
Co-Circulation of Leishmania Parasites and Phleboviruses in a Population of Sand Flies Collected in the South of Portugal
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
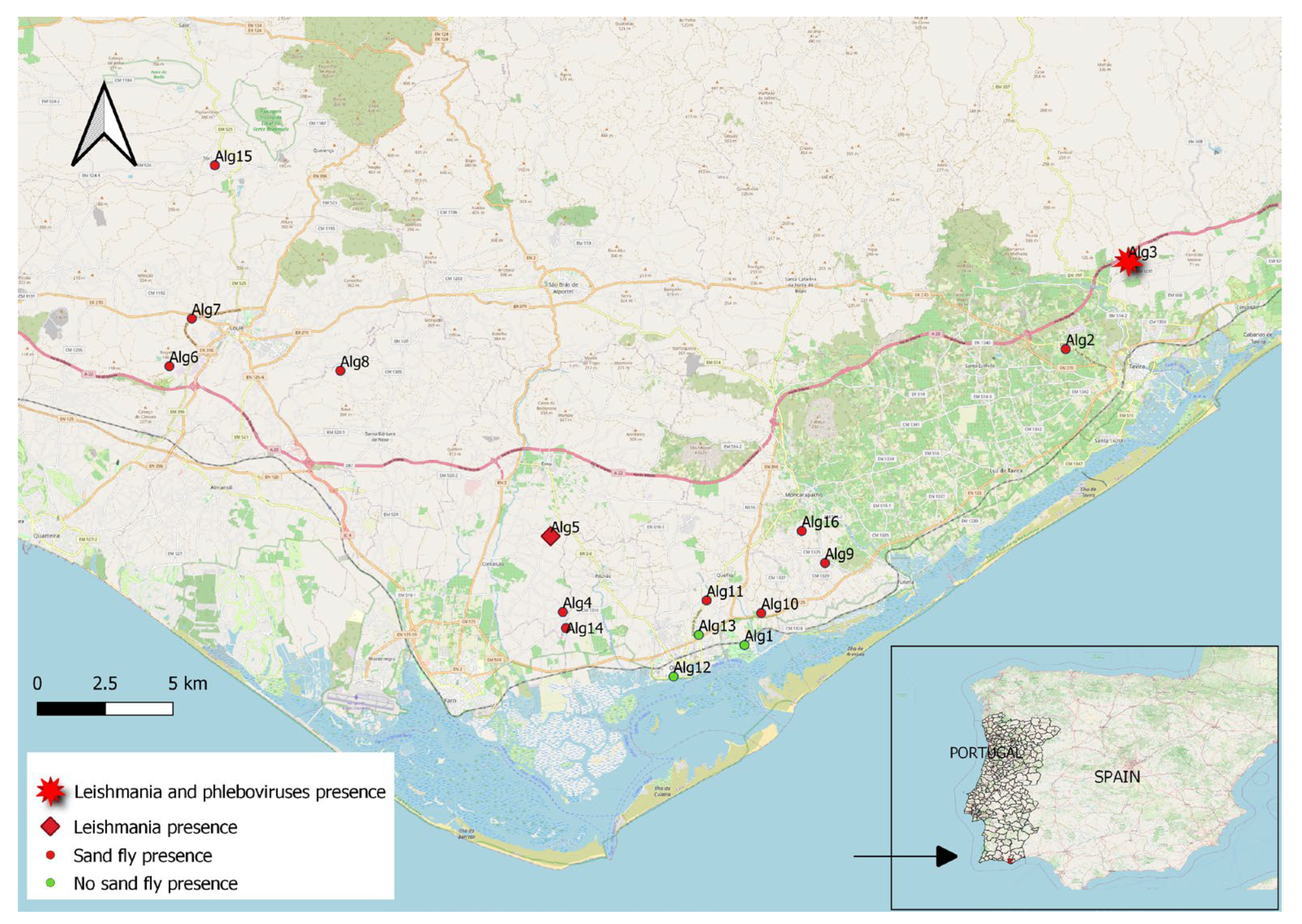
2.1. Field Survey
2.2. Identification of Sand flies
2.3. Phleboviruses and Leishmania spp. Detection and Sequencing
3. Results
3.1. Collected Sand Flies
3.2. Detection of Phleboviruses and Leishmania spp.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Center for Disease Prevention and Control. Phlebotomine Sand Flies. Factsheet for Experts. Available online: https://www.ecdc.europa.eu/en/disease-vectors/facts/phlebotomine-sand-flies (accessed on 28 March 2023).
