
Dichichthyidae, a New Family of Deepwater Sharks (Carcharhiniformes) from the Indo–West Pacific, with Description of a New Species


Abstract
1. Introduction
2. Materials and Methods
2.1. Morphology
2.2. Meristics
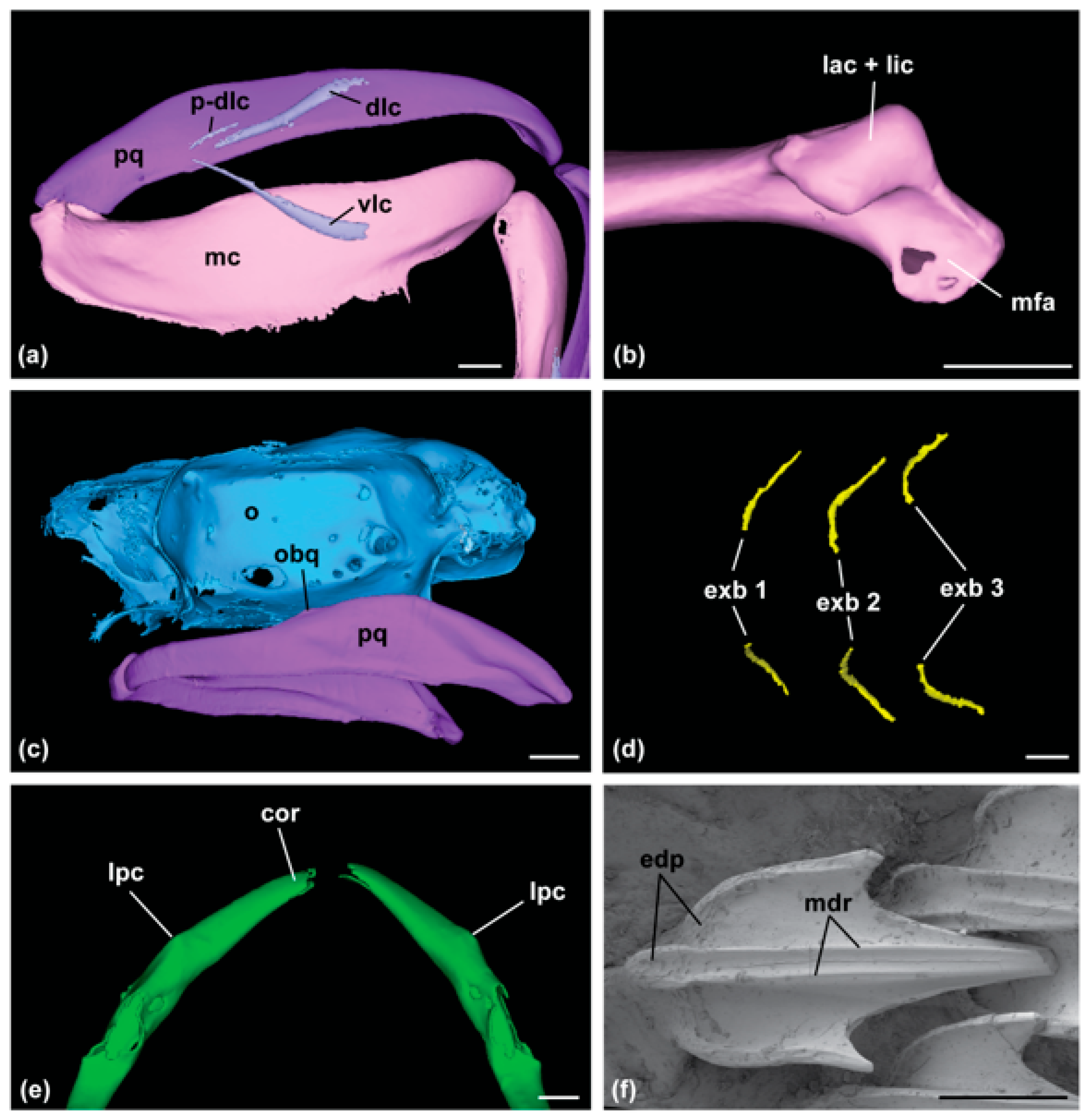
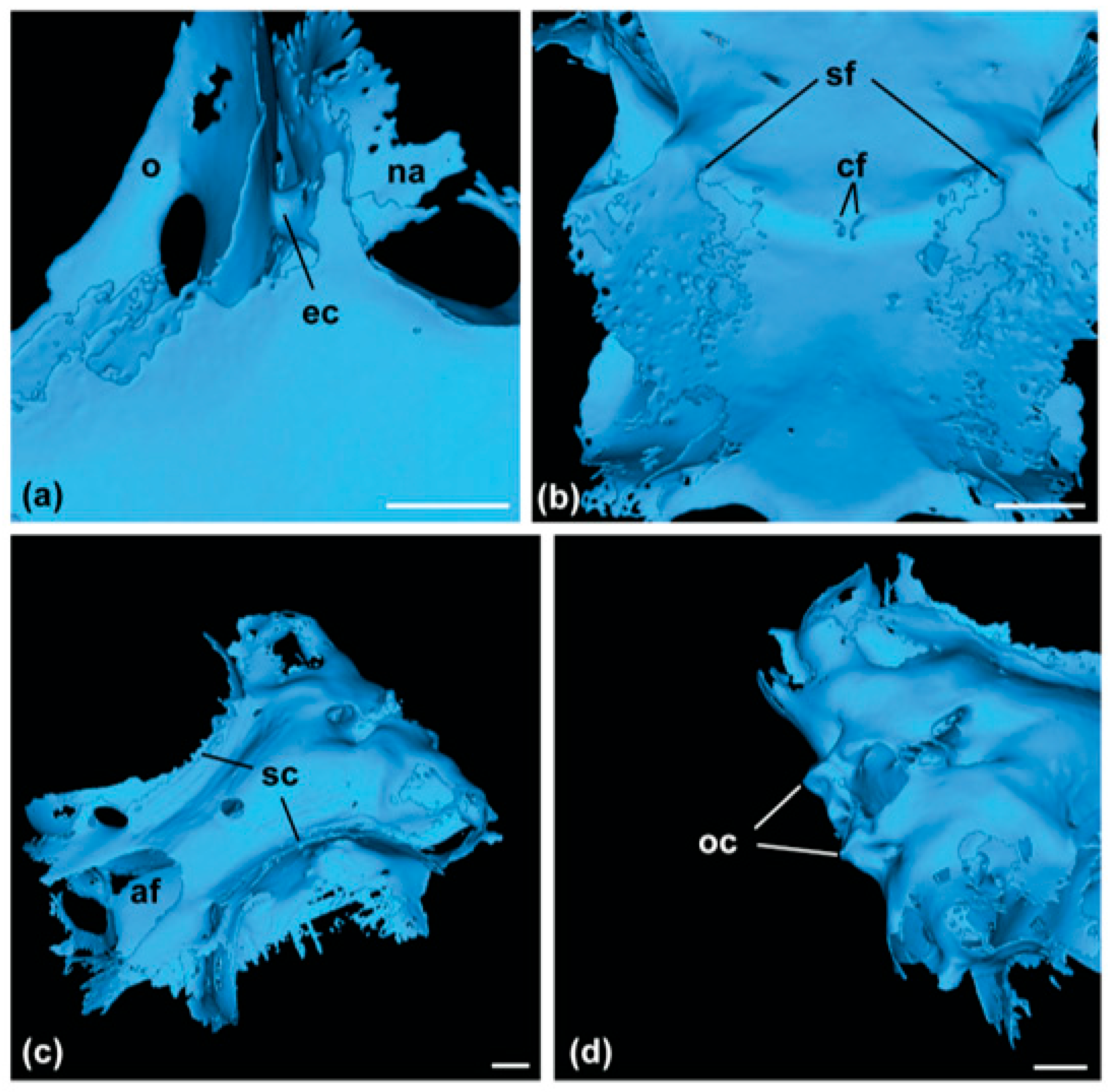
2.3. Skeletal Features
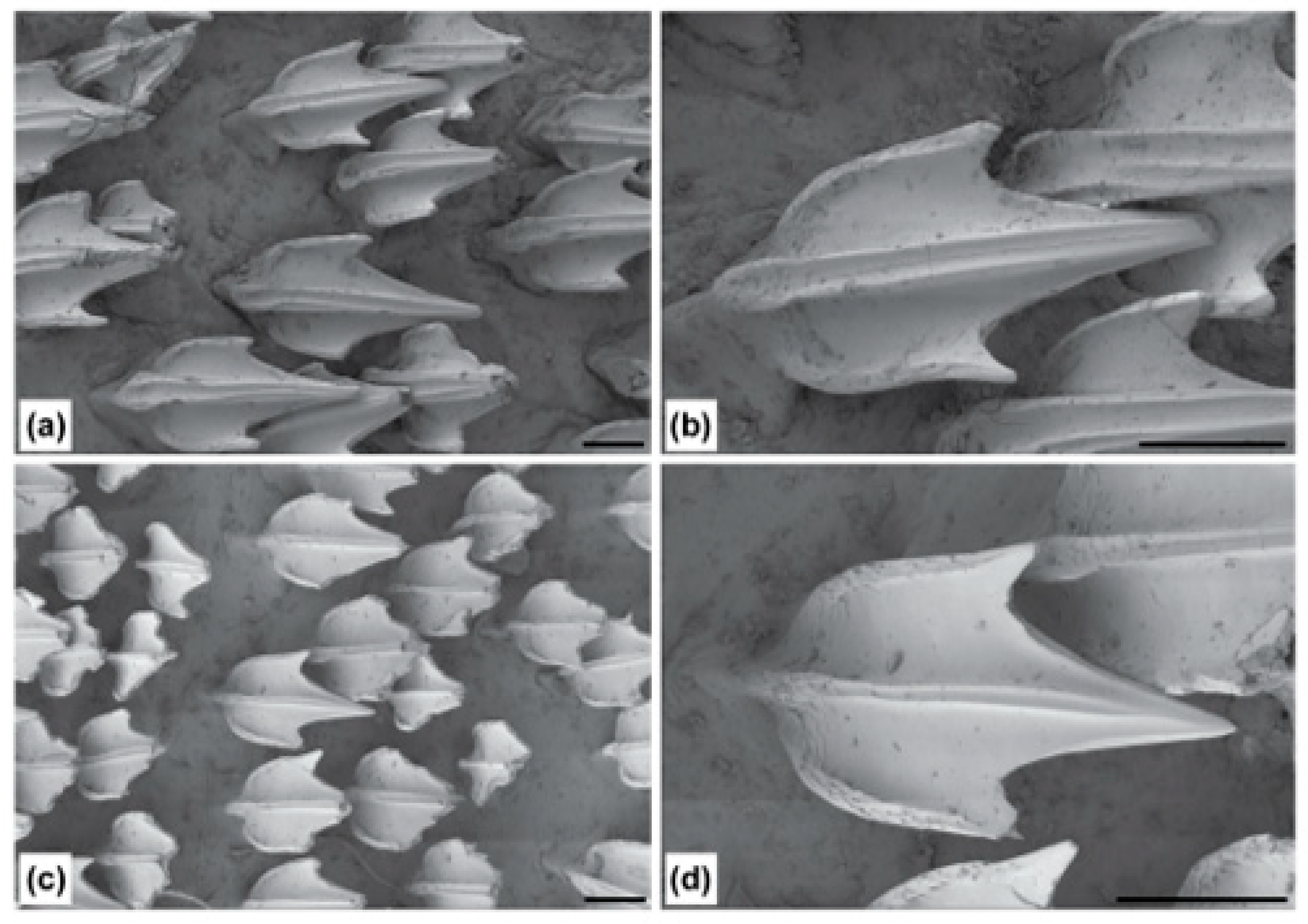
2.4. Denticle Morphology
2.5. Molecular Analyses
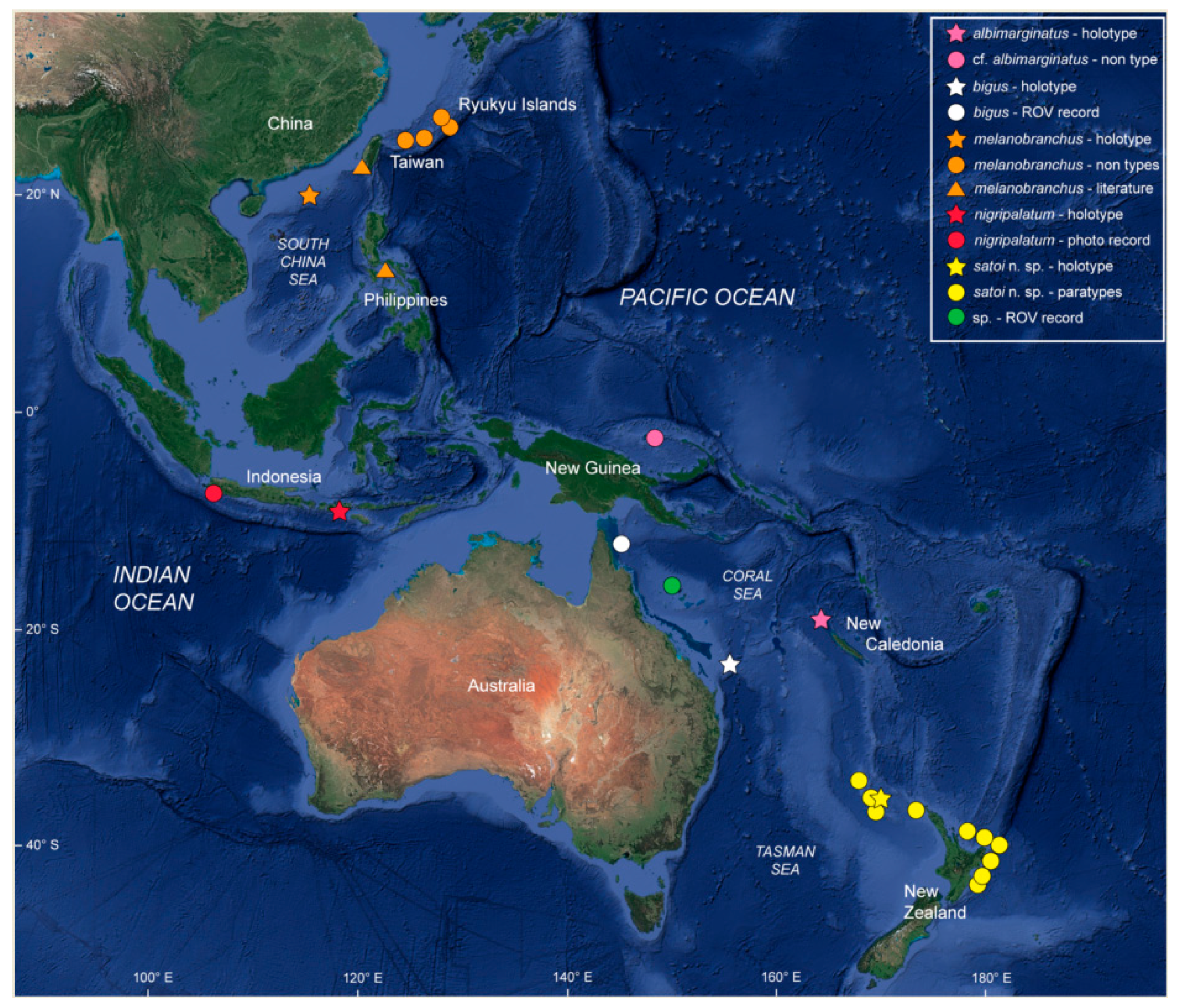
2.6. Distribution
2.7. Institutional Acronyms
3. Results
3.1. Family Dichichthyidae n. fam.
