
Cardiac Transcriptome and Histology of the Heart of the Male Chinese Mitten Crab (Eriocheir sinensis) under High-Temperature Stress


Abstract
1. Introduction
2. Materials and Methods
2.1. Crabs
2.2. Experimental Design and Sampling
2.3. Total RNA Extraction and cDNA Library Construction
2.4. RNA Sequencing and Transcriptome De Novo Assembly
2.5. Transcription Factor (TF) and Structure Analysis
2.6. Correlation and PCA Analysis
2.7. DEGs and Their Functional Analysis
2.8. Quantitative Real-Time PCR (qRT-PCR)
2.9. Analyses of Histological Sections of the Heart
3. Results and Discussion
3.1. Transcriptome and Unigene Annotation
3.2. SSR
3.3. TFs
3.4. DEG Analysis
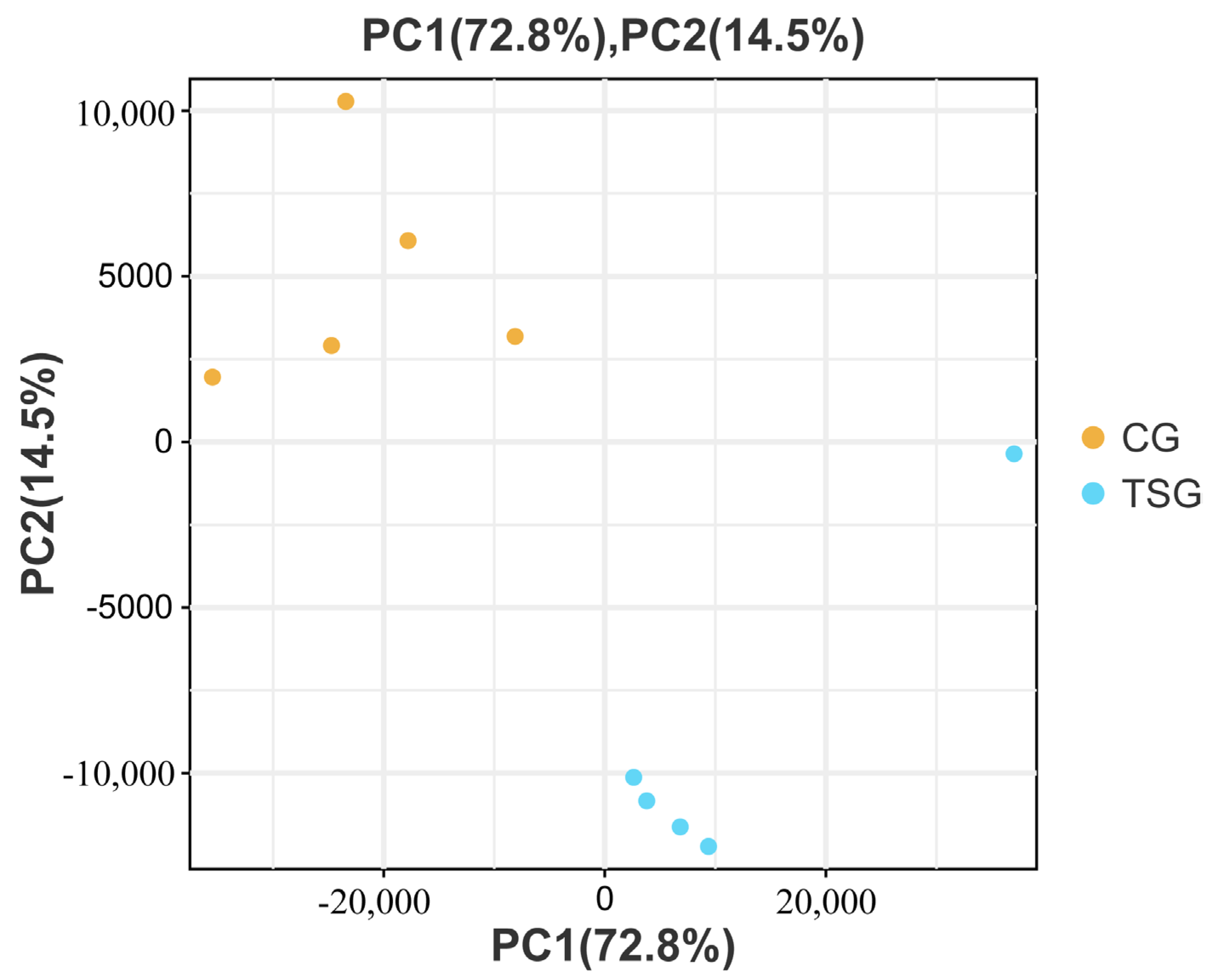
3.5. PCA
3.6. Functional Annotation
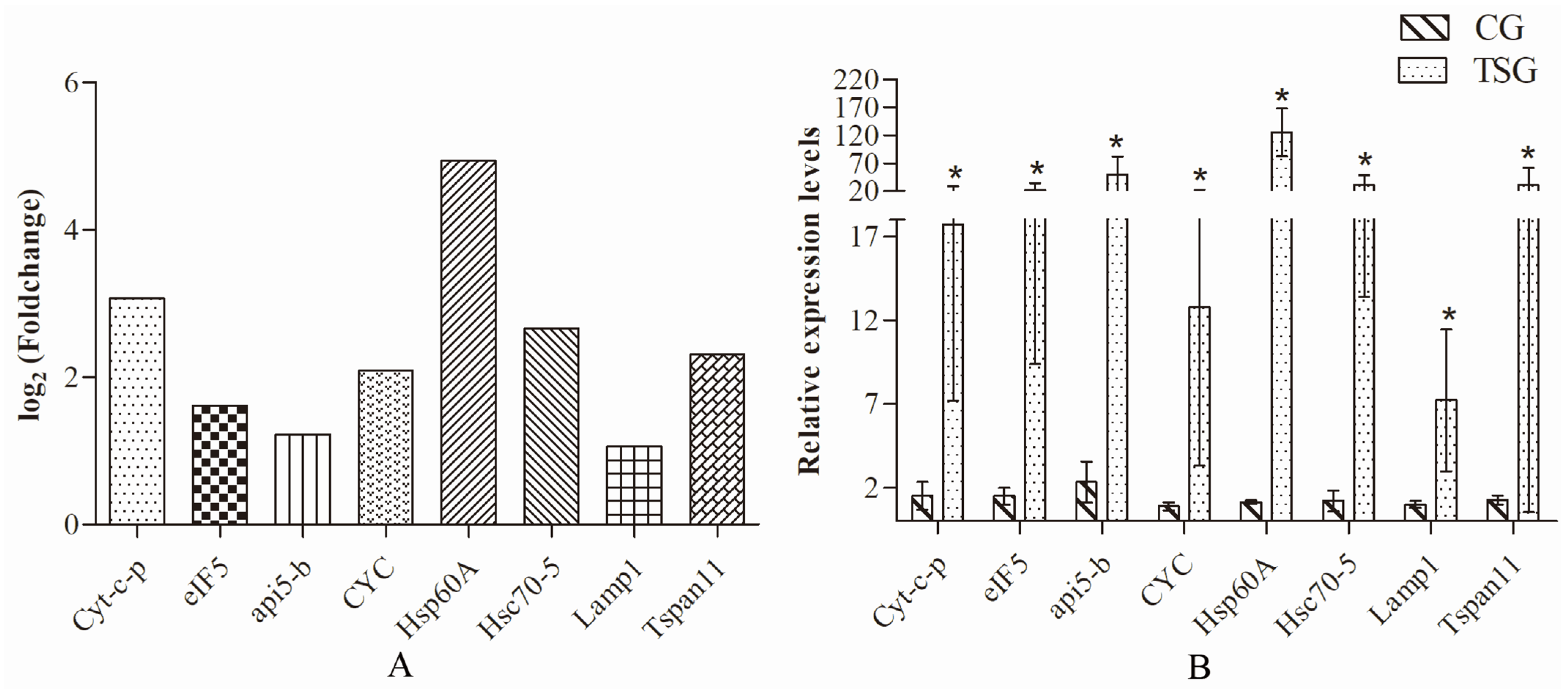
3.7. qRT-PCR
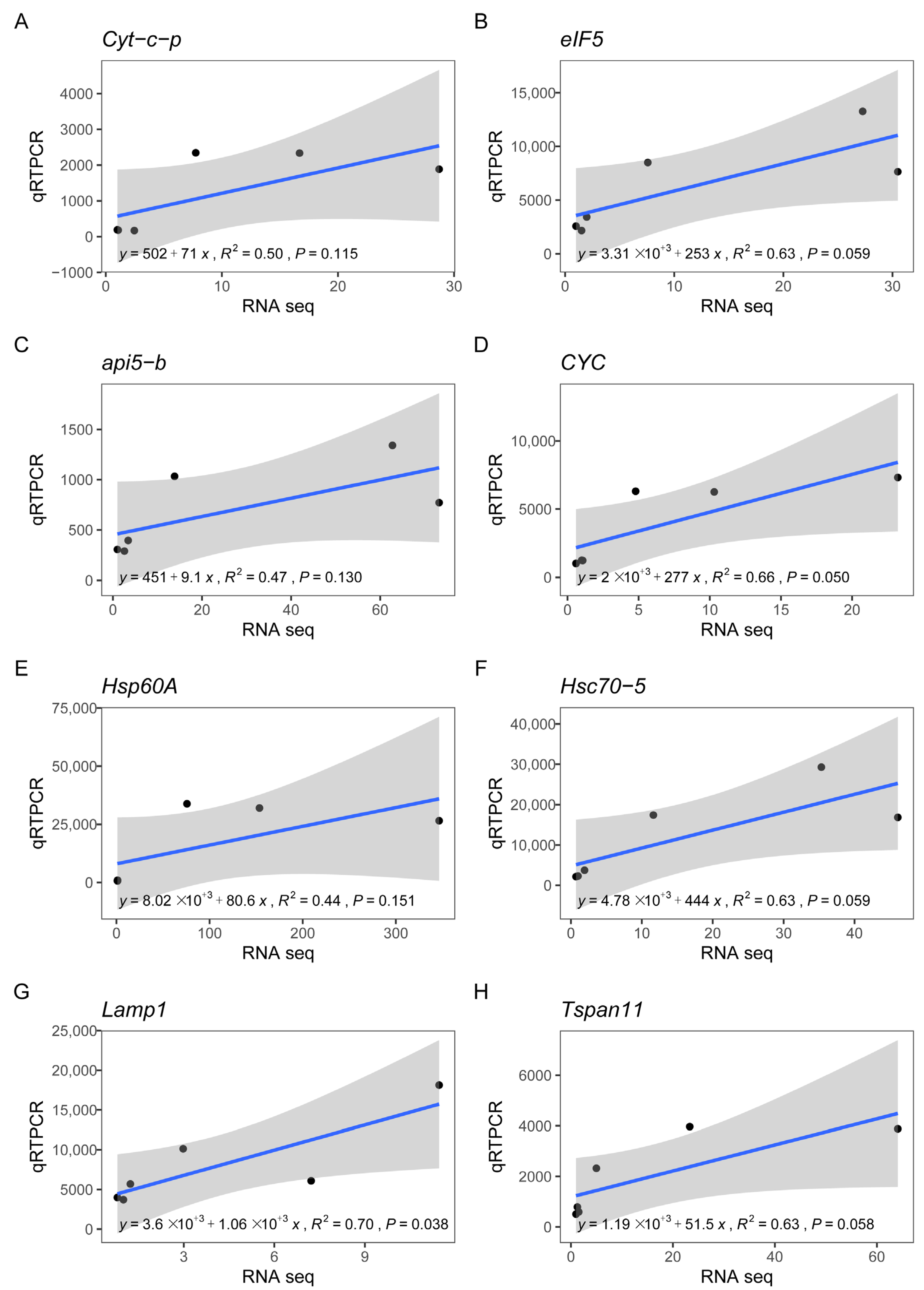
3.8. Correlation between RN-seq and qRT-PCR Analyses
3.9. Heart Tissue Histology
4. Conclusions and Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bowden, T. Modulation of the immune system of fish by their environment. Fish Shellfish. Immunol. 2008, 254, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zeng, G.; Yang, X.; Jiang, Z. Future changes in extreme high temperature over china at 1.5 C–5 C global warming based on cmip6 simulations. Adv. Atmos. Sci. 2021, 38, 253–267. [Google Scholar] [CrossRef]
- Gotthard, K. Growth strategies of ectothermic animals in temperate environments. Environ. Anim. Dev. 2001, 287–304. Available online: https://xueshu.baidu.com/usercenter/paper/show?paperid=5ca32de23cc3ed5113925ea0fe9c0aa7&site=xueshu_se&hitarticle=1 (accessed on 8 January 2024).