- Ramírez, J.D.; Hernández, C.; León, C.M.; Ayala, M.S.; Flórez, C.; González, C. Taxonomy, diversity, temporal and geographical distribution of Cutaneous Leishmaniasis in Colombia: A retrospective study. Sci. Rep. 2016, 6, 28266. [Google Scholar] [CrossRef] [PubMed]
- Vickerman, K. The diversity of the kinetoplastid flagellates. In Biology of the Kinetoplastida; Lumsden, W.H.R., Evans, D.A., Eds.; Academic Press: London, UK, 1976; pp. 1–34. [Google Scholar]
- Lainson, R.; Shaw, J.J. Leishmaniasis of the New World: Taxonomic problems. Br. Med. Bull. 1972, 28, 44. [Google Scholar] [CrossRef] [PubMed]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Surveillance, Prevention and Control of Leishmaniases in the European Union and Its Neighbouring Countries; ECDC: Stockholm, Sweden, 2022; p. 60. [Google Scholar]
- Özbilgin, A.; Harman, M.; Karakuş, M.; Bart, A.; Töz, S.; Kurt, Ö.; Çavuş, İ.; Polat, E.; Gündüz, C.; Van Gool, T.; et al. Leishmaniasis in Turkey: Visceral and cutaneous leishmaniasis caused by Leishmania donovani in Turkey. Acta Trop. 2017, 173, 90–96. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28587839 (accessed on 6 December 2023). [CrossRef] [PubMed]
- Antoniou, M.; Haralambous, C.; Mazeris, A.; Pratlong, F.; Dedet, J.P.; Soteriadou, K. Leishmania donovani leishmaniasis in Cyprus. Lancet Infect. Dis. 2008, 8, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Ntais, P.; Sifaki-Pistola, D.; Christodoulou, V.; Messaritakis, I.; Pratlong, F.; Poupalos, G.; Antoniou, M. Leishmaniases in Greece. Am. J. Trop. Med. Hyg. 2013, 89, 906–915. [Google Scholar] [CrossRef]
- Campino, L.; Maia, C. Epidemiology of leishmaniases in Portugal. Acta Med. Port. 2010, 23, 859–864. [Google Scholar]
- Martins, S.; Vilares, A.; Ferreira, I.; Reis, T.; Gargaté, M. Leishmaníase: Confirmação laboratorial de casos clínicos suspeitos de infeção entre 2008 e 2013. Bol. Epi. 2014, 3 (Suppl. S3), 23–25. [Google Scholar]
- Cortes, S.; Maurício, I.L.; Kuhls, K.; Nunes, M.; Lopes, C.; Marcos, M.; Cardoso, L.; Schönian, G.; Campino, L. Genetic diversity evaluation on Portuguese Leishmania infantum strains by multilocus microsatellite typing. Infect. Genet. Evol. 2014, 26, 20–31. [Google Scholar] [CrossRef]
- Remadi, L.; Farjallah, D.; Chargui, N.; Belgacem, S.; Baba, H.; Zrieq, R.; Alzain, M.A.; Babba, H.; Haouas, N. Blood meal analysis and molecular detection of mammalian Leishmania DNA in wild-caught Sergentomyia spp. from Tunisia and Saudi Arabia. Parasitol. Res. 2023, 122, 2181–2191. [Google Scholar] [CrossRef]
- Morales-Yuste, M.; Martín-Sánchez, J.; Corpas-Lopez, V. Canine Leishmaniasis: Update on Epidemiology, Diagnosis, Treatment, and Prevention. Vet. Sci. 2022, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Reguera, R.M.; Morán, M.; Pérez-Pertejo, Y.; García-Estrada, C.; Balaña-Fouce, R. Current status on prevention and treatment of canine leishmaniasis. Vet. Parasitol. 2016, 227, 98–114. [Google Scholar] [CrossRef] [PubMed]
- Bouloy, M. Molecular biology of phleboviruses. In Bunyaviridae Molecular and Cellular Biology; Plyusnin, A., Elliott, R.M., Eds.; Caister Academic Press: Norfolk, UK, 2011; pp. 95–128. [Google Scholar]
- International Committee on Taxonomy of Viruses. Virus Taxonomy: 2022 Release. Email Ratification March 2023 (MSL #38). Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 1 June 2023).