3.1.1. Definition
3.1.2. Genera
3.1.3. Distribution
3.1.4. Etymology
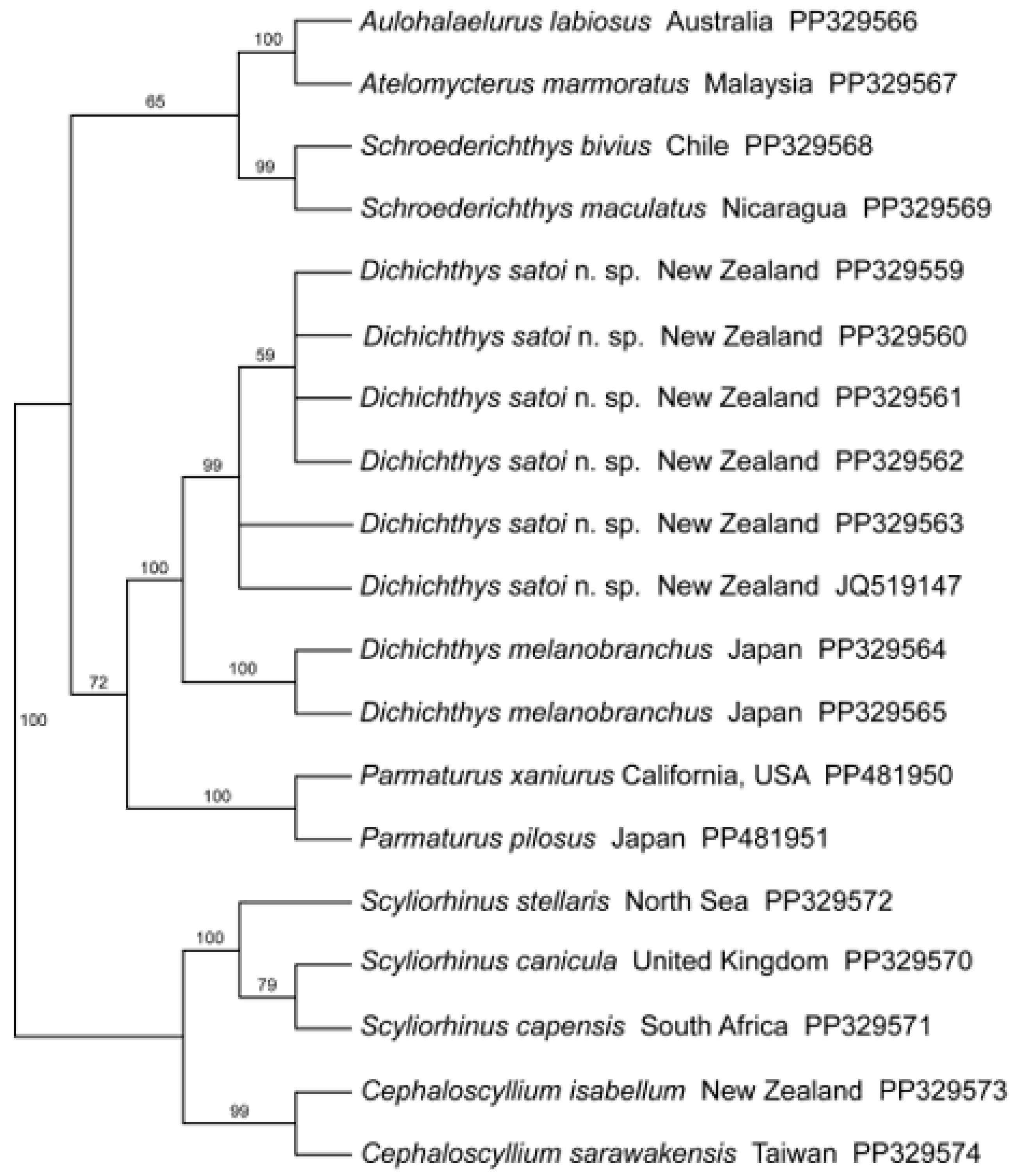
3.1.5. Molecular Analysis
3.1.6. Remarks
3.2. Genus Dichichthys (Chan, 1966 [19])
3.3. Dichichthys albimarginatus (Séret and Last, 2007 [25])
3.3.1. Synonymy
3.3.2. Specimens Examined
3.3.3. Diagnosis
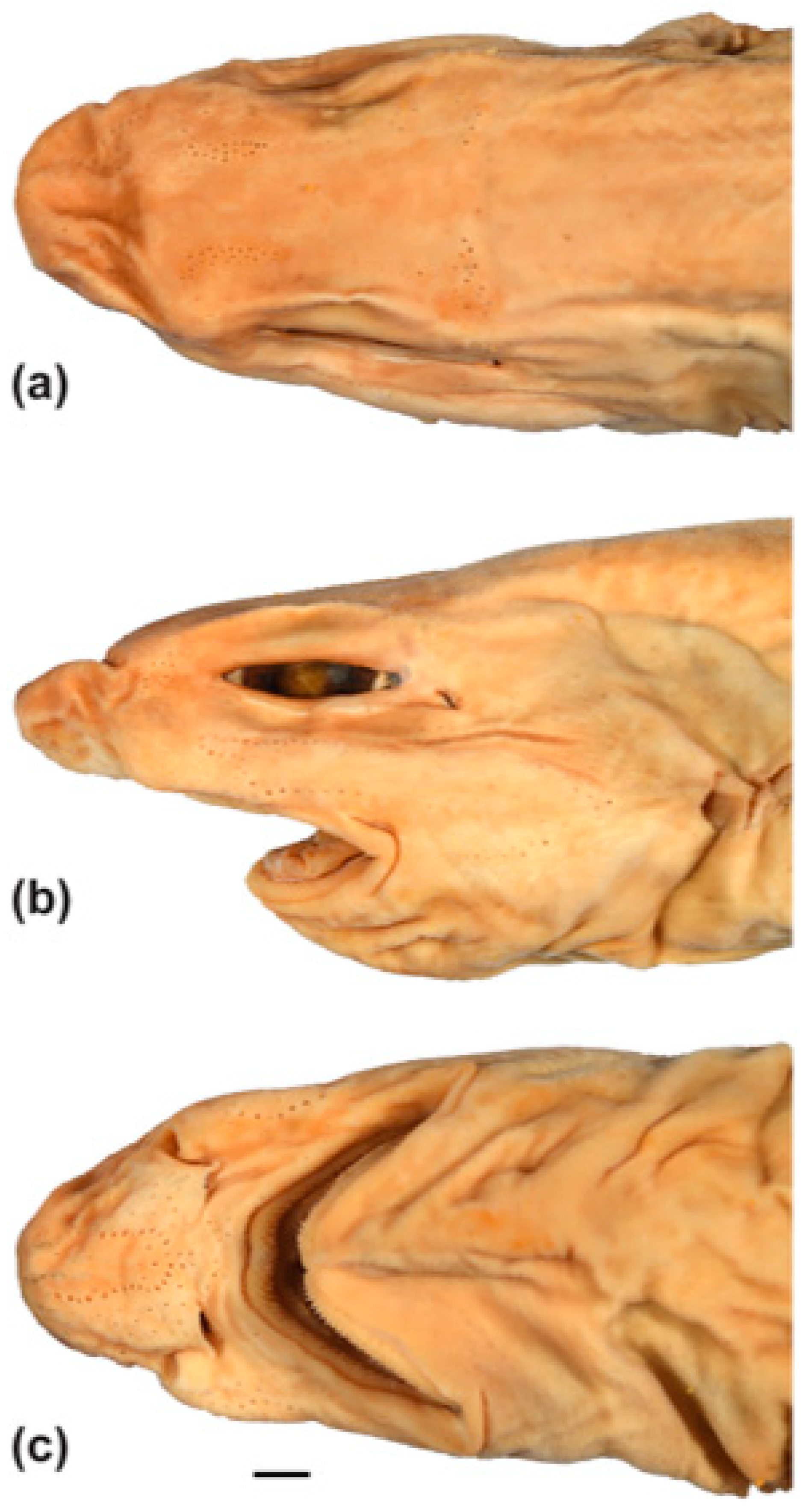
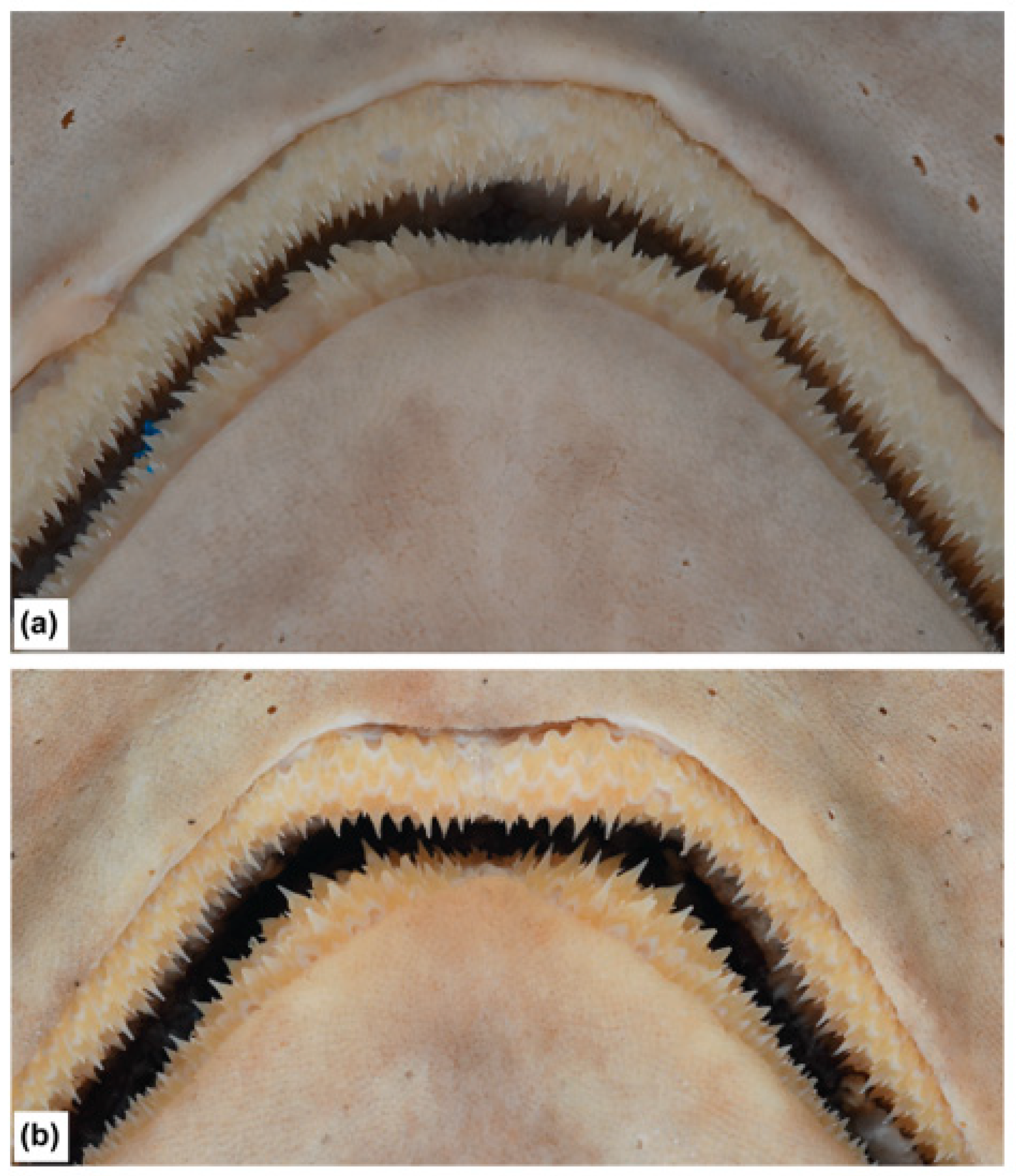
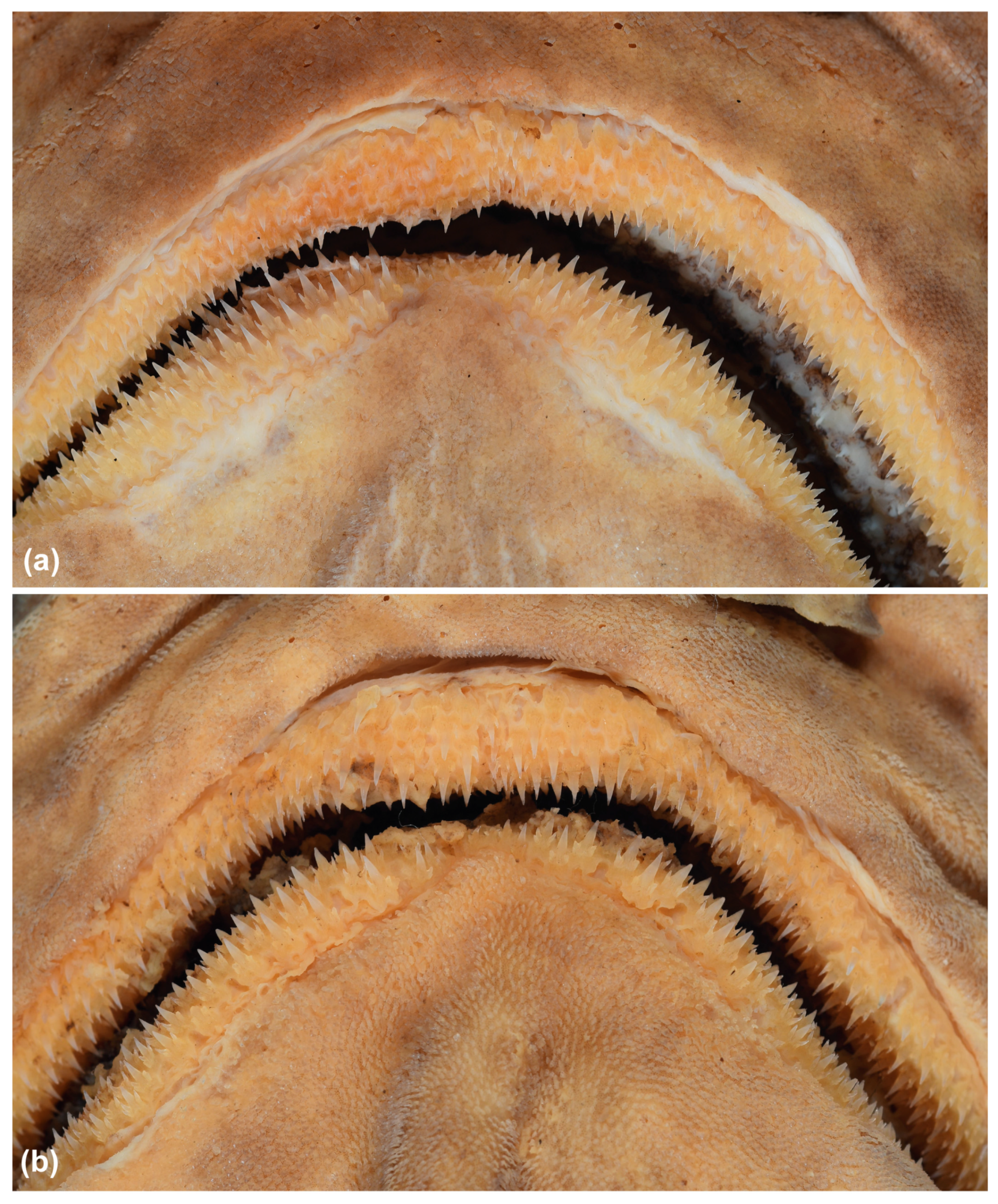
3.3.4. Description
3.3.5. Size
3.3.6. Distribution
3.3.7. Remarks