- Goikoetxea, A.; Sadoul, B.; Blondeau-Bidet, E.; Aerts, J.; Blanc, M.-O.; Parrinello, H.; Barrachina, C.; Pratlong, M.; Geffroy, B. Genetic pathways underpinning hormonal stress responses in fish exposed to short-and long-term warm ocean temperatures. Ecol. Indic. 2021, 120, 106937. [Google Scholar] [CrossRef]
- Alfonso, S.; Gesto, M.; Sadoul, B. Temperature increase and its effects on fish stress physiology in the context of global warming. J. Fish Biol. 2021, 98, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.-b.; Su, Y.q.; Mao, Y.; You, X.-x.; Ding, S.; Wang, J. Dietary supplementation with bacillus can improve the growth and survival of the kuruma shrimp Marsupenaeus japonicus in high-temperature environments. Aquac. Int. 2013, 22, 607–617. [Google Scholar] [CrossRef]
- Sanda, T.; Shimizu, T.; Iwasaki, T.; Dan, S.; Hamasaki, K. Effect of temperature on survival, intermoltperiod, and growth of juveniles of two mud crab species, Scylla paramamosain and Scylla serrata (Decapoda: Brachyura: Portunidae), under laboratory conditions. Nauplius 2022, 30, e2022012. [Google Scholar] [CrossRef]
- Yu, K.; Shi, C.; Ye, Y.; Li, R.; Mu, C.; Ren, Z.; Wang, C. The effects of overwintering temperature on the survival of female adult mud crab, Scylla paramamosain, under recirculating aquaculture systems as examined by histological analysis of the hepatopancreas and expression of apoptosis-related genes. Aquaculture 2022, 565, 739080. [Google Scholar] [CrossRef]
- Zarco-Perello, S.; Pratchett, M.; Liao, V. Temperature-growth performance curves for a coral reef fish, Acanthochromis polyacanthus. Galaxea 2012, 14, 97–103. [Google Scholar] [CrossRef]
- Chen, D.-W.; Zhang, M.; Shrestha, S. Compositional characteristics and nutritional quality of Chinese mitten crab (Eriocheir sinensis). Food Chem. 2007, 103, 1343–1349. [Google Scholar] [CrossRef]
- China Fishery Bureau. China Fisheries Yearbook; Chinese Agriculture Express: Beijing, China, 2023; p. 24. Available online: https://www.doc88.com/p-99029850248323.html (accessed on 8 January 2024).
