey, P., Ed.; British Archaeological Reports 54; BAR Publishing: Oxford, UK, 1978; pp. 39–50. [Google Scholar]
- Fullen, M.; Harris, J.; Kear, B.S. Soils of the Isle of Man; Centre for Manx Studies Research Report 5; Centre for Manx Studies: Douglas, Isle of Man, 1996. [Google Scholar]
- Tomlinson, P.R. The Manx hill-land: The palaeoenvironmental resource. In Proceedings of the Manx Hill-Land Seminar; Manx Nature Conservation Trust: Douglas, Isle of Man, 1997; pp. 57–62. [Google Scholar]
- Roberts, D.H.; Dackombe, R.V.; Thomas, G.S.P. Palaeo-ice streaming in the central sector of the British–Irish ice sheet during the last glacial maximum: Evidence from the northern Irish Sea basin. Boreas 2007, 36, 115–129. [Google Scholar] [CrossRef]
- Thomas, G.S.P. The Quaternary of the Isle of Man. In The Quaternary History of the Irish Sea; Kidson, C., Tooley, M.J., Eds.; Seel House Press: Liverpool, UK, 1977; pp. 155–178. [Google Scholar]
- Douglas, T.D. Periglacial involutions and the evidence for coversands in the English midlands. Proc. Yorks. Geol. Soc. 1982, 44, 131–143. [Google Scholar] [CrossRef]
- Chadwick, R.A.; Jackson, D.I.; Barnes, R.P.; Kimbell, G.S.; Johnson, H.; Chiverell, R.C.; Thomas, G.S.P.; Jones, N.S.; Riley, N.J.; Pickett, E.A.; et al. The Geology of the Isle of Man and Its Offshore Area: Quaternary; British Geological Survey Report RR/01/06; British Geological Survey and Treasury, Isle of Man: Keyworth, UK, 2001; pp. 107–116. [Google Scholar]
- Roberts, D.H.; Chiverrell, R.C.; Innes, J.B.; Horton, B.P.; Brooks, A.J.; Thomas, G.S.P.; Turner, S.; Gonzalez, S. Holocene sea levels, Last Glacial Maximum glaciomarine environments and geophysical models in the northern Irish Sea Basin, UK. Mar. Geol. 2006, 231, 113–128. [Google Scholar] [CrossRef]
- Shennan, I.; Horton, B.P. Holocene land- and sea-level changes in Great Britain. J. Quat. Sci. 2002, 17, 511–526. [Google Scholar] [CrossRef]
- Erdtman, G. Studies in the postarctic history of the forests of Northwestern Europe. I. Investigations in the British Isles. Geol. Fören. Stock. Förhand. 1928, 50, 123–192. [Google Scholar] [CrossRef]
- Mitchell, G.F. A Late-Glacial deposit near Ballaugh, Isle of Man. New Phytol. 1958, 57, 256–263. [Google Scholar] [CrossRef]
- Dickson, C.A.; Dickson, J.H.; Mitchell, G.F. The Late Weichselian flora of the Isle of Man. Philos. Trans. R. Soc. B 1970, 258, 31–79. [Google Scholar]
- Erdtman, G. Pollen statistics from The Curragh and Ballaugh, Isle of Man. Proc. Liverpool Geol. Soc. 1925, 14, 158–163. [Google Scholar]
- Tooley, M.J. Flandrian sea-level changes and vegetational history of the Isle of Man: A review. In Man and Environment in the Isle of Man; Davey, P., Ed.; British Archaeological Reports 54; BAR Publishing: Oxford, UK, 1978; pp. 15–24. [Google Scholar]
- Innes, J.B. The Dhoo Valley, Isle of Man: A Palaeo-Environmental Assessment; Centre for Manx Studies Research Report 2; Centre for Manx Studies: Douglas, Isle of Man, 1995. [Google Scholar]
- Gonzales, S.; Innes, J.B.; Huddart, D.; Davey, P.; Plater, A. Holocene coastal change in the north of the Isle of Man: Stratigraphy, palaeoenvironment and archaeological evidence. In Coastal and Estuarine Environments: Sedimentology, Geomorphology and Geoarchaeology; Geological Society Special Publication 175: London, UK, 2000; pp. 343–363. [Google Scholar]
- Innes, J.B.; Blackford, J.J.; Davey, P.J. Dating the introduction of cereal cultivation to the British Isles: Early palaeoecological evidence from the Isle of Man. J. Quat. Sci. 2003, 18, 603–613. [Google Scholar] [CrossRef]
- Innes, J.B.; Chiverrell, R.C.; Blackford, J.J.; Davey, P.J.; Gonzalez, S.; Rutherford, M.M.; Tomlinson, P. Earliest Holocene vegetation history and island biogeography of the Isle of Man, British Isles. J. Biogeogr. 2004, 31, 761–772. [Google Scholar] [CrossRef]
- Chiverrell, R.C.; Innes, J.B.; Blackford, J.J.; Woodcock, J.J.; Davey, P.J.; Tomlinson, P.R.; Rutherford, M.M.; Thomas, G.S.P. Palaeoecological and archaeological evidence for Bronze Age human activity on the Isle of Man. Holocene 2004, 14, 346–360. [Google Scholar] [CrossRef]
- Chiverrell, R.C.; Innes, J.B.; Blackford, J.J. Vegetation history of the Isle of Man. In A New History of the Isle of Man. Vol. I. The Evolution of the Natural Landscape; Chiverrell, R.C., Thomas, G.S.P., Eds.; Liverpool University Press: Liverpool, UK, 2006; pp. 300–326. [Google Scholar]
- Davey, P.J.; Tomlinson, P.R. Archaeology and landscape change at Port Cranstal, Bride, Isle of Man. Mes. Misc. 2017, 25, 11–25. [Google Scholar]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis; Blackwell Scientific Publications: Oxford, UK, 1991. [Google Scholar]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis; John Wiley & Sons: Chichester, UK, 1989. [Google Scholar]
- Stace, C.A. New Flora of the British Isles; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Bennett, K.D.; Whittington, G.; Edwards, K.J. Recent plant nomenclatural changes and pollen morphology in the British Isles. Quat. Newsl. 1994, 73, 1–6. [Google Scholar]
- Grimm, E.C. TILIA Software; Illinois State Museum: Chicago, IL, USA, 1993. [Google Scholar]
- Grimm, E.C. TGView v. 2.0.2, Software; Illinois State Museum, Research and Collections Center: Springfield, IL, USA, 2004. [Google Scholar]
- Reimer, P.; Austin, W.; Bard, E.; Bayliss, A.; Blackwell, P.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Lamplugh, G.W. The Geology of the Isle of Man; Memoir of the Geological Survey of England and Wales; HMSO: London, UK, 1903.