3.4. Dichichthys bigus (Séret and Last, 2007 [25])
3.4.1. Synonymy
3.4.2. Material Examined
3.4.3. Other Records
3.4.4. Diagnosis
3.4.5. Description
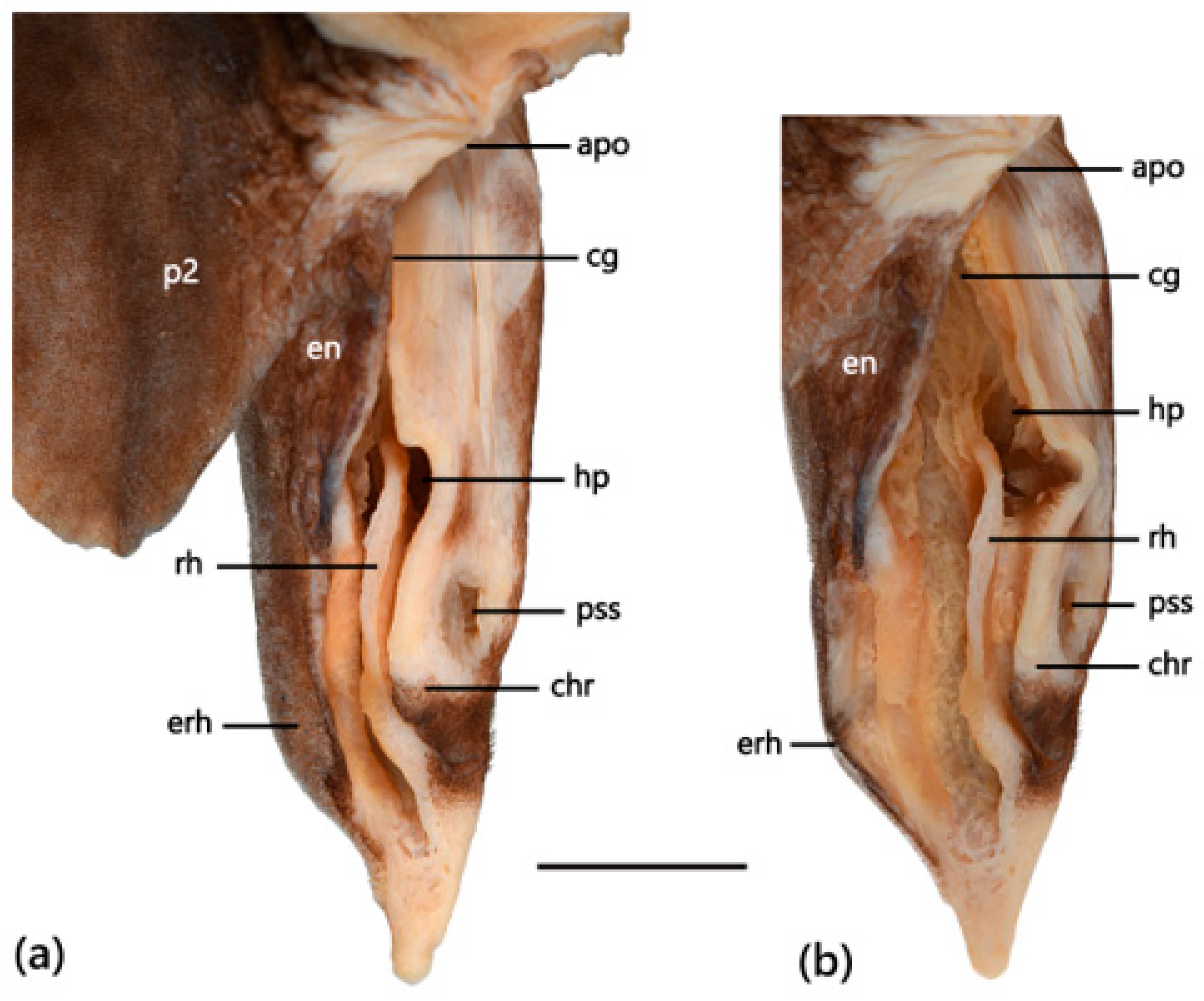
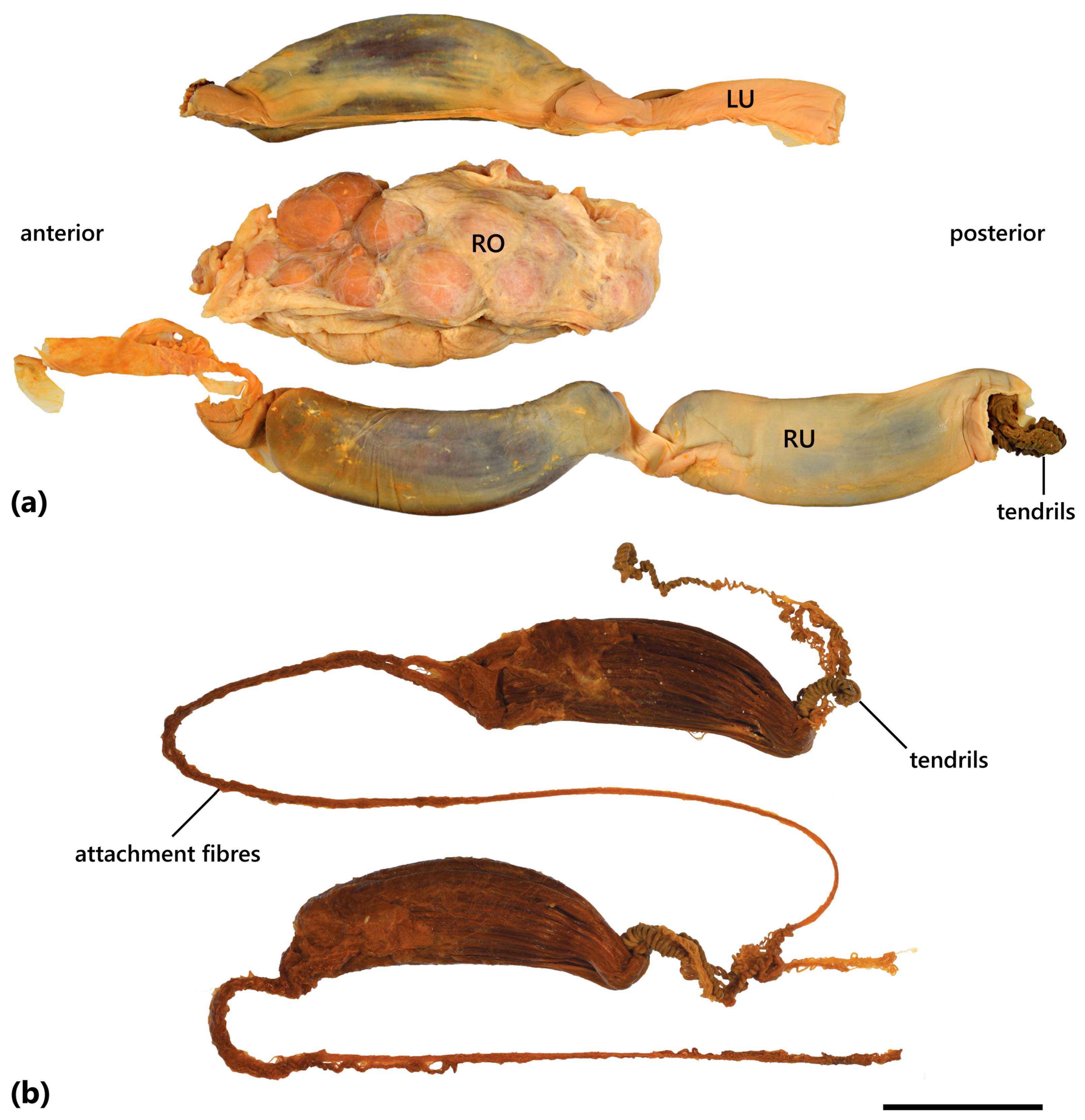
3.4.6. Egg Case
3.4.7. Size
3.4.8. Distribution
3.4.9. Remarks
3.5. Dichichthys melanobranchus (Chan, 1966 [19])
3.5.1. Synonymy
3.5.2. Material Examined
3.5.3. Diagnosis
3.5.4. Description
3.5.5. Egg Case
3.5.6. Colour
3.5.7. Size
3.5.8. Distribution
3.5.9. Remarks
3.6. Dichichthys nigripalatum (Fahmi and Ebert, 2018 [26])