- Charrel, R.N.; Bichaud, L.; de Lamballerie, X. Emergence of Toscana virus in the Mediterranean area. World J. Virol. 2012, 5, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Amaro, F.; Zé-Zé, L.; Alves, M.J. Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting. Viruses 2022, 14, 1768. [Google Scholar] [CrossRef]
- Bartels, S.; de Boni, L.; Kretzschmar, H.A.; Heckmann, J.G. Lethal encephalitis caused by the Toscana virus in an elderly patient. J. Neurol. 2012, 259, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Popescu, C.P.; Cotar, A.I.; Dinu, S.; Zaharia, M.; Tardei, G.; Ceausu, E.; Badescu, D.; Ruta, S.; Ceianu, C.S.; Florescu, S.A. Emergence of Toscana Virus, Romania, 2017–2018. Emerg. Infect. Dis. 2021, 27, 1482–1485. [Google Scholar] [CrossRef]
- Valassina, M.; Valentini, M.; Pugliese, A.; Valensin, P.E.; Cusi, M.G. Serological survey of Toscana virus infections in a high-risk population in Italy. Clin. Diag. Lab. Immunol. 2003, 10, 483–484. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Toscana Virus Infection. Available online: https://www.ecdc.europa.eu/en/toscana-virus-infection (accessed on 28 March 2023).
- Depaquit, J.; Grandadam, M.; Fouque, F.; Andry, P.E.; Peyrefitte, C. Arthropod-borne viruses transmitted by Phlebotomine sandflies in Europe: A review. Euro Surveill. 2010, 15, 19507. [Google Scholar] [CrossRef]
- Ehrnst, A.; Peters, C.J.; Niklasson, B.; Svedmyr, A.; Holmgren, B. Neurovirulent Toscana virus (a sandfly fever virus) in Swedish man after visit to Portugal. Lancet 1985, 1, 1212–1213. [Google Scholar] [CrossRef]
- Amaro, F.; Luz, T.; Parreira, P.; Marchi, A.; Ciufolini, M.G.; Alves, M.J. Serological evidence of Toscana virus infection in Portuguese patients. Epidemiol. Infect. 2012, 140, 1147–1150. [Google Scholar] [CrossRef]
- Amaro, F.; Zé-Zé, L.; Luz, M.T.; Alves, M.J. Toscana Virus: Ten Years of Diagnostics in Portugal. Acta Med. Port. 2021, 34, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Amaro, F.; Hanke, D.; Zé-Zé, L.; Alves, M.J.; Becker, S.C.; Höper, D. Genetic characterization of Arrabida virus, a novel phlebovirus isolated in South Portugal. Virus Res. 2016, 214, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Amaro, F.; Zé-Zé, L.; Alves, M.J.; Börstler, J.; Clos, J.; Lorenzen, S.; Becker, S.C.; Schmidt-Chanasit, J.; Cadar, D. Co-circulation of a novel phlebovirus and Massilia virus in sandflies, Portugal. Vir. J. 2015, 12, 174. [Google Scholar] [CrossRef]
- Filipe, A.R. Serological survey for antibodies to arboviruses in the human population of Portugal. Trans. R. Soc. Trop. Med. Hyg. 1974, 68, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.B.; Gouveia, C.; Zé-Zé, L.; Amaro, F.; Ferreira, G.C.; Brito, M.J. Prolonged Febrile Illness Caused by Sicilian Virus Infection in Portugal. In Proceedings of the 36th Annual Meeting of the European Society for Paediatric Infectious Diseases, Malmo, Sweden, 28 May–2 June 2018. [Google Scholar]
- Maia, C.; Ayhan, N.; Cristovão, J.M.; Pereira, A.; Charrel, R. Human seroprevalence of Toscana virus and Sicilian phlebovirus in the southwest of Portugal. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Ayhan, N.; Cristovão, J.M.; Vilhena, H.; Martins, A.; Cachola, P.; Henriques, J.; Coimbra, M.; Catarino, A.; Lestinova, T.; et al. Antibody response to Toscana Virus and Sandfly Fever Sicilian Virus in cats naturally exposed to phlebotomine Sand Fly bites in Portugal. Microorganisms 2019, 7, 339. [Google Scholar] [CrossRef]
- Alwassouf, S.; Maia, C.; Ayhan, N.; Coimbra, M.; Cristovão, J.M.; Richet, H.; Bichaud, L.; Campino, L.; Charrel, R.N. Neutralization-based seroprevalence of Toscana virus and sandfly fever Sicilian virus in dogs and cats from Portugal. J. Gen. Virol. 2016, 97, 2816–2823. [Google Scholar] [CrossRef]
- Instituto Português do Mar e da Atmosfera. Área Educativa—Clima de Portugal Continental. Available online: https://www.ipma.pt/pt/educativa/tempo.clima/ (accessed on 28 March 2023).