- Chiverrell, R.C.; Smedley, R.K.; Small, D.; Ballantyne, C.K.; Burke, M.J.; Callard, S.L.; Clark, C.D.; Duller, G.A.T.; Evans, D.J.A.; Fabel, D.; et al. Ice margin oscillations during deglaciation of the northern Irish Sea basin. J. Quat. Sci. 2018, 33, 739–762. [Google Scholar] [CrossRef]
- Scourse, J.D.; Chiverrell, R.C.; Smedley, R.K.; Small, D.; Burke, M.J.; Saher, M.; Van Landeghem, K.J.J.; Duller, G.A.T.; Ó Cofaigh, C.; Bateman, M.D.; et al. Maximum extent and ecosystem dynamics of the Irish Sea Ice Stream and Irish Sea glacier since the Last Glacial Maximum. J. Quat. Sci. 2021, 36, 780–804. [Google Scholar] [CrossRef]
- Davey, P.J.; Blackford, J.J.; Chiverrell, R.C.; Innes, J.B.; Tomlinson, P.R. The Curragh and other lowland wetlands of the Isle of Man. In Wetlands in the Landscape: Archaeology, Conservation, Heritage; Atherden, M., Ed.; PLACE Research Centre: York, UK, 2001; pp. 12–40. [Google Scholar]
- Chiverrell, C.; Thomas, G.S.P.; Harvey, A.M. Late Devensian and Holocene landscape change in the uplands of the Isle of Man. Geomorphology 2001, 40, 219–236. [Google Scholar] [CrossRef]
- Chiverrell, R.C. Climate change: The evidence from Beinn y Phott. In A New History of the Isle of Man. Vol. I. The Evolution of the Natural Landscape; Chiverrell, R.C., Thomas, G.S.P., Eds.; Liverpool University Press: Liverpool, UK, 2006; pp. 282–285. [Google Scholar]
- Blackford, J.J.; Innes, J.B. Peel Embayment Area Palynological Assessment; Report for the Centre for Manx Studies; University of Liverpool: Liverpool, UK, 1996. [Google Scholar]
- Chiverrell, R.C.; Davey, P.J.; Gowlett, J.A.J.; Woodcock, J.J. Radiocarbon dates for the Isle of Man. In Recent Archaeological Research on the Isle of Man; Davey, P.J., Ed.; BAR British Series 278; Archaeopress: Oxford, UK, 1999; pp. 321–336. [Google Scholar]
- Tipping, R.M. The prospects for establishing synchroneity in the early postglacial pollen peak of Juniperus in the British Isles. Boreas 1987, 16, 155–163. [Google Scholar] [CrossRef]
- Birks, H.J.B. Holocene (Flandrian) chronostratigraphy of the British Isles: A review. Striae 1982, 16, 99–105. [Google Scholar]
- Innes, J.B. Introduction to the Holocene (Flandrian) history and record of Northern England. In Quaternary of Northern England; Huddart, D., Glasser, N.F., Eds.; Geological Conservation Review Series 25; Joint Nature Conservation Committee: Peterborough, UK, 2002; pp. 351–365. [Google Scholar]
- Cowell, R.W.; Innes, J.B. The Wetlands of Merseyside; North-West Wetlands Survey 1; English Heritage and Lancaster University: Lancaster, UK, 1994. [Google Scholar]
- Godwin, H.; Walker, D.; Willis, E.H. Radiocarbon dating and post-glacial vegetational history: Scaleby Moss. Proc. R. Soc. Lond. B 1957, 147, 353–366. [Google Scholar]
- Moar, N.T. Late Weichselian and Flandrian pollen diagrams from South-West Scotland. New Phytol. 1969, 68, 433–467. [Google Scholar] [CrossRef]
- Hibbert, F.A.; Switsur, V.R.; West, R.G. Radiocarbon dating of Flandrian pollen zones at Red Moss, Lancashire. Proc. R. Soc. Lond. B. Biol. Sci. 1971, 177, 161–176. [Google Scholar]
- Innes, J.B. Radiocarbon (AMS) dating of the Betula and Alnus pollen rises and Holocene woodland history in the Isle of Man. Bull. Brit. Ecol. Soc. 2003, 34, 37–38. [Google Scholar]
- Pennington, W. Lags in adjustment of vegetation to climate caused by the pace of soil development: Evidence from Britain. Vegetatio 1986, 67, 105–118. [Google Scholar] [CrossRef]
- Birks, H.J.B. Late-Quaternary biotic changes in terrestrial and lacustrine environments, with particular reference to north-west Europe. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B.E., Ed.; John Wiley: Chichester, UK, 1986; pp. 3–65. [Google Scholar]
- Smith, A.G. The influence of Mesolithic and Neolithic man on British vegetation: A discussion. In Studies in the Vegetational History of the British Isles; Walker, D., West, R.G., Eds.; Cambridge University Press: Cambridge, UK, 1970; pp. 81–96. [Google Scholar]
- Huntley, B. Rapid early-Holocene migration and high abundance of hazel (Corylus avellana L.): Alternative hypotheses. In Climate Change and Human Impact on the Landscape; Chambers, F.M., Ed.; Chapman and Hall: London, UK, 1993; pp. 205–216. [Google Scholar]
- Linnman, G. Some aspects of the colonization of Corylus avellana L. in north-west Europe during early Flandrian times. Striae 1981, 14, 72–75. [Google Scholar]
- Tallantire, P.A. The early-Holocene spread of hazel (Corylus avellana L.) in Europe north and west of the Alps: An ecological hypothesis. Holocene 2002, 12, 81–96. [Google Scholar] [CrossRef]
- Finsinger, W.; Tinner, W.; van der Knaap, W.O.; Ammann, B. The expansion of hazel (Corylus avellana L.) in the southern Alps: A key for understanding its early Holocene history in Europe. Quat. Sci. Rev. 2006, 25, 612–631. [Google Scholar] [CrossRef]
- Morrison, M.E.S.; Stephens, N. A submerged late-Quaternary deposit at Roddans Port, on the north-east coast of Ireland. Philos. Trans. R. Soc. B 1965, 249, 221–255. [Google Scholar]
- Bennett, K.D. Post-glacial dynamics of pine (Pinus sylvestris L.) and pinewoods in Scotland. In Our Pinewood Heritage; Aldhous, J.R., Ed.; Forestry Commission, The Royal Society for the Protection of Birds, Scottish Natural Heritage: Inverness, UK, 1995; pp. 23–29. [Google Scholar]
