3.2.1. Subgenus Cryptocambeva Costa, 2021
Species of
Cryptocambeva may be diagnosed by a series of apomorphic conditions, including a small posttemporo-supracleithrum, resulting in a broad interspace between the posttemporo-supracleithrum and adjacent bones, and a colour pattern consisting of small dark brown to black dots scattered over all the body [
26,
41].
Cryptocambeva and its sister group, the subgenus
Humboldtglanis Costa, 2021, share a series of synapomorphies, including the presence of a long and narrow ridge on the dorsal surface of the autopalatine, the ventral surface of the lateral margin of the mesethmoid being ventrally folded around the vomer margin, and the lateral ethmoid being medially separated from its symmetrical homologous by an interspace. Species of
Cryptocambeva are easily distinguished from species of
Humboldtglanis by species of the former subgenus having six or seven pectoral-fin rays (vs. always eight). In the MRGD, two morphological patterns are present: a clade herein called the
Trichomycterus candidus complex, which contains three small species not surpassing about 60 mm SL and lacking pelvic fin and girdle (see discussion below), and an assemblage comprising three species not forming a monophyletic group, but sharing well-developed pelvic fin and girdle and a relatively robust body, often reaching about 100–120 mm SL. Species of the
Trichomycterus candidus complex are typically found among leaves of amphibious plants, thus contrasting with other congeners that are commonly found deeply buried in riverbanks, often close to marginal plant roots or within bottom leaf litter.
Trichomycterus (Cryptocambeva) candidus (Miranda Ribeiro, 1949)
Eremophilus candidus Miranda Ribeiro, 1949: [
19] (2) (original description; type locality: ‘pequeno córrego que cai no ribeirão Espírito Santo [21°05′44′′ S 46°08′46′′ W], afluente do Rio Claro e este do Sapucaí, que desagua no Rio Grande, Município de Conceição Aparecida, Estado de Minas Gerais” [small stream that flows into the Ribeirão Espírito Santo, a tributary of the Rio Claro, which is tributary of the Sapucaí, which flows into the Rio Grande, Municipality of Conceição Aparecida, State of Minas Gerais]; holotype: MNRJ 5209).
Trichomycterus candidus: [
41] (180) (new combination).
Figure 2.
Trichomycterus candidus, UFRJ 12824, Conceição da Aparecida Municipality, 43.1 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Figure 2.
Trichomycterus candidus, UFRJ 12824, Conceição da Aparecida Municipality, 43.1 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Diagnosis.
Trichomycterus candidus differs from all other congeners of
Cryptocambeva, except for
Trichomycterus listruroides Costa, Katz & Azevedo-Santos sp. nov. and
Trichomycterus uberabensis Costa, Azevedo-Santos & Katz sp. nov., by the pelvic fin and girdle that are absent in most specimens, or rudimentary in two specimens (UFRJ 12728, see discussion below), in contrast to being always well developed.
Trichomycterus candidus differs from
T. listruroides Costa, Katz & Azevedo-Santos sp. nov. and
T. uberabensis Costa, Azevedo-Santos & Katz sp. nov. by having a unique colour pattern, comprising minute dark brown dots, which are equal to or smaller than the orbital diameter, irregularly arranged over the flank, and separated by broad interspaces; a horizontal row of slightly larger dark brown dots along the longitudinal midline of the flank; and another similar horizontal row on the dorsal portion of the flank (vs. flank dots always larger than the orbital diameter, forming a vermiculate pattern in
T. listruroides, and small round spots in
T. uberabensis).
Trichomycterus candidus is also distinguished from
T. listruroides Costa, Katz & Azevedo-Santos sp. nov. by having a caudal fin margin not aligned with the caudal peduncle (vs. in a continuous line); fewer dorsal (19 or 20 vs. 24 or 25) and ventral (17 or 18 vs. 20) procurrent caudal-fin rays; fewer vertebrae (38 or 39 vs. 40); dorsal-fin origin at the vertical between the centrum of the 22nd or 23rd vertebra (vs. 24th); and anal-fin origin at vertical through the centrum of the 25th vertebra (vs. 26th or 27th). It is also distinguished from
T. uberabensis by having more ventral procurrent caudal-fin rays (17 or 18 vs. 13–16), fewer interopercular odontodes (16–19, vs. 21–24), and fewer teeth on the dentary (21–25, vs. 29–32). For a full description and anatomical illustrations, see Barbosa and Costa [
41].
Distribution.
Trichomycterus candidus occurs in the western tributaries of the Rio Sapucaí, Rio Grande drainage, upper Rio Paraná basin (
Figure 3B), at altitudes of about 800 m asl. Specimens from two populations, one from Ribeirão Antinha, Capetinga, and another from the Rio Itaci, an eastern tributary of the Rio Sapucaí (
Figure 3B), were tentatively identified as
Trichomycterus cf.
candidus. Specimens from these populations are similar to
T. candidus, except for some smaller specimens from the Ribeirão Antinha having a distinct colour pattern (i.e., dots coalesced to form two stripes on the flank), fewer vertebrae (37 vs. 38–39), and more opercular odontodes (12 or 13 vs. 8–10), and specimens from the Rio Itaci having flank dots not forming distinctive rows (vs. two distinctive longitudinal rows).
Material examined. Trichomycterus (Cryptocambeva) candidus: all localities in Brazil: Minas Gerais State: Rio Grande drainage, Rio Paraná basin. Conceição da Aparecida Municipality: MNRJ 5209, holotype; stream tributary of Rio Claro, with itself being a tributary of Rio Sapucaí, 21°05′44′′ S 46°08′46′′ W; J. C. M. Carvalho & C. Lako, October 1947. MNRJ 5356, 21 ex.; Renascença Farm; J. C. M. Carvalho & A. L. de Castro, November 1948. UFRJ 4926, 31 ex.; UFRJ 4928, 5 ex. (C&S); Riacho Cuiabá; D. Almeida, M. P. Gonçalves & M. A. Barbosa, 24 October 1999. UFRJ 12824, 6; stream near the road MG-184, 21°03′25′′ S 46°08′55′′ W, at about 800 m asl; A. M. Katz & V.M. de Azevedo Santos, 30 October 2021. Carmo do Rio Claro Municipality: UFRJ 12728, 15 ex.; Ribeirão Santa Quitéria, Cachoeira da Pedra Molhada, 21°00′18′′ S 46°14′49′′ W, at about 810 m asl; V.M. Azevedo-Santos et al., 21 July 2017. UFRJ 12826, 4 ex.; same locality as UFRJ 12728; A. M. Katz & V.M. Azevedo-Santos, 30 October 2021. Trichomycterus (Cryptocambeva) cf. candidus 1: UFRJ 8384, 16 ex.; 8391, 3 ex. (C&S); Capetinga Municipality: stream tributary of Ribeirão Antinha, Rio Grande drainage, Rio Paraná basin, 20°38′23′′ S 46°59′02′′ W, altitude about 750 m asl; V.M. Azevedo-Santos, 4 November 2011. UFRJ 8372, 5 ex.; same locality and collector as preceding, 21 August 2011. Trichomycterus (Cryptocambeva) cf. candidus 2: UFRJ 12823, 2 (C&S); stream tributary of Rio Itaci, a tributary of Rio Sapucaí, Rio Grande drainage, upper Rio Paraná basin, 20°54′58′′ S 45°56′21′′ W, about 840 m asl; A. M. Katz & V. M. Azevedo-Santos, 31 October 2021. UFRJ 12825, 2 ex.; same locality as preceding; A. M. Katz & V.M. Azevedo-Santos, 31 October 2021.
