Abstract
Aquaculture of marine ornamental fish could potentially reduce the fishing pressure on wild stocks by the global aquarium trade, but its expansion is often constrained by the limited understanding on the biology and early life history of candidate species. The orchid dottyback Pseudochromis fridmani is a valuable and popular marine ornamental fish, but scientific reports on its baseline biology, especially the egg filial cannibalistic behaviour, are scarce. The present study documented key aspects of reproductive biology and early ontogeny of P. fridmani in captivity, including reproductive behaviour, patterns of spawning and filial cannibalism throughout a 12-month period by seven pairs of brood fish, as well as the embryonic and larval development. The results showed that the captive spawning of the broodstock pairs generally occurred every 5–11 days, most commonly every 6–8 days, throughout a year. Despite feeding the brood fish to satiation throughout the data collection period, the average monthly rate of egg filial cannibalism reached 55 ± 37%, but the cannibalism frequency appeared to be pair-specific. The egg incubation duration was approximately 96 h at 27 ± 1 °C, and the development of embryos from cleavage, blastula, gastrula, and segmentation to the pharyngula stage is herein described in detail. The newly hatched P. fridmani larvae possessed important structures and organs for first feeding, including pigmented eyes, developed jaws, and a straight-tube gut. The 0 to 14 days post-hatching (DPH) period appeared to be an important larval stage, as P. fridmani larvae were observed to complete major changes in morphology, gut development, and phototactic behaviour by 14 DPH. Under the culture conditions of this study, the earliest transition to the juvenile stage was observed on 31 DPH, and the majority of fish became juveniles by 56 DPH. The results of this study inform baseline aquaculture production protocols and direct future research, particularly to reduce filial cannibalism through broodstock management and to improve larval culture through supporting the early ontogenetic development of P. fridmani. Additionally, these findings form a foundation for further studying the biology and ecology of P. fridmani in the wild.
Keywords:
captive breeding; spawning interval and behaviour; early ontogeny; larval rearing; marine ornamental fish Key Contribution:
To fill the knowledge gaps of the biology of Pseudochromidae, this study documented the reproductive events in captivity over a 12-month period as well as detailed the embryonic and larval development of orchid dottyback Pseudochromis fridmani. The outcomes facilitate a better understanding of the reproduction and early ontogeny of P. fridmani and serve as essential foundations not only for optimizing breeding techniques for dottyback species but also studying their ecology and population dynamics in the wild.
1. Introduction
There are at least 154 dottyback species (family Pseudochromidae) in the Indo-Pacific region grouped into 24 genera [1,2], and new species are still being described in recent years [3,4]. Dottybacks generally have an elongated body with bright colouration [5,6]. In some dottyback species, the coloration is sexually dimorphic [7], and they often display territorial and aggressive behaviour, particularly toward co-specifics [5,6,8]. In fact, due to their vibrant coloration, dottybacks are among the most popular ornamental fish families traded globally [9,10].
The orchid dottyback Pseudochromis fridmani (Klausewitz, 1968) is an endemic species in the Red Sea [11,12], and it is considered as one of the most commercially attractive dottyback species by aquarium hobbyists [10]. P. fridmani are commonly found near crevices or underneath overhangs in coral reef habitat below 10 m, but their distribution can range from 1 to 60 m [8,11,12]. In the wild, P. fridmani usually grow up to 6–7 cm [11,12] but in captivity can reach maximum 9–10 cm (pers. obs.). Two most iconic morphological features of this species are the vibrant metallic purple colouration throughout the body and the short black stripe across each eye. Interestingly, depending on the dietary nutrition, the intensity of the purple colouration of captive-bred P. fridmani can vary substantially among individuals [13]. Besides the attractive morphology, the relatively less aggressive nature of P. fridmani compared to other species of dottyback has also gained them popularity among aquarium hobbyists [6,8,10], driving the retail price to around USD 60 per fish in the aquarium trade market [14].
The global aquarium trade is a multi-billion dollar industry, in which around 90% of the marine fish sold are still collected from the wild, mainly the tropical coral reefs [15,16]. Hence, the development of marine ornamental aquaculture is driven by ecological, conservational, and socioeconomic imperatives with the aim to alleviate fishing pressure on wild stocks by supplying sufficient captive-bred fish to the trade [15,17]. The baseline knowledge of relevant reproductive biology and ontogenetic development is critical for achieving successful captive production of candidate species [18,19,20,21] as well as for studying their ecology and population dynamics in the wild. Nonetheless, spawning and early development of reef fish are often difficult to observe during field studies. As a result, relevant information is limited in the literature for most coral reef fish, which hinders the breeding breakthroughs.
Similarly, scientific data on the baseline biology of P. fridmani remains relatively limited. In the literature, field study was limited to report the species’ distribution, habitat, and morphology [8]. In a captive environment, Mies et al. [22] observed the spawning behaviour and patterns of seven dottyback species for 8 months and reported that the three P. fridmani pairs spawned every 5 to 9 days at 27 °C. Additionally, the authors found that the P. fridmani pairs often cannibalized their own eggs before the first successful hatching occurred, but this filial cannibalism pattern was not quantified over the 8-month period. To date, relevant research data on filial cannibalism of P. fridmani remain scarce, even though this cannibalistic behaviour is considered as an aquaculture bottleneck for P. fridmani as well as many other dottyback species [22,23]. For the early ontogeny of P. fridmani, Chen et al. [24] and Wittenrich and Turingan [25] documented the development of the digestive system and feeding apparatus of P. fridmani, respectively, while other recent studies focused on optimizing the larval-rearing techniques [24,26,27]. In general, most information on reproductive biology and the embryonic and larval developmental features of P. fridmani is reported by aquarium hobbyists [10,28]. Compared with the scientific data on the embryonic and larval development of other marine ornamental fishes [20,29,30,31], hobbyists’ documentation on P. fridmani lacks detailed morphological description as well as high-quality photographic detail at important developmental stages.
Therefore, the objectives of the present study were to fill these existing knowledge gaps by documenting relevant aspects of reproductive biology and the embryonic and larval development of P. fridmani. The spawning and filial cannibalism patterns of multiple P. fridmani pairs were documented throughout a full year in order to establish a baseline reproduction dynamic over time for this species. The insights gained serve the purposes of not only increased knowledge of fish biology and ecology but also improved aquaculture production protocols for P. fridmani, through informed description of broodstock management and assessment of larval development during rearing.
2. Materials and Methods
2.1. Brood Fish and Pair Formation
All orchid dottyback P. fridmani broodstock used in this study were more than 8 months old and captive-bred at the Marine Aquaculture Research Facility Unit (MARFU) of James Cook University, Townsville, Australia. Since P. fridmani lack conspicuous sexual dimorphic morphology, after the fish reached a total length between 50 and 70 mm, they were randomly paired; the pair bonding was considered completed when natural spawning occurred. In this study, each of the seven P. fridmani broodstock pairs was kept in a 100 L columnar fiberglass tank connected to a same recirculating system with 25 μm filtered seawater. All tanks were located outdoors under a semi-transparent roof to provide a natural photoperiod (March to August: 11–12 h daylight hours; September to February: 12–13 h). Within each tank, a capped PVC pipe (40 mm diameter, 20 cm in length) was provided as spawning site. A broodstock diet was prepared by blending a mix of seafood and nutritional supplements as described in Zeng et al. [32]. The broodstock pairs were fed twice a day (9–10 a.m. and 4–5 p.m.) until they reached apparent satiation (i.e., until no signs of feeding); before feeding ended, some excess food was also left in the tanks to ensure sufficient food supply for the pairs. Throughout the 12-month duration of this study, the seawater of the broodstock system was maintained at 27.5–29 °C, salinity 30–36 ppt, pH 8.0–8.3, total ammonia and nitrite < 0.2 ppm, and nitrate < 50 ppm.