- Gear, A.J.; Huntley, B. Rapid changes in the range limits of Scots Pine 4000 years ago. Science 1991, 251, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.D. The post-glacial history of Pinus sylvestris in the British Isles. Quat. Sci. Rev. 1984, 3, 133–155. [Google Scholar] [CrossRef]
- Bennett, K.D.; Birks, H.J.B. Postglacial history of alder (Alnus glutinosa (L.) Gaertn.) in the British Isles. J. Quat. Sci. 1990, 5, 123–133. [Google Scholar] [CrossRef]
- Smith, A.G.; Goddard, I.C. A 12 500 year record of vegetational history at Sluggan Bog, Co. Antrim, N. Ireland (incorporating a pollen zone scheme for the non-specialist). New Phytol. 1991, 118, 167–187. [Google Scholar] [CrossRef]
- Parker, A.G.; Goudie, A.S.; Anderson, D.E.; Robinson, M.A.; Bonsall, C. A review of the mid-Holocene elm decline in the British Isles. Prog. Phys. Geogr. 2002, 26, 1–45. [Google Scholar] [CrossRef]
- Perry, I.; Moore, P.D. Dutch elm disease as an analogue of Neolithic Elm Decline. Nature 1987, 326, 72–73. [Google Scholar] [CrossRef]
- Flynn, L.E.; Mitchell, F.J.G. Comparison of a recent Elm Decline with the mid-Holocene Elm Decline. Veget. Hist. Archaeobot. 2019, 28, 391–398. [Google Scholar] [CrossRef]
- Waller, M. Drought, disease, defoliation and death: Forest pathogens as agents of past vegetation change. J. Quat. Sci. 2013, 28, 336–342. [Google Scholar] [CrossRef]
- Kearney, K.; Gearey, B.J. The Elm Decline is dead! Long live declines in elm: Revisiting the chronology of the Elm Decline in Ireland and its association with the Mesolithic/Neolithic Transition. Env. Arch. 2020. [Google Scholar] [CrossRef]
- Lamb, H.; Thompson, A. Unusual mid-Holocene abundance of Ulmus in western Ireland—Human impact in the absence of a pathogen? Holocene 2005, 15, 447–452. [Google Scholar] [CrossRef]
- Pilcher, J.R. Pollen analysis and radiocarbon dating of a peat on Slieve Gallion, Co. Tyrone, N. Ireland. New Phytol. 1973, 72, 681–689. [Google Scholar] [CrossRef]
- Walker, R. Diatom and sediment studies of a sediment profile from Melynllyn, a mountain tarn in Snowdonia, North Wales. New Phytol. 1978, 81, 791–804. [Google Scholar] [CrossRef]
- Tallis, J.H.; Switsur, V.R. Forest and moorland in the South Pennine uplands in the mid-Flandrian period. II. The hillslope forests. J. Ecol. 1990, 78, 857–883. [Google Scholar] [CrossRef]
- Mighall, T.M.; Chambers, F.M. Holocene vegetation history and human impact at Bryn y Castell, Snowdonia, north Wales. New Phytol. 1995, 130, 299–321. [Google Scholar] [CrossRef]
- Pigott, C.D.; Huntley, J.P. Factors controlling the distribution of Tilia cordata at the northern limits of its geographical range. I. Distribution in north-west England. New Phytol. 1978, 81, 429–441. [Google Scholar] [CrossRef]
- Pigott, C.D.; Huntley, J.P. Factors controlling the distribution of Tilia cordata at the northern limits of its geographical range. II. History in North-West England. New Phytol. 1980, 84, 145–164. [Google Scholar] [CrossRef]
- Lloyd, J.M.; Zong, Y.; Fish, P.; Innes, J.B. Holocene and Lateglacial relative sea-level change in north-west England: Implications for glacial isostatic adjustment models. J. Quat. Sci. 2013, 28, 59–70. [Google Scholar] [CrossRef]
- Bradley, S.L.; Milne, G.A.; Shennan, I.; Edwards, R. An improved glacial isostatic adjustment model for the British Isles. J. Quat. Sci. 2011, 26, 541–552. [Google Scholar] [CrossRef]
- Wingfield, R.T.R. A model of sea-level in the Irish and Celtic seas during the end-Pleistocene to Holocene transition. In Island Britain: A Quaternary Perspective; Preece, R.C., Ed.; Special Publication 96; Geological Society London: London, UK, 1995; pp. 209–242. [Google Scholar]
- Lambeck, K. Glaciation and sea-level change for Ireland and the Irish Sea since Late Devensian time. J. Geol. Soc. Lond. 1996, 153, 853–872. [Google Scholar] [CrossRef]
- Innes, J.B.; Roberts, D.R.; Chiverrell, R.C. Coastal and sea level history. In A New History of the Isle of Man. Vol. I. The Evolution of the Natural Landscape; Chiverrell, R.C., Thomas, G.S.P., Eds.; Liverpool University Press: Liverpool, UK, 2006; pp. 286–296. [Google Scholar]
- Huddart, D.; Tooley, M.J.; Carter, P.A. The coasts of northwest England. In The Quaternary History of the Irish Sea; Kidson, C., Tooley, M.J., Eds.; Seel House Press: Liverpool, UK, 1977; pp. 119–154. [Google Scholar]
- Bennett, K.D. Insularity and the Quaternary tree and shrub flora of the British Isles. In Island Britain: A Quaternary Perspective; Preece, R.C., Ed.; Geological Society Special Publication 96; The Geological Society: London, UK, 1995; pp. 173–180. [Google Scholar]
- Boyd, W.E.; Dickson, J.H. Patterns in the geographical distribution of the early Flandrian Corylus rise in southwest Scotland. New Phytol. 1986, 102, 615–623. [Google Scholar] [CrossRef]
- Devoy, R.J.N. Deglaciation, Earth crustal behaviour and sea-level changes in the determination of insularity: A perspective from Ireland. In Island Britain: A Quaternary Perspective; Preece, R.C., Ed.; Special Publication 96; Geological Society London: London, UK, 1995; pp. 181–208. [Google Scholar]
- Petit, R.J.; Bialozyt, R.; Garnier-Géré, P.; Hampe, A. Ecology and genetics of tree invasions: From recent introductions to Quaternary migrations. Forest Ecol. Manag. 2004, 197, 117–137. [Google Scholar] [CrossRef]
