Abstract
The objective of this study was to analyze the relationship between the frequency of artemisinin-based combination (ACT) drug resistance molecular markers and clinical forms of P. falciparum malaria and parasitemia. A cross-sectional study was carried out between January and April 2014 at the Operational Clinical Research Unit of Melen in febrile children aged 12 to 240 months with a Plasmodium sp. infection. A total of 3 mL of peripheral blood collected from an EDTA tube was used for leukocyte depletion. DNA mutation detection was performed by next generation sequencing (NGS). A total of 1075 patients were screened for malaria. Among them, 384 had a Plasmodium infection. P. falciparum mono-infection was found in 98.9% of the patients. Pfcrt-326T mutation was found in all isolates, while 37.9% had Pfmdr2-484I mutant allele. The highest median parasite densities were found in patients infected by parasites carrying the CVIET haplotype of the Pfcrt gene. The different genetic profiles found here, and their variations according to clinical and biological signs of severe malaria, are additional arguments for the surveillance of P. falciparum strains.
1. Introduction
Malaria cases were estimated at 247 million in the world in 2021. During this year, the number of malaria deaths was 619,000, of which 80% were children under the age of 5 [1]. Plasmodium (P.) falciparum is responsible for a severe form of the disease. Since the start of the 2000s, the World Health Organization (WHO) has recommended the use of sulfadoxine-pyrimethamine combination for preventive treatment against malaria during pregnancy, and artemisinin-based combinations (ACTs) for the treatment of uncomplicated malaria. The main combinations are artemether-lumefantrine (AL), artesunate-amodiaquine (AS-AQ), dihydroartemisinin piperaquine (DH-PQ), artesunate-mefloquine (AS-MQ) and artesunate-sulfadoxine-pyrimethamine (AS-SP). Severe malaria is treated with injectable artesunate (intramuscular or intravenous), followed by a complete three-day ACT course once the patient can tolerate oral medicines. Artemether and quinine may also be used in the absence of artesunate [2].
These recommendations have been adopted after the spread of P. falciparum strains multiresistant to conventional antimalarials, some of which are partner’s molecules of artemisinin derivatives in ACTs. ACTs led to a significant reduction in malaria morbidity and mortality in populations at risk [1,3]. Thus, WHO recommends regular monitoring of the efficacy of these antimalarial medicines to ensure that the chosen treatments are still efficacious.
In Southeast Asia, the resistance of parasites against artemisinin (ART) derivatives, as well as Piperaquine, has been shown [4]. Resistance to ART and its derivatives is characterized by an extension of the parasite clearance time in treated individuals [4,5]. Indeed, the persistence of parasites after three days of ACT treatment has been observed [5].
Moreover, the genetic background of ART resistant P. falciparum strains has been analyzed through a genome-wide association study [5]. Mutations Y493H, R539T, R561H, I543T and C580Y, located on the P. falciparum klech 13 (Pfk13) gene in resistant parasites have been related to artemisinin resistance [6,7,8,9,10].
Other mutations have been found in other genes, such as P. falciparum ferrodoxine (Pffd- D193Y), P. falciparum apicoplast ribosomal precursor S10 (Pfarps-V127M), P. falciparum multidrug resistance protein 2 (Pfmdr2-T484I) and P. falciparum chloroquine resistance transporter (Pfcrt- I356T and Pfcrt- N326S) [11,12]. These mutations may contribute to the development of the parasite’s resistance to ART [13]. Likewise, the T484I mutation of P. falciparum multidrug resistance protein 2 (Pfmdr2) has been associated with artemisinin drug resistance [10,13]. Such resistance may be related to a higher risk of developing severe malaria linked to a change in the fitness of the P. falciparum strains.
Monitoring the resistance of malaria parasites to partner drugs is also necessary [14]. Two genes, Pfcrt and Pfmdr-1, are associated with resistance to the three main partner drugs: amodiaquine (AQ), lumefantrine and mefloquine. The 74I-75E-76T mutations of the Pfcrt gene and those of the Pfmdr1 gene, N86Y-F184Y-D1246Y, are related to the parasite’s resistance to chloroquine (CQ) and AQ [15,16]. The N86-F184-D1246 haplotype of the Pfmdr1 gene and the mutation detected at the E415G codon of the gene (exo-E415G) coding for exonuclease are associated with a decrease in parasite susceptibility to lumefantrine and piperaquine, respectively [17,18].
In Gabon, the treatment of uncomplicated malaria is based on the administration of AS-AQ and AL as first-line treatment and dihydroartemisinin-piperaquine phosphate (DHA-PQ) as second-line treatment. The injectable form of artesunate or quinine (if artesunate is unavailable) is recommended for the treatment of complicated malaria. Studies performed in different areas of this country reported a high frequency of molecular markers associated with resistance to aminoquinoline antimalarials and antifolates [19,20]. Otherwise, malaria prevalence in Gabon, after a drop between 2005 and 2011, has increased again in patients aged less than 20 years: 25% in 2011 and 36.3% in 2014 [21]. The prevalence of P. falciparum malaria was 32.8% among 1962 patients received in 2021 in the sentinel site for malaria surveillance of Melen (unpublished data). Children under 5 years old and those over 5 years old accounted for 68.9% and 54.5%, respectively (unpublished data). Among those aged over 5 years, 45.4% concerned children aged over 11 years (unpublished data). Likewise, the prevalence of severe malaria increased and a change in the clinical-biological profile was observed, with neurological forms being predominant [22].
Thus, the aim of this study was to analyze the relationship between the frequency of drug resistance molecular markers to ACTs and clinical forms of P. falciparum malaria according to parasite density level.
2. Materials and Methods
2.1. Study Area
A cross-sectional study was conducted between January and April 2014 in the Operational and Clinical Research Unit (OCRU), located in the Regional Hospital of Melen (RHM) in the north of Libreville. The RHM is located 11 km north of Libreville, as previously described [21]. Its equatorial climate is subdivided in four seasons: a short dry season from December to January, a long dry season from May to September, a short rainy season from October to November and a long rainy season from February to June. The malaria transmission is perennial in the country. P. falciparum is the cause of 99% of the symptomatic infections [20]. The major vector species are Anopheles (A.) gambiae and A. funestus [23].