2.2. Reproductive Behaviour, Spawning, and Filial Cannibalism
The reproductive activity of P. fridmani pairs was recorded by GoPro® HERO 7 cameras (GoPro, Inc., San Mateo, CA, USA). Considering the potential behavioural differences among pairs, the footage was recorded from three pairs of P. fridmani during spawning, egg incubation, and the period in-between, respectively. The spawning frequency, egg incubation duration, and incidences of filial cannibalism of seven P. fridmani pairs were documented from 1 February 2020 to 31 January 2021. During the 12-month period, only one fish was found dead, but the remaining fish was soon paired with a new P. fridmani adult to continue the experiment. For data collection, prior to daily morning feeding, the PVC pipe in each tank was checked for the presence of deposited eggs. Notes were also made when a significant amount or all the eggs disappeared during the incubation period (filial cannibalism). The rate of filial cannibalism (FC%) represented the monthly frequency of egg batches that were partially or completely cannibalized by the brood fish during the incubation period. FC was calculated as below:
Additionally, to understand when embryos were eaten, the timing of filial cannibalism, indicated by incidence (%) on a given day, within the 4-day egg incubation period was calculated for each pair.
2.3. Embryonic Development
To describe the embryonic development, two fertilized egg batches were sampled to cover the incubation period from 1 h post fertilization (PF) until 96 h PF (batch 1 from pair E: 1–48 h PF; batch 2 from pair C: 48–96 h PF). During incubation, approximately 300 embryos from each egg clutch were sampled and kept in seawater at 27 ± 1 °C, salinity 30 ppt, and under a photoperiod of 12 h L:12 h D in a 15 L cylindrical tank. The P. fridmani embryos were photographed under a dissection microscope (Olympus SZ61, Olympus, Tokyo, Japan) equipped with camera (Olympus DP26, Olympus, Tokyo, Japan). The sampling and observation of embryos occurred every 30 min between 1–5 h PF and at 5, 6, 7, 8, 9, 10, 12, 14, 16, 18, 20, 22, 24, 48, and 96 h PF. The embryonic development of P. fridmani was described based on the photographs taken, using stage identification and terminology from Kimmel et al. [33] for zebrafish Danio rerio.
2.4. Larval Development
2.4.1. Larval Rearing
The live prey super-small (ss)-type rotifers Brachionus rotundiformis were cultured on concentrated algae paste (RotiGrow®, Reed Mariculture Inc., Campbell, CA, USA), while the copepod Parvocalanus crassirostris were fed daily with live microalgae Isochrysis galbana (Tahitian strain, T-ISO), with all life stages collected upon harvesting to feed the larvae. The Artemia nauplii (Great Salt Lake strain, INVE Technologies, Dendermonde, Belgium) were hatched daily and enriched with S.Presso (SELCO®, INVE Technologies, Dendermonde, Belgium).
The newly hatched P. fridmani larvae were cultured in a cylindroconical 100 L fiberglass tank with black walls, connecting to a recirculation system with 25 μm filtration. From 0 to 12 days post hatching (DPH), larvae were fed a mix of ss-type rotifer B. rotundiformis (5–20 ind. mL−1) and the copepod P. crassirostris (0.5–2 ind. mL−1), and the prey densities were checked and replenished every morning. The “greenwater” condition was used during this early period of larval rearing [26]. Between 9–12 DPH, Artemia nauplii (0.1–1 ind. mL−1) were also added to feed the larvae. On 13 DPH, the larvae were solely fed 1 Artemia nauplii mL−1, but from 14 to 19 DPH, both Artemia nauplii and enriched metanauplii were fed to larvae at a combined density of 1–1.5 ind. mL−1. Between 20–40 DPH, the larvae were solely fed enriched Artemia at an increasing density (1.5–3 ind. mL−1), which were reduced to 1.5 ind. mL−1 from 41 DPH onwards. From 28 DPH onwards, formulated feed (O.range START-L, 150–315 μm, INVE Ltd., Nonthaburi, Thailand) was fed to the larvae twice daily with increasing quantity. Throughout larval rearing, the seawater was maintained at 27.5–28.5 °C, salinity 33.5–35 ppt, pH 8.0–8.2, total ammonia and nitrite < 0.2 ppm, and nitrate < 50 ppm. In addition, gradual sweater changes were achieved by maintaining a gentle and slow flow of fresh seawater into the larval-rearing tank. Fluorescent lamps were installed 2 m above the larval rearing tank to provide constant light (24 h L:0 h D).
P. fridmani became juveniles when the fish had completed flexion of the notochord and obtained typical adult-type appearance with purple coloration across the body and a black stripe diagonally across each eye. On 56 DPH, all surviving larvae and juveniles were counted to confirm the proportion of fish that had become juveniles.
2.4.2. Larvae Sampling
To document the larval development, P. fridmani larvae were randomly sampled in the morning on 0, 2, 4, 7, 10, 13, 16, 19, 22, 25, 28, 25, 38, and 40 DPH (0–2 DPH, n = 15; 4–38 DPH, n = 9; 40 DPH, n = 1). Immediately after being euthanized with anaesthetic (AQUI-S®, AQUI-S New Zealand Ltd., Melling, New Zealand), the larvae were photographed under a stereomicroscope (Olympus SZ61, Olympus, Tokyo, Japan) with a mounted camera (Olympus DP26, Olympus, Tokyo, Japan). The larval morphological development was described based on the photographs taken, while the notochord length (NL: the length between the snout tip and the notochord end) and body depth (BD: the distance from dorsal body muscle edge to anus) were measured on images using the software cellSens Standard 1.7.
3. Results
3.1. Reproductive Behaviour of Brood Pairs
Based on the video footage, although P. fridmani did not display conspicuous sexual dimorphism, the females generally had a shorter and wider body compared to the males, and their abdomens became swollen 1–2 days prior to spawning. While the males usually stayed inside the PVC pipe, the females swam in open areas and only entered the pipe in response to a potential threat or for spawning. On the day of spawning, the male initiated courtship by erecting the dorsal fins in front of the female and vigorously shaking the head and the tail from side to side before swimming back inside the PVC pipe soon after. This courting ritual was repeated many times until the female eventually followed the male into the pipe to lay eggs inside. The spawning usually started between 1–3 pm and lasted for 1–2 h; it was considered ended when either the male aggressively chased the female out of the pipe or when the female voluntarily left the spawning site. Notably, within 1–2 h after spawning, the males from all three broodstock pairs were recorded to attempt to court with the females again; however, the females did not appear to show any behavioural response in return. During the egg incubation period, the female freely swam in open areas of the tank as usual, while the male spent most of the time inside the pipe guarding the egg mass. Occasionally, the male appeared to defend the fertilized egg mass by chasing the female away when she was too close to the PVC pipe.