3.6.1. Synonymy
3.6.2. Type Material
3.6.3. Diagnosis
3.6.4. Description
3.6.5. Size
3.6.6. Distribution
3.6.7. Remarks
3.7. Dichichthys satoi n. sp.
3.7.1. Synonymy
3.7.2. Type Material
3.7.3. Diagnosis
3.7.4. Description
3.7.5. Egg Case
3.7.6. Colour
3.7.7. Size
3.7.8. Biology
3.7.9. Distribution
3.7.10. Etymology
3.7.11. Remarks
4. Discussion
4.1. Relationships within the Carcharhiniformes
4.2. Intraspecific Variation
4.3. Reproduction
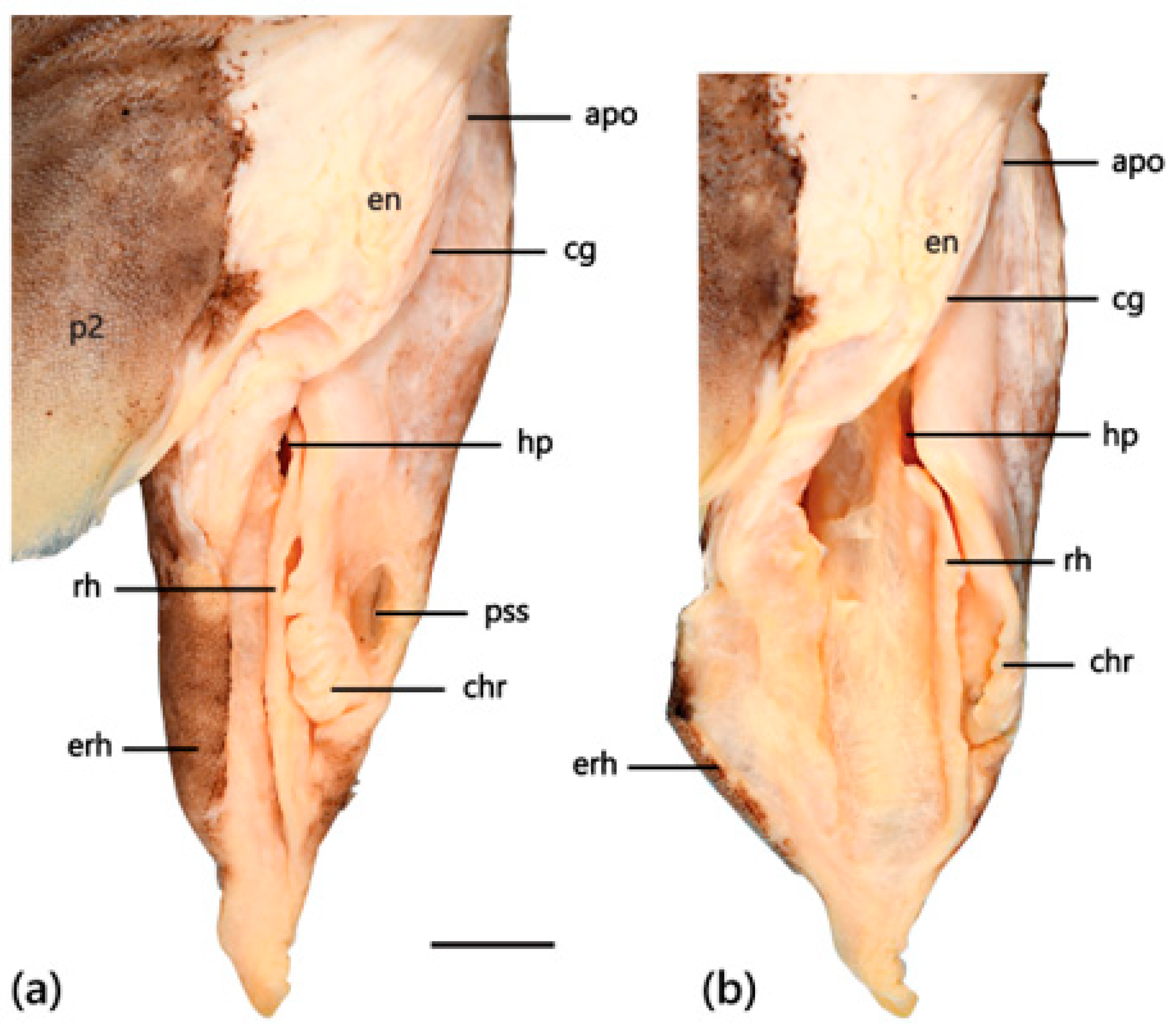
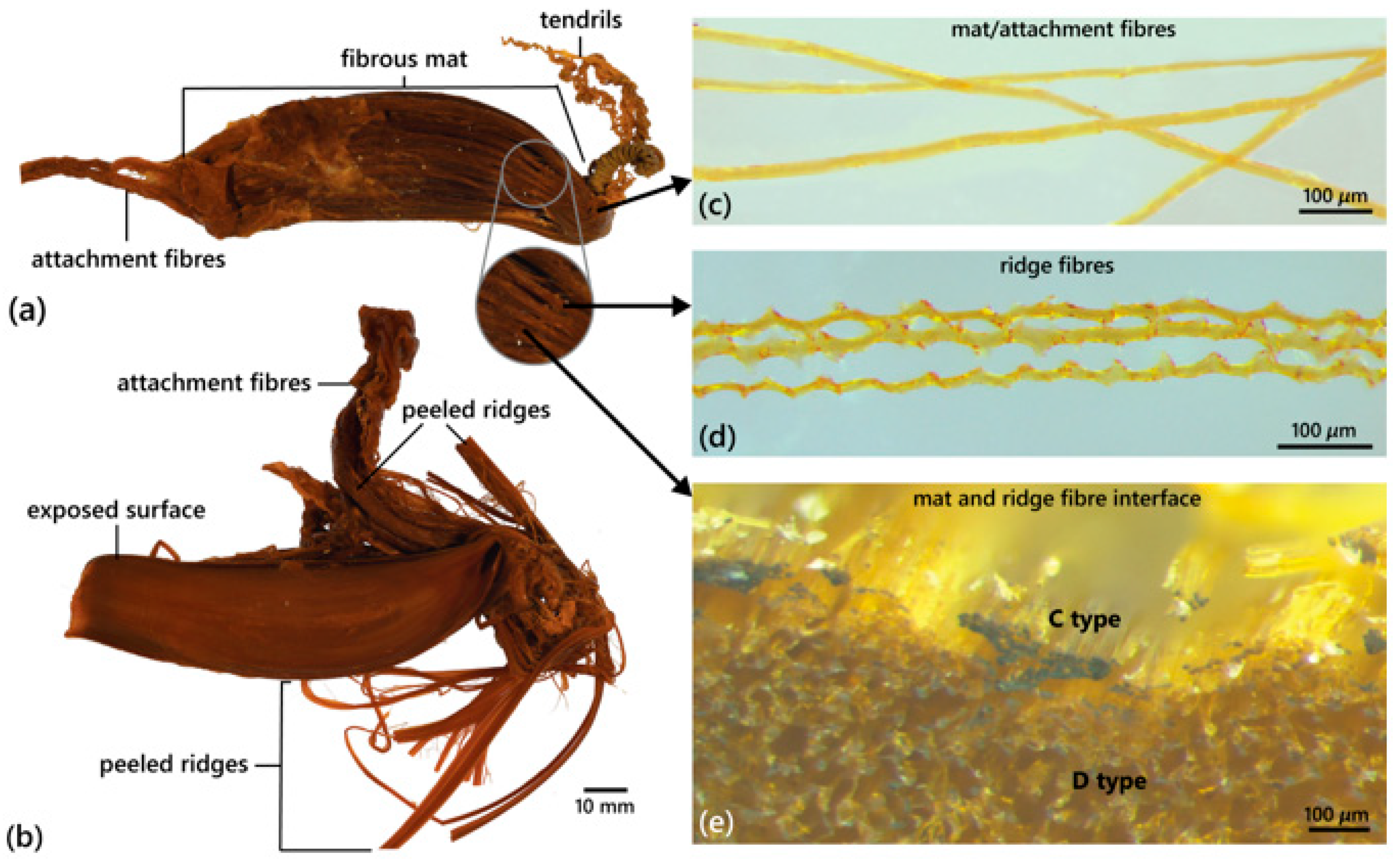
4.4. Egg Case Morphology
4.5. Parasites
- Skin irritation caused by an allergic or chemical exposure response. Chemical exposure is unlikely, given the depth and remote location. Allergic reaction to parasite bites is a possibility;
- Parasitic disease, e.g., metazoan trematode forming cysts (e.g., [69]);
- Bacterial infections, e.g., mycobacteria, can cause lumps of inflammatory cells (granulomas) (R. Chong and P. Quezada Rodriguez, CSIRO, pers. comm.);
- Fungal infection, although rare in sharks (R. Chong, CSIRO, pers. comm.).