2.2. Study Population
The participants were children aged 12 to 240 months mainly living in the district surrounding the RHM. Children meeting the following criteria were invited to participate: being febrile (with a tympanic temperature of >37.5 °C) or having a 24 h to 48 h history of fever before the day of consultation. A clinical examination was performed to identify the criteria of complicated malaria, looking for repeated convulsions, iterative vomiting and icterus in particular. Complicated malaria was defined and classified according to WHO indications [24]. Biological analyses were also performed for each patient. Indeed, health workers appointed in pediatric wards directed all patients towards the OCRU to benefit from the blood smear and cell blood count tests. Those who had a P. falciparum mono infection and parasite density of >2000 trophozoïtes per microliter of blood (p/µL) were included. Non-inclusion criteria were the presence of a mixed Plasmodium infection and non-falciparum infection.
Approximately 3 mL of blood sample in a EDTA tube were used for the malaria diagnoses.
2.3. Malaria Diagnosis
Giemsa-stained thick and thin blood smears of peripheral blood were taken for each patient, as previously described [25]. A smear was only considered negative if no malaria parasites were seen in 100 microscopic fields using immersion oil. The thin blood smear made it possible to identify Plasmodium species. All patients infected with Plasmodium species were treated according to national recommendations.
2.4. Leukocyte Depletion
Leukocyte depletion consists of the elimination of blood leukocytes before malaria parasite DNA extraction using CF11 columns containing cellulose powder. It reduces the amount of human DNA (contained in the nucleus of leukocytes) in a Plasmodium infected blood sample to obtain P. falciparum purified DNA. Leucocyte depletion was carried out at the Department of Parasitology-Mycology of the Faculty of Medicine. Venous blood sample from each patient was depleted of leucocytes within 6 hours following the sample collection. After the depletion of the leucocytes, a quality control of the sample was done based on microscopy.
2.5. Molecular Analysis
Parasite genomic DNA was then extracted from samples after leucocytes depletion using a commercial Kit QIAamp DNA Blood Midi (100), according to the manufacturer’s instructions. DNA was kept in 100 µL of elution buffer at −80 °C for long-term conservation. The genes analyzed are reported in Table 1.

Table 1.
Molecular markers of antimalarial resistance.
The NGS was carried out at the Wellcome Trust Sanger Institute (United Kingdom) for the detection of mutations related to resistance to ART and quinolines. To characterize ART, the resistance genes designated parasite genetic background (PGB) were sequenced (Table 1). Polymorphism on the Pfcrt (at codons 72–76) and Pfmdr1 (at codons 86, 184 and 1246) genes was also analyzed in order to characterize the genetic profile of Plasmodium strains resistant to quinolines. All single nucleotide polymorphisms (SNPs) were further assessed using the 3D7 strain as a reference genome.
This work was carried out as part of a multicenter study by different teams in 10 African countries for PfK13 gene polymorphism analysis [30]. These teams gathered in the "Plasmodium Diversity Network African (PDNA)" network, aiming to study the genetic diversity of P. falciparum. This includes the study of molecular markers associated with resistance to artemisinin and partner molecules in P. falciparum isolates [26]. This network has established a collaboration with the Wellcome Trust Sanger Institute in order to generate standardized data.
2.5.1. Genetic Report Card User Guide
The Genetic Report Card (GRC) contains malaria parasite genetic data, derived from the analysis of patient blood samples. For each sample, we determined the genotype for several variations (single nucleotide polymorphisms, or SNPs) that are known to be associated with resistance to antimalarials. The report also contains data on Plasmodium species found in the sample. The following is a guide to interpreting the Genetic Report Card data, linking the genotyped mutations (molecular markers) with resistance to different antimalarial drugs (Table A1 and Table A2).
2.5.2. Drug Resistance Genes and Haplotypes
Several columns in the GRC show the genotypes of drug resistance mutations. For consistency with the published literature, these genotypes are reported as amino acid positions and alleles. For genes that carry multiple SNPs of interest, we concatenated the genotypes to form a haplotype, which is also common in the literature. In the haplotypes, we also used the following special characters:
A dash (“-”) indicates a missing genotype, either because the sample could not be genotyped, or because the assay is still under development/refinement. An asterisk (“*”) indicates that two alleles were detected (a heterozygous call) (Table A2).
2.5.3. Sample Barcodes
The sample barcodes are formed by concatenated genotypes at 101 SNPs across the P. falciparum genome. These SNPs are all biallelic, i.e., only two alleles are observed. They were chosen for their usefulness in analyses of relationship between parasites, and are not associated with drug resistance. The full list of SNPs used is given in Appendix A. SNPs within the barcode are represented by the observed nucleotide (A, T, C and G). If the genotype is missing (could not be detected), the symbol “X” is used; the symbol “N” indicates that both alleles were observed (heterozygous call).
2.5.4. Species Co-Infections
We detected the presence of different Plasmodium species by testing an SNP in the parasite mitochondria (PfMIT: 270) that carries different alleles in three different parasite species: P. falciparum, P. vivax and P. knowlesi. This assay has not yet been published but has been validated with quantitative PCR assays for detecting P. vivax and P. falciparum.
2.5.5. Artemisinin Drug Resistance
Parasite genetic background (PGB) mutations showed evidence of a genetic background of mutations that allowed for the emergence of PfK13 mutations [11]. These mutations are: V127M in the Pfarp-s10 (PF3D7_1460900) protein, D193Y in ferredoxin (PF3D7_1318100), N326S and I356T in Pfcrt (PF3D7_0709000) and T484I in Pfmdr2 (PF3D7_1447900). These are displayed in a concatenated haplotype form, with the reference allele (WT) being VDNIT.
2.5.6. Chloroquine Drug Resistance
Chloroquine drug resistance is primarily mediated by mutations in the chloroquine resistance transporter (Pfcrt, PF3D7_0709000) [31]. An accessory mutation at position 86 in the multidrug resistance protein (Pfmdr1, PF3D7_0523000) has been shown to accentuate this resistance phenotype in parasites [32]. The loci in Pfcrt are represented as a 5-amino acid haplotype at positions 72–76, with the wild-type haplotype being CVMNK [29]. The CVIET haplotype is the most widespread resistant haplotype in Asia and Africa, while SVMNT is common in resistant parasites in South America and Oceania. The Pfmdr1 mutation at position 86 is the first in the three amino-acid haplotypes reported for this gene. The WT variant is N, while the Y variant in MDR-1 86 enhances resistance to chloroquine.
2.5.7. Amodiaquine Drug Resistance
Mutations in the multidrug resistance protein (Pfmdr1, PF3D7_0523000) have been associated with parasite response to amodiaquine. There is limited evidence that mutations at positions 86 and 1246 can mediate the response to this drug, and these positions constitute the haplotype reported for this gene [33]. In vitro experiments have shown that haplotypes containing the mutant 86Y have increased IC50’s to chloroquine and amodiaquine.
