
Southern Horse Mackerel (Trachurus trachurus) Spatio-Temporal Distribution Patterns Based on Fine-Scale Resolution Data


Abstract
1. Introduction
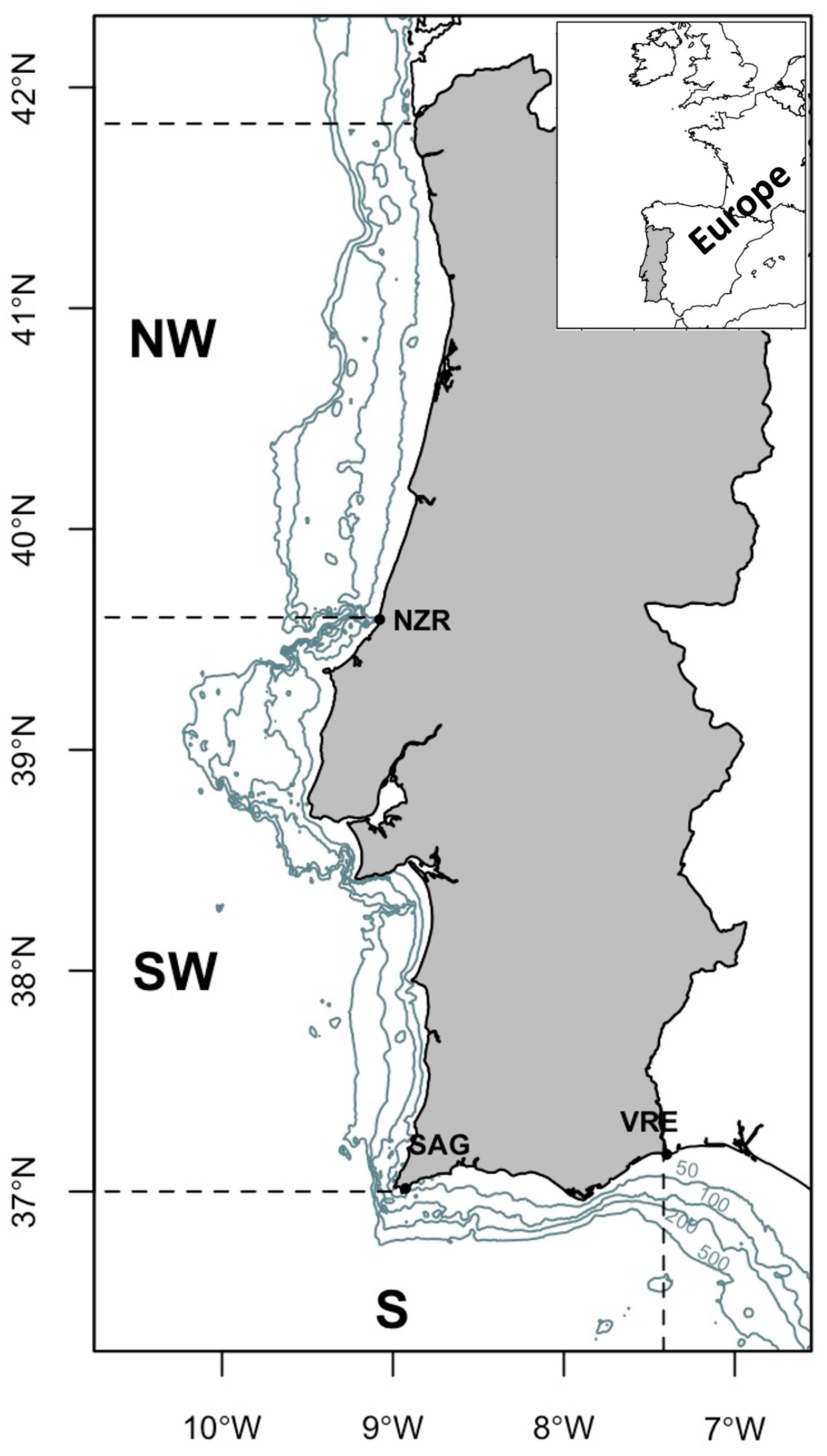
2. Materials and Methods
2.1. Data Exploration
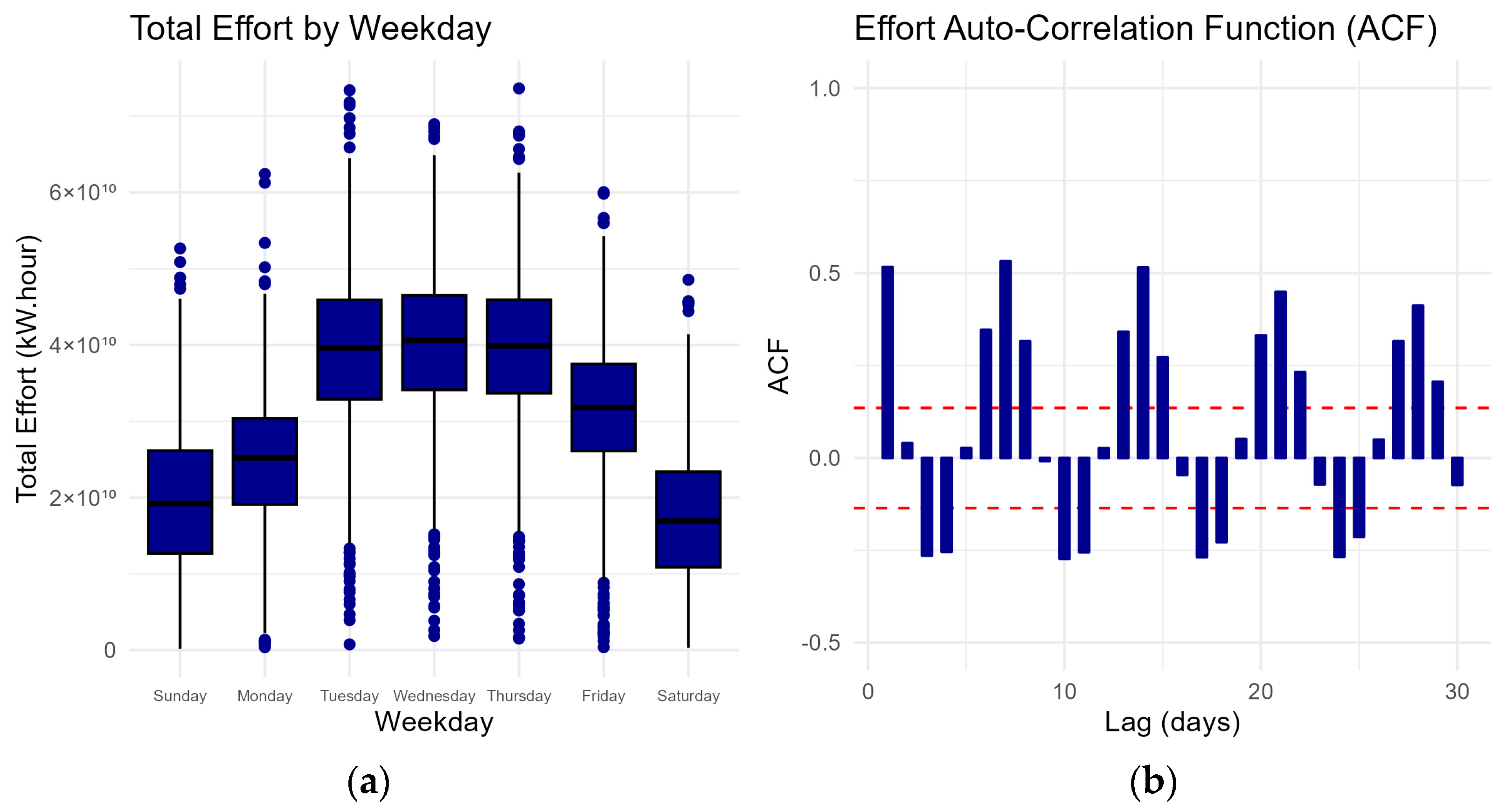
2.2. Effort Data Distribution
2.3. Statistical Analysis
3. Results
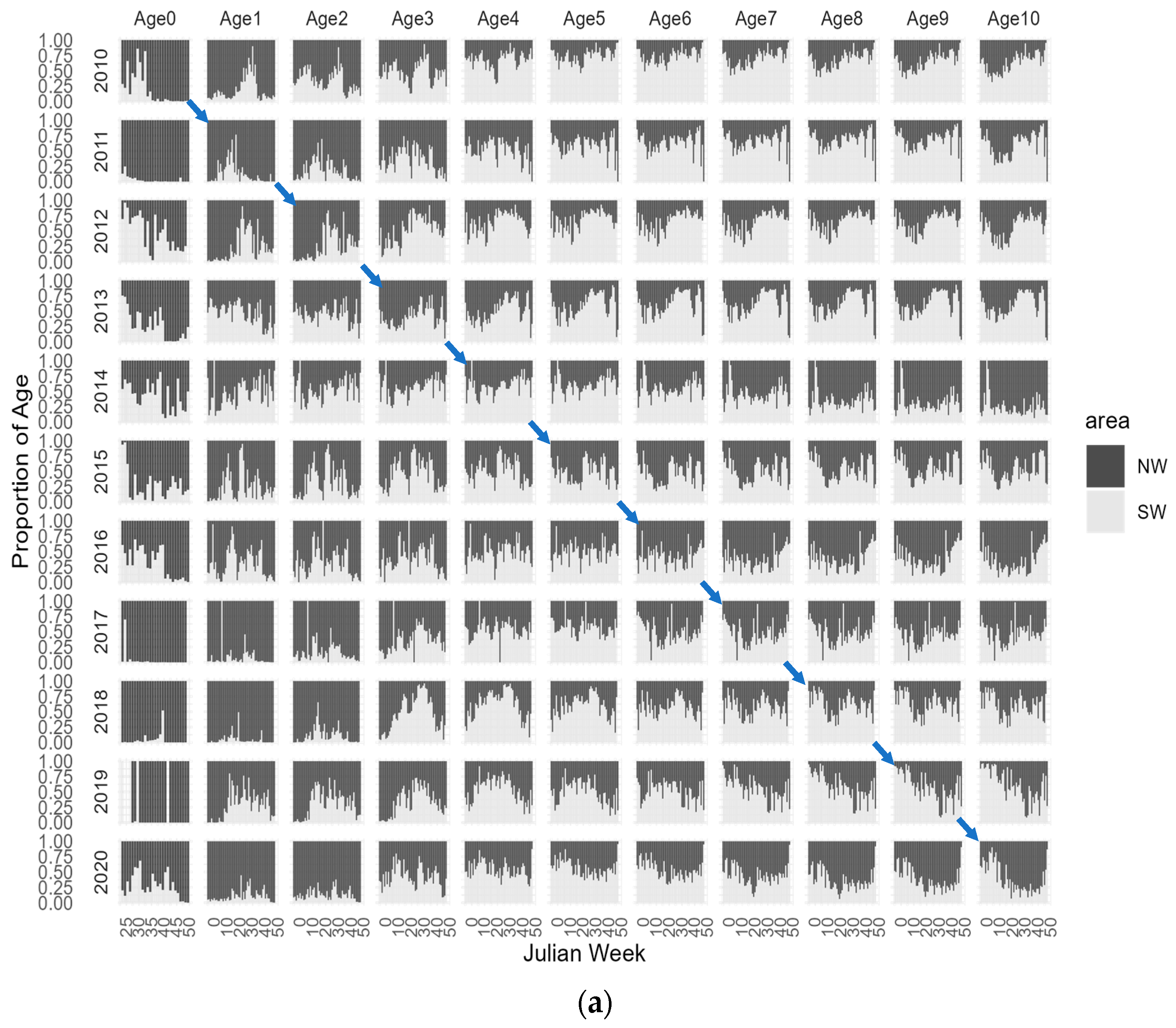
3.1. CPUE-at-Age Distribution
3.2. Model Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Núñez Riboni, I.; Akimova, A.; Sell, A.F. Effect of data spatial scale on the performance of fish habitat models. Fish Fish. 2021, 22, 955–973. [Google Scholar] [CrossRef]
- Zhou, X.; Ma, S.; Cai, Y.; Yu, J.; Chen, Z.; Fan, J. The Influence of Spatial and Temporal Scales on Fisheries Modeling-An Example of Sthenoteuthis oualaniensis in the Nansha Islands, South China Sea. J. Mar. Sci. Eng. 2022, 10, 1840. [Google Scholar] [CrossRef]
- Moura, T.; Chaves, C.; Figueiredo, I.; Mendes, H.; Moreno, A.; Silva, C.; Vasconcelos, R.P.; Azevedo, M. Assessing spatio-temporal changes in marine communities along the Portuguese continental shelf and upper slope based on 25 years of bottom trawl surveys. Mar. Environ. Res. 2020, 160, 105044. [Google Scholar] [CrossRef] [PubMed]
- Sousa, P.; Azevedo, M.; Gomes, M.C. Demersal assemblages off Portugal: Mapping, seasonal, and temporal patterns. Fish. Res. 2005, 75, 120–137. [Google Scholar] [CrossRef]