3.2. Captive Spawning and Filial Cannibalism
All seven pairs of P. fridmani naturally spawned during the one-year monitoring period (Figure 1). The spawning frequency of all pairs was relatively similar: each pair generally spawned every 5 to 11 days, and more commonly every 6 to 8 days. For all pairs, the number of spawn events per month ranged from one to five and was commonly between three and four times (3.7 ± 1.1 monthly spawns over all pairs, Table 1). An interrupted spawning period of over one month was observed from pair B (mid-February–March 2020) and pair F (July–August 2020), but both pairs resumed spawning within two months (Figure 1). The original male of pair D was found dead on 14 October 2020 and was replaced by a new male the next day. The new pair D readily started spawning on 18 October, and the spawning continued until the end of this study (Figure 1).
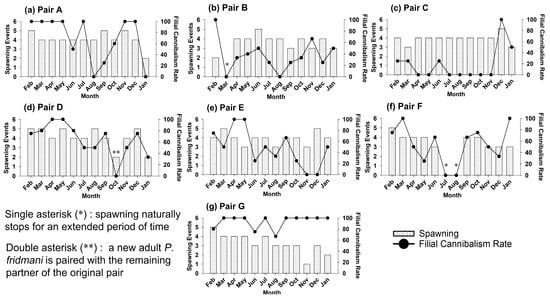
Figure 1.
The number of spawning events and the filial cannibalism rate (%) of seven broodstock pairs (a–g) of Pseudochromis fridmani recorded each month over one year (February 2020 to January 2021).

Table 1.
The average monthly spawning events and filial cannibalism rates of Pseudochromis fridmani pairs over one year (from February 2020 to January 2021). Data are expressed as mean ± SD.
The results also showed that cannibalism of the egg mass or filial cannibalism by the P. fridmani brood fish was very common throughout the study but highly variable among pairs (Table 1, Figure 1). By the end of the 12-month period, only three out of seven broodstock pairs maintained a mean filial cannibalism rate (FC) below 50% (pair C: 19 ± 30%, pair B: 37 ± 28%, and pair E: 48 ± 34%; Table 1). The mean FC of the other three pairs was between 54 and 70%, while pair G had the highest FC of 94 ± 12%. It is worth noting that within the same pair, the FC rates also fluctuated from month to month, and it did not appear to correlate with season change or monthly spawning frequencies (Figure 1).
Overall, filial cannibalism most commonly occurred on day 3 or 4 post fertilization (DPF; Figure 2). The pooled data of all pairs showed that on 2 DPF, incidence of filial cannibalism was only 5 ± 9%, but it increased sharply to 35 ± 17%, and 60 ± 20% on 3 and 4 DPF, respectively. Notably, among the seven pairs, only pairs F and G were found sometimes cannibalizing their eggs on 2 DPF. The incidence of filial cannibalism on 3 and 4 DPF was relatively even for pair A and C, while for pairs B, D, and E, filial cannibalism predominately occurred on 4 DPF (Figure 2).
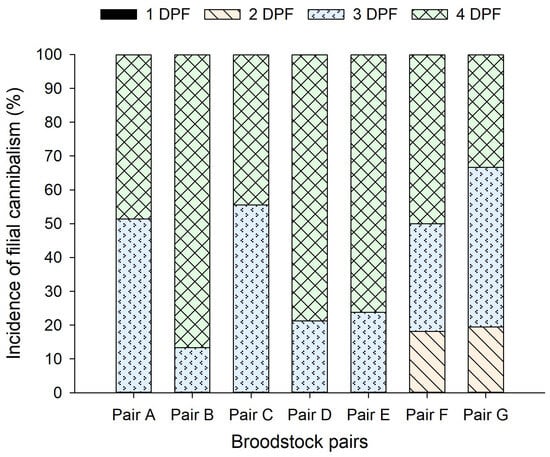
Figure 2.
The timing of filial cannibalism, indicated by incidence (%) on a given day, within the 4-day post-fertilization periods for each Pseudochromis fridmani broodstock pair over a 12-month period (from February 2020 to January 2021). DPF, day post fertilization.
3.3. Embryonic Development
The embryonic development of P. fridmani is summarized in Table 2. The newly fertilized P. fridmani eggs were demersal and spherical (average diameter: 1.086 ± 0.043 mm, n = 40). Under the microscope, it was observed that there were numerous thin and adhesive threads on the egg surface that held the egg mass together, and predominantly, a large single oil droplet was present on the yolk of each embryo (Figure 3a).
Table 2.
Embryonic development of Pseudochromis fridmani incubated at 27 ± 1 °C.
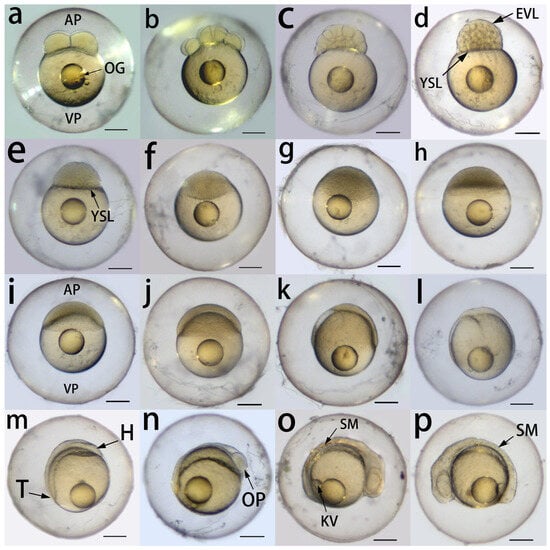
Figure 3.
Embryonic development of Pseudochromis fridmani from 1 to 22 h post fertilization (PF) at 27 ± 1 °C. (a) The 4-cell stage (1 h PF); (b) 16-cell stage (1 h 30 min PF); (c) 64-cell stage (2 h PF); (d) 512-cell stage (3 h PF); (e) high stage (4 h PF); (f) oblong stage (5 h PF); (g) between oblong and sphere stage (7 h PF); (h) sphere stage (8 h PF); (i) dome stage, beginning of epiboly (9 h PF); (j) 30% epiboly (10 h PF); (k) 60% epiboly (12 h PF); (l) 90% epiboly (14 h PF); (m) bud stage, blastoderm completely covers the yolk (16 h PF); (n) optic primordium appears in the prospective head region (18 h PF); (o) 6-somite stage, Kupffer’s vesicle appears at the base of the prospective tail region (20 h PF); (p) 10-somite stage (22 h PF). AP, animal pole; EVL, enveloping layer; H, prospective head region; KV, Kupffer’s vesicle; OG, oil globule; OP, optic primordium; SM, somite; T, prospective tail region; VP, vegetal pole; YSL, yolk syncytial layer. The scale bar at the bottom right corner in each figure represents 200 μm.