2.5.8. Mefloquine and Lumefantrine Drug Resistance
Mutations in the multidrug resistance protein (Pfmdr1, PF3D7_0523000) have been associated with parasite response to the drugs mefloquine and lumefantrine [28]. There is limited evidence that variations at positions 86, 184 and 1246 increase susceptibility to these drugs [28].
2.5.9. Piperaquine Drug Resistance
In a recent genome-wide association study (GWAS), an SNP in a putative exonuclease gene (PF3D7_1362500) was associated with ex vivo piperaquine IC50 of parasite isolates from Cambodia [17]. This molecular marker is at position 415, and a G allele was shown to be significantly associated with increased tolerance of piperaquine with respect to the WT allele E.
2.6. Ethical Considerations
In Gabon, the Gabonese Ministry of Health, represented by the Malaria National Control Program, has committed the Department of Parasitology-Mycology and Tropical Medicine of the Faculty of Medicine of Libreville to perform an evaluation of malaria prevalence and antimalarial drug resistance monitoring in sentinel sites throughout the country. In this study, all patients were enrolled in the sentinel site for malaria surveillance in the RHM (Melen). In this context, patients and the guardian or parent of the children included were informed of the activities in this sentinel site and their signed consent was obtained. Children over the age of 12 gave their consent to participate in the study. In addition, in this site, a standardized procedure on the conservation of blood aliquots of febrile patients in the biobank was established. Being in a sentinel site, the participants were treated according to national recommendations.
2.7. Statistical Analysis
Data were analyzed using Epi InfoTM7. All variables were compared using Chi 2 or the Fisher test to compare the exact proportions, and Student’s t-test and analysis of variance (ANOVA) or the Kruskal-Wallis test, as appropriate for continuous variables. p-value < 0.05 was considered statistically significant.
3. Results
3.1. Characteristics of the Study Population
A total of 1074 patients were screened for malaria. Median age of the population was 69 (66–72) months, with a sex ratio of 1.23 (with 55.1% of patients of male sex). Among them, 384 had a Plasmodium infection. P. falciparum mono-infection was found in 98.9% of blood samples. Among those, sixty-one samples were selected for molecular studies (Figure 1).
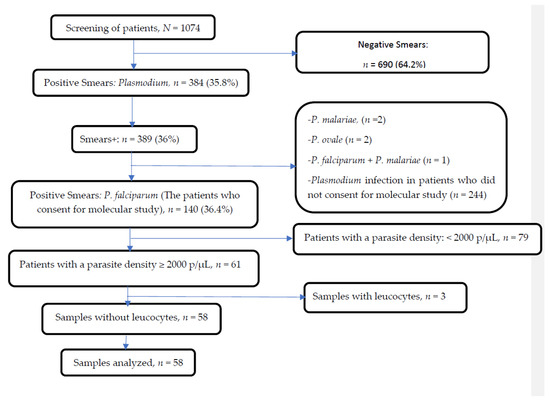
Figure 1.
Profile of outcomes obtained before sequencing.
Amplification and sequencing of the Pfcrt, Pfmdr2, Pfarps10, Pffd and Pfmdr1 genes were successfully performed in 58 samples. These samples were from febrile patients, of whom 70.7% (n = 41/58) had uncomplicated malaria (UM) (Table 2). The median parasitemia was 32,200 (25,900–39,200) p/µL in clinical samples. The highest median parasite density was found in samples from patients with complicated malaria (CM) (p < 0.01) (Table 2). Hyperparasitemia (parasite density ≥200,000 p/µL) was found in six samples from patients with CM. Likewise, diarrhea, vomiting and convulsions were observed in two, three and six patients with CM, respectively. The use of antimalarial drugs before consultation applied to 17.6% of patients with CM and 82.3% of those with UM.
Table 2.
Characteristics of the study population.
3.2. Artemisinin Resistance Related Molecular Markers Prevalence
Analysis of PGB Pfarps10, ferredoxin, Pfcrt and Pfmdr2 genes showed that 37.9% of samples carried the mutant allele Pfmdr2-484I. The Pfcrt-326T mutant allele was found in all samples, while the mutant Pfarps10-127M and Pffd-193Y were not detected. The haplotypes identified were the double mutant VDNTI (27.5%, n = 16/58) and the single mutant VDNTT (62.0%, n = 36/58) (p < 0.01). Six isolates carried both haplotypes.
3.3. Piperaquine, Amodiaquine and Lumefantrine Resistance Related Molecular Markers Prevalence
Exonuclease mutation E415G related to piperaquine resistance was not found in any isolates. Analysis of Pfcrt and Pfmdr1 genes polymorphism showed that the Pfmdr1-86N allele and Pfmdr1-184Y allele were detected in 22 (37.9%) and 3 (5.2%) isolates, respectively. A single (1.72%) isolate had a Pfmdr1-1246Y mutant type allele. Three-quarters of the isolates had mutant alleles Pfcrt-76T, Pfcrt-74I and Pfcrt-75E (Figure 2a). No mutations were found at codon Pfcrt-72 and Pfcrt-73.
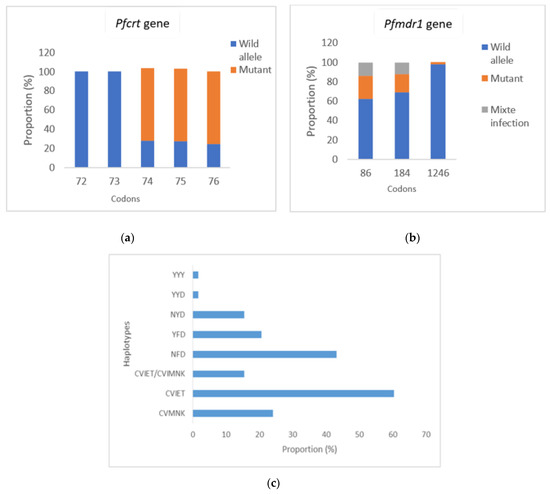
Figure 2.
Frequency of molecular markers of amino-4-quinoleine resistance (Pfcrt-72-76 + Pfmdr1-86-184-1246). (a) Frequency of Pfcrt mutations related to amino-4-quinoleines resistance; (b) Frequency of Pfmdr1 mutations related to AQ and lumefantrine resistance; (c) Frequency of haplotypes of Pfcrt gene, Pfmdr1 gene and Pfcrt + Pfmdr1 genes. YYY: 86Y-184Y-1246Y; YYD: 86Y-184Y-D1246; NYD: N86-184Y-D1246; YFD: 86Y-F184-D1246; NFD: N86-F184-D1246; CVMNK: 72C-73V-74M-75N-76K; CVIET: 72C-73V-74I-75E-76T; CVIET/CVMNK: mixed infection.