- Maroli, M.; Feliciangeli, M.D.; Arias, J. Metodos de Captura, Conservacion y Montaje de los Flebotomos (Diptera: Psychodidae); Organización Panamericana de la Salud: Washington, DC, USA, 1997; p. 72. [Google Scholar]
- Pires, C. Os Flebótomos (Diptera, Psychodidae) dos Focos Zoonóticos de Leishmanioses em Portugal. Ph.D. Thesis, Instituto de Higiene e Medicina Tropical, Universidade Nova de Lisboa, Lisboa, Portugal, 2000; p. 228. (In Portuguese). [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial Cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Lambert, A.J.; Lanciotti, R.S. Consensus amplification and novel multiplex sequencing method for S segment species identification of 47 viruses of the Orthobunyavirus, Phlebovirus, and Nairovirus genera of the family Bunyaviridae. J. Clin. Microbiol. 2009, 4, 2398–2404. [Google Scholar] [CrossRef]
- Cruz, I.; Cañavate, C.; Rubio, J.M.; Morales, M.A.; Chicharro, C.; Laguna, F.; Jiménez-Mejías, M.; Sirera, G.; Videla, S.; Alvar, J. Spanish HIV-Leishmania Study Group. A nested polymerase chain reaction (Ln-PCR) for diagnosing and monitoring Leishmania infantum infection in patients co-infected with human immunodeficiency virus. Trans. R. Soc. Trop. Med. Hyg. 2002, 96 (Suppl. S1), S185–S189. [Google Scholar] [CrossRef]
- León, C.M.; Muñoz, M.; Hernández, C.; Ayala, M.S.; Flórez, C.; Teherán, A.; Cubides, J.R.; Ramírez, J.D. Analytical performance of four polymerase chain reaction (PCR) and real time PCR (qPCR) assays for the detection of six Leishmania species DNA in Colombia. Front. Microbiol. 2007, 8, 1907. [Google Scholar] [CrossRef] [PubMed]
- Baverstock, P.R.; Illana, S.; Christy, P.E.; Robinson, B.S.; Johnson, A.M. srRNA evolution and phylogenetic relationships of the genus Naegleria (Protista: Rhizopoda). Mol. Biol. Evol. 1989, 6, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Schrey, C.F.; Pires, C.A.; Macvean, D.W. Distribution of phlebotomine sandflies and the rate of their infection with Leishmania promastigotes in the Algarve, Portugal. Med. Vet. Entomol. 1989, 3, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Maia, C.; Dionísio, L.; Afonso, M.O.; Neto, L.; Cristóvão, J.M.; Campino, L. Leishmania infection and host-blood feeding preferences of phlebotomine sandflies and canine leishmaniasis in an endemic European area, the Algarve Region in Portugal. Mem. Inst. Oswaldo Cruz 2013, 108, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Zé-Zé, L.; Amaro, F.; Osório, H.C.; Giovanetti, M.; Lourenço, J.; Alves, M.J. Molecular Identification and Ecology of Portuguese Wild-Caught Phlebotomine Sandfly Specimens. Zoonotic Dis. 2022, 2, 19–31. [Google Scholar] [CrossRef]
- Alves-Pires, C.; Ribeiro, H. The phlebotomine sandflies of Portugal. V—Observations of the ecology of the vectors of leishmaniasis in the Alto-Douro region. Parasitologia 1991, 33, 63–68. [Google Scholar]