Figure 3.
Geographical distribution of Trichomycterus in the Middle Rio Grande drainage. (A) T. garbei Costa, Azevedo-Santos & Katz sp. nov. (triangle), T. pirabitira (dots), and T. saturaturatus Costa, Katz & Azevedo-Santos sp. nov. (star); (B) T. candidus (dots), T. cf. candidus 1 (triangle), T. cf. candidus 2 (square), T. listruroides Costa, Katz & Azevedo-Santos sp. nov. (star), and T. uberabensis Costa, azevedo-Santos & Katz sp. nov. (lozenge); (C) T. adautoleitei Costa, Azevedo-Santos & Katz sp. nov. (square), T. coelhorum Costa, Azevedo-Santos & Katz sp. nov. (lozenge), T. sainthilairei (star), and T. septemradiatus (dots). T indicates type localities.
Figure 3.
Geographical distribution of Trichomycterus in the Middle Rio Grande drainage. (A) T. garbei Costa, Azevedo-Santos & Katz sp. nov. (triangle), T. pirabitira (dots), and T. saturaturatus Costa, Katz & Azevedo-Santos sp. nov. (star); (B) T. candidus (dots), T. cf. candidus 1 (triangle), T. cf. candidus 2 (square), T. listruroides Costa, Katz & Azevedo-Santos sp. nov. (star), and T. uberabensis Costa, azevedo-Santos & Katz sp. nov. (lozenge); (C) T. adautoleitei Costa, Azevedo-Santos & Katz sp. nov. (square), T. coelhorum Costa, Azevedo-Santos & Katz sp. nov. (lozenge), T. sainthilairei (star), and T. septemradiatus (dots). T indicates type localities.
Trichomycterus (Cryptocambeva) garbei Costa, Azevedo-Santos & Katz sp. nov.
LSID: urn:lsid:zoobank.org:act:4F240F1F-C0F4-40D0-9778-6C2B9521F799
Holotype. UFRJ 12917, 60.8 mm SL; Brazil: São Paulo State: Cristais Paulista Municipality: stream tributary of Rio Canoas, which is a tributary of Rio Grande at Estreito dam, road Cristais Paulista to Mascarenhas, 20°18′07′′S 47°18′45′′W, at about 715 m asl; V. M. Azevedo-Santos et al., 16 April 2022.
Paratypes. UFRJ 12918, 11 ex., 32.3–70.5 mm SL; UFRJ 12960, 3 ex., 38.8–53.3 mm SL (C&S); CICCAA 07645, 4 ex., 42.6–49.6 mm SL; UFRJ 12896, 9 ex. (DNA), 34.3–62.5 mm SL; collected with holotype.
Figure 5.
Mesethmoidal region, with middle and left portions, and in dorsal view: (A) Trichomycterus (Cryptocambeva) garbei Costa, Azevedo-Santos & Katz sp. nov.; (B) Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov.; (C) Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov.; (D) Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov.; (E) Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov.; and (F) Trichomycterus (Paracambeva) coelhorum Costa, Azevedo-Santos & Katz sp. nov. Larger stippling represents cartilage.
Figure 5.
Mesethmoidal region, with middle and left portions, and in dorsal view: (A) Trichomycterus (Cryptocambeva) garbei Costa, Azevedo-Santos & Katz sp. nov.; (B) Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov.; (C) Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov.; (D) Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov.; (E) Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov.; and (F) Trichomycterus (Paracambeva) coelhorum Costa, Azevedo-Santos & Katz sp. nov. Larger stippling represents cartilage.
Diagnosis. Trichomycterus garbei is distinguished from all other congeners of the subgenus Cryptocambeva by possessing a dorsally expanded caudal peduncle as a result of long and numerous dorsal procurrent caudal-fin rays (29–32 vs. 15–25). Additionally, it is distinguished from other species of Cryptocambeva from the Rio Grande drainage by a combination of the presence of well-developed pelvic fin and girdle (vs. absence in species of the T. candidus complex), the absence of the anterior segment of the infraorbital canal (vs. presence in T. pirabitira and T. saturatus Costa, Katz & Azevedo-Santos sp. nov.), and 19 or 20 ventral procurrent caudal-fin rays (vs. 13–18 in T. pirabitira and T. saturatus).
Description. General morphology: Morphometric data are presented in
Table 1. Body relatively deep, compressed. Greatest body depth in area immediately anterior to pelvic-fin base, or sometimes at middle portion of caudal peduncle. Dorsal and ventral profiles slightly convex between snout and anterior limit of caudal peduncle. Dorsal margin of caudal peduncle slightly expanded on its middle portion resulting in convex dorsal profile; ventral profile of caudal peduncle nearly straight. Anus and urogenital papilla opening at vertical just anterior to middle portion of dorsal-fin base. Head sub-trapezoidal in dorsal view, with anterior profile of snout convex. Eye small, dorsally positioned on head, nearer snout tip than posterior margin of opercle. Posterior nostril nearer anterior nostril than orbit. Tip of nasal barbel posteriorly reaching area between orbit and opercle, sometimes reaching middle of opercle; tip of maxillary barbel reaching between middle of interopercular patch of odontodes and area between interopercle and pectoral-fin base; rictal barbel reaching between middle and posterior portion of interopercular patch of odontodes. Mouth subterminal. Jaw teeth pointed, irregularly arranged, 46–70 on premaxilla, 50–59 on dentary. Minute skin papillae on dorsal and ventral surfaces of head. Branchial membrane attached to isthmus only at its anterior-most point, in ventral midline.
Table 1.
Morphometric data of Trichomycterus garbei Costa, Azevedo-Santos & Katz sp. nov.
Table 1.
Morphometric data of Trichomycterus garbei Costa, Azevedo-Santos & Katz sp. nov.
| | Holotype | Paratypes (n = 10) |
|---|
| | | |
| Standard length (SL) | 60.8 | 43.6–70.5 |
| | | |
Percentage of standard length | |
| Body depth | 17.1 | 15.6–20.1 |
| Caudal peduncle depth | 17.3 | 14.9–17.6 |
| Body width | 10.9 | 9.8–12.9 |
| Caudal peduncle width | 2.9 | 2.2–3.2 |
| Pre-dorsal length | 63.6 | 58.7–63.8 |
| Pre-pelvic length | 59.7 | 55.9–59.3 |
| Dorsal-fin base length | 12.0 | 10.4–12.9 |
| Anal-fin base length | 10.2 | 9.0–11.4 |
| Caudal-fin length | 17.6 | 17.4–19.9 |
| Pectoral-fin length | 13.8 | 13.6–15.8 |
| Pelvic-fin length | 9.4 | 8.9–10.7 |
| Head length | 20.0 | 18.8–21.2 |
| | | |
Percentage of head length | |
| Head depth | 49.3 | 44.0–51.5 |
| Head width | 88.7 | 81.6–89.7 |
| Snout length | 40.1 | 38.1–44.5 |
| Interorbital width | 28.3 | 23.9–29.4 |
| Preorbital length | 12.6 | 10.0–13.3 |
| Eye diameter | 9.2 | 9.0–11.8 |
Dorsal and anal fins subtriangular. Total dorsal-fin rays 11 (ii + II + 7), total anal-fin rays 9 (ii + II + 5). Anal-fin origin at vertical through posterior portion of dorsal-fin base, at vertical through base of 5th bifid dorsal-fin ray. Dorsal-fin origin at vertical between centrum of 16th and 18th vertebra; anal-fin origin at vertical between centrum of 20th and 22nd vertebra. Pectoral fin subtriangular in dorsal view, first pectoral-fin ray terminating in filament about 15–30 % pectoral-fin length excluding filament. Total pectoral-fin rays 7 (I + 6). Pelvic fin subtruncate, posteriorly overlapping anus and urogenital papilla, its posterior extremity at vertical just posterior to middle portion of dorsal-fin base. Pelvic-fin bases medially separated by small interspace, about one third of pelvic-fin base. Total pelvic-fin rays 5 (I + 4). Caudal fin subtruncate, posterior corners rounded. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 29–32 (xxviii–xxxi + I), total ventral procurrent rays 19 or 20 (xviii–xix + I).