The Pfcrt mutant haplotypes CVIET and wild type CVMNK, based on codons 72–76, and the mutant Pfmdr1 haplotypes YFD, NFD and wild type NYD, based on codons 86, 184 and 1246 were the main haplotypes detected (Figure 2c). The triple mutant haplotype CVIET was found in 60.4% of the samples. One sample contained a mixed infection (CVIET/CVMNK) (Figure 2c). The NFD single mutant allele was found more frequently than the double mutant haplotype YFD (43.1% vs. 20.7%) (p = 0.01) and the wild type haplotype NYD (43.2% vs. 15.5%) (p = 0.001).
3.4. Frequency of Haplotypes According to Malaria Clinical Forms
Considering the different haplotypes identified, the double mutant VDNTI haplotype was 1.4-fold (35.3% vs. 24.4%) more frequent in isolates from patients with CM, although the difference was not statistically significant (p = 0.3). Similarly, wild type haplotype NYD was almost twice as frequent in the isolates of these patients (23.5%; n = 4/17 vs. 12%; n = 5/41 in isolates of patients with uncomplicated malaria) (p = 0.4; OR 2.2154 [0.5146–9.5372]).
The haplotype CVMNK was found in the majority of the isolates of patients with uncomplicated malaria. In these, the proportion of the NFD haplotype tends to be more frequent: 58.8% (n = 10/17) vs. 48.7% (n = 20/41) (p = 0.5). Likewise, the double mutant allele YFD was twice as frequent (24.4%; n = 10/41 vs. 11.7%; n = 2/17 in isolates of patients with CM) (p = 0.4; OR 2.4194 (0.4700–12.4547)).
The frequency of the triple mutant haplotype CVIET was comparable between patients with CM (58.8%; n = 10/17) and those with uncomplicated malaria (61%; n = 25/41).
3.5. Haplotypes and Clinical and Biological Signs of Severe Malaria
Patients with hyperparasitemia or hyperparasitemia associated with convulsion were frequently infected by parasites with the CVIET triple mutant haplotype, VDNTT haplotype, NFD and YFD haplotypes of Pfmdr1 gene. The VDNTI haplotype was frequent in isolates from patients with icterus and convulsions.
The combination of PGB and Pfcrt 72-76 haplotypes showed the presence of five haplotypes (H0 to H4) (Table 3). H0 and H2 were the most frequent (Table 3).
Table 3.
Genetic profile of P. falciparum isolates according to median parasite density.
4. Discussion
In the present study, the monitoring of drug resistance molecular markers related to antimalarial drugs used in Gabon revealed mutations T484I (>37.8%) and I356T (100%) on the Pfmdr2 and Pfcrt genes, respectively. In Asia, these are associated with a delayed clearance of parasites in patients treated with AL, DHA-PQ and AS-AQ, respectively [13]. The appearance of these mutations suggests that P. falciparum strains are under drug pressure. Thus, the occurrence of additional mutations on the Pfarps and Pffd genes may be expected, although none were found in these isolates. Similarly, molecular markers such as Exo-E415G SNP related to piperaquine resistance and DH-PQ treatment failure were not detected in the isolates. Nevertheless, P. falciparum strains carrying multiple copies of the Plasmepsin 2 gene have been collected in previous studies carried out in Gabon, Burkina Faso, the Democratic Republic of Congo, Mozambique and Uganda [34]. Taken altogether, these data suggest a delayed appearance of the Exo-E415G mutation, compared to the Plasmepsin 2 gene’s amplification in isolates circulating in areas where these molecules are used. Furthermore, the mutant Pfmdr1-86Y (43%) allele, which is less frequent than the mutant Pfcrt-76T allele (60%) in P. falciparum isolates from Melen was also detected. Its frequency confirms a decreasing frequency of this haplotype since the implementation of ACTs [19,35].
It is notable that multiple mutations are generally required to functionally confer and increase the level of drug resistance. The Pfcrt CVIET triple mutation was found in 60.4% of P. falciparum isolates from Melen. This proportion is similar to those reported in the Congo (71%) and south-east Gabon (70.6%) [36,37]. This haplotype was reported to be more frequently detected in isolates from West Africa (44%) and East Africa (38%), than in those from South Africa (21%) and Central Africa (26%) [38]. Among the Pfmdr1 haplotypes, the NFD haplotype related to lumefantrine resistance was the most prevalent, found in almost one out of two isolates, several years after the introduction of AL and AS-AQ in Gabon. Similar frequencies, varying from 35% to 46%, were found in other areas of Gabon and in Tanzania [20,36]. In contrast, a higher prevalence of the NFD haplotype was found in Senegal (62.26%), Kenya (51%) and Ghana (60%) [38]. The circulation of P. falciparum strains carrying the NFD haplotype is frequently related to pressure due to the use of AL [20,39].
In addition to the use of antimalarial drugs as self-medication, as well as the non-observance of the recommended dosage, other factors, such as the parasite’s genetic background, may lead to an increase in the number of complicated malaria cases in Gabon [22].
In the present study, specific haplotypes varied according to malaria severity, as well as clinical and biological signs. Haplotypes NFD, YFD and CVMNK were more common in isolates from patients with uncomplicated malaria. In contrast, in those from patients with severe malaria, NYD and VDNTI were the most frequent, although these differences were not statistically significant. The CVIET haplotype was common in both groups of patients. The relationship between these haplotypes and the occurrence of severe malaria needs to be confirmed. Specific haplotypes have been identified, but no significant relationship between the clinical forms of malaria—or biological modifications and the frequency of drug resistance molecular markers—has been found. More specifically, patients with hyperparasitemia were infected by parasites with the CVIET and VDNTT haplotypes. The VDNTI haplotype was found in isolates from patients with icterus and convulsion. NFD and YFD were detected in parasites from patients with convulsion. A relationship between NYD and high parasitemia (p = 0.04) has been reported elsewhere [40]. In the current study, the combined haplotypes H2 (VDNTT + CVIET), H3 (VDNTT + CVIET/CVMNK) and H4 (VDNTT + CVIET/CVMNK) were found in patients infected with high parasite densities, suggesting their involvement in the parasite’s ability to multiply.