- Gomes, M.C.; Serrão, E.; Borges, M.F. Spatial patterns of groundfish assemblages on the continental shelf of Portugal. ICES J. Mar. Sci. 2001, 58, 633–647. [Google Scholar] [CrossRef]
- Whitehead, P.J.P.; Bauchot, M.L.; Hureau, J.C.; Nielsen, J.; Tortonese, E. Fishes of the North-Eastern Atlantic and the Mediterranean; UNESCO: Paris, France, 1986; Volume 2. [Google Scholar]
- Abaunza, P.; Santos, M.B.; Murta, A.G.; Cimmaruta, R.; Cariani, A.; Tinti, F.; Deflorio, M. Stock identity of horse mackerel (Trachurus trachurus) in the Northeast Atlantic and Mediterranean Sea: Integrating the results from different stock identification approaches. Fish. Res. 2008, 89, 196–209. [Google Scholar] [CrossRef]
- Murta, A.G.; Abaunza, P.; Cardador, F.; Sánchez, F. Ontogenic migrations of horse mackerel along the Iberian coast. Fish. Res. 2008, 89, 186–195. [Google Scholar] [CrossRef]
- ICES. Report of the Benchmark Workshop on Pelagic Stocks (WKPELA), 6–10 February 2017, Lisbon, Portugal; ICES CM 2017/ACOM:35; ICES: Copenhagen, Denmark, 2017; p. 278. [Google Scholar]
- Relvas, P.; Barton, E.D.; Dubert, J.; Oliveira, P.B.; Peliz, A.; Da Silva, J.C.B.; Santos, A.M.P. Physical oceanography of the western Iberia ecosystem: Latest views and challenges. Prog. Oceanogr. 2007, 74, 149–173. [Google Scholar] [CrossRef]
- Ministério do Mar, República Portuguesa. Reavaliação do Estado Ambiental e Definição de Metas: Parte D, Subdivisão do Continente. In Estratégia Marinha, Relatório do 2º Ciclo; Ministério do Mar, República Portuguesa: Algés, Portugal, 2020; p. 458. [Google Scholar]