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gill, T. Analytical synopsis of the order Squali; a revision of the nomenclature of the genera. Ann. Lyceum Nat. Hist. 1862, 7, 367–413. [Google Scholar] [CrossRef]
- Springer, S. A revision of the catsharks, family Scyliorhinidae. NOAA Tech. Rep. NMFS Circ. 1979, 422, 1–152. [Google Scholar] [CrossRef]
- Compagno, L.J.V. Sharks of the Order Carcharhiniformes; Princeton University Press: Princeton, NJ, USA, 1988; pp. 1–486. [Google Scholar]
- White, E.G. A classification and phylogeny of the elasmobranch fishes. Am. Mus. Novit. 1936, 837, 1–16. [Google Scholar]
- Smith, H.M. Description of a new notidanoid shark from the Philippine Islands representing a new family. Proc. U. S. Nat. Mus. 1912, 41, 489–491. [Google Scholar] [CrossRef]
- Iglésias, S.P.; Lecointre, G.; Sellos, D.Y. Extensive paraphylies within sharks of the order Carcharhiniformes inferred from nuclear and mitochondrial genes. Mol. Phylo. Evol. 2005, 34, 569–583. [Google Scholar] [CrossRef]
- Last, P.R.; Stevens, J.D. Sharks and Rays of Australia, 2nd ed.; CSIRO Publishing: Melbourne, Australia, 2009; pp. 1–644. [Google Scholar]
- Ebert, D.A.; Fowler, S.; Compagno, L. Sharks of the World: A Fully Illustrated Guide; Wild Nature Press: Plymouth, UK, 2013; pp. 1–528. [Google Scholar]
- Soares, K.D.A.; Mathubara, K. Combined phylogeny and new classification of catsharks (Chondrichthyes: Elasmobranchii: Carcharhiniformes). Zool. J. Linn. Soc. 2022, 195, 761–814. [Google Scholar] [CrossRef]
- Garman, S. The Plagiostomia (shark, skates and rays). Mem. Mus. Comp. Zool. 1913, 36, 1–515. [Google Scholar] [CrossRef]
- Whitley, G.P. Taxonomic notes on sharks and rays. Aust. Zoologist 1939, 9, 227–262. [Google Scholar]
- Bigelow, H.B.; Schroeder, W.C. Cephalurus, a new genus of scyliorhinid shark with redescription of the genotype, Catulus cephalus Gilbert. Copeia 1941, 1941, 73–76. [Google Scholar] [CrossRef]
- Whitley, G.P. Studies in ichthyology. No. 2. Rec. Aust. Mus. 1929, 16, 211–240. [Google Scholar] [CrossRef][Green Version]
- Rafinesque, C.S. Caratteri di Alcuni Nuovi Generi e Nuove Specie di Animali e Piante Della Sicilia, con Varie Osservazioni Sopra i Medisimi; Per le stampe di Sanfilippo: Palermo, Italy, 1810. [Google Scholar]
- Fowler, H.W. Descriptions of new fishes obtained 1907–1910, chiefly in the Philippine Islands and adjacent seas. Proc. Acad. Nat. Sci. Phila. 1934, 85, 233–367. [Google Scholar]
- Garman, S. New Plagiostomia. Bull. Mus. Comp. Zool. 1906, 46, 203–208. [Google Scholar]
- White, W.T.; Fahmi, F.; Weigmann, S. A new genus and species of catshark (Carcharhiniformes: Scyliorhinidae) from eastern Indonesia. Zootaxa 2019, 4691, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Sato, K. Skeletal morphology and systematics of Parmaturus melanobranchus (Scyliorhinidae). In Proceedings of the 8th Indo Pacific Fish Conference and 2009 ASFB Workshop and Conference, Fremantle, Australia, 31 May–5 June 2009. [Google Scholar]
- Chan, W.L. New sharks from the South China Sea. J. Zool. 1966, 148, 218–237. [Google Scholar] [CrossRef]
- Naylor, G.J.P.; Caira, J.N.; Jensen, K.; Rosana, K.A.M.; White, W.T.; Last, P.R. A DNA sequence-based approach to the identification of shark and ray species and its implications for global elasmobranch diversity and parasitology. Bull. Am. Mus. Nat. Hist. 2012, 367, 1–263. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.H. Scientific results of explorations by the U. S. Fish Commission Steamer Albatross. No. XXII. Descriptions of thirty-four new species of fishes collected in 1888 and 1889, principally among the Santa Barbara Islands and in the Gulf of California. Proc. U. S. Nat. Mus. 1892, 14, 539–566. [Google Scholar] [CrossRef]
- Jordan, D.S.; Richardson, R.E. A catalogue of the fishes of the island of Formosa, or Taiwan, based on the collections of Dr. Hans Sauter. Mem. Carnegie Mus. 1909, 4, 159–204. [Google Scholar] [CrossRef]
- Nakaya, K.; Sato, K.; Kawauchi, J.; Stewart, A.L. 14 Family Scyliorhinidae. In The Fishes of New Zealand; Roberts, C.D., Stewart, A.L., Struthers, C.D., Eds.; Te Papa Press: Wellington, New Zealand, 2015; pp. 75–89. [Google Scholar]
- Compagno, L.J.V. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Bullhead, mackerel and carpet sharks (Heterodontiformes, Lamniformes and Orectolobiformes); FAO: Rome, Italy, 2001; Volume 2, pp. 1–269. [Google Scholar]
- Séret, B.; Last, P.R. Four new species of deep-water catsharks of the genus Parmaturus (Carcharhiniformes: Scyliorhinidae) from New Caledonia, Indonesia and Australia. Zootaxa 2007, 1657, 23–39. [Google Scholar] [CrossRef]
- Fahmi, F.; Ebert, D.A. Parmaturus nigripalatum n. sp., a new species of deep-sea catshark (Chondrichthyes: Carcharhiniformes: Scyliorhinidae) from Indonesia. Zootaxa 2018, 4413, 531–540. [Google Scholar] [CrossRef]
- O’Neill, H.L.; Avila, C.; White, W.T. Description of the egg cases and juvenile colouration in two catsharks of the genus Atelomycterus (Carcharhiniformes: Scyliorhinidae). J. Fish Biol. 2020, 97, 1724–1732. [Google Scholar] [CrossRef]
- White, W.T.; O’Neill, H.L.; Cleeland, J.; Lamb, T.D.; Iglésias, S.P. Further description of the Kerguelen sandpaper skate Bathyraja irrasa (Rajiformes: Arhynchobatidae) based on additional specimens including egg cases and embryos. J. Fish Biol. 2022, 101, 659–675. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Press: Cold Spring Harbor, NY, USA, 1989; Volume 1, pp. 1–1659. [Google Scholar]
- Swofford, D. PAUP*. Phylogenetic Analysis Using Parsimony (* and Other Methods). Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougement, J. A rapid bootstrap algorithm for the RAxML web-servers. Sys. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Sabaj, M.H. Codes for natural history collections in ichthyology and herpetology. Copeia 2020, 108, 593–669. [Google Scholar] [CrossRef]
- Grandperrin, R.; Bargibant, G.; Menou, J.L. Campagne HALICAL 1 de Pêche à la Palangre de Fond Dans le Nord et sur la Ride des Loayauté, en Nouvelle-Calédonie. N.O. Alis, 21 Novembre–1er Décembre et 12–23 Décembre: Rapport Final; ORSTOM: Nouméa, New Caledonia, 1995; pp. 1–67. [Google Scholar]
- Weigmann, S. Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity. J. Fish Biol. 2016, 88, 837–1037. [Google Scholar] [CrossRef] [PubMed]
- Ebert, D.A.; Dando, M.; Fowler, S. Sharks of the World: A Complete Guide; Princeton University Press: Princeton, NJ, USA, 2021; pp. 1–607. [Google Scholar]
- White, W.T.; Ko’ou, A. An annotated checklist of the chondrichthyans of Papua New Guinea. Zootaxa 2018, 4411, 1–82. [Google Scholar] [CrossRef] [PubMed]
- White, W.T.; Baje, L.; Sabub, B.; Appleyard, S.A.; Pogonoski, J.J.; Mana, R.R. Sharks and Rays of Papua New Guinea; Australian Centre for International Agricultural Research: Canberra, Australia, 2018; pp. 1–327. [Google Scholar]
- White, W.T.; Mana, R.R.; Naylor, G.J.P. Description of a new species of deepwater catshark Apristurus yangi n.sp. (Carcharhiniformes: Pentanchidae) from Papua New Guinea. Zootaxa 2017, 4320, 25–40. [Google Scholar] [CrossRef]
- Last, P.R.; Stevens, J.D. Sharks and Rays of Australia; CSIRO: Melbourne, Australia, 1994; pp. 1–513. [Google Scholar]
- Last, P.R.; Pogonoski, J.J.; Gledhill, D.C.; White, W.T.; Walker, C.J. The deepwater demersal ichthyofauna of the western Coral Sea. Zootaxa 2014, 3887, 191–224. [Google Scholar] [CrossRef]