5. Conclusions
More than one decade after the withdrawal of CQ and the introduction of ACTs in Gabon, monitoring of drug resistance molecular markers remains critical to providing data on the propagation of artemisinin resistant parasites, as done in the present study. Moreover, the different genetic profiles found here and their variations according to the clinical and biological signs of severe malaria are additional arguments for their monitoring, and for an in-depth analysis of their role of the CIVIET haplotype of the Pfcrt gene on the fitness of the parasite.
6. Patents
This section is not mandatory but may be added if there are patents resulting from the work reported in this manuscript.
Author Contributions
Conceptualization, M.K.B.-A.; methodology, D.P.M.-M.; software N.P.M.; validation, N.P.M.; formal analysis, J.V.K.L.; investigation, J.M.N.N., N.P.M., B.M.D. and F.B.B.O.; data curation, J.V.K.L.; writing—original draft preparation, J.M.N.N.; writing—review and editing, J.M.N.N.; supervision, D.P.M.-M.; project administration, M.K.B.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Medical Research Council UK (grant G0600718) and the Wellcome Trust (grant 098051) and carried out by the Plasmodium Diversity Network African (PDNA).
Institutional Review Board Statement
The Gabonese Ministry of Health, but represented by the Malaria National Control Program. The study was conducted according to the guidelines of the Declaration of Helsinki.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the study participants, Plasmodium Diversity Network African (PDNA)/DELGEME which provided the funds to carry out this project, all the authors as well as the team of the Department of Parasitology-Mycology.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
List of barcode SNPs.
Table A1.
List of barcode SNPs.
| Multiplex | Chromosome | Position | Mutation | Multiplex | Chromosome | Position | Mutation |
|---|---|---|---|---|---|---|---|
| W36960 | 2 | 376222 | K1929E | W36962 | 7 | 704373 | 389E |
| W36960 | 2 | 470013 | G75E | W36962 | 7 | 1066698 | G483S |
| W36960 | 3 | 656861 | 129V | W36962 | 7 | 1213486 | S543N |
| W36960 | 4 | 110442 | G285E | W36962 | 8 | 339406 | 1283C |
| W36960 | 4 | 881571 | 1081R | W36962 | 8 | 701557 | 394G |
| W36960 | 5 | 350933 | 1369N | W36962 | 8 | 1313202 | 799F |
| W36960 | 5 | 369740 | 907L | W36962 | 9 | 452690 | 1018I |
| W36960 | 6 | 900278 | P696S | W36962 | 9 | 599655 | E654D |
| W36960 | 7 | 1044052 | 686K | W36962 | 10 | 1383789 | N114H |
| W36960 | 8 | 413067 | 1044V | W36962 | 10 | 1385894 | 815P |
| W36960 | 8 | 1314831 | 1342K | W36962 | 11 | 1006911 | D124E |
| W36960 | 9 | 900277 | 1534E | W36962 | 11 | 1295068 | E405K |
| W36960 | 11 | 1018899 | 1199L | W36962 | 11 | 1802201 | 450S |
| W36960 | 11 | 1815412 | E765Q | W36962 | 12 | 858501 | Q469K |
| W36960 | 13 | 1056452 | 1234D | W36962 | 12 | 1667593 | 2381N |
| W36960 | 13 | 1466422 | N66K | W36962 | 12 | 1934745 | 241L |
| W36960 | 14 | 137622 | 1179V | W36962 | 13 | 159086 | 21R |
| W36960 | 14 | 2164225 | 2830S | W36962 | 13 | 388365 | S1236R |
| W36961 | 1 | 145515 | 294I | W36962 | 13 | 1419519 | Q208R |
| W36961 | 3 | 548178 | R2L | W36962 | 13 | 2161975 | D252V |
| W36961 | 4 | 139051 | K438N | W36962 | 13 | 2573828 | I1153M |
| W36961 | 4 | 286542 | H586N | W36962 | 14 | 438592 | N348T |
| W36961 | 4 | 529500 | 1477Y | W36962 | 14 | 2625887 | M238I |
| W36961 | 4 | 1102392 | E808D | W36962 | 14 | 3126219 | S628F |
| W36961 | 5 | 796714 | 396K | W36963 | 1 | 179347 | 311G |
| W36961 | 7 | 461139 | M361I | W36963 | 1 | 180554 | D714N |
| W36961 | 7 | 619957 | 675R | W36963 | 1 | 283144 | H664D |
| W36961 | 7 | 1256331 | L321F | W36963 | 1 | 535211 | 2521F |
| W36961 | 8 | 417335 | R244K | W36963 | 2 | 839620 | 260L |
| W36961 | 8 | 417335 | R244K | W36963 | 4 | 426436 | D560A |
| W36961 | 10 | 317581 | 311I | W36963 | 4 | 531138 | A992E |
| W36961 | 10 | 336274 | I1677V | W36963 | 4 | 891732 | R4468S |
| W36961 | 11 | 477922 | H147Y | W36963 | 5 | 172801 | E218K |
| W36961 | 11 | 1020397 | G700E | W36963 | 6 | 574938 | I2934L |
| W36961 | 11 | 1294107 | 84A | W36963 | 7 | 635985 | T598A |
| W36961 | 11 | 1935227 | R73S | W36963 | 7 | 1308383 | G1945R |
| W36961 | 12 | 1663492 | 1014E | W36963 | 7 | 1358910 | K286E |
| W36961 | 12 | 2171901 | V140D | W36963 | 7 | 1359218 | K388N |
| W36961 | 13 | 1233218 | N277S | W36963 | 8 | 150033 | 1315I |
| W36961 | 13 | 1867630 | M4911I | W36963 | 8 | 399774 | 421K |
| W36961 | 13 | 2377887 | 2002S | W36963 | 8 | 1056829 | L474I |
| W36961 | 14 | 2355751 | H1589Q | W36963 | 9 | 1379145 | R398Q |
| W36961 | 14 | 3046108 | 417V | W36963 | 10 | 1386850 | 927K |
| W36962 | 2 | 529709 | F487L | W36963 | 11 | 408668 | 1058I |
| W36962 | 2 | 714480 | D258G | W36963 | 11 | 828596 | K240E |
| W36962 | 3 | 155697 | 150P | W36963 | 11 | 1935031 | I139L |
| W36962 | 4 | 648101 | 51V | W36963 | 12 | 857245 | E50G |
| W36962 | 4 | 1037656 | 2776I | W36963 | 14 | 107014 | 215K |
| W36962 | 5 | 1204155 | 1338I | W36963 | 14 | 1757603 | D1365G |
| W36962 | 6 | 1282691 | 803K | W36963 | 14 | 2733656 | 557C |
| W36962 | 6 | 1289212 | 125T |
Table A2.