- Murta, A.G.; Borges, M.F. Factors Affecting the Abundance Distribution of Horse Mackerel Trachurus trachurus (Linnaeus, 1758) in Portuguese Waters; ICES CM 1994/H:20; ICES: Copenhagen, Denmark, 1994. [Google Scholar]
- Borges, M.F.; Gordo, L.S. Spatial Distribution by Season and Some Biological Parameters of Horse Mackerel (Trachurus trachurus L.) in the Portuguese Continental Waters (Division IXa); ICES CM 1991/H:54; ICES: Copenhagen, Denmark, 1991. [Google Scholar]
- EC. Commission Regulation (EC) No. 1489/97 of 29 July 1997 laying down detailed rules for the application of Council Regulation (EEC) No. 2847/93 as regards satellite-based vessel monitoring systems. Off. J. Eur. Union 1997, L202, 18–23. [Google Scholar]
- EC. Commission Regulation (EC) No. 2244/2003 of 18 December 2003 laying down detailed provisions regarding satellite based vessel monitoring systems. Off. J. Eur. Union 2003, L333, 17–27. [Google Scholar]
- Gerritsen, H.; Lordan, C. Integrating vessel monitoring systems (VMS) data with daily catch data from logbooks to explore the spatial distribution of catch and effort at high resolution. ICES J. Mar. Sci. 2010, 68, 245–252. [Google Scholar] [CrossRef]