3.3.1. Cleavage Period (2- to 64-Cell Stage)
The embryos progressed from the 4-cell stage at 1 h post fertilization (PF; Figure 3a) to the 16-cell stage by 1 h 30 min PF (Figure 3b). However, from this point onwards, the exact number of blastomeres could not be counted. At 2 h PF, the presence of three blastomere layers suggested that the embryos had reached the 64-cell stage (Figure 3c), marking the end of the cleavage period and the beginning of blastula formation.
3.3.2. Blastula Period (128-Cell Stage to 30% Epiboly)
At 2 h 30 min PF, the embryos reached between the 128- and 256-cell stages, as indicated by the presence of 5–7 layers of blastomeres, and the outer blastomere layer formed the enveloping layer (EVL). At 3 h PF, the yolk syncytial layer (YSL) was formed between the inner layer of blastomeres and the yolk, and the embryos reached the approximately 512-cell stage (8–9 layers of blastomeres; Figure 3d). At 3 h 30 min PF, the embryos reached >1-k-cell stage; blastomeres also became much smaller as they continued to divide. The embryos reached the high stage at 4 h PF, and the blastodisc started to flatten (Figure 3e). At 5 h PF, the blastula developed an ellipsoidal shape with the flattened blastodisc (oblong stage; Figure 3f), and the boundary between the blastodisc and the yolk became much less distinguishable by 7 h PF (Figure 3g). At 8 h PF, the embryos were almost spherical, and the surface of the blastomeres became smooth (Figure 3h). At 9 h PF, part of the yolk protruded towards the animal pole and formed a dome-like structure, suggesting the beginning of epiboly (Figure 3i). From this point onwards, the blastoderm spread across the yolk from the animal to vegetal pole and covered 30% of the yolk by 10 h PF (30% epiboly stage; Figure 3j).
3.3.3. Gastrula Period (50% Epiboly to Bud Stage)
Epiboly continued during the gastrula period (Figure 3k), and approximately 90% of the yolk was covered by blastoderm at 14 h PF (Figure 3l). Concurrently, the blastoderm became thicker at the dorsal region (facing the animal pole) than the ventral region. By 16 h PF, the blastoderm had completely covered the yolk, and the prospective head region and the tail bud were both formed on the yolk surface (Figure 3m).
3.3.4. Segmentation and Pharyngula Period
At 18 h PF, the optic primordium was distinguishable in the prospective head region (Figure 3n). At 20 h PF, the Kupffer’s vesicle appeared near the base of the tail region, while the head region thickened (Figure 3o); additionally, the embryos reached the 6-somite stage, and the somite number increased to 10 by 22 h PF (Figure 3p). At 24 h PF, the rudiments of the eyes and lenses were developed at the head region, and the tail began to detach and protrude away from the yolk (Figure 4a).

Figure 4.
Embryonic development of Pseudochromis fridmani from 24 to 96 h post fertilization (PF) at 27 ± 1 °C. (a) At 24 h PF, showing rudiments of eyes and lens; (b) 48 h PF, showing the development of otic vesicles, otoliths, circulatory system, and pigmentation on the body; note the further development of the head and tail; (c) 72 h PF, showing the eyes pigmented with metallic green colour and differentiation of the gall bladder; blood vessels near the yolk are clearly visible, and more pigment cells are developed on the body; (d) 72 h PF, showing the olfactory bulbs, early development of the brain, and the enlarged otic vesicles; the tail has elongated; (e) 96 h PF, showing the reflective eyes, heart, operculum, gall bladder, and pectoral fin buds; most of the yolk and oil globule have been absorbed; (f) 96 h PF, showing the development of forebrain, midbrain and hindbrain. B, brain; BV, blood vessel; FB, forebrain; GB: gall bladder; H, heart; HB, hindbrain; L, lens; MB, midbrain; OB, olfactory blub; OPL, operculum; OT, otolith; OV, otic vesicle; PF, pectoral fin buds. The scale bar at the bottom right corner in each figure represents 200 μm.
The embryos underwent further organogenetic development from 24 to 72 h PF. At 48 h PF (Figure 4b), the head started to resemble that of a larva, and the rudiments of the eyes became lightly pigmented. The otic vesicles with two tiny otoliths each were differentiated posterior to the prospective brain region. The somites had developed into myotomes, and the tail substantially elongated compared to 24 h PF. Pigmented cells, i.e., the melanophores, also appeared along the posterior body of the embryos. At the same time, blood flowing through a beating heart was visible, and sporadic movements of the embryos within the chorion were also observed.
At 72 h PF, the iridophores were present to give a reflective metallic green pigmentation in the eyes (Figure 4c). Both the olfactory bulbs at the tip of the head and the brain had developed, and the otic vesicles also substantially enlarged (Figure 4d). The frequency of heartbeat increased, and more blood vessels were clearly visible near the yolk (Figure 4c). Moreover, the gall bladder was differentiated as a small green spherical structure posterior to the yolk, and pigmentation of light orange, black, and green was also scattered along the elongated embryo body and tail (Figure 4c). At this stage, sporadic twitches of the eyes and body inside the chorion were observed. When the embryos were handled for microscopy, the threads on the egg surface were also found to become much less adhesive.
At 96 h PF, the eyes developed a more reflective metallic green colour (Figure 4e). The operculum was distinguishable (Figure 4e), and the brain was clearly differentiated into forebrain, midbrain, and hindbrain (Figure 4f). The heart developed into a chamber shape with stronger and quicker heartbeats (Figure 4e). Furthermore, the pectoral fin buds also appeared, and substantial proportions of the yolk and the oil globule were absorbed (Figure 4e). The embryo body was now coiled more tightly within the chorion due to the continuous elongation of the tail. At this stage, frequent movements of the eyes, pectoral fin buds, and the tail were observed, suggesting that hatching was imminent. In general, the larvae hatched out in darkness by 102 h PF.
3.4. Larval Development and Associated Behavioural Observations
Newly hatched P. fridmani larvae (0 DPH) were 3.19 ± 0.05 mm and 0.34 ± 0.02 mm in notochord length (NL) and body depth (DP), respectively, and possessed pigmented eyes, developed jaws, and a gut in the form of a straight tube (Figure 5a and Figure 6a). At this stage, most of the yolk sac was reabsorbed, and the larvae actively swam with the pectoral fins and fin folds. On 1 DPH, the larvae showed positive phototactic behaviour, as they were observed to be attracted by torch light. On 2 DPH (Figure 5b), the yolk sac had been completely absorbed, and more small dots of orange pigmentation developed across the larval body. The larval intestine also widened with increased intestinal folds, and an intestinal valve clearly separated the anterior and posterior intestine regions (Figure 6b). The development of gill filaments behind the operculum was distinguishable on 4 DPH (Figure 6c). On 7 DPH, a coiled gut was developed (Figure 6d); the larval notochord tip flexed dorsally, and caudal fin buds were evident (flexion stage; Figure 5d). At this stage, positive phototaxis of larvae was reduced.

Figure 5.