- Ebert, D.A.; White, W.T.; Ho, H.C.; Last, P.R.; Nakaya, K.; Seret, B.; Straube, N.; Naylor, G.J.P.; Carvalho, M.R.d. An annotated checklist of the chondrichthyans of Taiwan. Zootaxa 2013, 3752, 279–386. [Google Scholar] [CrossRef]
- Chu, Y.T.; Meng, Q.W.; Liu, J.X. Description of a new species of Scyliorhinidae from China. Acta Zootax. Sin. 1983, 8, 104–107. [Google Scholar]
- Compagno, L.J.V. FAO Species Catalogue. Sharks of the World, An Annotated and Illustrated Catalogue of Shark Species Known to Date; FAO: Rome, Italy, 1984; Volume 4, Part 2 (Carcharhiniformes). [Google Scholar]
- Compagno, L.J.V.; Niem, V.H. Scyliorhinidae: Catsharks. In The Living Marine Resources of the Western Central Pacific. FAO Species Identification Guide for Fishery Purposes. Vol. 2. Cephalopods, Crustaceans, Holothurians and Sharks; Carpenter, K.E., Niem, V.H., Eds.; FAO: Rome, Italy, 1998; pp. 1279–1292. [Google Scholar]
- Compagno, L.J.V. Checklist of living elasmobranchs. In Sharks, Skates, and Rays: The Biology of Elasmobranch Fishes; Hamlett, W.C., Ed.; Johns Hopkins University Press: Baltimore, MY, USA, 1999; pp. 471–498. [Google Scholar]
- Yano, K. Chondrichthyans of the Ryukyu Islands, Japan. In Proceedings of the 5th Indo-Pacific Fish Conference, Nouméa, New Caledonia, 3–8 November 1997; Séret, B., Sire, J.Y., Eds.; Societe Francaise d’Ichthyologie & Institut de Recherche pour le Developpement: Paris, France, 1999; pp. 351–365. [Google Scholar]
- Gledhill, D.C.; Last, P.R.; White, W.T. Resurrection of the genus Figaro Whitley (Carcharhiniformes: Scyliorhinidae) with the description of a new species from northeastern Australia. In Descriptions of New Australian Chondrichthyans; Last, P.R., White, W.T., Pogonoski, J.J., Eds.; CSIRO Marine and Atmospheric Research Paper 022; CSIRO: Hobart, Australia, 2008; pp. 179–187. [Google Scholar]
- Lee, P.F.; Shao, K.T. New record of the rare shark Parmaturus melanobranchius (Scyliorhinidae) from Taiwan. Taiwanica 2010, 55, 386–390. [Google Scholar] [CrossRef]
- Yano, K.; Kugai, K. Deep-sea chondrichthyans collected from the waters around the Okinawa Islands: Results of catch analysis of bottom longlines. Bull. Seikai Nat. Fish. Res. Inst. 1993, 71, 51–65. [Google Scholar]
- Soares, K.D.A.; Carvalho, M.R.d.; Schwingel, P.R.; Gadig, O.B.F. A new species of Parmaturus (Chondrichthyes: Carcharhiniformes: Scyliorhinidae) from Brazil, southwestern Atlantic. Copeia 2019, 107, 314–322. [Google Scholar] [CrossRef]
- Hardy, G.S. A new species of catshark in the genus Parmaturus Garman (Scyliorhinidae) from New Zealand. N. Z. J. Zool. 1985, 12, 119–124. [Google Scholar] [CrossRef]
- White, W.T.; O’Neill, H.L.; Devloo-Delva, F.; Nakaya, K.; Iglésias, S.P. What came first, the shark or the egg? Discovery of a new species of deepwater shark by investigation of egg case morphology. J. Fish Biol. 2023, 103, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Weigmann, S.; Ebert, D.A.; Clerkin, P.J.; Stehmann, M.F.W.; Naylor, G.J.P. Bythaelurus bachi n. sp., a new deep-water catshark (Carcharhiniformes, Scyliorhinidae) from the southwestern Indian Ocean, with a review of Bythaelurus species and a key to their identification. Zootaxa 2016, 4208, 401–432. [Google Scholar] [CrossRef] [PubMed]
- Human, B.A. A taxonomic revision of the catshark genus Poroderma Smith, 1837 (Chondrichthyes: Carcharhiniformes: Scyliorhinidae). Zootaxa 2006, 1229, 1–32. [Google Scholar] [CrossRef]
- Cooper, R.L.; Thiery, A.P.; Fletcher, A.G.; Delbarre, D.J.; Rasch, L.J.; Fraser, G.J. An ancient Turing-like patterning mechanism regulates skin denticle development in sharks. Sci. Adv. 2018, 4, eaau5484. [Google Scholar] [CrossRef]
- Nakaya, K.; White, W.T.; Ho, H.C. Discovery of a new mode of oviparous reproduction in sharks and its evolutionary implications. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Francis, M.P. Distribution and biology of the New Zealand endemic catshark, Halaelurus dawsoni. Environ. Biol. Fish. 2006, 75, 295–306. [Google Scholar] [CrossRef]
- White, W.T.; Last, P.R.; Stevens, J.D. Halaelurus maculosus n. sp. and H. sellus n. sp., two new species of catshark (Carcharhiniformes: Scyliorhinidae) from the Indo-West Pacific. Zootaxa 2007, 1639, 1–21. [Google Scholar] [CrossRef]
- Flammang, B.E.; Ebert, D.A.; Cailliet, G.M. Egg cases of the genus Apristurus (Chondrichthyes: Scyliorhinidae): Phylogenetic and ecological implications. Zoology 2007, 110, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Mabragaña, E.; Figueroa, D.E.; Scenna, L.B.; Díaz de Astarloa, J.M.; Colonello, J.H.; Delpiani, G. Chondrichthyan egg cases from the south-west Atlantic Ocean. J. Fish Biol. 2011, 79, 1261–1290. [Google Scholar] [CrossRef]
- Duméril, A.H.A. Monographie de la tribu des Scylliens ou Roussettes (poissons plagiostomes) comprenant deux espèces nouvelles. Rev. Mag. Zool. (Sér. 2) 1853, 5, 8–25, 73–87, 119–130. [Google Scholar]
- Günther, A. Preliminary notices of deep-sea fishes collected during the voyage of H.M.S. ‘Challenger’. Ann. Mag. Nat. Hist. (Ser. 5) 1878, 2, 17–28, 179–187, 248–251. [Google Scholar] [CrossRef]
- Müller, J.; Henle, F.G.J. Systematische Beschreibung der Plagiostomen; Veit und Comp: Berlin, Germany, 1838; pp. 1–28. [Google Scholar]
- Vazquez, D.M.; Belleggia, M.; Schejter, L.; Mabragana, E. Avoiding being dragged away: Finding egg cases of Schroederichthys bivius (Chondrichthyes: Scyliorhinidae) associated with benthic invertebrates. J. Fish Biol. 2018, 92, 248–253. [Google Scholar] [CrossRef]
- Dill, J.A.; Camus, A.C.; Leary, J.H.; Ng, T.F.F. Microscopic and molecular evidence of the first elasmobranch adomavirus, the cause of skin disease in a giant guitarfish, Rhynchobatus djiddensis. mBio 2018, 9, e00185-18. [Google Scholar] [CrossRef]
- Chong, R.S.M. Chapter 76—Oncogenic finfish viral diseases. In Aquatic Pathophysiology. Volume I. Finfish Diseases; Kibenge, F.S.B., Baldisserotto, B., Chong, R.S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 839–843. [Google Scholar]
- Volpatti, D.; Ciulli, S. Chapter 15—Lymphocystis viral disease. In Aquatic Pathophysiology. Volume I. Finfish Diseases; Kibenge, F.S.B., Baldisserotto, B., Chong, R.S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 201–216. [Google Scholar]
- Chong, R.S.M. Chapter 46—Digenetic trematode infections. In Aquatic Pathophysiology. Volume I. Finfish Diseases; Kibenge, F.S.B., Baldisserotto, B., Chong, R.S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 569–590. [Google Scholar]






























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| albimarginatus | bigus | melanobranchus | nigripal. | satoi n. sp. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Holo | PNG | Holo | Holo | Adults | Holo | Holo | Paratypes | |||
| Character | Min | Max | Min | Max | ||||||
| Morphometrics | ||||||||||
| Total length (mm) | 577 | 250 | 711 | 235 | 650 | 762 | 548 | 954 | 632 | 1046 |
| Pre-first dorsal length | 51.1 | 46.2 | 54.4 | 45.9 | 52.1 | 53.9 | 53.8 | 53.2 | 50.5 | 53.4 |
| Pre-second dorsal length | 66.9 | 62.0 | 69.4 | 59.6 | 67.6 | 69.6 | 69.7 | 67.1 | 65.2 | 68.5 |
| Precaudal length | 78.9 | 78.8 | 84.2 | 71.5 | 82.0 | 83.5 | 75.6 | 81.8 | 80.4 | 83.2 |
| Prepectoral length | 19.2 | 19.1 | 18.8 | 19.2 | 18.0 | 21.0 | 19.4 | 19.1 | 18.3 | 21.3 |