- Bastardie, F.; Nielsen, J.R.; Ulrich, C.; Egekvist, J.; Degel, H. Detailed mapping of fishing effort and landings by coupling fishing logbooks with satellite-recorded vessel geo-location. Fish. Res. 2010, 106, 41–53. [Google Scholar] [CrossRef]
- Palmer, M.C.; Wigley, S.E. Using Positional Data from Vessel Monitoring Systems to Validate the Logbook-Reported Area Fished and the Stock Allocation of Commercial Fisheries Landings. N. Am. J. Fish. Manag. 2009, 29, 928–942. [Google Scholar] [CrossRef]
- Chang, S.; Yuan, T. Deriving high-resolution spatiotemporal fishing effort of large-scale longline fishery from vessel monitoring system (VMS) data and validated by observer data. Can. J. Fish. Aquat. Sci. 2014, 71, 1363–1370. [Google Scholar] [CrossRef]
- Bez, N.; Walker, E.; Gaertner, D.; Rivoirard, J.; Gaspar, P. Fishing activity of tuna purse seiners estimated from vessel monitoring system (VMS) data. Can. J. Fish. Aquat. Sci. 2011, 68, 1998–2010. [Google Scholar] [CrossRef]
- Watson, J.T.; Haynie, A.C.; Sullivan, P.J.; Perruso, L.; O’Farrell, S.; Sanchirico, J.N.; Mueter, F.J. Vessel monitoring systems (VMS) reveal an increase in fishing efficiency following regulatory changes in a demersal longline fishery. Fish. Res. 2018, 207, 85–94. [Google Scholar] [CrossRef]
- Enguehard, R.A.; Devillers, R.; Hoeber, O. Comparing interactive and automated mapping systems for supporting fisheries enforcement activities—A case study on vessel monitoring systems (VMS). J. Coast. Conserv. 2012, 17, 105–119. [Google Scholar] [CrossRef]