- Xu, Z.; Fan, N.; Hou, X.; Wang, J.; Fu, S.; Song, J.; Shi, M.; Liang, G. Isolation and Identification of a Novel Phlebovirus, Hedi Virus, from Sandflies Collected in China. Viruses 2021, 13, 772. [Google Scholar] [CrossRef]
- Charrel, R.N.; Moureau, G.; Temmam, S.; Izri, A.; Marty, P.; Parola, P.; da Rosa, A.T.; Tesh, R.B.; de Lamballerie, X. Massilia virus, a novel Phlebovirus (Bunyaviridae) isolated from sandflies in the Mediterranean. Vector Borne Zoonotic Dis. 2009, 9, 519–530. [Google Scholar] [CrossRef]
- Pires, C.A. Les Phlébotomes du Portugal. I—Infestation naturelle de Phlebotomus ariasi Tonnoir, 1921 et Phlebotomus perniciosus Newstead, 1911, par Leishmania dans le foyer zoonotique de Arrábida (Portugal) [Phlebotomus of Portugal. I-Natural infestation of Phlebotomus ariasi Tonnoir, 1921 and Phlebotomus perniciosus Newstead, 1911, by Leishmania in the zoonotic focus of Arrábida (Portugal)]. Ann. Parasitol. Hum. Comp. 1984, 59, 521–524. [Google Scholar] [PubMed]
- Maia, C.; Afonso, M.O.; Neto, L.; Dionísio, L.; Campino, L. Molecular detection of Leishmania infantum in naturally infected Phlebotomus perniciosus from Algarve region, Portugal. J. Vector Borne Dis. 2009, 46, 268–272. [Google Scholar] [PubMed]
- Branco, S.; Alves-Pires, C.; Maia, C.; Cortes, S.; Cristovão, J.M.; Gonçalves, L.; Campino, L.; Afonso, M.O. Entomological and ecological studies in a new potential zoonotic leishmaniasis focus in Torres Novas municipality, Central Region, Portugal. Acta Trop. 2013, 125, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Fares, W.; Dachraoui, K.; Barhoumi, W.; Cherni, S.; Chelbi, I.; Zhioua, E. Co-circulation of Toscana virus and Leishmania infantum in a focus of zoonotic visceral leishmaniasis from Central Tunisia. Acta Trop. 2020, 204, 105342. [Google Scholar] [CrossRef] [PubMed]
- Ergunay, K.; Kasap, O.E.; Orsten, S.; Oter, K.; Gunay, F.; Yoldar, A.Z.; Dincer, E.; Alten, B.; Ozkul, A. Phlebovirus and Leishmania detection in sandflies from eastern Thrace and Northern Cyprus. Parasit. Vectors. 2014, 7, 575. [Google Scholar] [CrossRef] [PubMed]
- Faucher, B.; Bichaud, L.; Charrel, R.; Mary, C.; Izri, A.; de Lamballerie, X.; Piarroux, R. Presence of sandflies infected with Leishmania infantum and Massilia virus in the Marseille urban area. Clin. Microbiol. Infect. 2014, 20, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Romeo, G.; Callegari, E.; Bonilauri, P.; Chiapponi, C.; Carra, E.; Rugna, G.; Taddei, R.; Lelli, D.; Dottori, M. Co-Circulation of Phleboviruses and Leishmania Parasites in Sand Flies from a Single Site in Italy Monitored between 2017 and 2020. Viruses 2021, 13, 1660. [Google Scholar] [CrossRef]
- Dincer, E.; Gargari, S.; Ozkul, A.; Ergunay, K. Potential animal reservoirs of Toscana virus and coinfections with Leishmania infantum in Turkey. Amer J. Trop. Med. Hyg. 2015, 92, 690–697. [Google Scholar] [CrossRef]