- Zhang, T.; Li, T. Ecological observations on molting of juveniles of the Chinese mitten crab, Eriocheir sinensis. J. Lake Sci. 1999, 11, 333–337. [Google Scholar]
- Li, Z.; Zhao, Z.; Luo, L.; Wang, S.; Zhang, R.; Guo, K.; Yang, Y. Immune and intestinal microbiota responses to heat stress in Chinese mitten crab (Eriocheir sinensis). Aquaculture 2023, 563, 738965. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, Q.; Zhang, T.; Li, Z.; Liu, J. Effects of water temperature on growth, feeding and molting of juvenile Chinese mitten crab Eriocheir sinensis. Aquaculture 2017, 468, 169–174. [Google Scholar] [CrossRef]
- Peng, J.; Zhao, Y.; Xu, Z.; Liu, B.; Duan, C.; Tang, Y. Effect of temperature stress on the survival of juvenile Chinese mitten crab (Eriocheir sinensis). Iran. J. Fish. Sci. 2019, 18, 763–774. [Google Scholar]
- McMahon, B.R. Control of cardiovascular function and its evolution in Crustacea. J. Exp. Biol. 2001, 204, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Keen, A.N.; Klaiman, J.M.; Shiels, H.A.; Gillis, T.E. Temperature-induced cardiac remodeling in fish. J. Exp. Biol. 2017, 220, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.; Qadri, S.A.; Wang, H.; Worden, M.K. Temperature acclimation alters cardiac performance in the lobster Homarus americanus. J. Comp. Physiol. A 2006, 192, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Kushinsky, D.; Morozova, E.O.; Marder, E. In vivo effects of temperature on the heart and pyloric rhythms in the crab Cancer borealis. J. Exp. Biol. 2019, 222, jeb199190. [Google Scholar] [CrossRef] [PubMed]
- DeWachter, B.; Wilkens, J.L. Comparison of temperature effects on heart performance of the Dungeness crab, Cancer magister, in vitro and in vivo. Biol. Bull. 1996, 190, 385–395. [Google Scholar] [CrossRef]
- Duan, P.; Tian, Y.; Li, Z.; Chen, S.; Li, L.; Wang, X.; Wang, L.; Liu, Y.; Zhai, J.; Li, W. Comparative transcriptome analysis of hybrid jinhu grouper (Epinephelus fuscoguttatus♀ × Epinephelus tukula♂) and Epinephelus fuscoguttatus under temperature stress. Aquaculture 2024, 578, 740037. [Google Scholar] [CrossRef]
- Pan, T.; Yang, M.; Jiang, H.; Li, T.; Duan, G.; Ling, J.; Gao, Q. Effect of Astragalus membranaceus on transcriptome and survival of hybrid yellow catfish (Pseudobagrus vachellii♂ × Tachysurus fulvidraco♀) inresponse to Aeromonas hydrophila challenge. Fishes 2023, 8, 454. [Google Scholar] [CrossRef]
- Bao, J.; Wang, X.; Feng, C.; Li, X.; Jiang, H. Trehalose metabolism in the Chinese mitten crab Eriocheir sinensis: Molecular cloning of trehalase and its expression during temperature stress. Aquac. Rep. 2021, 20, 100770. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one fastq preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.; Haas, B.; Yassour, M.; Levin, J.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from rna-seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. Busco update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef] [PubMed]
- Becker, R.A.; Chambers, J.M.; Wilks, A.R. The New S Language; Wadsworth&Brooks/Cole: London, UK, 1988. [Google Scholar]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by rna-seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression of rna-seq data at the gene level the deseq package. Eur. Mol. Biol. Lab. (EMBL) 2012, 10, f1000research. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Guo, X.-F.; Liu, M.; Zhou, Y.-L.; Wei, W.-Y.; Li, Z.; Zhou, L.; Wang, Z.-W.; Gui, J.-F. Genetic diversity evaluation and population structure analysis of red swamp crayfish (Procambarus clarkii) from lakes and rice fields by ssr markers. Fishes 2022, 7, 142. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, M.; Deng, Y.; Yang, Y.; Li, X.; Lu, Q.; Ge, J.; Pan, J.; Xu, Z. Molecular cloning, characterization and expression analysis of two juvenile hormone esterase-like carboxylesterase cDNAs in Chinese mitten crab, Eriocheir sinensis. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 205, 46–53. [Google Scholar] [CrossRef]