Figure 6.
Left jaw suspensorium and opercular series, in lateral view: (A) Trichomycterus (Cryptocambeva) garbei Costa, Azevedo-Santos & Katz sp. nov.; (B) Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov.; (C) Trichomycterus (Cryptocambeva) pirabitira; (D) Trichomycterus (Cryptocambeva) mirissumba; (E) Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov.; (F) Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov.; (G) Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov.; and (H) Trichomycterus (Paracambeva) coelhorum Costa, Azevedo-Santos & Katz sp. nov. Larger stippling represents cartilage.
Figure 6.
Left jaw suspensorium and opercular series, in lateral view: (A) Trichomycterus (Cryptocambeva) garbei Costa, Azevedo-Santos & Katz sp. nov.; (B) Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov.; (C) Trichomycterus (Cryptocambeva) pirabitira; (D) Trichomycterus (Cryptocambeva) mirissumba; (E) Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov.; (F) Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov.; (G) Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov.; and (H) Trichomycterus (Paracambeva) coelhorum Costa, Azevedo-Santos & Katz sp. nov. Larger stippling represents cartilage.
Laterosensory system: Supraorbital, posterior section of infraorbital canal and postorbital canal continuous. Supraorbital sensory canal pores 3: s1, adjacent to medial margin of anterior nostril; s3, adjacent and just posterior to medial margin of posterior nostril; s6, in transverse line through posterior half of orbit; pore s6 slightly nearer its homologous pore than orbit. Anterior infraorbital sensory canal absent. Posterior infraorbital sensory canal pores 2: pore i10, adjacent to ventral margin of orbit, and pore i11, posterior to orbit. Postorbital canal pores 2: po1, at vertical through posterior portion of interopercular patch of odontodes, and po2, at vertical through posterior portion of opercular patch of odontodes. Lateral line pores 2; posterior-most pore at vertical just posterior to pectoral-fin base.
Figure 7.
Parurohyal, in ventral view, of (A) Trichomycterus (Cryptocambeva) garbei Costa, Azevedo-Santos & Katz sp. nov.; (B) Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov.; (C) Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov.; (D) Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov.; (E) Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov.; and (F) Trichomycterus (Paracambeva) coelhorum Costa, Azevedo-Santos & Katz sp. nov.
Figure 7.
Parurohyal, in ventral view, of (A) Trichomycterus (Cryptocambeva) garbei Costa, Azevedo-Santos & Katz sp. nov.; (B) Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov.; (C) Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov.; (D) Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov.; (E) Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov.; and (F) Trichomycterus (Paracambeva) coelhorum Costa, Azevedo-Santos & Katz sp. nov.
Osteology: Mesethmoid slender, anterior margin slightly about straight, main axis gradually widening posteriorly. Mesethmoid cornu narrow, tip rounded. Postero-lateral margin of lateral ethmoid with pronounced projection towards middle portion of sesamoid supraorbital. Antorbital thin, small, drop-shaped, separated from sesamoid supraorbital by interspace larger than antorbital length. Sesamoid supraorbital slender, without lateral projections, its length about four times antorbital length. Premaxilla sub-trapezoidal in dorsal view, long, longer than maxilla. Maxilla slightly curved. Autopalatine sub-rectangular in dorsal view when excluding its postero-lateral process, its largest width about two thirds of its length including anterior cartilage; medial margin about straight. Autopalatine posterolateral process well-developed, short, its length about half autopalatine length excluding anterior cartilage. Metapterygoid subtriangular, deeper than long, dorsal extremity blunt, anterior margin convex, posterior portion without distinctive posterior projection. Quadrate compact, dorsal margin with weak projection posterior to articulation to metapterygoid, anterodorsal tip slightly projected dorsally, posterodorsal margin in contact with hyomandibula outgrowth. Hyomandibula long, with well-developed anterior outgrowth; dorsal margin of hyomandibula outgrowth with weak concavity on its posterior portion. Opercle long and slender, longer than interopercle. Opercular odontode patch slender, its width about half length of dorsal hyomandibula articular facet. Opercular odontodes 13–15, narrow, nearly straight, arranged in irregular transverse rows. Dorsal process of opercle short, subtriangular. Opercular articular facet for hyomandibula with rounded lateral shield, articular facet for preopercle small, rounded. Interopercle moderate in length, about three fourths longitudinal length of hyomandibula, anterior portion convex; dorsal process placed near anterior margin of interopercle. Interopercular odontodes 28–32, nearly straight, pointed, arranged in irregular longitudinal rows. Preopercle compact, with minute ventral projection. Parurohyal robust, lateral process truncate, posteriorly curved. Parurohyal head well-developed, with pronounced anterolateral paired process. Middle parurohyal foramen elliptical. Posterior process of parurohyal long, approximately equal to distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 8. Vertebrae 34 – 36. Ribs 12. Two dorsal hypural plates corresponding to hypurals 3 + 4 + 5; single ventral hypural plate corresponding to hypurals 1 + 2 + parhypural.
Colouration in alcohol: Flank, dorsum and head side pale brown, densely covered by dark brown dots with size variable among specimens, since about half to four times orbit diameter; dots irregularly arranged. Venter and ventral surface of head white. Barbels pale brown. Fins pale brown to hyaline on extremities, with dark brown dots on basal region.
Etymology. The name garbei is in honour of Ernst Garbe (1853–1925), a German naturalist who was Brazilian naturalized and who, between 1882 and the period just before his death, travelled through several Brazilian regions, including the Rio Grande drainage, making a rich biological collection. He collected the type specimens of Imparfinis longicauda Borodin, 1927 [=Heptapterus longicauda (Borodin, 1927)] in the type locality of T. garbei.
Distribution.
Trichomycterus garbei is only known from its type locality, a small stream tributary of the Rio Canoas, Rio Grande drainage, upper Rio Paraná basin, at about 715 m asl (
Figure 3B).
Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov.
LSID: urn:lsid:zoobank.org:act: A9BAED1D-5828-4545-B9F8-CCA9A97D9358
Holotype. UFRJ 11845, 55.1 mm SL; Brazil: Minas Gerais State: Capitólio Municipality: Córrego Tamborete, a tributary of Rio Grande at the Furnas dam, upper Rio Paraná basin, 20°38′53′′ S 46°09′55′′ W, about 875 m asl; A.M. Katz & P.H.N. Bragança, 14 February 2014.