- Froyd, C.A. Fossil stomata reveal early pine presence in Scotland: Implications for postglacial colonization analyses. Ecology 2005, 86, 579–586. [Google Scholar] [CrossRef]
- Fossitt, J.A. Late-glacial and Holocene vegetation history of western Donegal, Ireland. Biol. Environ. Proc. R. Irish Acad. 1994, 94, 1–31. [Google Scholar]
- Bush, M.B.; Hall, A.R. Flandrian Alnus: Expansion or immigration? J. Biogeogr. 1987, 14, 479–481. [Google Scholar] [CrossRef]
- Oldfield, F. Studies in the post-glacial history of the British vegetation: Lowland Lonsdale. New Phytol. 1960, 59, 192–217. [Google Scholar] [CrossRef]
- Stolze, S.; Moneke, T. Holocene history of ‘non-native’ trees in Ireland. Rev. Palaeobot. Palynol. 2017, 244, 347–355. [Google Scholar] [CrossRef]
- Lisitsyna, O.V.; Gieseke, T.; Hicks, S. Exploring pollen percentage pollen threshold values as an indication for the regional presence of major European trees. Rev. Palaeobot. Palynol. 2011, 166, 311–324. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Madsen, B.J. Flandrian vegetational history of Little Loch Roag, Isle of Lewis, Scotland. J. Ecol. 1979, 67, 825–842. [Google Scholar] [CrossRef]
- Keatinge, T.H.; Dickson, J.H. Mid-Flandrian changes in vegetation on Mainland Orkney. New Phytol. 1979, 82, 585–612. [Google Scholar] [CrossRef]
- Tipping, R.M. Storminess as an explanation for the decline of pine woodland ca. 7400 years ago at Loch Tulla, western Scotland. Veget. Hist. Archaeobot. 2008, 17, 345–350. [Google Scholar] [CrossRef]
- Coope, G.R. Fossil Coleoptera from Glen Ballyre and their bearing upon the interpretation of late-Glacial environments. In Field Guide to the Isle of Man; Thomas, G.P.S., Ed.; Quaternary Research Association: Cambridge, UK, 1971; pp. 13–15. [Google Scholar]
- Coope, G.R.; Lemdahl, G.; Lowe, J.J.; Walking, A. Temperature gradients in northern Europe during the last glacial-Holocene transition (14–9 14C kyr BP) interpreted from coleopteran assemblages. J. Quat. Sci. 1998, 13, 419–433. [Google Scholar] [CrossRef]
- Walker, M.J.C.; Björck, S.; Lowe, J.J.; Cwynar, L.C.; Johnsen, S.; Knudsen, K.-L.; Wohlfarth, B.; INTIMATE Group. Isotopic ‘events’ in the GRIP ice core: A stratotype for the Late Pleistocene. Quat. Sci. Rev. 1999, 18, 1143–1150. [Google Scholar] [CrossRef]
- Feurdean, A.; Bhagwat, S.A.; Willis, K.J.; Birks, H.J.B.; Lischke, H.; Hickler, T. Tree migration-rates: Narrowing the gap between inferred post-glacial rates and projected rates. PLoS ONE 2013, 8, e71797. [Google Scholar] [CrossRef] [PubMed]
- Roberts, N.; Fyfe, R.M.; Woodbridge, J.; Gaillard, M.-J.; Davis, B.A.S.; Kaplan, J.O.; Marquer, L.; Mazier, F.; Nielsen, A.B.; Sugita, S.; et al. Europe’s lost forests: A pollen-based synthesis for the last 11,000 years. Sci. Rep. 2018, 8, 716. [Google Scholar] [CrossRef] [PubMed]
- Silvertown, J. History of a latitudinal diversity gradient: Woody plants in Europe 13,000–1000 years BP. J. Biogeogr. 1985, 12, 519–525. [Google Scholar] [CrossRef]
- Bradshaw, R.H.W. Forest response to Holocene climatic change: Equilibrium or non-equilibrium. In Climate Change and Human Impact on the Landscape; Chambers, F.M., Ed.; Chapman and Hall: London, UK, 1993; pp. 57–65. [Google Scholar]
- Geirsdóttir, A.; Miller, G.; Larsen, D.J.; Ólafsdóttir, S. Abrupt Holocene climate transitions in the northern North Atlantic region recorded by synchronized lacustrine records in Iceland. Quat. Sci. Rev. 2013, 70, 48–62. [Google Scholar] [CrossRef]
- Birks, H.J.B. Mind the gap: How open were European primeval forests? TREE 2005, 20, 154–156. [Google Scholar] [CrossRef]
- Mitchell, F.J.G. How open were European primeval forests? Hypothesis testing using palaeoecological data. J. Ecol. 2005, 93, 168–177. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Birks, H.H. The rise and fall of forests. Science 2004, 305, 484–485. [Google Scholar] [CrossRef]
- Kirby, K.J. A model of a natural wooded landscape in Britain as influenced by large herbivore activity. Forestry 2004, 77, 405–420. [Google Scholar] [CrossRef]
- Rasmussen, S.O.; Vinther, B.M.; Clausen, H.B.; Andersen, K.K. Early Holocene climate oscillations recorded in three Greenland ice cores, Quat. Sci. Rev. 2007, 26, 1907–1914. [Google Scholar] [CrossRef]
- Alley, R.B.; Ágứstsdóttir, A.M. The 8k event: Cause and consequences of a major Holocene abrupt climate change. Quat. Sci. Rev. 2005, 24, 1123–1149. [Google Scholar] [CrossRef]
- Fleitmann, D.; Mudelsee, M.; Burns, S.J.; Bradley, R.S.; Kramers, J.; Matter, A. Evidence for a widespread climatic anomaly at around 9.2 ka before present. Paleoceanography 2008, 23, PA1102. [Google Scholar] [CrossRef]
- Swindles, G.T.; Lawson, I.T.; Matthews, I.P.; Blaauw, M.; Daley, T.J.; Charman, D.J.; Roland, T.P.; Plunkett, G.; Schettler, G.; Gearey, B.R.; et al. Centennial-scale climate change in Ireland during the Holocene. Earth-Sci. Rev. 2013, 126, 300–320. [Google Scholar] [CrossRef]
- Marshall, J.D.; Jones, R.T.; Crowley, S.F.; Oldfield, F.; Nash, S.; Bedford, A. A high-resolution Lateglacial isotopic record from Hawes Water, Northwest England: Climatic oscillations, calibration and comparison of palaeotemperature proxies. Palaeogeogr. Palaeoclimat. Palaeoecol. 2002, 185, 25–40. [Google Scholar] [CrossRef]