- Rossi, M.; Castiglioni, P.; Hartley, M.A.; Eren, R.O.; Prével, F.; Desponds, C.; Utzschneider, D.T.; Zehn, D.; Cusi, M.G.; Kuhlmann, F.M.; et al. Type I interferons induced by endogenous or exogenous viral infections promote metastasis and relapse of leishmaniasis. Proc. Natl. Acad. Sci. USA 2017, 114, 4987–4992. [Google Scholar] [CrossRef]
- Rath, C.T.; Schnellrath, L.C.; Damaso, C.R.; de Arruda, L.B.; da Costa Vasconcelos, P.F.; Gomes, C.; Laurenti, M.D.; Calegari Silva, T.C.; de Carvalho Vivarini, Á.; Fasel, N.; et al. Amazonian Phlebovirus (Bunyaviridae) potentiates the infection of Leishmania (Leishmania) amazonensis: Role of the PKR/IFN1/IL-10 axis. PLoS Negl. Trop. Dis. 2019, 13, e0007500. [Google Scholar] [CrossRef]
- Heirwegh, E.; MacLean, E.; He, J.; Kamhawi, S.; Sagan, S.M.; Olivier, M. Sandfly Fever Sicilian Virus-Leishmania major co-infection modulates innate inflammatory response favoring myeloid cell infections and skin hyperinflammation. PLoS Negl. Trop. Dis. 2021, 15, e0009638. [Google Scholar] [CrossRef] [PubMed]
- Chelbi, I.; Maghraoui, K.; Zhioua, S.; Cherni, S.; Labidi, I.; Satoskar, A.; Hamilton, J.G.C.; Zhioua, E. Enhanced attraction of sand fly vectors of Leishmania infantum to dogs infected with zoonotic visceral leishmaniasis. PLoS Negl. Trop. Dis. 2021, 15, e0009647, Erratum in PLoS Negl. Trop. Dis. 2021, 15, e0009947. [Google Scholar] [CrossRef]
- Dachraoui, K.; Chelbi, I.; Labidi, I.; Ben Osman, R.; Sayadi, A.; Ben Said, M.; Cherni, S.; Abbas, M.A.S.; Charrel, R.; Zhioua, E. The Role of the Leishmania infantum Infected Dogs as a Potential Reservoir Host for Toscana Virus in a Zoonotic Visceral Leishmaniasis Focus of Northern Tunisia. Viruses 2023, 15, 1012. [Google Scholar] [CrossRef] [PubMed]
- Maia, C.; Alwassouf, S.; Cristóvão, J.M.; Ayhan, N.; Pereira, A.; Charrel, R.N.; Campino, L. Serological association between Leishmania infantum and sand fly fever Sicilian (but not Toscana) virus in sheltered dogs from southern Portugal. Parasit Vectors. 2017, 10, 92. [Google Scholar] [CrossRef]
- Bichaud, L.; Souris, M.; Mary, C.; Ninove, L.; Thirion, L.; Piarroux, R.P.; Piarroux, R.; De Lamballerie, X.; Charrel, R.N. Epidemiologic relationship between Toscana virus infection and Leishmania infantum due to common exposure to Phlebotomus perniciosus sandfly vector. PLoS Negl. Trop. Dis. 2011, 5, e1328. [Google Scholar] [CrossRef]

{kind=link}
| Collection Site | Parish, Municipality/Month | Number of Collected Sand Flies | Total Number of Studied Pools (f/m) | Identified Species |
|---|---|---|---|---|
| Alg1 | Quelfes, Olhão/May | 0 | - | - |
| Alg2 | Santiago, Tavira/May | 13 | 7 (6/1) | Ph. perniciosus (3f; 1m) |
| Alg3 | Santa Maria, Tavira/May, June | 438 | 57 (48/9) | Ph. ariasi (2f; 2m), Ph. perniciosus (25f; 7m) Se. minuta (1f) |
| Alg4 | Conceição, Olhão/June | 50 | 12 (1/11) | Ph. perniciosus (7f) |
| Alg5 | Conceição, Olhão/June | 57 | 12 (9/3) | Ph. perniciosus (5f; 1m) |
| Alg6 | Loulé, Loulé/July | 3 | 2 (0/2) | Ph. perniciosus (1f) |