Paratypes. UFRJ 10020, 4 ex., 30.8–56.2 mm SL; UFRJ 13345, 3 ex., 30.9–43.0 mm SL (C&S); CICCAA 07646, 2 ex., 42.7–42.9 mm SL; collected with holotype.
Diagnosis. Trichomycterus listruroides is distinguished from all other species of Cryptocambeva, except for T. candidus and Trichomycterus uberabensis Costa, Azevedo-Santos & Katz sp. nov., by the absence of pelvic fin and girdle (vs. presence). Trichomycterus listruroides differs from T. candidus and T. uberabensis Costa, Azevedo-Santos & Katz sp. Nov. by having the caudal fin margin aligned with the caudal peduncle, forming a spatula-like tail (vs. not aligned); more dorsal (24 or 25 vs. 17–20) and ventral (20 vs. 13–18) procurrent caudal-fin rays; fewer vertebrae (40 vs. 37–39); the dorsal-fin origin at the vertical between the centrum of the 24th vertebra (vs. 22nd or 23rd); the anal-fin origin at vertical through the centrum of the 26th or 27th vertebra (vs. 24th or 25th); and a vermiculate colour pattern on the flank (vs. minute dots in T. candidus and small round spots in T. uberabensis).
Description. General morphology: Morphometric data are presented in
Table 2. Body relatively slender, subcylindrical anteriorly, compressed posteriorly. Greatest body depth in area approximately at midway between pectoral-fin base and anal-fin origin. Dorsal and ventral profiles slightly convex between snout and anterior limit of caudal peduncle, slightly convex on caudal peduncle due to slight expansion on area supported by procurrent caudal-fin rays. Anus and urogenital papilla opening at vertical just posterior dorsal-fin origin. Head sub-trapezoidal in dorsal view, with anterior profile of snout convex. Eye small, dorsally positioned on head, nearer snout tip than posterior margin of opercle. Posterior nostril nearer anterior nostril than orbit. Tip of nasal barbel posteriorly reaching area just anterior to orbit; tip of maxillary barbel reaching between interopercular patch of odontodes and pectoral-fin base; rictal barbel reaching posterior portion of interopercular patch of odontodes. Mouth subterminal. Jaw teeth pointed, irregularly arranged, 19–26 on premaxilla, 22–25 on dentary. Minute skin papillae on snout and ventral surface of head. Branchial membrane attached to isthmus only at its anterior-most point, in ventral midline.
Figure 8.
Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov. Holotype, UFRJ 11845, Capitólio, 55.1 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Figure 8.
Trichomycterus (Cryptocambeva) listruroides Costa, Katz & Azevedo-Santos sp. nov. Holotype, UFRJ 11845, Capitólio, 55.1 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Dorsal and anal fins rounded, dorsal fin slender, longest ray shorter than anal-fin base length. Total dorsal-fin rays 11 (ii + II + 7), total anal-fin rays 9 (ii + II + 5). Anal-fin origin at vertical through middle portion of dorsal-fin base, or slightly anterior to it, at vertical through base of 5th segmented dorsal-fin ray. Dorsal-fin origin at vertical through centrum of 24th vertebra; anal-fin origin at vertical between centrum of 26th and 27th vertebra. Pectoral fin subtriangular in dorsal view, first pectoral-fin ray terminating in filament about 20–30 % pectoral-fin length excluding filament. Total pectoral-fin rays 6 (I + 5). Pelvic fin and girdle absent. Caudal fin rounded, continuous with caudal peduncle, forming spatula-shaped tail. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 24 or 25 (xxiii–xxiv + I), total ventral procurrent rays 20 (xix + I).
Table 2.
Morphometric data of Trichomycterus listruroides Costa, Katz & Azevedo-Santos sp. nov.
Table 2.
Morphometric data of Trichomycterus listruroides Costa, Katz & Azevedo-Santos sp. nov.
| | Holotype | Paratypes (n = 5) |
|---|
| | | |
| Standard length (SL) | 55.1 | 42.6–56.2 |
| | | |
Percentage of standard length | |
| Body depth | 18.1 | 11.3–18.1 |
| Caudal peduncle depth | 16.0 | 10.5–16.0 |
| Body width | 10.3 | 7.4–10.3 |
| Caudal peduncle width | 3.3 | 2.5–3.3 |
| Pre-dorsal length | 67.3 | 52.3–67.3 |
| Dorsal-fin base length | 11.8 | 9.1–11.8 |
| Anal-fin base length | 8.7 | 7.4–8.9 |
| Caudal-fin length | 20.3 | 15.1–20.7 |
| Pectoral-fin length | 11.1 | 7.6–11.1 |
| Head length | 17.2 | 13.2–17.2 |
| | | |
Percentage of head length | |
| Head depth | 54.7 | 43.2–54.7 |
| Head width | 84.2 | 64.2–84.2 |
| Snout length | 36.8 | 31.6–42.1 |
| Interorbital width | 28.4 | 22.1–28.4 |
| Preorbital length | 12.6 | 8.4–12.6 |
| Eye diameter | 8.4 | 7.3–9.3 |
Laterosensory system: Supraorbital, posterior section of infraorbital canal and postorbital canal continuous. Supraorbital sensory canal pores 3: s1, adjacent to medial margin of anterior nostril; s3, adjacent and just posterior to medial margin of posterior nostril; s6, in transverse line through posterior half of orbit; pore s6 approximately equidistant from its homologous pore and adjacent orbit. Anterior infraorbital sensory canal absent. Posterior infraorbital sensory canal pores 2: pore i10, adjacent to ventral margin of orbit, and pore i11, posterior to orbit. Postorbital canal pores 2: po1, at vertical through posterior portion of interopercular patch of odontodes, and po2, at vertical through posterior portion of opercular patch of odontodes. Lateral line pores 2; posterior-most pore at vertical just posterior to pectoral-fin base.
Osteology: Mesethmoid slender, anterior margin slightly concave, main axis gradually widening posteriorly. Mesethmoid cornu narrow, slightly curved posteriorly, extremity pointed. Postero-lateral margin of lateral ethmoid with small projection. Antorbital thin, small, drop-shaped, separated from sesamoid supraorbital by interspace larger than antorbital length. Sesamoid supraorbital slender, without lateral projections, its length about four times antorbital length. Premaxilla sub-trapezoidal in dorsal view, laterally tapering, shorter than maxilla. Maxilla slightly curved. Autopalatine sub-rectangular in dorsal view when excluding its postero-lateral process, its largest width about half its length including anterior cartilage; medial margin concave. Autopalatine posterolateral process well-developed, its length slightly larger than autopalatine largest width. Metapterygoid subtriangular to subtrapezoidal, longer than deep, dorsal extremity rounded or truncate, anterior margin convex, with small ventral projection, posterior portion with distinctive posterior projection. Quadrate compact, dorsal margin with weak projection posterior to articulation to metapterygoid, anterodorsal tip slightly projected dorsally, posterodorsal margin separated from hyomandibula outgrowth by small interspace. Hyomandibula long, with well-developed anterior outgrowth; dorsal margin of hyomandibula outgrowth with weak concavity. Opercle long and moderately slender, longer than interopercle. Opercular odontode patch slender, its width about two thirds of dorsal hyomandibula articular facet length. Opercular odontodes 9 or 10, narrow, nearly straight to slightly curved, irregularly arranged. Dorsal process of opercle short, slightly curved, extremity rounded. Opercular articular facet for hyomandibula with rounded lateral shield, articular facet for preopercle small, rounded. Interopercle moderate in length, about three fourths longitudinal length of hyomandibula, with stick-like anterior portion; dorsal process placed near anterior margin of interopercle, anterior socket for ligament connecting interopercle to lower jaw with small anterior projection. Interopercular odontodes 21 or 22, nearly straight, pointed, arranged in irregular longitudinal rows. Preopercle compact, with minute ventral projection. Parurohyal robust, lateral process pointed, slightly curved. Parurohyal head well-developed, with pronounced anterolateral paired process. Middle parurohyal foramen minute. Posterior process of parurohyal moderately long, about three fourths of distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 8. Vertebrae 40. Ribs 12 or 13. Two dorsal hypural plates corresponding to hypurals 3 + 4 + 5; single ventral hypural plate corresponding to hypurals 1 + 2 + parhypural.