- Lee, J.; South, A.; Jennings, S. Developing reliable, repeatable and accessible methods to provide high-resolution estimates of fishing-effort distributions from vessel monitoring system (VMS) data. ICES J. Mar. Sci. 2010, 67, 1260–1271. [Google Scholar] [CrossRef]
- Azevedo, M.; Silva, C. A framework to investigate fishery dynamics and species size and age spatio-temporal distribution patterns based on daily resolution data: A case study using Northeast Atlantic horse mackerel. ICES J. Mar. Sci. 2020, 77, 2933–2944. [Google Scholar] [CrossRef]
- EC. Council Regulation (EC) No. 2406/96 of 23 December 1996 laying down common marketing standards for certain fishery products. Off. J. Eur. Union 1996, L334, 1–15. [Google Scholar]
- Azevedo, M.; Silva, C.; Vølstad, J.H. Onshore biological sampling of landings by species and size category within auction sites can be more efficient than trip-based concurrent sampling. ICES J. Mar. Sci. 2021, 78, 2757–2773. [Google Scholar] [CrossRef]
- ICES. Working Group on Southern Horse Mackerel, Anchovy and Sardine (WGHANSA). ICES Sci. Rep. 2022, 4, 518. [Google Scholar] [CrossRef]
- Afonso-Dias, M.; Pinto, C. Análise da Distribuição Espacial do Esforço e Rendimentos de Pesca das Frotas Portuguesas de Arrasto Costeiro. Projecto GeoPesca. Relatório Final Projecto MARE 22-05-01-00025. 2008. Available online: http://w3.ualg.pt/madias/geopesca/GeoPescasRelatorioFinal08.pdf (accessed on 15 November 2021).