- Marshall, J.D.; Lang, B.; Crowley, S.F.; Weedon, G.P.; van Calsteren, P.; Fisher, E.H.; Holme, R.; Holmes, J.A.; Jones, R.T.; Bedford, A.; et al. Terrestrial impact of abrupt changes in the North Atlantic thermohaline circulation: Early Holocene, UK. Geology 2007, 35, 639–642. [Google Scholar] [CrossRef]
- Jones, R.T.; Marshall, J.D.; Fisher, E.; Hatton, J.; Patrick, C.; Anderson, K.; Lang, B.; Bedford, A.; Oldfield, F. Controls on lake level in the early to mid-Holocene, Hawes Water, Lancashire, UK. Holocene 2011, 21, 1061–1072. [Google Scholar] [CrossRef]
- van der Plicht, J.; van Geel, B.; Bohncke, S.J.P.; Bos, J.A.A.; Blaauw, M.; Speranza, A.O.M.; Muscheler, R.; Björck, S. The Preboreal climate reversal and a subsequent solar-forced climate shift. J. Quat. Sci. 2004, 19, 263–269. [Google Scholar] [CrossRef]
- Bos, J.; van Geel, B.; van der Plicht, J.; Bohncke, S. Preboreal climate oscillations in Europe: Wiggle-match dating and synthesis of Dutch high-resolution multi-proxy records. Quat. Sci. Rev. 2007, 26, 1927–1950. [Google Scholar] [CrossRef]
- Renssen, H.; Goosse, H.; Fichefet, T.; Campin, J.-M. The 8.2 kyr BP event simulated by a global atmosphere-sea-ice-ocean model. Geophys. Res. Lett. 2001, 28, 1567–1570. [Google Scholar] [CrossRef]
- Li, H.; Renssen, H.; Roche, D.M.; Miller, P.A. Modelling the vegetation response to the 8.2 ka BP cooling event in Europe and northern Africa. J. Quat. Sci. 2019, 34, 650–661. [Google Scholar] [CrossRef]
- Andrews, M.V.; Beck, R.B.; Birks, H.J.B.; Gilbertson, D.D.; Switsur, V.R. The past and present vegetation of Oronsay and Colonsay. In Excavations on Oronsay, Prehistoric Human Ecology on a Small Island; Mellars, P., Ed.; Edinburgh University Press: Edinburgh, UK, 1987; Volume 1, pp. 52–77. [Google Scholar]
- Ghilardi, B.; O’Connell, M. Early Holocene vegetation and climate dynamics with particular reference to the 8.2 ka event: Pollen and macrofossil evidence from a small lake in western Ireland. Veget. Hist. Archaeobot. 2013, 22, 99–114. [Google Scholar] [CrossRef]
- O’Connell, M.; Ghilardi, B.; Morrison, L. A 7000-year record of environmental change, including early farming impact, based on lake-sediment geochemistry and pollen data from County Sligo, western Ireland. Quat. Res. 2014, 81, 35–49. [Google Scholar] [CrossRef]
- Tinner, W.; Lotter, A.F. Central European vegetation response to abrupt climate change at 8.2 ka. Geology 2001, 29, 551–554. [Google Scholar] [CrossRef]
- Veski, S.; Seppä, H.; Ojala, A.E.K. Cold event at 8200 yr BP recorded in annually laminated lake sediments in eastern Europe. Geology 2004, 32, 681–684. [Google Scholar] [CrossRef]
- Hede, M.U.; Rasmussen, P.; Noe-Nygaard, N.; Clarke, A.L.; Vinebrooke, R.D.; Olsen, J. Multiproxy evidence for terrestrial and aquatic ecosystem responses during the 8.2 ka cold event as recorded at Højby Sø, Denmark. Quat. Res. 2010, 73, 485–496. [Google Scholar] [CrossRef]
- Crombé, P. Abrupt cooling events in the early Holocene and their potential impact on the environment and human behaviour along the southern North Sea basin (NW Europe). J. Quat. Sci. 2018, 33, 353–367. [Google Scholar] [CrossRef]
- Paus, A.; Haflidason, H.; Routh, J.; Naafs, B.; Thoen, M.W. Environmental responses to the 9.7 and 8.2 events at two ecotonal sites in the Dovre Mountains, mid-Norway. Quat. Sci. Rev. 2019, 205, 45–61. [Google Scholar] [CrossRef]
- Edwards, K.J.; Langdon, P.G.; Sugden, H. Separating climatic and possible human impacts in the early Holocene: Biotic response around the time of the 8200 cal. BP event. J. Quat. Sci. 2007, 22, 77–84. [Google Scholar] [CrossRef]
- Seppä, H.; Birks, H.J.B.; Gieseke, T.; Hammarlund, D.; Alenius, T.; Antonsson, K.; Bjune, A.E.; Heikkilä, M.; MacDonald, G.M.; Ojala, A.E.K.; et al. Spatial structure of the 8200 cal yr BP event in northern Europe. Clim. Past 2007, 3, 225–236. [Google Scholar] [CrossRef]
- Davis, M.B.; Botkin, D.B. Sensitivity of cool-temperate forests and their fossil pollen record to rapid temperature change. Quat. Res. 1985, 23, 327–340. [Google Scholar] [CrossRef]
- Hughes, P.D.M.; Mauquoy, D.; Barber, K.E.; Langdon, P.G. Mire-development pathways and palaeoclimatic records from a full Holocene peat archive at Walton Moss, Cumbria, England. Holocene 2000, 10, 465–479. [Google Scholar] [CrossRef]
- Seppä, H.; Schurgers, G.; Miller, P.A.; Bjune, A.E.; Giesecke, T.; Kühl, N.; Renssen, H.; Salonen, J.S. Trees tracking a warmer climate: The Holocene range shift of hazel (Corylus avellana) in northern Europe. Holocene 2015, 25, 53–63. [Google Scholar] [CrossRef]
- Pennington, W. Palaeolimnology in the English Lakes—Some questions and answers over fifty years. Hydrobiologia 1991, 214, 9–24. [Google Scholar] [CrossRef]
- Chambers, F.M.; Elliott, L. Spread and expansion of Alnus Mill. In the British Isles: Timing, agencies and possible vectors. J. Biogeogr. 1989, 16, 541–550. [Google Scholar] [CrossRef]
- Barber, K.E.; Chambers, F.M.; Maddy, D. Holocene palaeoclimates from peat stratigraphy: Macrofossil proxy climate records from three oceanic raised bogs in England and Ireland. Quat. Sci. Rev. 2003, 22, 521–539. [Google Scholar] [CrossRef]