Colouration in alcohol: Dorsal part of flank, dorsum and head side pale brown, ventral part of flank pale brownish yellow, with great concentration of minute, diffuse brown dots, their size about equal or smaller than orbit, sometimes more concentrated and darker along longitudinal midline of flank. Venter and ventral surface of head pale yellow. Barbels pale brown. Fins pale brown to hyaline on extremities.
Etymology. The name listruroides is an allusion to the superficial similarity of the new species with species of the catfish genus Listrura Pinna, 1988 (Trichomycteridae: Microcambevinae), including an elongate body, a rounded caudal fin that is continuous with the caudal peduncle forming a spatula-shaped tail, and an absence of pelvic fin and girdle.
Distribution.
Trichomycterus listruroides is only known from the type locality, the Córrego Tamborete, Rio Grande drainage, upper Rio Paraná basin, at about 875 m asl (
Figure 3B).
Trichomycterus (Cryptocambeva) pirabitira Barbosa & Azevedo-Santos, 2012
Trichomycterus pirabitira Barbosa & Azevedo-Santos, 2012: [
7] (358) (original description; type locality: Brazil: Estado de Minas Gerais: Município de Conceição da Aparecida: small stream between Carmo do rio Claro and Conceição da Aparecida, a tributary of the rio Grande, rio Paraná basin, approximately 21°8′53′′ S, 46°14′95′′ W, with altitude approximately 960 m [correctly: stream about 8.5 km SW of Conceição da Aparecida, 21°07′37′′ S, 46°16′59′′ W, about 965 m asl]; holotype: UFRJ 8335).
Diagnosis.
Trichomycterus pirabitira differs from all other congeners of the subgenus
Cryptocambeva by the presence of a foramen in the posterior portion of the dorsal process of the interopercle (
Figure 6C; vs. foramen absent,
Figure 6D).
Trichomycterus pirabitira is distinguished from other species of
Cryptocambeva, except for
Trichomycterus claudiae Barbosa & Costa, 2010,
Trichomycterus mirissumba Costa, 1992,
Trichomycterus mariamole Barbosa & Costa, 2010,
Trichomycterus saturatus Costa, Katz & Azevedo-Santos sp. Nov., and
Trichomycterus vermiculatus (Eigenmann, 1917), by a combination of the anterior segment of the infraorbital canal being present (vs. absent), seven pectoral-fin rays (vs. six), well-developed pelvic fin and girdle (vs. absent or rudimentary), and three procurrent dorsal-fin rays (vs. two).
Trichomycterus pirabitira differs from
T. claudiae,
T. mirissumba,
T. mariamole, and
T. vermiculatus by having a relatively robust metapterygoid, which is about as long as deep (
Figure 6C; vs. slender, deeper than long,
Figure 6D); from
T. claudiae,
T. mirissumba, and
T. vermiculatus by having 12–14 ribs (vs. 15 or 16); from
T. claudiae and
T. vermiculatus by having the anal-fin origin at vertical through the centrum of the 23rd or 24th vertebra (vs. 25th or 26th); from
T. claudiae by the absence of a longitudinal black stripe along the midline of the body side (vs. presence); from
T. mirissumba by having well-delimited spots on the flank (vs. spots diffuse); from
T. vermiculatus by having 36–38 vertebrae (vs. 39 or 40); and from
T. saturatus Costa, Katz & Azevedo-Santos sp. Nov. by having flank dots separated by interspaces that are larger than the dots (vs. flank with a great concentration of dark brown dots making the interspace areas smaller than the areas occupied by the dots), an absence of a rounded anterior projection in the interopercle (
Figure 6C; vs. presence,
Figure 6), a subrectangular basibranchial 3 (
Figure 10A; vs. goblet-shaped due to a strong posterior constriction,
Figure 10B), and a rudimentary hypobranchial 3 (
Figure 10A; vs. well-developed,
Figure 10B). For a full description and anatomical illustrations, see Barbosa and Azevedo-Santos [
7].
Figure 9.
Trichomycterus (Cryptocambeva) pirabitira, UFRJ 7279, Conceição da Aparecida, 65.7 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Figure 9.
Trichomycterus (Cryptocambeva) pirabitira, UFRJ 7279, Conceição da Aparecida, 65.7 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Distribution. Streams at the left margin of the Rio Grande, between the Rio São João and Rio Sapucaí subdrainages, and at altitudes between about 750 and 965 m asl (
Figure 3A).
Material examined. All specimens from Brazil: Minas Gerais State: Rio Paraná basin, Rio Grande drainage. Conceição da Aparecida Municipality: UFRJ 8335, holotype; UFRJ 8140, six paratypes; UFRJ 8264, three paratypes (C&S); a small stream about 8.5 km SW of Conceição da Aparecida, 21°07′37′′ S 46°16′59′′ W, at about 965 m asl; V.M. Azevedo-Santos, 30 April 2011. UFRJ 4927, 11 paratypes; UFRJ 7881, 1 ex. (C&S); a small stream tributary of Córrego Cuiabá, on the way to the village of Nova Resende; D. D’Almeida, M. Gonçalves & M.A. Barbosa, 24 September 1999. UFRJ 7279, 11 paratypes; UFRJ 5829, 3 paratypes (C&S); 7885, 1 ex. (C&S); same locality as UFRJ 4927; J.P.B. Barata, R. Paiva & M.A. Barbosa; 24 September 2006. UFRJ 9914, 7; Córrego Cuiabá, road Conceição da Aparecida-Nova Rezende, 21°05′22′′ S 46°13′43′′ W, at about 910 m asl; A. M. Katz & P. Bragança, 14 February 2014. Carmo do Rio Claro Municipality: UFRJ 8283, 7; Rio Pedra Molhada, 21°00′23′′ S 46°15′00′′ W, at about 855 m asl; V. M. Azevedo-Santos & I. C. Azevedo-Santos, 3 August 2011. UFRJ 12729, 8; Rio Pedra Molhada, 21°00′18′′ S 46°14′49′′ W, about 810 m asl; V. M. Azevedo-Santos & A. M. Katz, 30 October 2021. UFRJ 12853, 1; stream tributary of Ribeirão Itaci, 20°55′20′′ S 45°58′05′′ W, at about 795 m asl; V. M. Azevedo-Santos & A. M. Katz, 1 November 2021. Fortaleza de Minas Municipality: UFRJ 7273, 47 ex.; UFRJ 7884, 3 ex. (C&S); Riacho Fortaleza, road Jacuí-Fazenda Fortaleza, 20°53′57′′ S 46°44′09′′ W, at about 890 m asl; M. A. Barbosa, J. Prata & R. Paiva, 25 September 2006. Capetinga Municipality: UFRJ 8372, 4; stream tributary of Ribeirão Antinha, Rio Grande drainage, Rio Paraná basin, 20°38′23′′ S 46°59′02′′ W, at an altitude about 750 m asl; V.M. Azevedo-Santos, 21 August 2011.