Larval development of Pseudochromis fridmani from newly hatched larvae to juveniles (i.e., from 0 to 40 days post hatching, DPH). (a) 0-DPH larva with fin folds and a straight-tube gut, with most of the yolk sac absorbed; (b) 2-DPH larva; (c) 4-DPH larva with rotifers in the gut and the developing gill filaments behind the operculum; (d) 7-DPH larva, showing the development of an intestinal loop and the beginning of notochord flexion; (e) 10-DPH larva with a gut full of Artemia nauplii and caudal fin ray development showing the completion of flexion; (f) 13-DPH larva with well-developed caudal fin; (g) 16-DPH larva; (h) 19-DPH larva, showing most fins are well developed; (i) 22 DPH larva; (j) 25 DPH larva with tiny dots of pink pigmentation on the body; (k) 28-DPH larva with more small patches of pink pigmentation on the body; (l) 35-DPH larva; note that the larva becomes less transparent with increased pigmentation developed on the body; (m) 38-DPH larva undergoing pigmentation development on the body with visible melanophores around the eye; (n) 40-DPH juvenile with adult-type pigmentation pattern. The scale bar at the bottom right corner in each figure represents 1 mm.
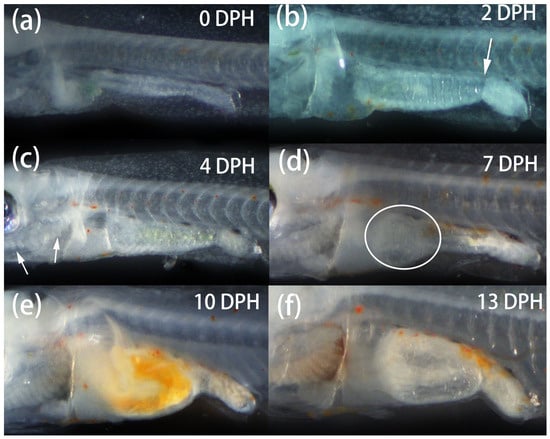
Figure 6.
Development in the ventral region of Pseudochromis fridmani larvae from 0 to 40 days post hatching (DPH). (a) 0-DPH larva with a straight-tube gut; (b) 2-DPH larva, showing a widened intestine with folds and an intestinal valve (arrowhead); (c) 4-DPH larva; note the intestine was filled with ingested rotifers, and the arrowheads indicate the development of gill filaments behind the operculum; (d) 7-DPH larva, the circle indicates a coiled gut forming in the anterior intestine region; (e) 10-DPH larva with a dilated and coiled intestine; note the ingested Artemia nauplii in the gut; (f) 13-DPH larva; note the large lumen formed by the coiled intestine with many folds on the intestinal wall.
On 10 DPH (Figure 6e), the coiled larval intestine started to dilate, and the P. fridmani larvae actively ingested Artemia nauplii as shown by the bright orange colour of gut contents. Most fin folds had reduced, and fin buds were differentiated at the posterior dorsal and anal fin region. Fin rays were differentiated in a short caudal fin, and notochord flexion was completed (Figure 5e). By 13 DPH, the coiled intestine of larvae formed a large lumen with many folds on the intestinal wall, occupying a significant amount of space in the abdomen (Figure 6f). The dorsal and anal fin rays were distinguishable, and the caudal fin had enlarged (Figure 5f). Under the stereomicroscope, small patches of light pink pigmentation were observed on the larval bodies.
The P. fridmani larvae shifted to negative phototaxis to strong light by 14 DPH, as they were observed to dart away from the bright torch light. By 19 DPH, most fins were well developed (Figure 5h), and there were no significant morphological changes between 19 and 28 DPH. The larval bodies became less transparent between 25 and 28 DPH, and the small patches of pink pigmentation were more prevalent across the bodies (Figure 5j,k). During this period, more larvae were also observed to swim close to the tank wall or tank bottom.
The transition to the P. fridmani juvenile stage, characterized by the adult-type pigmentation development, was observed to occur progressively from 31 to 55 DPH. By 35 DPH, the larvae developed a body shape similar to that of juveniles. While many larvae only showed a pale flash of light pink or orange colour across their bodies at this stage (Figure 5l), a small percentage (<5%) of larvae had turned purple. For most larvae, the melanophores first appeared near the eyes on 38 DPH (Figure 5m), when the average NL and BD reached 14.40 ± 1.23 mm and 3.52 ± 0.49 mm, respectively (Figure 7). Approximately 50% of P. fridmani developed the typical adult-type pigmentation on 45 DPH, and such a proportion increased to around 90% by 55 DPH.
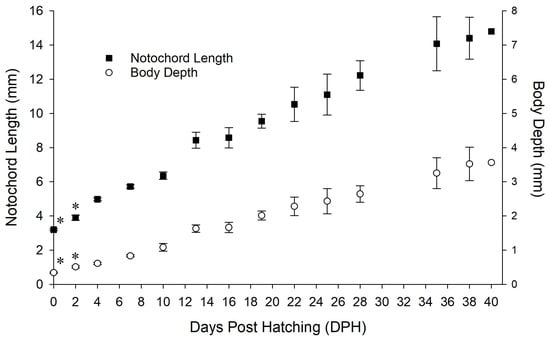
Figure 7.
Larval growth of Pseudochromis fridmani from 0 to 2, and from 4 to 40 days post hatching (DPH). Data are expressed as mean ± SD. Larvae from the same batch was sampled between 4 and 40 DPH (4–38 DPH: n = 9; 40 DPH: n = 1); growth data on 0 and 2 DPH (n = 15), as indicated by asterisk (*), were collected from a separate batch of larvae cultured under same rearing conditions.
On 56 DPH, despite reaching similar size to those individuals showed adult-like pigmentation pattern, around 5% of P. fridmani (out of a total of 387) larvae remained relatively transparent, with a very low level of pink and/or purple pigmentation.
4. Discussion
The present study documents several aspects of reproductive biology, with emphasis on spawning and filial cannibalism, and the early ontogeny of orchid dottyback P. fridmani in captivity. These findings build a baseline knowledge on the life cycle of P. fridmani and provide valuable insights for the aquaculture industry to manage and further improve the commercial production of P. fridmani for the marine ornamental fish trade.
The footages on the courting behaviour of P. fridmani provided a useful baseline to understand the bonding dynamics of this species. Similar to reports on sunrise dottyback P. flavivertex [34], readhead dottyback P. dilectus [29], and other dottyback species [10], P. fridmani males also displayed the typical courting behaviour of guiding the female to the spawning site. Interestingly, the P. fridmani males in present study were also observed to attempted to court with the females within 1–2 h after spawning ended, although the female apparently did not show a courting response. Similar dottyback behaviour has not been reported in the past. To date, little is known about the reproductive capability of dottyback males, as researchers [22,24,26,27,29,34] and aquarium hobbyists [10,28] generally keep dottybacks in pairs for captive breeding. However, P. fridmani were reported to live in small groups in the wild [11,12] and were able to be kept relatively peacefully in groups in an aquarium setting ([8], pers. obs.). To explore the potential of reproduction efficiency and capability of P. fridmani for commercial production purposes, it may be worthwhile to investigate this post-spawning courting behaviour of P. fridmani in a harem instead of pairs.