| Alg7 | São Sebastião, Loulé/July | 78 | 16 (13/3) | Ph. ariasi (1f), Ph. perniciosus (7f; 2m) |
| Alg8 | Santa Bárbara de Nexe, Loulé/July | 3 | 18 (12/6) | Ph. perniciosus (1f) |
| Alg9 | Moncarapacho, Olhão/August | 85 | 16 (10/6) | Ph. perniciosus (3f; 3m), Ph. sergenti (1f) |
| Alg10 | Quelfes, Olhão/August, October | 229 | 25 (17/8) | Ph. perniciosus (10f; 8m) |
| Alg11 | Quelfes, Olhão/August | 15 | 3 (2/1) | Ph. perniciosus (1m) |
| Alg12 | Olhão, Olhão/September | 0 | - | - |
| Alg13 | Quelfes, Olhão/September | 0 | - | - |
| Alg14 | Conceição, Olhão/September | 98 | 8 (6/2) | Ph. perniciosus (2f; 4m) |
| Alg15 | Tôr, Loulé/October | 24 | 8 (6/2) | Ph. perniciosus (3f; 2m), Ph. sergenti (1f) |
| Alg16 | Moncarapacho, Olhão/September, October | 68 | 10 (8/2) | Ph. perniciosus (5f; 1m) |
| Collection Station | Sampling Date in 2018 | Pool Size (f)/#Pool Number | DNA Barcoding | Identified Pathogen/GenBank Accession Number | Reference |
|---|---|---|---|---|---|
| Alg3 | 16 May | 20/#11 | - | New phlebovirus?/ON807199 | [19] |
| Alg3 | 16 May | 1/#19 | Ph. perniciosus | L. donovani complex OR783264 | This work |
| Alg5 | 5 June | 1/#32 | Ph. perniciosus | L. donovani complex OR783265 | This work |
| Alg3 | 5 June | 20/#38 | - | Massilia phlebovirus/ON807200 | [19] |
| Alg3 | 5 June | 1/#45 | Ph. perniciosus | L. donovani complex * OR783266 | This work |
| Alg3 | 7 June | 13/#57 | - | L. donovani complex * | This work |
| Alg3 | 7 June | 1/#61 | Ph. perniciosus | L. donovani complex * | This work |
| Alg5 | 7 June | 1/#86 | Ph. perniciosus | L. donovani complex * | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaro, F.; Vilares, A.; Martins, S.; Reis, T.; Osório, H.C.; Alves, M.J.; Gargaté, M.J. Co-Circulation of Leishmania Parasites and Phleboviruses in a Population of Sand Flies Collected in the South of Portugal. Trop. Med. Infect. Dis. 2024, 9, 3. https://doi.org/10.3390/tropicalmed9010003
Amaro F, Vilares A, Martins S, Reis T, Osório HC, Alves MJ, Gargaté MJ. Co-Circulation of Leishmania Parasites and Phleboviruses in a Population of Sand Flies Collected in the South of Portugal. Tropical Medicine and Infectious Disease. 2024; 9(1):3. https://doi.org/10.3390/tropicalmed9010003
Chicago/Turabian StyleAmaro, Fátima, Anabela Vilares, Susana Martins, Tânia Reis, Hugo Costa Osório, Maria João Alves, and Maria João Gargaté. 2024. "Co-Circulation of Leishmania Parasites and Phleboviruses in a Population of Sand Flies Collected in the South of Portugal" Tropical Medicine and Infectious Disease 9, no. 1: 3. https://doi.org/10.3390/tropicalmed9010003
APA StyleAmaro, F., Vilares, A., Martins, S., Reis, T., Osório, H. C., Alves, M. J., & Gargaté, M. J. (2024). Co-Circulation of Leishmania Parasites and Phleboviruses in a Population of Sand Flies Collected in the South of Portugal. Tropical Medicine and Infectious Disease, 9(1), 3. https://doi.org/10.3390/tropicalmed9010003