| Prepelvic length | 46.9 | 42.4 | 48.1 | 39.0 | 46.7 | 49.5 | 49.3 | 47.3 | 44.4 | 49.4 |
| Preanal length | 59.1 | 55.2 | 62.4 | 51.1 | 60.6 | 62.3 | 62.2 | 60.1 | 58.1 | 62.1 |
| Snout-vent length | 49.7 | 44.8 | 50.8 | – | 49.7 | 51.8 | 51.6 | 50.5 | 47.8 | 52.4 |
| Preorbital length (direct) | 6.0 | 6.7 | 6.0 | 7.0 | 6.3 | 7.3 | 6.8 | 5.8 | 5.7 | 7.5 |
| Eye length | 3.9 | 5.6 | 4.1 | 4.5 | 4.5 | 4.8 | 4.2 | 4.2 | 3.8 | 4.6 |
| Eye height | 0.5 | 1.5 | 1.2 | – | 0.8 | 1.1 | 2.2 | 1.0 | 0.7 | 1.3 |
| Interorbital space | 7.7 | 8.6 | 5.4 | – | 6.1 | 6.6 | 6.1 | 6.3 | 5.8 | 7.4 |
| Prespiracular length | 11.8 | 12.2 | 10.9 | – | 11.2 | 12.3 | 11.3 | 11.0 | 10.0 | 12.2 |
| Spiracle length | 0.2 | 0.7 | 0.6 | – | 0.6 | 0.6 | 0.3 | 0.4 | 0.3 | 0.6 |
| Prenarial length | 3.7 | 5.2 | 4.2 | 4.0 | 4.8 | 5.4 | 4.7 | 4.2 | 4.0 | 5.2 |
| Nostril width | 2.5 | 3.0 | 2.3 | – | 2.4 | 3.0 | 2.6 | 2.6 | 2.5 | 3.1 |
| Internarial space | 2.1 | 2.8 | 2.2 | 2.5 | 2.1 | 2.6 | 2.6 | 2.2 | 2.0 | 2.7 |
| Anterior nasal flap length | 1.1 | 0.9 | 1.1 | – | 1.0 | 1.3 | 1.4 | 1.0 | 0.8 | 1.2 |
| Preoral length | 6.3 | 7.0 | 6.1 | 8.5 | 6.4 | 7.1 | 5.0 | 5.4 | 5.3 | 6.9 |
| Mouth width | 9.1 | 9.6 | 7.7 | 8.9 | 9.2 | 9.9 | 9.0 | 9.4 | 9.0 | 11.4 |
| Mouth length | 3.7 | 4.1 | 3.2 | 7.0 | 3.4 | 4.4 | 4.2 | 3.7 | 3.2 | 4.1 |
| Upper labial furrow length | 1.7 | 1.7 | 1.6 | 1.0 | 1.6 | 1.9 | 1.9 | 1.4 | 1.4 | 2.0 |
| Lower labial furrow length | 2.0 | 2.1 | 1.6 | 2.3 | 1.8 | 2.0 | 1.9 | 2.5 | 2.0 | 2.8 |
| Prebranchial length | 15.8 | 16.2 | 17.5 | 17.0 | 16.3 | 18.5 | 15.5 | 16.9 | 15.5 | 18.0 |
| Head length | 19.2 | 19.7 | 20.0 | – | 19.6 | 21.4 | 20.7 | 19.4 | 18.8 | 21.6 |
| First gill slit height | 1.7 | 2.2 | 2.3 | 1.9 | 1.4 | 2.2 | 2.2 | 2.1 | 1.8 | 2.7 |
| Fifth gill slit height | 0.8 | 1.9 | 1.5 | 1.2 | 0.8 | 1.2 | 1.3 | 1.2 | 1.0 | 1.9 |
| First dorsal anterior margin | 8.2 | 9.9 | 5.8 | – | 7.2 | 8.0 | 8.6 | 7.9 | 8.0 | 9.1 |
| First dorsal base | 6.8 | 7.3 | 5.1 | 6.1 | 6.0 | 6.7 | 6.5 | 6.5 | 6.1 | 7.3 |
| First dorsal height | 3.2 | 2.6 | 3.7 | 3.4 | 3.4 | 3.9 | 3.8 | 3.7 | 2.9 | 3.8 |
| First dorsal inner margin | 1.6 | 3.1 | 1.7 | – | 1.1 | 1.9 | 2.1 | 1.9 | 1.5 | 2.1 |
| First dorsal posterior margin | 3.1 | 2.6 | 3.1 | – | 2.7 | 3.1 | 3.3 | 3.0 | 2.3 | 3.3 |
| Interdorsal space | 9.8 | 9.7 | 9.1 | – | 8.4 | 10.0 | 9.5 | 8.0 | 7.7 | 9.9 |
| Second dorsal anterior margin | 10.4 | 13.0 | 9.9 | – | 9.6 | 10.9 | 10.8 | 11.1 | – | – |
| Second dorsal base | 8.0 | 10.7 | 7.8 | 9.4 | 8.2 | 9.2 | 8.6 | 9.4 | 8.6 | 10.2 |
| Second dorsal height | 4.6 | 5.2 | 5.1 | 3.4 | 4.7 | 5.2 | 4.9 | 4.8 | – | – |
| Second dorsal inner margin | 1.8 | 2.2 | 1.9 | – | 0.9 | 1.8 | 1.7 | 1.9 | – | – |
| Second dorsal posterior margin | 3.9 | 4.0 | 4.2 | – | 4.4 | 4.6 | 4.1 | 4.3 | – | – |
| Pectoral anterior margin | 9.4 | 10.6 | 8.1 | 10.3 | 7.7 | 9.1 | 9.6 | 7.7 | 8.3 | 9.6 |
| Pectoral base | 4.4 | 4.8 | 4.6 | 4.7 | 4.8 | 5.2 | 5.3 | 5.0 | 5.1 | 5.8 |
| Pectoral inner margin | 4.3 | 4.4 | 2.6 | – | 2.9 | 4.0 | 4.0 | 3.0 | 2.6 | 4.6 |
| Pectoral posterior margin | 6.4 | 5.7 | 6.2 | – | 6.0 | 6.7 | 6.4 | 6.1 | 5.4 | 6.5 |
| Pectoral–pelvic space | 24.8 | 18.4 | 25.0 | – | 24.0 | 26.0 | 27.2 | 24.6 | 21.2 | 26.1 |
| Pelvic anterior margin | 7.6 | 7.1 | 6.8 | 8.9 | 5.9 | 8.0 | 6.8 | 7.0 | 6.4 | 9.6 |
| Pelvic base | 5.7 | 6.1 | 7.5 | 6.2 | 6.2 | 8.2 | 5.5 | 6.0 | 6.4 | 8.8 |
| Pelvic length | 9.4 | 9.4 | 10.2 | – | 9.1 | 10.7 | 9.2 | 9.8 | 9.2 | 12.2 |
| Pelvic inner margin | 2.4 | 3.8 | 2.8 | – | 2.9 | 3.8 | 3.6 | 4.1 | 2.8 | 4.2 |
| Pelvic posterior margin | 3.7 | 5.0 | 5.3 | 3.2 | 4.1 | 5.5 | 4.6 | 4.3 | 4.1 | 6.1 |
| Pelvic–anal space | 7.2 | 7.7 | 6.9 | – | 5.6 | 8.0 | 7.0 | 7.1 | 5.3 | 7.4 |
| Anal anterior margin | 9.0 | 10.4 | 8.4 | – | 7.4 | 9.1 | 9.3 | 9.2 | 9.2 | 10.5 |
| Anal base | 10.7 | 10.9 | 9.8 | 12.9 | 11.2 | 11.9 | 10.7 | 11.5 | 11.0 | 12.0 |
| Anal length | 11.6 | 11.7 | 11.2 | – | 11.8 | 13.2 | 11.5 | 13.1 | 12.0 | 13.4 |
| Anal Inner margin | 1.3 | 1.5 | 1.3 | – | 0.6 | 1.3 | 0.8 | 1.4 | 1.1 | 1.7 |
| Anal posterior margin | 4.8 | 4.6 | 5.8 | 1.5 | 5.6 | 7.0 | 5.4 | 7.2 | 5.4 | 7.4 |
| Dorsal–caudal space | 2.8 | 7.3 | 7.0 | – | 5.5 | 7.0 | 2.0 | 5.2 | 4.7 | 6.5 |
| Anal–caudal space | 11.4 | 10.1 | 10.4 | – | 7.1 | 9.2 | 4.5 | 7.5 | 6.6 | 9.5 |
| Caudal peduncle height | 2.9 | 3.4 | 2.6 | – | 2.6 | 2.8 | 2.2 | 2.8 | 2.2 | 2.9 |
| Caudal peduncle width | 1.8 | 1.6 | 1.3 | – | 1.5 | 2.0 | 1.5 | 1.2 | 1.3 | 1.6 |
| Dorsal caudal margin | 21.9 | 21.8 | 14.3 | 27.0 | 15.6 | 17.8 | 20.5 | 18.1 | 16.3 | 19.7 |
| Preventral caudal margin | 7.2 | 12.6 | 6.8 | 23.2 | 8.2 | 10.1 | 12.4 | 7.7 | 7.8 | 10.8 |
| Subterminal caudal margin | 3.3 | 4.6 | 3.8 | 6.2 | 2.8 | 3.8 | 3.6 | 3.8 | 2.9 | 4.0 |
| Clasper outer length | 5.1 | – | – | – | 4.3 | 4.9 | 5.4 | 4.6 | 0.0 | 4.7 |
| Clasper inner length | 5.9 | – | – | – | 6.0 | 6.1 | 8.8 | 6.4 | 0.0 | 6.3 |
| Clasper base width | 1.7 | – | – | – | 1.6 | 2.0 | 1.6 | 1.9 | 0.0 | 2.1 |
| Head height | 6.9 | 8.0 | 8.6 | – | 6.5 | 9.0 | 7.2 | 12.3 | 5.1 | 12.7 |
| Head width | 12.9 | 10.9 | 9.6 | – | 10.5 | 13.0 | 10.8 | 10.8 | 11.3 | 13.9 |
| Anal height | 3.8 | 4.5 | 4.1 | 4.6 | 3.9 | 4.3 | 3.9 | 5.2 | 4.3 | 5.7 |
| Terminal caudal margin | 3.8 | 3.7 | 3.8 | – | 3.4 | 4.3 | 3.5 | 3.8 | 3.2 | 4.2 |
| Meristics | ||||||||||
| Intestinal spiral valve turns | – | – | 9 | – | 10 | 10 | – | – | 13 | 14 |
| Monospondylous centra | 43 | 46 | 47 | – | 45 | 46 | 42 | 47 | 45 | 47 |
| Precaudal centra (to lower caudal fin origin) | 95 | 92 | 102 | 102 | 96 | 99 | 90 | 96 | 95 | 101 |
| Total vertebral centra | 136 | 133 | 144 | – | 133 | 145 | 130 | 140 | 133 | 143 |
| bigus | satoi n. sp. | |||
|---|---|---|---|---|
| Egg Case 1 | Egg Case 2 | Min. | Max. | |
| Egg case length (mm) | 89.4 | 85.4 | 92.0 | 112.3 |
| Body length | 79.6 | 74.5 | 67.2 | 73.8 |
| Posterior case width | 15.5 | 13.3 | 15.4 | 17.4 |
| Anterior waist width | 24.6 | 25.6 | 23.2 | 25.5 |
| Posterior waist width | 20.1 | 19.4 | 19.2 | 21.7 |
| Anterior case width | 23.5 | 21.9 | 22.1 | 24.4 |
| Anterior border width | 19.2 | 18.2 | 16.4 | 20.8 |
| Height | 22.0 | 22.2 | 19.2 | 24.2 |
| Maximum width | 31.2 | 32.5 | 28.6 | 33.0 |
| Left single keel depth | 5.5 | 5.5 | 4.4 | 5.8 |
| Right lower keel depth | 2.7 | 2.7 | 4.7 | 6.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, W.T.; Stewart, A.L.; O’Neill, H.L.; Naylor, G.J.P. Dichichthyidae, a New Family of Deepwater Sharks (Carcharhiniformes) from the Indo–West Pacific, with Description of a New Species. Fishes 2024, 9, 121. https://doi.org/10.3390/fishes9040121
White WT, Stewart AL, O’Neill HL, Naylor GJP. Dichichthyidae, a New Family of Deepwater Sharks (Carcharhiniformes) from the Indo–West Pacific, with Description of a New Species. Fishes. 2024; 9(4):121. https://doi.org/10.3390/fishes9040121
Chicago/Turabian StyleWhite, William T., Andrew L. Stewart, Helen L. O’Neill, and Gavin J. P. Naylor. 2024. "Dichichthyidae, a New Family of Deepwater Sharks (Carcharhiniformes) from the Indo–West Pacific, with Description of a New Species" Fishes 9, no. 4: 121. https://doi.org/10.3390/fishes9040121
APA StyleWhite, W. T., Stewart, A. L., O’Neill, H. L., & Naylor, G. J. P. (2024). Dichichthyidae, a New Family of Deepwater Sharks (Carcharhiniformes) from the Indo–West Pacific, with Description of a New Species. Fishes, 9(4), 121. https://doi.org/10.3390/fishes9040121