- Shen, H.; Ma, Y.; Hu, Y.; Zhou, X. Cloning of the ecdysone receptor gene from the Chinese mitten crab, Eriocheir sinensis, and sexually dimorphic expression of two splice variants. J. World Aquac. Soc. 2015, 46, 421–433. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, H.; Li, H.; Wang, A.; Yu, H.-y. Effect of high temperature on immune response of grass carp (Ctenopharyngodon idellus) by transcriptome analysis. Fish Shellfish. Immunol. 2016, 58, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Ma, A.; Yang, S.; Huang, Z. Integrated metabolome and transcriptome analyses revealing the effects of thermal stress on lipid metabolism in juvenile turbot Scophthalmus maximus. J. Therm. Biol. 2021, 99, 102937. [Google Scholar] [CrossRef] [PubMed]
- Colson-Proch, C.; Morales, A.; Hervant, F.; Konecny, L.; Moulin, C.; Douady, C.J. First cellular approach of the effects of global warming on ground water organisms: A study of the hsp70 gene expression. Cell Stress Chaperones 2010, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Han, J.; Chen, P.; Chang, Z.; He, Y.; Liu, P.; Wang, Q.; Li, J. Cloning of a heat shock protein 90 (hsp90) gene and expression analysis in the ridgetail white prawn Exopalaemon carinicauda. Fish Shellfish. Immunol. 2012, 32, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ye, H.; Huang, H.; Li, S.; Liu, X.; Zeng, X.; Gong, J. Expression of hsp70 in the mud crab, Scylla paramamosain in response to bacterial, osmotic, and thermal stress. Cell Stress Chaperones 2013, 18, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Oksala, N.K.; Ekmekçi, F.G.; Özsoy, E.; Kirankaya, Ş.; Kokkola, T.; Emecen, G.; Lappalainen, J.; Kaarniranta, K.; Atalay, M. Natural thermal adaptation increases heat shock protein levels and decreases oxidative stress. Redox Biol. 2014, 3, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Mahanty, A.; Mohanty, S.; Mohanty, B.P. Dietary supplementation of curcumin augments heat stress tolerance through upregulation of nrf-2-mediated antioxidative enzymes and hsps in Puntius sophore. Fish Physiol. Biochem. 2017, 43, 1131–1141. [Google Scholar] [CrossRef]
- Ma, F.; Luo, L. Genome-wide identification of hsp70/110 genes in rainbow trout and their regulated expression in response to heat stress. PeerJ 2020, 8, e10022. [Google Scholar] [CrossRef]
- Chowdary, T.K.; Raman, B.; Ramakrishna, T.; Rao, C.M. Mammalian hsp22 is a heat-inducible small heat-shock protein with chaperone-like activity. Biochem. J. 2004, 381, 379–387. [Google Scholar] [CrossRef]
- Gober, M.D.; Smith, C.C.; Ueda, K.; Toretsky, J.A.; Aurelian, L. Forced expression of the h11 heat shock protein can be regulated by DNA methylation and trigger apoptosis in human cells. J. Biol. Chem. 2003, 278, 37600–37609. [Google Scholar] [CrossRef]
- Huang, J.; Li, Y.; Liu, Z.; Kang, Y.; Wang, J. Transcriptomic responses to heat stress in rainbow trout Oncorhynchus mykiss head kidney. Fish Shellfish. Immunol. 2018, 82, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Luo, L.T.; Wang, Q. Hsp60/10 and shsp families of heat shock protein genes in rainbow trout (Oncorhynchus mykiss) and their expression under heat stress. Aquac. Int. 2022, 30, 1–18. [Google Scholar] [CrossRef]
- Rogon, C.; Ulbricht, A.; Hesse, M.; Alberti, S.; Vijayaraj, P.; Best, D.; Adams, I.R.; Magin, T.M.; Fleischmann, B.K.; Höhfeld, J. Hsp70-binding protein hspbp1 regulates chaperone expression at a posttranslational level and is essential for spermatogenesis. Mol. Biol. Cell 2014, 25, 2260–2271. [Google Scholar] [CrossRef]