- Chambers, F.M.; Price, S.-M. Palaeoecology of Alnus (Alder): Early post-glacial rise in a valley mire, north-west Wales. New Phytol. 1985, 101, 333–344. [Google Scholar] [CrossRef]
- Tipping, R.M. Holocene evolution of a lowland Scottish landscape: Kirkpatrick Fleming. Part I. Peat and pollen-stratigraphic evidence for raised moss development and climatic change. Holocene 1995, 5, 69–81. [Google Scholar] [CrossRef]
- Cayliss, S.M.; Tipping, R.M. Data on mid-Holocene climatic, vegetation and anthropogenic interactions at Stanshiel Rig, southern Scotland. Veget. Hist. Archaeobot. 2002, 11, 201–210. [Google Scholar] [CrossRef]
- Langdon, P.G.; Barber, K.E.; Hughes, P.D.M. A 7500-year, peat-based palaeoclimatic reconstruction and evidence for an 1100-year cyclicity in bog surface wetness from Temple Hill Moss, Pentland Hills, SE Scotland. Quat. Sci. Rev. 2003, 22, 259–274. [Google Scholar] [CrossRef]
- McCarroll, J.; Chambers, F.M.; Webb, J.C.; Thom, T. Informing innovative peatland conservation in light of palaeoecological evidence for the demise of Sphagnum imbricatum: The case of Oxenhope Moor, Yorkshire, UK. Mires Peat 2016, 18, 1–24. [Google Scholar]
- Bonsall, C.; Macklin, M.G.; Anderson, C.E.; Payton, R.W. Climate change and the adoption of agriculture in north-west Europe. Eur. J. Arch. 2002, 5, 9–23. [Google Scholar] [CrossRef]
- Tipping, R.M. The case for climatic stress forcing choice in the adoption of agriculture in the British Isles. In Landscapes in Transition; Finlayson, B., Warren, G., Eds.; Levant Supplementary Series 8; Oxbow Books: Oxford, UK, 2010; pp. 66–76. [Google Scholar]
- Kear, B.S. Soil development and soil patterns in north-west England. In The Geomorphology of North-West England; Johnson, R.H., Ed.; Manchester University Press: Manchester, UK, 1985; pp. 80–93. [Google Scholar]
- Wilson, P.; Bateman, R.M.; Catt, J.A. Petrography, origin and environment of deposition of the Shirdley Hill Sand of south-west Lancashire, England. Proc. Geol. Assoc. 1981, 92, 211–229. [Google Scholar] [CrossRef]
- Innes, J.B.; Tooley, M.J.; Tomlinson, P.R. A comparison of the age and palaeoecology of some sub-Shirdley Hill Sand peat deposits from Merseyside and south-west Lancashire. Naturalist 1989, 114, 65–69. [Google Scholar]
- Theuerkauf, M.; Bos, J.A.A.; Jahns, S.; Janke, W.; Kuparinen, A.; Stebich, M.; Joosten, H. Corylus expansion and persistent openness in the early Holocene vegetation of northern central Europe. Quat. Sci. Rev. 2014, 90, 183–198. [Google Scholar] [CrossRef]
- Hodgkinson, D.; Huckerby, E.; Middleton, R.; Wells, C.E. The Lowland Wetlands of Cumbria; North-West Wetlands Survey 6; English Heritage and Lancaster University: Lancaster, UK, 2000. [Google Scholar]
- Birks, H.J.B. Mid-Flandrian forest history of Roudsea Wood National Nature Reserve, Cumbria. New Phytol. 1982, 90, 339–354. [Google Scholar] [CrossRef]
- Bradshaw, R.H.W.; Browne, P. Changing patterns in the post-glacial distribution of Pinus sylvestris in Ireland. J. Biogeogr. 1987, 14, 237–248. [Google Scholar] [CrossRef]
- Green, D.G. Fire and stability in the postglacial forests of southwest Nova Scotia. J. Biogeogr 1982, 9, 29–40. [Google Scholar] [CrossRef]
- Bennett, K.D. Postglacial population expansion of forest trees in Norfolk, UK. Nature 1983, 303, 164–167. [Google Scholar] [CrossRef]
- Bennett, K.D. Competitive interactions among forest tree populations in Norfolk, England during the last 10,000 years. New Phytol. 1986, 103, 603–620. [Google Scholar] [CrossRef]
- Bennett, K.D.; Lamb, H.F. Holocene pollen sequences as a record of competitive interactions among tree populations. TREE 1988, 3, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, R.; Hannon, G. The Holocene structure of north-west European forest induced from palaeoecological data. In Forest Biodiversity: Lessons from History for Conservation; Honnay, O., Verheyen, K., Bossuyt, B., Hermy, M., Eds.; CAB International: Oxford, UK, 2004; pp. 11–25. [Google Scholar]
- Mitchell, F.J.G. The investigation of long-term successions in temperate woodland using fine spatial resolution pollen analysis. In The Ecological History of European Forests; Kirby, K.J., Watkins, C., Eds.; CAB International: Wallingford, UK, 1998; pp. 213–223. [Google Scholar]
- Bennett, K.D. Post-glacial vegetation history: Ecological considerations. In Vegetation History; Huntley, B., Webb, T., III, Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 699–724. [Google Scholar]
- Smith, A.G. Problems of inertia and threshold related to post-glacial habitat changes. Proc. Roy. Soc. Lond. B 1965, 161, 331–342. [Google Scholar]
- Davis, M.B. Quaternary history and the stability of forest communities. In Forest Succession: Concepts and applications; West, D.C., Shugart, H.H., Botkin, D.B., Eds.; Springer: New York, NY, USA, 1981; pp. 132–153. [Google Scholar]
- Von Holle, B.; Delcourt, H.R.; Simberloff, D. The importance of biological inertia in plant community resistance to invasion. J. Veg. Sci. 2003, 14, 425–432. [Google Scholar] [CrossRef]
- Barclay, R.; Ferreira, C.; Ballantyne, E.; Tipping, R.; Tisdall, E. Possibly climatically driven, later prehistoric woodland decline on Ben Lomond, central Scotland. Veget. Hist. Archaeobot. 2022. [Google Scholar] [CrossRef]