Suitable water temperature and photoperiod are essential for the successful captive spawning of reef fish [35,36], which is a prerequisite of breeding these fish to reduce collection pressure by the marine aquarium trade on wild stocks. In previous studies, relatively warm water temperatures and fixed photoperiods were used to achieve captive spawning of sunrise dottyback P. flavivertex (27 ± 0.5 °C, 14 h light, Olivotto et al. [34]), orchid dottyback P. fridmani (27 °C, 10 h light, Mies et al. [22]), and redhead dottyback P. dilectus (29 ± 1 °C, 14 h light, Madhu et al. [29]). The results of the present study showed P. fridmani pairs were able to spawn year-round at similar temperatures (27.5–29 °C), with tolerance of some photoperiodic changes. Over the 12-month period of the present study, the natural photoperiod fluctuated between 11 and 13 h of daylight, but the spawning frequencies of the seven P. fridmani pairs (every 5–11 days, most commonly every 6–8 days) were still in agreement with findings reported by Mies et al. [22]. Although extended periods of interrupted spawning were observed from pair B (mid-February–March) and F (July–August), they occurred at different times of the year, suggesting that it was not related to seasonal changes in photoperiod. Overall, the continuous and relatively stable spawnings of most broodstock pairs observed in this study suggest that P. fridmani is a year-round spawner. While little is known about the spawning seasonality of P. fridmani in the wild, it is possible that such a year-round spawning pattern might be related to specific captive conditions (e.g., temperature control, ambient tropical photoperiod, and sufficient feeding) used in this study. Nonetheless, the observed year-round spawning feature in captivity allows hatcheries to potentially breed and sell P. fridmani without seasonal fluctuations, closing the gaps between supply and demand of this popular species in the marine aquarium trade market.
Notably, the frequent spawnings of P. fridmani observed were also counteracted by the filial cannibalism, as all P. fridmani broodstock pairs in this study often partially or completely ate the eggs before hatching (Table 1). In general, the degree of filial cannibalism appeared to be highly pair-specific, as some pairs (e.g., pair A and G) were much more prone to filial cannibalism than others (e.g., pair C and E; Figure 1). However, even for a same pair (e.g., pair B and E), the pattern of filial cannibalism could highly vary month by month (Figure 1). In the literature, filial cannibalism has been reported from several species of dottyback kept in captivity [22,29]. In fact, it is considered as a major challenge for the egg management of dottybacks [23], as the unexpected egg loss during the incubation period could cause lasting disruptions in research and production plans. In an extreme case, it was reported that a P. fridmani pair cannibalized their eggs of 24 batches over a 6-month period before the eggs were successfully hatched for the first time [22].
Different hypotheses have been proposed to explain the filial cannibalistic behaviour, including to fulfill the energy cost of parental care [37,38,39,40], to remove embryos that are unlikely to hatch [41,42,43,44], and to restart courtship with a partner [45]. In the study done by Mies et al. [22], the authors observed that the eggs of seven dottyback species, including P. fridmani, were often eaten on 2 or 3 day post spawning, and the eaten eggs appeared to have developmental issues. In this study, filial cannibalism by P. fridmani was generally observed during the late phase of the incubation period (i.e., 3 and 4 DPF; Figure 2). Since signs of P. fridmani embryonic development (i.e., change of egg colour and appearance) were regularly observed by naked eyes during incubation, fertilization problems are an unlikely reason for the filial cannibalism observed. However, it could be possible that the P. fridmani broodstocks cannibalized the eggs due to potential egg developmental issues that were not investigated in this study. To date, the specific mechanism behind the filial cannibalism of dottyback species remains to be investigated. To advance breeding and support the research on dottybacks in captivity, relevant studies in the near future are much needed to understand the filial cannibalism of dottybacks and hence mitigate the associated risks of egg loss during larval production.
The detailed description of embryonic development of P. fridmani from the present study serves as an important benchmark to understand the early ontogeny of this species. The first 48 h post fertilization appears to be a crucial developmental period for P. fridmani, as its embryos drastically developed from the single-cell stage of blastomeres (Figure 3) to embryos with a head, tail, rudiments of the eye and lens, and a circulatory system (Figure 4). To enhance the hatching and subsequent larval-rearing success this species, care must be taken in first 48 h period to ensure proper embryonic development at the provided incubation condition. The embryo incubation of P. fridmani lasted for 96–102 h, which was similar to the 96 h reported from P. flavivertex [34] and P. dilectus [29]. The stage of embryonic development achieved by 96 h PF (a few hours before hatching) was also highly comparable among these three species. However, despite the similar mean incubation temperature of 27 °C, the embryonic developmental rate appeared to vary among these dottyback species (Table 3). For instance, while the embryos of P. dilectus reached 75% epiboly stage at 7 h 30 min PF, the embryos of P. fridmani only reached 60% epiboly by 12 h PF. At 72 h PF, the retina of P. fridmani embryos was pigmented with a metallic green colour, but it was only partially pigmented for P. flavivertex embryos. Differences in the rate of embryonic development among species within a same genus have also been reported from marine angelfish Centropyge. spp. [20], and they likely represent species-specific features.
Table 3.
Comparison of the timing (hour post fertilization) of embryonic development events among three dottyback species from the genus Pseudochromis.
Understanding the larval development of marine fish can provide valuable insights into optimizing the larval-rearing techniques for candidate species. In the present study, substantial developmental changes occurred in P. fridmani between 0 and 14 DPH. In terms of morphology, the development of fin folds into fin rays or fin was completed by 13 DPH. Larval length (NL) increased approximately 70% within 9 days from 4 to 13 DPH, while it took the following 25 days for larvae to reach similar growth again (71% increment, 13 to 38 DPH; Figure 7). The gut of P. fridmani had differentiated from a straight tube post hatching to a coiled gut (7 DPH), which furthered formed a large lumen by 13 DPH. The timings of these noticeable and observed gut developments of P. fridmani are also in agreement with Chen et al. [24], suggesting that P. fridmani have gained significantly improved digestive capabilities by 13 DPH [24]. Furthermore, the phototaxis of P. fridmani shifted from positive on 1 DPH to negative by 14 DPH, indicating major developments in the larval visual system [46,47]. From the larval-rearing perspective, supplying adequate dietary nutrients during this critical stage of 0 to 14 DPH is thus likely a key to supporting development as well as improving survival for P. fridmani larvae. Additionally, providing suitable rearing conditions to accommodate the phototactic behaviour change of P. fridmani, such as adjusting lighting conditions in the tank at different larval stages, may also be important for optimizing larval-rearing success [48,49].
5. Conclusions
The captive breeding success of P. fridmani helps secure a reliable supply of embryos and larvae for relevant scientific research and aquaculture production. The present study adds to the limited literature on the biology of Pseudochromidae by expanding the knowledge base on the reproductive biology and early ontogeny of P. fridmani in captivity, whereas relevant information is difficult to obtain in field studies. The findings provide an important foundation not only for optimizing breeding techniques for P. fridmani and other dottyback species but also for studying dottyback ecology and population dynamics in the wild. To deepen the understanding of biology and optimize aquaculture production of P. fridmani, the results of present study encourage future studies to (1) understand and manage filial cannibalism by broodstock, (2) explore the spawning dynamics in a harem structure, and (3) optimize larval dietary nutrition and rearing conditions, especially during the first 14 DPH.