- Wang, C.; Chen, X.; Dai, Y.; Zhang, Y.; Sun, Y.; Cui, X. Comparative transcriptome analysis of heat-induced domesticated zebrafish during gonadal differentiation. BMC Genom. Data 2022, 23, 39. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-H.; Yang, F.-F.; Liao, S.-A.; Miao, Y.-T.; Ye, C.-X.; Wang, A.-L.; Tan, J.-W.; Chen, X.-Y. High temperature induces apoptosis and oxidative stress in pufferfish (Takifugu obscurus) blood cells. J. Therm. Biol. 2015, 53, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Arama, E.; Bader, M.; Srivastava, M.; Bergmann, A.; Steller, H. The two Drosophila cytochrome c proteins can function in both respiration and caspase activation. EMBO J. 2006, 25, 232–243. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, X. Cytochrome c-mediated apoptosis. Annu. Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef]
- Vornanen, M.; Hassinen, M.; Koskinen, H.; Krasnov, A. Steady-state effects of temperature acclimation on the transcriptome of the rainbow trout heart. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2005, 289, R1177–R1184. [Google Scholar] [CrossRef]
- Jiang, X.; Jiang, X.; Feng, Y.; Xu, R.; Wang, Q.; Deng, H. Proteomic analysis of eif5b silencing-modulated proteostasis. PLoS ONE 2016, 11, e0168387. [Google Scholar] [CrossRef]
- Krupovic, M.; Koonin, E.V. Multiple origins of viral capsid proteins from cellular ancestors. Proc. Natl. Acad. Sci. USA 2017, 114, E2401–E2410. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full Name of Gene | Abbreviated Name | Primer Sequences |
|---|---|---|
| cytochrome c’ | Cyt-c-p | F: CAAGGCGTCGGGTTATGTGT |
| R: AGGTAGGCGATGAGGTCTGC | ||
| Eukaryotic translation initiation factor 5 | eIF5 | F: CAAGGTAGAGGGCAAAGGCA |
| R: GGTCTCAGGGTTCTCGCACT | ||
| Apoptosis inhibitor 5 | api5-b | F: GCCCCCAAAGGAACGCT |
| R: ACGAGATGAGGCGATGAACC | ||
| Cytochrome c | CYC | F: CCAACAAGTCCAAGGGCATC |
| R: CGAGGTAGGCGATAAGGTCTG | ||
| Member of heat shock protein 60 family | Hsp60A | F: ACCCCAGCCAATGAAGACC |
| R: ATGATTCCCGCCTCCACAAG | ||
| Heat shock 70 kDa proteincognate 5 | Hsc70-5 | F: ATGCCCAAGGTGATTTCGCT |
| R: TCGTGTTCCTGTTGATGAGC | ||
| Lysosome-associated membrane glycoprotein 1 | Lamp1 | F: CATCTGCCGCTCCTCCATTAC |
| R: CCGACTGGTATGCTCCCGAC | ||
| Member of the transmembrane tetraspanin protein family | Tspan11 | ACGACATCTGGGAGGAAGC |
| ACATCCAGACGCCGACCA | ||
| beta-Actin | β-Actin | F: GGCTCGGGGATGGTCAA |
| R: CCAGTTGGTAATGATGCCGT |
| Number of Genes | BUSCO Categorization |
|---|---|
| 904 | Complete (C) |
| 810 | Complete and single copy (S) |
| 94 | Complete and duplicated (D) |
| 34 | Fragmented (F) |
| 40 | Missing (M) |
| 978 | Total BUSCO groups searched (n) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, T.; Li, T.; Yang, M.; Jiang, H.; Ling, J.; Gao, Q. Cardiac Transcriptome and Histology of the Heart of the Male Chinese Mitten Crab (Eriocheir sinensis) under High-Temperature Stress. Fishes 2024, 9, 92. https://doi.org/10.3390/fishes9030092
Pan T, Li T, Yang M, Jiang H, Ling J, Gao Q. Cardiac Transcriptome and Histology of the Heart of the Male Chinese Mitten Crab (Eriocheir sinensis) under High-Temperature Stress. Fishes. 2024; 9(3):92. https://doi.org/10.3390/fishes9030092
Chicago/Turabian StylePan, Tingshuang, Tong Li, Min Yang, He Jiang, Jun Ling, and Qian Gao. 2024. "Cardiac Transcriptome and Histology of the Heart of the Male Chinese Mitten Crab (Eriocheir sinensis) under High-Temperature Stress" Fishes 9, no. 3: 92. https://doi.org/10.3390/fishes9030092
APA StylePan, T., Li, T., Yang, M., Jiang, H., Ling, J., & Gao, Q. (2024). Cardiac Transcriptome and Histology of the Heart of the Male Chinese Mitten Crab (Eriocheir sinensis) under High-Temperature Stress. Fishes, 9(3), 92. https://doi.org/10.3390/fishes9030092