Figure 10.
Ventral branchial arches, with left-central portion and in dorsal view: (A) Trichomycterus pirabitira; (B) Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov. Larger stippling represents cartilage.
Figure 10.
Ventral branchial arches, with left-central portion and in dorsal view: (A) Trichomycterus pirabitira; (B) Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov. Larger stippling represents cartilage.
Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. Nov.
LSID: urn:lsid:zoobank.org:act:31E2B7D1-CEEB-43C3-814D-A7F8AF0185CF
Holotype. UFRJ 13378, 85.8 mm SL; Brazil: Minas Gerais State: Capitólio Municipality: Córrego Tamborete, a tributary of Rio Grande at the Furnas dam, upper Rio Paraná basin, 20°38′53′′ S 46°09′55′′ W, at about 875 m asl; A.M. Katz & P.H.N. Bragança, 14 February 2014.
Paratypes. UFRJ 9912, 3 ex., 33.8–59.6 mm SL; UFRJ 13463, 3 ex., 37.1–70.3 mm SL (C&S); CICCAA 07647, 2 ex., 51.5–77.0 mm SL; collected with holotype. UFRJ 12720, 1 ex., 49.3 mm SL; same locality as holotype; V. M. Azevedo-Santos, 13 July 2017. UFRJ 12827, 1 ex., 116.3 mm SL; Córrego Tamborete, 20°38′38′′ S 46°10′13′′ W, at about 890 m asl; A. M. Katz & V. M. Azevedo-Santos, 2 November 2021.
Figure 11.
Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov. Holotype, UFRJ 13378, Capitólio, 85.8 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Figure 11.
Trichomycterus (Cryptocambeva) saturatus Costa, Katz & Azevedo-Santos sp. nov. Holotype, UFRJ 13378, Capitólio, 85.8 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Diagnosis.
Trichomycterus saturatus is distinguished from all other congeners of the subgenus
Cryptocambeva by having a goblet-shaped basibranchial 3 as a result of a strong posterior constriction (
Figure 10B; vs. subrectangular,
Figure 10A).
Trichomycterus saturatus is distinguished from other species of
Cryptocambeva, except for
T. claudiae,
T. mirissumba,
T. mariamole,
T. pirabitira, and
T. vermiculatus, by a combination of an anterior segment of the infraorbital canal being present (vs. absent), seven pectoral-fin rays (vs. six), well-developed pelvic fin and girdle (vs. absent or rudimentary), and three procurrent dorsal-fin rays (vs. two).
Trichomycterus saturatus differs from
T. claudiae,
T. mirissumba,
T. mariamole, and
T. vermiculatus by having a relatively robust metapterygoid, which is about as long as deep (
Figure 6E; vs. slender, deeper than long,
Figure 6D); from
T. claudiae,
T. mirissumba, and
T. vermiculatus by having 14 ribs (vs. 15 or 16); from
T. claudiae and
T. vermiculatus by having the anal-fin origin at vertical through the centrum of the 23rd or 24th vertebra (vs. 25th or 26th); from
T. claudiae by the absence of a longitudinal black stripe along the midline of the body side (vs. presence); from
T. mirissumba by having well-delimited spots on the flank (vs. spots diffuse); from
T. pirabitira by having the flank with a great concentration of dark brown dots making the interspace areas smaller than the areas occupied by the dots (vs. flank dots separated by interspaces that are larger than the dots), the absence of a foramen in the posterior portion of the dorsal process of the interopercle (vs. presence,
Figure 6C), the presence of a rounded anterior projection in the interopercle (
Figure 6E; vs. absence,
Figure 6C), and a well-developed hypobranchial 3 (
Figure 10B; vs. rudimentary,
Figure 10A).
Table 3.
Morphometric data of Trichomycterus saturatus Costa, Katz & Azevedo-Santos sp. nov.
Table 3.
Morphometric data of Trichomycterus saturatus Costa, Katz & Azevedo-Santos sp. nov.
| | Holotype | Paratypes (n = 7) |
|---|
| | | |
| Standard length (SL) | 85.8 | 40.2–116.3 |
| | | |
Percentage of standard length | |
| Body depth | 17.3 | 8.2–26.7 |
| Caudal peduncle depth | 14.3 | 7.1–22.4 |
| Body width | 11.6 | 5.4–18.3 |
| Caudal peduncle width | 3.9 | 1.9–6.9 |
| Pre-dorsal length | 61.9 | 28.4–86.1 |
| Pre-pelvic length | 55.8 | 28.7–81.1 |
| Dorsal-fin base length | 12.5 | 5.0–18.0 |
| Anal-fin base length | 10.6 | 9.2–14.0 |
| Caudal-fin length | 17.7 | 7.3–23.2 |
| Pectoral-fin length | 13.6 | 5.0–16.0 |
| Pelvic-fin length | 10.2 | 8.9–15.2 |
| Head length | 19.9 | 10.4–27.9 |
| | | |
Percentage of head length | |
| Head depth | 60.1 | 29.2–79.6 |
| Head width | 89.9 | 45.2–123.8 |
| Snout length | 43.6 | 22.5–64.4 |
| Interorbital width | 30.3 | 16.0–42.8 |
| Preorbital length | 13.3 | 5.5–20.3 |
| Eye diameter | 9.6 | 6.0–16.3 |
Description. General morphology: Morphometric data are presented in
Table 3. Body relatively deep, subcylindrical anteriorly, compressed posteriorly. Greatest body depth in area immediately anterior to pelvic-fin base. Dorsal and ventral profiles slightly convex between snout and anterior limit of caudal peduncle, nearly straight on caudal peduncle. Anus and urogenital papilla opening at vertical just anterior to middle portion of dorsal-fin base. Head sub-trapezoidal in dorsal view, with anterior profile of snout convex. Eye minute, dorsally positioned on head, nearer snout tip than posterior margin of opercle. Posterior nostril nearer anterior nostril than orbit. Tip of nasal barbel posteriorly reaching opercular patch of odontodes; tip of maxillary and rictal barbels reaching pectoral-fin base. Mouth subterminal. Jaw teeth pointed, irregularly arranged, 51–61 on premaxilla, 36–47 on dentary. Minute skin papillae on dorsal and ventral surfaces of head. Branchial membrane attached to isthmus only at its anterior-most point, in ventral midline.
Dorsal and anal fins subtriangular. Total dorsal-fin rays 11 (ii + II + 7), total anal-fin rays 10 (iii + II + 5). Anal-fin origin at vertical through posterior portion of dorsal-fin base, at vertical through base of 5th bifid dorsal-fin ray. Dorsal-fin origin at vertical through centrum of 19th or 20th vertebra; anal-fin origin at vertical through centrum of 23rd or 24th vertebra. Pectoral fin subtriangular in dorsal view, first pectoral-fin ray terminating in filament about 25–40 % pectoral-fin length excluding filament. Total pectoral-fin rays 7 (I + 6). Pelvic fin subtruncate, posteriorly overlapping anus and urogenital papilla, its posterior extremity at vertical just posterior to middle portion of dorsal-fin base. Pelvic-fin bases medially separated by minute interspace. Total pelvic-fin rays 5 (I + 4). Caudal fin truncate, posterior corners slightly rounded. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 21 (xx + I), total ventral procurrent rays 16 or 17 (xv–xvi + I).