Author Contributions
J.Y.C. contributed to project ideas, data generation, data analysis, manuscript preparation, and editing; C.Z. contributed to project ideas, supervision, manuscript editing, and funding; J.M.C. contributed to project ideas, data interpretation, and manuscript editing. All authors have read and agreed to the published version of the manuscript.
Funding
The first author was supported by the James Cook University postgraduate research scholarship when the research was conducted.
Institutional Review Board Statement
The research presented was conducted within the guidelines of the Australian code for care and use of animals for scientific purposes and the Animal Care and Protection Act (2001, Queensland Government), Animal Care and Protection Regulation (2012, Queensland Government). The proposed methods, use of animals, and research practice (application number: A2467) were examined and approved by the Animal Welfare and Ethics Committee at James Cook University.
Data Availability Statement
The authors confirm that the data supporting the findings are all available in form of text, graphs, tables and figures presented on this manuscript.
Acknowledgments
We would like to thank the Centre for Macroalgal Resources and Biotechnology, Centre for Sustainable Tropical Fisheries and Aquaculture at James Cook University for providing access to the equipment needed for data collection. Special thanks to Da Li for providing technical assistance with image editing, and Yau Ming Michael Lai for assisting with data collection.
Conflicts of Interest
We confirm that there are no conflict of interest to disclose.
References
- Froese, R.; Pauly, D. Family Pseudochromidae—Dottybacks. Available online: https://www.fishbase.se/summary/FamilySummary.php?ID=291 (accessed on 6 January 2023).
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Family PSEUDOCHROMIDAE (289)—Dottybacks. In Fishes of the World; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 339–340. [Google Scholar]
- Gill, A.C.; Allen, G.R.; Erdmann, M.V. Pseudochromis stellatus, a New Species of Dottyback from Indonesia (Teleostei: Pseudochromidae). Zootaxa 2017, 4338, 333–340. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gill, A.C.; Allen, G.R.; Erdmann, M. Two New Dottyback Species of the Genus Pseudochromis from Southern Indonesia (Teleostei: Pseudochromidae). Zootaxa 2012, 60, 53–60. [Google Scholar] [CrossRef]
- Lieske, E.; Myers, R. Dottybacks. In Coral Reef Fishes: Indo-Pacific & Caribbean; HarperCollins Publishers: London, UK, 1993; pp. 31–33. ISBN 0002199742. [Google Scholar]
- Kuiter, R.H. Dottybacks—Pseudochromidae. In Guide to Sea Fishes of Australia: A Comprehensive Reference for Divers and Fishermen; New Holland Publisher (Australia) Pty Ltd.: Sydney, Australia, 2016; pp. 124–127. [Google Scholar]
- Randall, J.E.; Allen, G.R.; Steene, R.C. Dottybacks and Eel Blennies. In Fishes of the Great Barrier Reef and Coral Sea; University of Hawaii Press: Honolulu, HI, USA, 1997; pp. 124–132. [Google Scholar]
- Lubbock, R. Fishes of the Family Pseudochromidae (Perciformes) in the Northwest Indian Ocean and Red Sea. J. Zool. 1975, 176, 115–157. [Google Scholar] [CrossRef]
- Rhyne, A.L.; Tlusty, M.F.; Schofield, P.J.; Kaufman, L.; Morris, J.A.; Bruckner, A.W. Revealing the Appetite of the Marine Aquarium Fish Trade: The Volume and Biodiversity of Fish Imported into the United States. PLoS ONE 2012, 7, e35808. [Google Scholar] [CrossRef] [PubMed]
- Wittenrich, M.L. The Complete Illustrated Breeder’s Guide to Marine Aquarium Fishes; T.F.H. Publications: Neptune City, NJ, USA, 2007. [Google Scholar]
- Field, R.; Field, M. Dottybacks. In Reef Fish of the Red Sea; Routledge Ltd.: New York, NY, USA, 2016; pp. 76–78. [Google Scholar]
- Taquet, M.; Diringer, A. Dottybacks. In Fishes of the Indian Ocean and Red Sea; Quae: Versailles, France, 2012; pp. 233–236. ISBN 9782759218899. [Google Scholar]
- Jiang, J.; Nuez-Ortin, W.; Angell, A.; Zeng, C.; de Nys, R.; Vucko, M.J. Enhancing the Colouration of the Marine Ornamental Fish Pseudochromis fridmani Using Natural and Synthetic Sources of Astaxanthin. Algal Res. 2019, 42, 101596. [Google Scholar] [CrossRef]
- LiveAquaria ORA® Captive-Bred Orchid Dottyback. Available online: https://www.liveaquaria.com/product/prod_display.cfm?c=15+38+3465&pcatid=3465 (accessed on 10 April 2023).
- Palmtag, M.R. The Marine Ornamental Species Trade. In Marine Ornamental Species Aquaculture; Calado, R., Olivotto, I., Oliver, M.P., Holt, G.J., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2017; pp. 3–14. [Google Scholar]
- Wabnitz, C.; Taylor, M.; Green, E.; Razak, T. From Ocean to Aquarium: The Global Trade in Marine Ornamental Species; UNEP–WCMC: Cambridge, UK, 2003. [Google Scholar]
- Olivotto, I.; Chemello, G.; Vargas, A.; Randazzo, B.; Piccinetti, C.C.; Carnevali, O. Marine Ornamental Species Culture: From the Past to “Finding Dory”. Gen. Comp. Endocrinol. 2017, 245, 116–121. [Google Scholar] [CrossRef]
- Degidio, J.M.L.A.; Yanong, R.P.E.; Watson, C.A.; Ohs, C.L.; Cassiano, E.J.; Barden, K. Spawning, Embryology, and Larval Development of the Milletseed Butterflyfish Chaetodon miliaris in the Laboratory. N. Am. J. Aquac. 2017, 79, 205–215. [Google Scholar] [CrossRef]
- Majoris, J.E.; Francisco, F.A.; Atema, J.; Buston, P.M. Reproduction, Early Development, and Larval Rearing Strategies for Two Sponge-Dwelling Neon Gobies, Elacatinus lori and E. colini. Aquaculture 2018, 483, 286–295. [Google Scholar] [CrossRef]
- Mendoncą, R.C.; Chen, J.Y.; Zeng, C.; Tsuzuki, M.Y. Embryonic and Early Larval Development of Two Marine Angelfish, Centropyge bicolor and Centropyge bispinosa. Zygote 2020, 28, 196–202. [Google Scholar] [CrossRef]