- EC. Council Regulation (EC) No. 2017/1004 of 17 May, on the Establishment of a Union Framework for the Collection, Management and Use of Data in the Fisheries Sector and Support for Scientific Advice Regarding the Common Fisheries Policy and Repealing Council Regulation (EC) No. 199/2008; European Council: Brussels, Belgium, 2017. [Google Scholar]
- ICES. Workshop on Age Reading of Horse Mackerel, Mediterranean Horse Mackerel and Blue Jack Mackerel (Trachurus trachurus, T. mediterraneus and T. picturatus) (WKARHOM3), 5–9 November 2018, Livorno, Italy; ICES CM 2018/EOSG:28; ICES: Copenhagen, Denmark, 2018; p. 186. [Google Scholar] [CrossRef]
- Panfili, J.; Troadec, H.; Pontual, H.D.; Wright, P.J. Manual of Fish Sclerochronology; Ifremer-lRD Coedition: Brest, France, 2002; p. 464. [Google Scholar]
- Williams, T.; Bedford, B.C. The use of otoliths for age determination. In Ageing of Fish: Proceedings of an International Symposium at University of Reading, 19–20 July 1973; Bagenal, T.B., Ed.; Unwin Brothers: Surrey, UK, 1974; pp. 114–123. [Google Scholar]
- Maunder, M.N.; Thorson, J.T.; Xu, H.; Oliveros-Ramos, R.; Hoyle, S.D.; Tremblay-Boyer, L.; Lee, H.H.; Kai, M.; Chang, S.-K.; Kitakado, T.; et al. The need for spatio-temporal modeling to determine catch-per-unit effort based indices of abundance and associated composition data for inclusion in stock assessment models. Fish. Res. 2020, 229, 105594. [Google Scholar] [CrossRef]
- Thorson, J.T.; Barnett, L.A.K. Comparing estimates of abundance trends and distribution shifts using single- and multispecies models of fishes and biogenic habitat. ICES J. Mar. Sci. 2017, 74, 1311–1321. [Google Scholar] [CrossRef]
- Azevedo, M.; Mendes, H.; Costas, G.; Jardim, E.; Mosqueira, I.; Scott, F. Report on the Assessment of a Long-term Management Strategy for Southern Horse Mackerel (hom27.9a), 15–16 February 2018; ICES CM 2018/ACOM:42; ICES: Copenhagen, Denmark, 2018; p. 36. [Google Scholar]
- Rindorf, A.; Lewy, P. Analyses of length and age distributions using continuation-ratio logits. Can. J. Fish. Aquat. Sci. 2001, 58, 1141–1152. [Google Scholar] [CrossRef]
- Agresti, A. Categorical data analysis. In Wiley Series in Probability and Statistics; Wiley: Hoboken, NJ, USA, 2002. [Google Scholar] [CrossRef]
- Kvist, T.; Gislason, H.; Thyregod, P. Using continuation-ratio logits to analyze the variation of the age composition of fish catches. J. Appl. Stat. 2000, 27, 303–319. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017. [Google Scholar] [CrossRef]
- Yee, T.W. Vector Generalized Linear and Additive Models: With an Implementation in R; Springer Series in Statistics; Springer: New York, NY, USA, 2015; ISBN 978-1-4939-2818-7. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 10 January 2024).
- Bivand, R.; Pebesma, E.; Gomez-Rubio, V. Applied Spatial Data Analysis with R. Use R! Series, 2nd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-7618-4. Available online: https://asdar-book.org/ (accessed on 15 November 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 10 January 2024).
- Bauldry, S.; Xu, J.; Fullerton, A.S. GenCRM: A new command for Generalized Continuation-Ratio Models. Stata J. 2018, 18, 924–936. [Google Scholar] [CrossRef]
- Ivy, L.; Agresti, A. The analysis of ordered categorical data: An overview and a survey of recent developments. TEST 2005, 14, 1–73. [Google Scholar]
- Cabral, H.N.; Murta, A. The diet of blue whiting, hake, horse mackerel and mackerel off Portugal. J. Appl. Ichthyol. 2002, 18, 14–23. [Google Scholar] [CrossRef]
- Villamor, B.; Abaunza, P.; Lucio, P.; Porteiro, C. Distribution and age structure of mackerel (Scomber scombrus, L.) and horse mackerel (Trachurus trachurus, L.) in the northern coast of Spain, 1989–1994. Sci. Mar. 1997, 61, 345–366. Available online: http://www.icm.csic.es/scimar/pdf/61/sm61n3345.pdf (accessed on 15 November 2021).