Laterosensory system: Supraorbital, posterior section of infraorbital canal and postorbital canal continuous. Supraorbital sensory canal pores 3: s1, adjacent to medial margin of anterior nostril; s3, adjacent and just posterior to medial margin of posterior nostril; s6, in transverse line through posterior half of orbit; pore s6 slightly nearer its homologous pore than orbit. Infraorbital sensory canal arranged in 2 segments; anterior section isolated, with two pores: i1, at transverse line through anterior nostril, i3, at transverse line just anterior to posterior nostril; posterior segment posteriorly connected to supraorbital and postorbital canal, with 2 pores: i10, adjacent to ventral margin of orbit, i11, posterior to orbit. Postorbital canal pores 2: po1, at vertical through posterior portion of interopercular patch of odontodes, and po2, at vertical through posterior portion of opercular patch of odontodes. Lateral line pores 2; posterior-most pore at vertical just posterior to pectoral-fin base.
Osteology: Mesethmoid slender, anterior margin straight to slightly concave, main axis gradually widening posteriorly. Mesethmoid cornu narrow, tip rounded. Postero-lateral margin of lateral ethmoid with small projection. Antorbital thin, sub-elliptical, separated from sesamoid supraorbital by interspace shorter than antorbital length. Sesamoid supraorbital slender, without lateral projections, its length about two times antorbital length. Premaxilla sub-trapezoidal in dorsal view, laterally tapering, longer than maxilla. Maxilla slightly curved. Autopalatine sub-rectangular in dorsal view when excluding its postero-lateral process, its largest width about half its length including anterior cartilage; medial margin weakly concave. Autopalatine posterolateral process well-developed, its length slightly larger than autopalatine largest width. Metapterygoid subtriangular, deeper than long, dorsal extremity rounded, anterior margin convex, continuous, posterior portion without distinctive projection. Quadrate compact, dorsal margin with weak projection posterior to articulation to metapterygoid, posterodorsal margin separated from hyomandibula outgrowth by minute interspace. Hyomandibula long, with well-developed anterior outgrowth; dorsal margin of hyomandibula outgrowth with small concavity on its posterior portion. Opercle long and slender, slightly longer than interopercle. Opercular odontode patch slender, its width about one third of dorsal hyomandibula articular facet. Opercular odontodes 10–13, narrow, nearly straight, arranged in irregular oblique rows. Dorsal process of opercle short, slightly curved, extremity pointed. Opercular articular facet for hyomandibula with rounded lateral shield, articular facet for preopercle rudimentary, rounded. Interopercle moderate in length, about three fourths longitudinal length of hyomandibula, with rounded anterior projection; dorsal process posteriorly placed to anterior margin of interopercle. Interopercular odontodes 36 or 39, nearly straight, pointed, arranged in irregular longitudinal rows. Preopercle compact, with minute ventral projection. Basibranchial 2 robust, anterior portion slightly wider; basibranchial 3 goblet-shaped, with posterior constriction. Hypobranchial ossifications well-developed. Parurohyal robust, lateral process pointed, slightly curved. Parurohyal head well-developed, with almost indistinct anterolateral paired process. Middle parurohyal foramen small, rounded. Posterior process of parurohyal moderately long, about three fourths of distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 8 or 9. Vertebrae 37–39. Ribs 14. One or two dorsal hypural plates corresponding to hypurals 3 + 4 + 5; single ventral hypural plate corresponding to hypurals 1 + 2 + parhypural.
Colouration in alcohol: Flank with great concentration of dark brown dots over pale brownish yellow ground, making areas without dots smaller than areas occupied by dots; dots darker on dorsal portion of flank. Dorsum and dorsal surface of head brown with dots slightly darker than colour ground, venter and ventral surface of head light yellow. Barbels brown. Fins hyaline with brown dots on basal portion; brown chromatophores concentrate along rays of pectoral and unpaired fins. Smallest specimen (33.1 mm SL), with dots darker and highly coalesced along median longitudinal line of flank; dots smaller and more concentrated on dorsal portion of flank.
Etymology. From the Latin word saturatus (=saturated), its name is an allusion to the colour pattern consisting of numerous dark brown dots concentrated over the whole flank, making the interspaces smaller than the areas occupied by overlapped dots.
Distribution.
Trichomycterus saturatus is only known from its type locality, the Córrego Tamborete, Rio Grande drainage, Rio Paraná basin, at about 875 m asl (
Figure 3B).
Remarks. An examination of the gut content of the largest (70.3 mm SL) cleared and stained paratype (UFRJ 13463) revealed the presence of a chironomid larva, an unidentified insect larva, and a juvenile (about 20 mm SL) of Phalloceros sp. (Cypridontiformes: Poeciliidae), consisting of the first record of ichthyophagy in eastern South American trichomycterines.
Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov.
LSID: urn:lsid:zoobank.org:act: CADF9135-EEEB-4E46-A124-90F99B70EA3C
Holotype. UFRJ 13366, 42.3 mm SL; Brazil: Minas Gerais State: Veríssimo Municipality: stream tributary of Rio Uberaba, Rio Grande drainage, upper Rio Paraná basin, 19°39′21′′ S 48°15′21′′ W, at about 665 m asl; V.M. Azevedo-Santos et al., 15 April 2022.
Paratypes. UFRJ 12921, 8 ex., 23.3–39.9 mm SL; UFRJ 12957, 3 ex., 31.3–40.3 mm SL (C&S); CICCAA 07648, 2 ex., 31.9–32.7 mm SL; UFRJ 12894, 4 ex., 12.2–26.0 mm SL (DNA); collected with holotype.
Diagnosis. Trichomycterus uberabensis differs from all other congeners of Cryptocambeva, except for T. candidus and T. listruroides, by the absence of pelvic fin and girdle (vs. presence). Trichomycterus uberabensis is distinguished from T. candidus and T. listruroides by having fewer ventral procurrent caudal-fin rays (13–16 vs. 20), more teeth on the dentary (29–32 vs. 21–25), and a unique colour pattern on the flank, comprising small dark brown to black spots that are larger than the orbital diameter (vs. minute dark brown dots equal to or smaller than the orbital diameter in T. candidus, and small vermiculate marks in T. listruroides). Trichomycterus uberabensis also differs from T. listruroides by having the caudal-fin margin not aligned with the caudal peduncle, which is not dorsally expanded (vs. in a continuous line, with the dorsal margin of the caudal peduncle being slightly expanded), fewer dorsal procurrent caudal-fin rays (17–19 vs. 24 or 25), fewer vertebrae (37 or 38 vs. 40), the dorsal-fin origin at vertical through the centrum of the 22nd vertebra (vs. 24th), and the anal-fin origin at vertical through the centrum of the 24th or 25th vertebra (vs. 26th or 27th). It differs from T. candidus by having more interopercular odontodes (21–24 vs. 16–19).