- Allen, J.R.L. Trees and their response to wind: Mid-Flandrian strong winds, Severn Estuary and inner Bristol Channel, Southwest Britain. Philos. Trans. R. Soc. Lond. 1992, B338, 335–364. [Google Scholar]
- Sacheverell, W. An Account of the Isle of Man: Its Inhabitants, Language, Soil, Remarkable Curiosities, the Succession of Its Kings and Bishops, down to the Present Time; Hartley: London, England, 1702. [Google Scholar]
- Innes, J.B.; Blackford, J.J.; Simmons, I.G. Woodland disturbance and possible land-use regimes during the Late Mesolithic in the English uplands: Pollen, charcoal and non-pollen palynomorph evidence from Bluewath Beck, North York Moors, UK. Veget. Hist. Archaeobot. 2010, 19, 439–452. [Google Scholar] [CrossRef]
- Ryan, P.; Blackford, J.J. Late Mesolithic environmental change at Black Heath, South Pennines, UK: A test of Mesolithic woodland management models using pollen, charcoal and non-palynomorph data. Veget. Hist. Archaeobot. 2010, 19, 545–558. [Google Scholar] [CrossRef]
- Innes, J.B.; Blackford, J.J.; Simmons, I.G. Late Mesolithic and early Neolithic forest disturbance: A high-resolution palaeoecological test of human impact hypotheses. Quat. Sci. Rev. 2013, 77, 80–100. [Google Scholar] [CrossRef]
- Bennett, K.D.; Simonson, A.B.; Peglar, S.M. Fire and man in postglacial woodlands of eastern England. J. Archaeol. Sci. 1990, 17, 635–642. [Google Scholar] [CrossRef]
- Simmons, I.G.; Innes, J.B. Mid-Holocene adaptations and Later Mesolithic forest disturbance in northern England. J. Archaeol. Sci. 1987, 14, 385–403. [Google Scholar] [CrossRef]
- Simmons, I.G. The Environmental Impact of Later Mesolithic Cultures; Edinburgh University Press: Edinburgh, UK, 1996. [Google Scholar]
- Rowley-Conwy, P.A.; Layton, R. Foraging and farming as niche construction: Stable and unstable adaptations. Philos. Trans. R. Soc. B 2011, 366, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Woodman, P.C. Excavations at Cass ny Hawin, a Manx Mesolithic site, and the position of the Manx microlithic industries. Proc. Prehist. Soc. 1987, 53, 1–22. [Google Scholar] [CrossRef]
- McCartan, S.B. The Mesolithic in the Isle of Man: An Island Perspective. In Mesolithic Scotland and Its Neighbours; Savile, A., Ed.; Society of Antiquaries of Scotland: Edinburgh, UK, 2004; pp. 271–283. [Google Scholar]
- Woodman, P.C.; McCarthy, M.; Monaghan, N. The Irish Quaternary fauna project. Quat. Sci. Rev. 1997, 16, 129–159. [Google Scholar] [CrossRef]
- Bishop, R.R.; Church, M.J.; Rowley-Conwy, P.A. Firewood, food and human niche construction: The potential role of Mesolithic hunter-gatherers in actively structuring Scotland’s woodlands. Quat Sci. Rev. 2015, 108, 51–75. [Google Scholar] [CrossRef]
- Atkinson, M.D. Betula pendula Roth. (B. verrucosa Ehrh.) and B. pubescens Ehrh. J. Ecol. 1992, 80, 837–870. [Google Scholar] [CrossRef]
- Birks, H.H. Studies in the vegetational history of Scotland. IV. Pine stumps in the Scottish blanket peats. Philos. Trans. R. Soc. Lond. B 1975, 270, 181–226. [Google Scholar]
- Smith, A.G. Newferry and the Boreal-Atlantic Transition. New Phytol. 1984, 98, 35–55. [Google Scholar] [CrossRef] [PubMed]
- Middleton, R.; Wells, C.E.; Huckerby, E. The Wetlands of North Lancashire; North West Wetlands Survey 7, Lancaster Imprints 20; English Heritage & Lancaster University: Lancaster, UK, 1995. [Google Scholar]
- McCartan, S.B. Lhen Trench: Aspects of the prehistoric heritage. In The Quaternary of the Isle of Man & North West England: Field Guide; Chiverrell, R.C., Plater, A.J., Thomas, G.S.P., Eds.; Quaternary Research Association: London, UK, 2004; pp. 106–111. [Google Scholar]
- Tipping, R.M.; Milburn, P. Mid-Holocene charcoal fall in southern Scotland—Temporal and spatial variability. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 164, 177–193. [Google Scholar] [CrossRef]
- Darvill, T. Billown Neolithic enclosures, Isle of Man. In Enclosures in Neolithic Europe: Essays on Causewayed and Non-Causewayed Sites; Varndell, G., Topping, P., Eds.; Oxbow Books: Oxford, UK, 2002; pp. 83–90. [Google Scholar]
- Woodbridge, J.; Fyfe, R.M.; Roberts, N.; Downey, S.; Edinborough, K.; Shennan, S. The impact of the Neolithic agricultural transition in Britain: A comparison of pollen-based land-cover and archaeological 14C date-inferred population change. J. Archaeol. Sci. 2014, 51, 216–224. [Google Scholar] [CrossRef]
- Davey, P.J.; Innes, J.B. The early Neolithic and the Manx environment. In Neolithic Settlement in Ireland and Western Britain; Armit, I., Murphy, E., Nelis, E., Simpson, D., Eds.; Oxbow Books: Oxford, UK, 2003; pp. 120–127. [Google Scholar]
- Garrow, D.; Sturt, F. Grey waters bright with Neolithic argonauts? Maritime connections and the Mesolithic-Neolithic Transition within the ‘western seaways’ of Britain, c.5000–3500 BC. Antiquity 2011, 85, 59–72. [Google Scholar] [CrossRef]
- Edwards, K.J. The anthropogenic factor in vegetational history. In The Quaternary History of Ireland; Edwards, K.J., Warren, W.P., Eds.; Academic Press: London, UK, 1985; pp. 187–220. [Google Scholar]
- Williams, E. Dating the introduction of food production into Britain and Ireland. Antiquity 1989, 63, 510–521. [Google Scholar] [CrossRef]
- Tipping, R.M. The form and fate of Scotland’s woodlands. Proc. Soc. Antiq. Scot. 1994, 124, 1–54. [Google Scholar]