- Gonçalves, P.; Costa, A.M.; Murta, A.G. Estimates of batch fecundity and spawning fraction for the southern stock of horse mackerel (Trachurus trachurus) in ICES Division IXa. ICES J. Mar. Sci. 2009, 66, 617–622. [Google Scholar] [CrossRef]
- Hočevar, S.; Hutchings, J.A.; Kuparinen, A. Multiple-batch spawning: A risk-spreading strategy disarmed by highly intensive size-selective fishing rate. Proc. R. Soc. B Biol. Sci. 2022, 289, 20221172. [Google Scholar] [CrossRef] [PubMed]
- Chust, G.; Taboada, F.G.; Álvarez, P.; Ibaibarriaga, L. Species acclimatization pathways: Latitudinal shifts and timing adjustments to track ocean warming. Ecol. Indic. 2023, 146, 109752. [Google Scholar] [CrossRef]
- Lavín, A.; Moreno-Ventas, X.; De Zárate, V.O.; Abaunza, P.; Cabanas, J.M.O. Environmental variability in the North Atlantic and Iberian waters and its influence on horse mackerel (Trachurus trachurus) and albacore (Thunnus alalunga) dynamics. ICES J. Mar. Sci. 2007, 64, 425–438. [Google Scholar] [CrossRef][Green Version]
- Santos, A.M.P.; Borges, M.F.; Groom, S. Sardine and horse mackerel recruitment and upwelling off Portugal. ICES J. Mar. Sci. 2001, 58, 589–596. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Time Aggregation | Spatial Aggregation | Basis |
|---|---|---|---|
| Catch-at-length (number) | Daily | 0.05° × 0.05° | Landings (weight) by trip recorded at auction by size category × length distribution by size category |
| Catch-at-age (number) | Catch-at-length by trip × semester Age-Length Keys by year | ||
| Effort (kW × hour) | VMS data (trawl hours by fishing position) × vessel power information (from EU Fleet Register) | ||
| Depth (meters) | Satellite Global Topography |
| Year | Number of Vessels | Total Number of Trips | Total Trawl Hours | Average Engine Power (kW) | Average Depth (m) |
|---|---|---|---|---|---|
| 2010 | 38 | 4352 | 53,683.1 | 538.9 | 97.6 |
| 2011 | 35 | 3842 | 52,479.5 | 529.1 | 99 |
| 2012 | 37 | 4412 | 54,449.0 | 529.3 | 107.5 |
| 2013 | 34 | 4093 | 46,158.2 | 523.8 | 114.1 |
| 2014 | 36 | 4279 | 53,240.3 | 526.8 | 119 |
| 2015 | 43 | 4537 | 59,361.8 | 519.6 | 111.8 |
| 2016 | 43 | 4778 | 52,682.5 | 516.7 | 114.4 |
| 2017 | 39 | 4577 | 61,092.6 | 515.3 | 110.9 |
| 2018 | 42 | 4511 | 64,709.4 | 514.1 | 102.3 |
| 2019 | 43 | 4721 | 65,473.4 | 495.5 | 94.9 |
| 2020 | 42 | 4736 | 68,020.0 | 465.1 | 100.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendes, H.; Silva, C.; Azevedo, M. Southern Horse Mackerel (Trachurus trachurus) Spatio-Temporal Distribution Patterns Based on Fine-Scale Resolution Data. Fishes 2024, 9, 93. https://doi.org/10.3390/fishes9030093
Mendes H, Silva C, Azevedo M. Southern Horse Mackerel (Trachurus trachurus) Spatio-Temporal Distribution Patterns Based on Fine-Scale Resolution Data. Fishes. 2024; 9(3):93. https://doi.org/10.3390/fishes9030093
Chicago/Turabian StyleMendes, Hugo, Cristina Silva, and Manuela Azevedo. 2024. "Southern Horse Mackerel (Trachurus trachurus) Spatio-Temporal Distribution Patterns Based on Fine-Scale Resolution Data" Fishes 9, no. 3: 93. https://doi.org/10.3390/fishes9030093
APA StyleMendes, H., Silva, C., & Azevedo, M. (2024). Southern Horse Mackerel (Trachurus trachurus) Spatio-Temporal Distribution Patterns Based on Fine-Scale Resolution Data. Fishes, 9(3), 93. https://doi.org/10.3390/fishes9030093