Data base after sequencing.
Table A2.
Data base after sequencing.
| ID | Species | CRT | MDR1 | PGB | EXO |
|---|---|---|---|---|---|
| QW0009-CW | Pf | CVMNK | NYD | VDNTI | E |
| QW0010-CW | Pf | CVIET | YYY | VDNTT | E |
| QW0010-CxW | Pf | CVIET | YFD | VDNTT | E |
| QW0014-CW | Pf | CVMNK | NYD | VDNTI | E |
| QW0016-CW | Pf | CVMNK | NYD | VDNTI | E |
| QW0018-CW | Pf | CVIET,CVMNK | **D | VDNT* | E |
| QW0019-CW | Pf | CVIET,CVMNK | NYD | VDNT* | E |
| QW0020-CW | Pf | CVMNK | NFD | VDNTI | E |
| QW0021-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0022-CW | Pf | CVIET | NYD | VDNTT | E |
| QW0023-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0025-CW | Pf | CVIET | YFD | VDNTT | E |
| QW0027-CW | Pf | CVIET | YYD | VDNTT | E |
| QW0031-CW | Pf | CVIET,CVMNK | YFD | VDNT* | E |
| QW0033-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0035-CW | Pf | CVIET | **D | VDNTT | E |
| QW0038-CW | Pf | CVMNK | YFD | VDNTI | E |
| QW0041-CW | Pf | CVIET | **D | VDNTT | E |
| QW0043-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0046-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0049-CW | Pf | CVIET | YFD | VDNTT | E |
| QW0050-CW | Pf | CVIET | YFD | VDNTT | E |
| QW0057-CW | Pf | CVMNK | NYD | VDNTI | E |
| QW0058-CW | Pf | CVIET | NFD | VDNTI | E |
| QW0064-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0067-CW | Pf | CVMNK | NFD | VDNTI | E |
| QW0071-CW | Pf | CVIET,CVMNK | *FD | VDNT* | E |
| QW0072-CW | Pf | CVIET | N*D | VDNTI | E |
| QW0074-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0076-CxW | Pf | CVIET | NFD | VDNTT | E |
| QW0077-CW | - | - | --- | ----- | - |
| QW0077-CxW | Pf | CVIET | NFD | VDNTT | E |
| QW0081-CW | Pf | CVIET | N*D | VDNTT | E |
| QW0083-CxW | Pf | CVIET | NFD | VDNTT | E |
| QW0084-CxW | Pf | CVIET | YFD | VDNTT | E |
| QW0087-CW | Pf | CVIET | **D | VDNTT | E |
| QW0088-CW | Pf | CVMNK | YFD | VDNTI | E |
| QW0089-CxW | Pf | CVIET | NFD | VDNTT | E |
| QW0090-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0090-CxW | Pf | CVIET | NFD | VDNTT | E |
| QW0091-CxW | Pf | CVIET,CVMNK | **D | VDNTT | E |
| QW0092-CxW | Pf | CVIET | NFD | VDNTT | E |
| QW0096-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0100-CW | Pf | CVMNK | NFD | VDNTI | E |
| QW0101-CW | Pf | CVIET | YFD | VDNTT | E |
| QW0107-CW | Pf | CVMNK | NYD | VDNTI | E |
| QW0109-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0115-CW | Pf | CVMNK | NFD | VDNTI | E |
| QW0118-CW | Pf | CVMNK | YFD | VDNTI | E |
| QW0119-CxW | Pf | CVIET | NYD | VDNTT | E |
| QW0120-CxW | Pf | CVMNK,CVIET | NYD | VDNT* | E |
| QW0125-CxW | Pf | CVIET | YFD | VDNTT | E |
| QW0127-CW | Pf | CVIET,CVMNK | *FD | VDNTT | E |
| QW0127-CxW | Pf | CVIET,CVMNK | YFD | VDNTT | E |
| QW0129-CW | Pf | CVIET | NFD | VDNTT | E |
| QW0139-CxW | Pf | CVIET | *FD | VDNTT | E |
| QW0144-CxW | Pf | CVMNK | NFD | VDNTI | E |
| QW0146-CW | Pf | CVMNK | NFD | VDNTI | E |
| QW0148-CW | Pf | CVMNK,CVIET | NFD | VDNT* | E |
References
- WHO. World Malaria Report 2022; WHO: Geneva, Switzerland, 2022; ISBN 9789240064898.
- Lalloo, D.G.; Shingadia, D.; Bell, D.J.; Beeching, N.J.; Whitty, C.J.M.; Chiodini, P.L. UK Malaria Treatment Guidelines 2016. J. Infect. 2016, 72, 635–649. [Google Scholar] [CrossRef] [PubMed]
- WHO. Word Malaria Report 2021; WHO: Geneva, Switzerland, 2021; ISBN 9789240040496.