- Innes, J.B.; Blackford, J.J.; Chambers, F.M. Kretzschmaria deusta and the North-West European mid-Holocene Ulmus decline at Moel y Gerddi, North Wales, UK. Palynology 2006, 30, 121–132. [Google Scholar]
- Rowley-Conwy, P.A. Forest grazing and clearance in temperate Europe with special reference to Denmark: An archaeological view. In Archaeological Aspects of Woodland Ecology; Bell, M., Limbrey, S., Eds.; British Archaeological Reports International Series, 146; BAR Oxford: Oxford, UK, 1982; pp. 199–215. [Google Scholar]
- Edwards, K.J. Models of forest farming for north-west Europe. In Climate Change and Human Impact on the Landscape; Chambers, F.M., Ed.; Chapman and Hall: London, UK, 1993; pp. 134–155. [Google Scholar]
- Darvill, T. Billown Neolithic Landscape Project 1995–1997. In Recent Archaeological Research on the Isle of Man; Davey, P.J., Ed.; BAR British Series 278; Archaeopress: Oxford, UK, 1999; pp. 13–26. [Google Scholar]
- Darvill, T. Billown and the Neolithic of the Isle of Man. In Neolithic Settlement in Ireland and Western Britain; Armit, I., Murphy, E., Nelis, E., Simpson, D., Eds.; Oxbow Books: Oxford, UK, 2003; pp. 112–119. [Google Scholar]
- Peglar, S.M.; Birks, H.J.B. The mid-Holocene Ulmus fall at Diss Mere, southeast England—Disease and human impact. Veget. Hist. Archaeobot. 1993, 2, 61–68. [Google Scholar] [CrossRef]
- Grosvenor, M.J.; Jones, R.T.; Turney, C.S.M.; Charman, D.J.; Hogg, A.; Coward, D.; Wilson, R. Human activity was a major driver of the mid-Holocene vegetation change in southern Cumbria: Implications for the elm decline in the British Isles. J. Quat. Sci. 2017, 32, 934–945. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollen Feature | 14C Date BP | Lab. Code | 2δ Age Range (cal. BP) | Mid-Range Age (cal. BP) | Site |
|---|---|---|---|---|---|
| Ulmus decline | 4785 ± 55 | AA-39112 | 5320–5600 | 5460 | Montpellier Bog |
| >4980 ± 50 | AA-34512 | 5590–5900 | 5745 | Port-y-Candas | |
| 5200 ± 35 | Poz-88363 | 5900–6170 | 6035 | Ballachrink LT12 | |
| 5310 ± 70 | AA-28379 | 5930–6280 | 6105 | The Cronk | |
| 5313 ± 38 | UB-3555 | 5940–6270 | 6105 | Dhoo Valley 16 | |
| Pinus rise | <6860 ± 55 | AA-28383 | 7580–7830 | 7705 | Port Cranstal |
| 7020 ± 35 | SUERC-2603 | 7750–7940 | 7845 | Quarry Bends | |
| 7045 ± 35 | SUERC-2604 | 7790–7960 | 7875 | Pollies | |
| 7470± 35 | SUERC-2620 | 8200–8370 | 8285 | Montpellier Bog | |
| Main Alnus rise | 6680 ± 55 | AA-45405 | 7430–7670 | 7550 | Montpellier Bog |
| >6730 ± 100 | AA-29738 | 7420–7780 | 7600 | Ballaclucas LT5 | |
| 6865 ± 45 | AA-52529 | 7600–7830 | 7715 | Lezayre Curragh | |
| <6860 ± 55 | AA-28383 | 7580–7830 | 7705 | Port Cranstal | |
| 7045 ± 35 | SUERC-2604 | 7790–7960 | 7875 | Pollies | |
| 7105 ± 80 | AA-29740 | 7730–8170 | 7950 | Ballachrink LT12 | |
| 7370 ± 35 | SUERC-2602 | 8030–8320 | 8175 | Quarry Bends | |
| Main Ulmus and | 7540 ± 75 | AA-29741 | 8180–8520 | 8350 | Ballachrink LT12 |
| Quercus rises * | 8115 ± 65 | AA-45404 | 8770–9290 | 9030 | Montpellier Bog |
| 8220 ± 35 | SUERC-2605 | 9020–9400 | 9210 | Pollies | |
| 8265 ± 40 | SUERC-2600 | 9030–9420 | 9225 | Curragh y Cowle | |
| 8290 ± 35 | AA-29335 | 9130–9430 | 9280 | Ballaugh Curragh 21 | |
| <8770 ± 50 | AA-52525 | 9550–10120 | 9835 | Ramsey Harbour | |
| Main Corylus rise | 8160 ± 75 | AA-29742 | 8780–9410 | 9095 | Ballachrink LT12 |
| 8870 ± 85 | AA-29737 | 9670–10,230 | 9950 | Curragh y Cowle | |
| 9030 ± 40 | SUERC-2611 | 9960–10,250 | 10,105 | Ballaugh Curragh 21 | |
| 9110 ± 35 | SUERC-2606 | 10,190–10,380 | 10,285 | Pollies | |
| <9150 ± 70 | AA-36488 | 10,200–10,500 | 10,350 | Montpellier Bog | |
| >9390 ± 40 | SUERC-2615 | 10,500–10,720 | 10,610 | Ramsey Harbour | |
| Main Betula rise | 9275 ± 50 | AA-48013 | 10,260–10,580 | 10,420 | Curragh y Cowle |
| 9320 ± 55 | AA-48012 | 10,300–10,690 | 10,495 | Pollies | |
| Juniperus peak | 9450 ± 100 | AA-32034 | 10,420–11,150 | 10,785 | Pollies |
| Juniperus rise | 9745 ± 40 | SUERC-2614 | 10,900–11,250 | 11,075 | Ballaugh Curragh 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiverrell, R.C.; Innes, J.B.; Blackford, J.J.; Davey, P.J.; Roberts, D.H.; Rutherford, M.M.; Tomlinson, P.R.; Turner, S.D. Early to Mid-Holocene Tree Immigration and Spread in the Isle of Man: The Roles of Climate and Other Factors. Quaternary 2023, 6, 3. https://doi.org/10.3390/quat6010003
Chiverrell RC, Innes JB, Blackford JJ, Davey PJ, Roberts DH, Rutherford MM, Tomlinson PR, Turner SD. Early to Mid-Holocene Tree Immigration and Spread in the Isle of Man: The Roles of Climate and Other Factors. Quaternary. 2023; 6(1):3. https://doi.org/10.3390/quat6010003
Chicago/Turabian StyleChiverrell, Richard C., James B. Innes, Jeff J. Blackford, Peter J. Davey, David H. Roberts, Mairead M. Rutherford, Philippa R. Tomlinson, and Simon D. Turner. 2023. "Early to Mid-Holocene Tree Immigration and Spread in the Isle of Man: The Roles of Climate and Other Factors" Quaternary 6, no. 1: 3. https://doi.org/10.3390/quat6010003
APA StyleChiverrell, R. C., Innes, J. B., Blackford, J. J., Davey, P. J., Roberts, D. H., Rutherford, M. M., Tomlinson, P. R., & Turner, S. D. (2023). Early to Mid-Holocene Tree Immigration and Spread in the Isle of Man: The Roles of Climate and Other Factors. Quaternary, 6(1), 3. https://doi.org/10.3390/quat6010003