- Moorhead, J.A. Broodstock Management, Development of Rearing Systems, and Feeding Regimes for Larvae of the Forktail Blenny, Meiacanthus atrodorsalis. Ph.D. Thesis, James Cook University, Townsville, Australia, 2017. [Google Scholar]
- Mies, M.; Güth, A.Z.; Scozzafave, M.S.; Sumida, P.Y.G. Spawning Behaviour and Activity in Seven Species of Ornamental Dottybacks. J. Zoo Aquar. Res. 2014, 2, 117–122. [Google Scholar]
- Shei, M.; Mies, M.; Olivotto, I. Other Demersal Spawners and Mouthbrooders. In Marine Ornamental Species Aquaculture; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 223–250. [Google Scholar] [CrossRef]
- Chen, J.Y.; Zeng, C.; Cobcroft, J.M. Digestive System Ontogeny and the Effects of Weaning Time on Larval Survival, Growth and Pigmentation Development of Orchid Dottyback Pseudochromis fridmani. Aquaculture 2022, 549, 737737. [Google Scholar] [CrossRef]
- Wittenrich, M.L.; Turingan, R.G. Linking Functional Morphology and Feeding Performance in Larvae of Two Coral-Reef Fishes. Environ. Biol. Fishes 2011, 92, 295–312. [Google Scholar] [CrossRef]
- Chen, J.Y.; Zeng, C. The Effects of Live Prey and Greenwater on the Early Larval Rearing of Orchid Dottyback Pseudochromis fridmani. Aquaculture 2021, 543, 737008. [Google Scholar] [CrossRef]
- Chen, J.Y.; Zeng, C. Transition to Artemia Feeding Phase for Orchid Dottyback Pseudochromis fridmani Larvae: Establishing Suitable Prey Shift Time and Strategy. Aquaculture 2021, 545, 737180. [Google Scholar] [CrossRef]
- Moe, M.A. Breeding the Orchid Dottyback, Pseudochromis fridmani: An Aquarist’s Journal; Green Turtle Publications: Plantation, FL, USA, 1997; ISBN 9780939960095. [Google Scholar]
- Madhu, K.; Madhu, R.; Retheesh, T. Spawning, Embryonic Development and Larval Culture of Redhead Dottyback Pseudochromis dilectus Lubbock, 1976 under Captivity. Aquaculture 2016, 459, 73–83. [Google Scholar] [CrossRef]
- Madhu, K.; Madhu, R. Captive Spawning and Embryonic Development of Marine Ornamental Purple Firefish Nemateleotris decora (Randall & Allen, 1973). Aquaculture 2014, 424–425, 1–9. [Google Scholar] [CrossRef]
- Leu, M.Y.; Tai, K.; Meng, P.; Tang, C.; Wang, P. Embryonic, Larval and Juvenile Development of the Long Fin Batfish, Platax teira (Forsskål,1775) under Controlled Conditions with Special Regard to Mitigate Cannibalism for Larviculture. Aquaculture 2018, 493, 204–213. [Google Scholar] [CrossRef]
- Zeng, C.; Shao, L.; Ricketts, A.; Moorhead, J. The Importance of Copepods as Live Feed for Larval Rearing of the Green Mandarin Fish Synchiropus splendidus. Aquaculture 2018, 491, 65–71. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Olivotto, I.; Rollo, A.; Sulpizio, R.; Avella, M.; Tosti, L.; Carnevali, O. Breeding and Rearing the Sunrise Dottyback Pseudochromis flavivertex: The Importance of Live Prey Enrichment during Larval Development. Aquaculture 2006, 255, 480–487. [Google Scholar] [CrossRef]
- Olivotto, I.; Holt, S.A.; Carnevali, O.; Holt, G.J. Spawning, Early Development, and First Feeding in the Lemonpeel Angelfish Centropyge Flavissimus. Aquaculture 2006, 253, 270–278. [Google Scholar] [CrossRef]
- Baensch, F.U.; Tamaru, C.S. Spawning and Development of Larvae and Juveniles of the Rare Blue Mauritius Angelfish, Centropyge debelius (1988), in the Hatchery. J. World Aquac. Soc. 2009, 40, 425–439. [Google Scholar] [CrossRef]
- Manica, A. Parental Fish Change Their Cannibalistic Behaviour in Response to the Cost-to-Benefit Ratio of Parental Care. Anim. Behav. 2004, 67, 1015–1021. [Google Scholar] [CrossRef]
- Mehlis, M.; Bakker, T.C.M.; Frommen, J.G. Nutritional Benefits of Filial Cannibalism in Three-Spined Sticklebacks (Gasterosteus aculeatus). Naturwissenschaften 2009, 96, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Manica, A. Filial Cannibalism in Teleost Fish. Biol. Rev. 2002, 77, 261–277. [Google Scholar] [CrossRef]
- Rohwer, S. Parent Cannibalism of Offspring and Egg Raiding as a Courtship Strategy. Am. Nat. 1978, 112, 429–440. [Google Scholar] [CrossRef]
- Petersen, C.W.; Marchetti, K. Filial Cannibalism in the Cortez Damselfish Stegastes rectifraenum. Evolution 1989, 43, 158–168. [Google Scholar] [CrossRef]
- Vallon, M.; Heubel, K.U. Old but Gold: Males Preferentially Cannibalize Young Eggs. Behav. Ecol. Sociobiol. 2016, 70, 569–573. [Google Scholar] [CrossRef]
- Payne, A.G.; Smith, C.; Campbell, A.C. Filial Cannibalism Improves Survival and Development of Beaugregory Damselfish Embryos. Proc. R. Soc. B Biol. Sci. 2002, 269, 2095–2102. [Google Scholar] [CrossRef]
- Mrowka, W. Filial Cannibalism and Reproductive Success in the Maternal Mouthbrooding Cichlid Fish Pseudocrenilabrus multicolor. Behav. Ecol. Sociobiol. 1987, 21, 257–265. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Tateishi, T.; Terada, R.; Soyano, K.; Takegaki, T. Filial Cannibalism by Male Fish as an Infanticide to Restart Courtship by Self-Regulating Androgen Levels. Curr. Biol. 2018, 28, 2831–2836. [Google Scholar] [CrossRef] [PubMed]
- Stearns, D.E.; Holt, J.G.; Forward Jr, R.B.; Pickering, P.L. Ontogeny of Phototactic Behavior in Red Drum Larvae (Sciaenidae: Sciaenops ocellatus). Mar. Ecol. Prog. Ser. 1994, 104, 1–11. [Google Scholar] [CrossRef]
- Hilder, P.E.; Battaglene, S.C.; Hart, N.S.; Collin, S.P.; Cobcroft, J.M. Retinal Adaptations of Southern Bluefin Tuna Larvae: Implications for Culture. Aquaculture 2019, 507, 222–232. [Google Scholar] [CrossRef]
- Papadakis, I.E.; Kentouri, M.; Divanach, P.; Mylonas, C.C. Ontogeny of the Eye of Meagre (Argyrosomus regius) from Hatching to Juvenile and Implications to Commercial Larval Rearing. Aquaculture 2018, 484, 32–43. [Google Scholar] [CrossRef]
- Ina, Y.; Sakakura, Y.; Tanaka, Y.; Yamada, T.; Kumon, K.; Eba, T.; Hashimoto, H.; Konishi, J.; Takashi, T.; Gen, K. Development of Phototaxis in the Early Life Stages of Pacific Bluefin Tuna Thunnus orientalis. Fish. Sci. 2017, 83, 537–542. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).