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin Resistance in Plasmodium Falciparum Malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef]
- Fairhurst, R.M.; Dondorp, A.M.; Medicine, M.T.; Kingdom, U. Artemisinin-Resistant Plasmodium falciparum Malaria. Microbiol. Spectr. 2016, 4, 1–25. [Google Scholar] [CrossRef]
- Duru, V.; Khim, N.; Leang, R.; Kim, S.; Domergue, A.; Kloeung, N.; Ke, S.; Chy, S.; Eam, R.; Khean, C.; et al. Plasmodium Falciparum Dihydroartemisinin-Piperaquine Failures in Cambodia Are Associated with Mutant K13 Parasites Presenting High Survival Rates in Novel Piperaquine in Vitro Assays: Retrospective and Prospective Investigations. BMC Med. 2015, 13, 305. [Google Scholar] [CrossRef]
- Plasmodium, M. Genomic Epidemiology of Artemisinin Resistant Malaria. Epidemiol. Glob. Health Microbiol. Infect. Dis. 2016. [CrossRef]
- Straimer, J.; Gnädig, N.F.; Witkowski, B.; Amaratunga, C.; Duru, V.; Ramadani, A.P.; Dacheux, M.; Khim, N.; Zhang, L.; Lam, S.; et al. K13-Propeller Mutations Confer Artemisinin Resistance in Plasmodium Falciparum Clinical Isolates. Science 2015, 347, 428–431. [Google Scholar] [CrossRef]
- Ariey, F.; Witkowski, B.; Amaratunga, C.; Beghain, J.; Ma, L.; Lim, P.; Leang, R.; Duong, S.; Sreng, S. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 2016, 505, 50–55. [Google Scholar] [CrossRef]
- Miotto, O.; Amato, R.; Ashley, E.A.; Macinnis, B.; Dhorda, M.; Imwong, M.; Woodrow, C.; Manske, M.; Stalker, J. Genetic architecture of artemisinin-resistant Plasmodium falciparum. Nat. Genet. 2015, 47, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Nyunt, M.H.; Wang, B.; Aye, K.M.; Aye, K.H.; Han, J.H.; Lee, S.K.; Han, K.T.; Htut, Y.; Han, E.T. Molecular Surveillance of Artemisinin Resistance Falciparum Malaria among Migrant Goldmine Workers in Myanmar. Malar. J. 2017, 16, 97. [Google Scholar] [CrossRef]
- Nyunt, M.H.; Soe, M.T.; Myint, H.W.; Oo, H.W.; Aye, M.M.; Han, S.S.; Zaw, N.N.; Cho, C.; Aung, P.Z.; Kyaw, K.T.; et al. Clinical and Molecular Surveillance of Artemisinin Resistant Falciparum Malaria in Myanmar (2009–2013). Malar. J. 2017, 16, 333. [Google Scholar] [CrossRef]
- Otienoburu, S.D.; Suay, I.; Garcia, S.; Thomas, N.V.; Srisutham, S.; Björkman, A.; Humphreys, G.S. An Online Mapping Database of Molecular Markers of Drug Resistance in Plasmodium Falciparum: The ACT Partner Drug Molecular Surveyor. Malar. J. 2019, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Wootton, J.C.; Feng, X.; Ferdig, M.T.; Cooper, R.A.; Mu, J.; Baruch, D.I.; Magill, A.J.; Su, X.Z. Genetic Diversity and Chloroquine Selective Sweeps in Plasmodium Falciparum. Nature 2002, 418, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.P.; Stein, W.D.; Lanzer, M. Is PfCRT a Channel or a Carrier? Two Competing Models Explaining Chloroquine Resistance in Plasmodium Falciparum. Trends Parasitol. 2007, 23, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Amato, R.; Lim, P.; Miotto, O.; Neal, A.T.; Sreng, S.; Suon, S.; Drury, E. Europe PMC Funders Group Genetic Markers Associated with Dihydroartemisinin—Piperaquine Failure in Plasmodium Falciparum Malaria in Cambodia: A Genotype-Phenotype Association Study. Lancet Infect. Dis. 2017, 17, 164–173. [Google Scholar] [CrossRef]
- Otienoburu, S.D.; Ascofaré, O.M.; Schramm, B.; Jullien, V.; Jones, J.J.; Zolia, Y.M.; Houzé, P.; Ashley, E.A.; Kiechel, J.R.; Guérin, P.J. Selection of Plasmodium Falciparum Pfcrt and Pfmdr1 Polymorphisms after Treatment with Artesunate—Amodiaquine Fixed Dose Combination or Artemether—Lumefantrine in Liberia. Malar. J. 2016, 15, 452. [Google Scholar] [CrossRef]
- Frank, M.; Lehners, N.; Mayengue, P.I.; Gabor, J.; Dal-bianco, M.; Kombila, D.U.; Ngoma, G.M.; Supan, C.; Lell, B.; Ntoumi, F.; et al. A Thirteen-Year Analysis of Plasmodium Falciparum Populations Reveals High Conservation of the Mutant Pfcrt Haplotype despite the Withdrawal of Chloroquine from National Treatment Guidelines in Gabon. Malar. J. 2011, 10, 304. [Google Scholar] [CrossRef]
- Nzondo, S.M.; Kouna, L.C.; Mourembou, G.; Boundenga, L.; Karl, R.; Limoukou, I.; Matsiegui, P.B.; Zoleko, R.M.; Mbatchi, B.; Raoult, D.; et al. Malaria in Urban, Semi-Urban and Rural Areas of Southern of Gabon: Comparison of the Pfmdr 1 and Pfcrt Genotypes from Symptomatic Children. Malar. J. 2016, 15, 420. [Google Scholar] [CrossRef]
- Mawili-mboumba, D.P.; Akotet, M.K.B.; Kendjo, E.; Nzamba, J.; Medang, M.O. Increase in Malaria Prevalence and Age of at Risk Population in Different Areas of Gabon. Malar. J. 2013, 12, 3. [Google Scholar] [CrossRef]
- Ndong Ngomo, M.J.; Abang Ekouaghe, P.L.; M’boundoukowe, N.P.; Koumba Lengongo, J.V.; Mawili-Mboumba, D.P.; Bouyou-Akotet, M.K. Profil Clinico-Biologique Du Paludisme Grave Dans Un Contexte de Transition Épidémiologique, Congress of the Ivorian Society of Parasitology and Mycology, 2nd ed.; Université Félix Houphouet-Boigny: Abidjan, Ivory Coast, 2016; p. 39. [Google Scholar]
- Mourou, J.R.; Coffinet, T.; Jarjaval, F.; Pradines, B.; Amalvict, R.; Rogier, C.; Kombila, M.; Pagès, F. Malaria Transmission and Insecticide Resistance of Anopheles Gambiae in Libreville and Port-Gentil, Gabon. Malar. J. 2010, 9, 321. [Google Scholar] [CrossRef]
- WHO World Malaria Report 2005 (WHO/HTM/MAL/2005.1102). 2005. Available online: https://www.rbm.who.int/wmr2005/html/1----2.htm (accessed on 16 January 2023).
- Planche, T.; Krishna, S.; Kombila, M.; Engel, K.; Faucher, J.F.; Kremsner, P.G.; Kingdom, U.; Hospital, A.S.; Humanparasitologie, S. Comparison of Methods for the Rapid Laboratory Assessment of Children with Malaria. Am. J. Trop. Med. Hyg. 2001, 65, 599–602. [Google Scholar] [CrossRef]
- Chenet, S.M.; Okoth, S.A.; Kelley, J.; Lucchi, N.; Huber, C.S.; Vreden, S.; Macedo De Oliveira, A.; Barnwell, J.W.; Udhayakumar, V.; Adhin, M.R. Molecular Profile of Malaria Drug Resistance Markers of Plasmodium Falciparum in Suriname. Antimicrob. Agents Chemother. 2017, 61, 1–8. [Google Scholar] [CrossRef]
- Price, R.N.; Cassar, C.; Brockman, A.; Duraisingh, M.; White, N.J.; Nosten, F.; Krishna, S.; Al, P.E.T.; Hemother, A.N.A.G.C. The Pfmdr1 Gene Is Associated with a Multidrug-Resistant Phenotype in Plasmodium Falciparum from the Western Border of Thailand. Antimicrob. Agents Chemother. 1999, 43, 2943–2949. [Google Scholar] [CrossRef] [PubMed]
- Veiga, M.I.; Dhingra, S.K.; Henrich, P.P.; Straimer, J.; Gna, N.; Uhlemann, A.; Martin, R.E.; Lehane, A.M.; Fidock, D.A. Globally Prevalent PfMDR1 Mutations Modulate Plasmodium falciparum susceptibility to artemisinin-based combination therapies. Nat. Commun. 2016, 7, 11553. [Google Scholar] [CrossRef] [PubMed]
- Fidock, D.A.; Nomura, T.; Talley, A.K.; Cooper, R.A.; Dzekunov, S.M.; Ferdig, M.T.; Ursos, L.M.B.; Sidhu, S.; Deitsch, K.W.; Su, X.; et al. Mutations in the P. falciparum Digestive Vacuole Transmembrane Protein PfCRT and Evidence for Their Role in Chloroquine Resistance. Mol. Cell 2000, 6, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Kamau, E.; Campino, S.; Amenga-etego, L.; Drury, E.; Ishengoma, D.; Johnson, K.; Mumba, D.; Kekre, M.; Yavo, W.; Mead, D.; et al. K13-Propeller Polymorphisms in Plasmodium Falciparum Parasites from Sub-Saharan Africa. J. Infect. Dis. 2015, 211, 1352–1355. [Google Scholar] [CrossRef] [PubMed]
- Wellems, T.E.; Plowe, C. V Chloroquine-Resistant Malaria; Nature Publishing Group: Berlin, Germany, 1990; pp. 770–776. [Google Scholar]
- Venkatesan, M.; Gadalla, N.B.; Stepniewska, K.; Dahal, P.; Nsanzabana, C.; Moriera, C.; Price, R.N.; Rosenthal, P.J.; Dorsey, G.; Sutherland, C.J.; et al. Polymorphisms in Plasmodium Falciparum Chloroquine Resistance Transporter and Multidrug Resistance 1 Genes: Parasite Risk Factors That Affect Treatment Outcomes for P. falciparum Malaria after Artemether-Lumefantrine and Artesunate-Amodiaquine. Am. J. Trop. Med. Hyg. 2014, 91, 833–843. [Google Scholar] [CrossRef]
- Malmberg, M.; Ferreira, P.E.; Tarning, J.; Ursing, J.; Ngasala, B.; Björkman, A.; Mårtensson, A.; Gil, J.P. Plasmodium Falciparum Drug Resistance Phenotype as Assessed by Patient Antimalarial Drug Levels and Its Association with Pfmdr1 Polymorphisms. J. Infect. Dis. 2013, 207, 842–847. [Google Scholar] [CrossRef]
- Leroy, D.; Macintyre, F.; Adoke, Y.; Ouoba, S.; Barry, A.; Mombo-Ngoma, G.; Ndong Ngomo, J.M.; Varo, R.; Dossou, Y.; Tshefu, A.K.; et al. African Isolates Show a High Proportion of Multiple Copies of the Plasmodium Falciparum Plasmepsin-2 Gene, a Piperaquine Resistance Marker. Malar. J. 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Mawili-Mboumba, D.P.; Ngomo, J.M.N.; Maboko, F.; Guiyedi, V.; Mbina, J.R.M.; Kombila, M.; Akotet, M.K.B. Pfcrt 76T and Pfmdr1 86Y Allele Frequency in Plasmodium Falciparum Isolates and Use of Self-Medication in a Rural Area of Gabon. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 729–734. [Google Scholar] [CrossRef]
- Zhou, R.; Zhang, H.; Yang, C.; Liu, Y.; Zhao, Y.; Li, S.; Qian, D.; Xu, B. Molecular Mutation Profile of Pfcrt in Plasmodium Falciparum Isolates Imported from Africa in Henan Province. Malar. J. 2016, 15, 265. [Google Scholar] [CrossRef]
- Ishengoma, D.S.; Mandara, C.I.; Francis, F.; Talundzic, E.; Lucchi, N.W.; Ngasala, B.; Kabanywanyi, A.M.; Mahende, M.K.; Kamugisha, E.; Kavishe, R.A.; et al. Efficacy and Safety of Artemether-Lumefantrine for the Treatment of Uncomplicated Malaria and Prevalence of Pfk13 and Pfmdr1 Polymorphisms after a Decade of Using Artemisinin—Based Combination Therapy in Mainland Tanzania. Malar. J. 2019, 18, 88. [Google Scholar] [CrossRef] [PubMed]
- Voumbo-Matoumona, D.F.; Kouna, L.C.; Madamet, M.; Maghendji-Nzondo, S.; Pradines, B.; Lekana-Douki, J.B. Prevalence of Plasmodium Falciparum Antimalarial Drug Resistance Genes in Southeastern Gabon from 2011 to 2014. Infect. Drug Resist. 2018, 11, 1329–1338. [Google Scholar] [CrossRef]
- Koussounda, F.K.; Jeyaraj, S.; Nguetse, C.N.; Nkonganyi, C.N.; Kokou, K.C.; Beka, M.K.E.; Ntoumi, F. Molecular Surveillance of Plasmodium Falciparum Drug Resistance in the Republic of Congo: Four and Nine Years after the Introduction of Artemisinin - Based Combination Therapy. Malar. J. 2017, 16, 155. [Google Scholar] [CrossRef] [PubMed]
- Mbaye, A.; Dieye, B.; Ndiaye, Y.D.; Bei, A.K.; Muna, A.; Deme, A.B.; Yade, M.S.; Diongue, K.; Gaye, A.; Ndiaye, I.M.; et al. Selection of N86F184D1246 Haplotype of Pfmrd1 Gene by Artemether—Lumefantrine Drug Pressure on Plasmodium Falciparum Populations in Senegal. Malar. J. 2016, 15, 433. [Google Scholar] [CrossRef]
- Nguetse, C.N.; Adegnika, A.A.; Agbenyega, T.; Ogutu, B.R.; Krishna, S.; Kremsner, P.G.; Velavan, T.P. Molecular Markers of Anti-Malarial Drug Resistance in Central, West and East African Children with Severe Malaria. Malar. J. 2017, 16, 217. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).