Description. General morphology: Morphometric data are presented in
Table 4. Body slender, subcylindrical anteriorly, compressed posteriorly. Greatest body depth in area approximately at midway between pectoral-fin base and vertical through dorsal-fin origin. Dorsal and ventral profiles slightly convex between snout and anterior limit of caudal peduncle, about straight on caudal peduncle. Anus and urogenital papilla opening at vertical just posterior of dorsal-fin origin. Head sub-trapezoidal in dorsal view, with anterior profile of snout convex. Eye small, dorsally positioned on head, nearer snout tip than posterior margin of opercle. Posterior nostril nearer anterior nostril than orbit. Tip of nasal barbel posteriorly reaching between middle of opercle or area immediately posterior to it; tip of maxillary barbel reaching area between interopercle and pectoral-fin base, sometimes reaching pectoral-fin base; rictal barbel reaching between posterior portion of interopercular patch of odontodes and area between interopercle and pectoral-fin base. Mouth subterminal. Jaw teeth pointed, irregularly arranged, 26–28 on premaxilla, 29–32 on dentary. Minute skin papillae on dorsal and ventral surfaces of head. Branchial membrane attached to isthmus only at its anterior-most point, in ventral midline.
Figure 12.
Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov. Holotype, UFRJ 13366, Veríssimo, 42.3 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Figure 12.
Trichomycterus (Cryptocambeva) uberabensis Costa, Azevedo-Santos & Katz sp. nov. Holotype, UFRJ 13366, Veríssimo, 42.3 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Dorsal and anal fins slender, margin slightly convex. Total dorsal-fin rays 11 (ii + II + 7), total anal-fin rays 9 (ii + II + 5). Anal-fin origin at vertical through anterior half of dorsal-fin base, at vertical through base of 2nd or 3rd branched dorsal-fin ray. Dorsal-fin origin at vertical through centrum of 22nd vertebra; anal-fin origin at vertical between centrum of 24th or 25th vertebra. Pectoral fin subtriangular in dorsal view, first pectoral-fin ray terminating in short filament about 10–20 % pectoral-fin length excluding filament. Total pectoral-fin rays 6 (I + 5). Pelvic fin and girdle absent. Caudal fin rounded, nearly continuous with caudal peduncle. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 17–19 (xvi–xviii + I), total ventral procurrent rays 13–16 (xii–xv + I).
Laterosensory system: Supraorbital, posterior section of infraorbital canal and postorbital canal continuous. Supraorbital sensory canal pores 3: s1, adjacent to medial margin of anterior nostril; s3, adjacent and just posterior to medial margin of posterior nostril; s6, in transverse line through posterior half of orbit; pore s6 nearer adjacent orbit than its homologous pore pair. Anterior infraorbital sensory canal absent. Posterior infraorbital sensory canal pores 2: pore i10, adjacent to ventral margin of orbit, and pore i11, posterior to orbit. Postorbital canal pores 2: po1, at vertical through posterior portion of interopercular patch of odontodes, and po2, at vertical through posterior portion of opercular patch of odontodes. Lateral line pores 2; posterior-most pore at vertical just posterior to pectoral-fin base.
Table 4.
Morphometric data of Trichomycterus uberabensis Costa, Azevedo-Santos & Katz sp. nov.
Table 4.
Morphometric data of Trichomycterus uberabensis Costa, Azevedo-Santos & Katz sp. nov.
| | Holotype | Paratypes (n = 8) |
|---|
| | | |
| Standard length (SL) | 60.8 | 31.3–40.3 |
| | | |
Percentage of standard length | |
| Body depth | 14.9 | 12.1–14.6 |
| Caudal peduncle depth | 12.7 | 9.3–12.2 |
| Body width | 9.8 | 7.3–10.2 |
| Caudal peduncle width | 5.0 | 2.5–4.2 |
| Pre-dorsal length | 66.7 | 49.6–63.1 |
| Dorsal-fin base length | 11.5 | 8.4–11.2 |
| Anal-fin base length | 9.3 | 7.3–9.6 |
| Caudal-fin length | 19.1 | 14.4–18.7 |
| Pectoral-fin length | 11.8 | 9.3–12.4 |
| Head length | 17.7 | 14.0–17.7 |
| | | |
Percentage of head length | |
| Head depth | 53.3 | 49.7–55.1 |
| Head width | 84.1 | 77.2–87.0 |
| Snout length | 41.0 | 38.2–40.9 |
| Interorbital width | 23.0 | 19.3–26.7 |
| Preorbital length | 10.0 | 7.4–10.4 |
| Eye diameter | 11.6 | 8.6–18.9 |
Osteology: Mesethmoid slender, anterior margin slightly concave, main axis abruptly widening posteriorly. Mesethmoid cornu narrow, tip rounded. Postero-lateral margin of lateral ethmoid with inconspicuous projection. Antorbital thin, small, drop-shaped, separated from sesamoid supraorbital by interspace larger than antorbital length. Sesamoid supraorbital slender, without lateral projections, its length about three times antorbital length. Premaxilla sub-trapezoidal in dorsal view, laterally tapering, shorter than maxilla. Maxilla slightly curved. Autopalatine sub-rectangular in dorsal view when excluding its postero-lateral process, its largest width about half its length including anterior cartilage; medial margin concave. Autopalatine posterolateral process well-developed, its length slightly larger than autopalatine largest width. Metapterygoid subtriangular, deeper than long, dorsal extremity slightly pointed, anterior margin convex, posterior margin with concavity, posterior portion with distinctive posterior projection. Quadrate compact, dorsal margin with weak projection posterior to articulation to metapterygoid, anterodorsal tip slightly projected dorsally, posterodorsal margin separated from hyomandibula outgrowth by small interspace. Hyomandibula long, with well-developed anterior outgrowth; dorsal margin of hyomandibula outgrowth with weak concavity. Opercle long and moderately slender, longer than interopercle. Opercular odontode patch slender, its width about half length of dorsal hyomandibula articular facet. Opercular odontodes 8 or 9, narrow, nearly straight to slightly curved, irregularly arranged. Dorsal process of opercle short, slightly curved, extremity pointed. Opercular articular facet for hyomandibula with rounded lateral shield, articular facet for preopercle minute, rounded. Interopercle moderate in length, about three fourths longitudinal length of hyomandibula, with broad, rounded anterior portion; dorsal process placed near anterior margin of interopercle, anterior socket for ligament connecting interopercle to lower jaw with small anterior projection. Interopercular odontodes 21–24, nearly straight, pointed, arranged in irregular longitudinal rows. Preopercle compact, with small ventral projection. Parurohyal robust, lateral process pointed, nearly straight. Parurohyal head well-developed, with pronounced anterolateral paired process. Middle parurohyal foramen small, rounded. Posterior process of parurohyal moderately long, about four fifths of distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 7 or 8. Vertebrae 37 or 38. Ribs 11 or 12. Two dorsal hypural plates corresponding to hypurals 3 + 4 + 5; single ventral hypural plate corresponding to hypurals 1 + 2 + parhypural.
Colouration in alcohol: Flank, dorsum and head side brownish grey, with black dots, about equal or slightly larger than orbital diameter, sometimes coalesced on lateral midline of trunk. Venter and ventral surface of head white. Barbels pale brown. Fins hyaline, with dark brown dots on basal region.
Etymology. The name uberabensis is a reference to the occurrence of this new species in the Rio Uberaba drainage.
Distribution.
Trichomycterus uberabensis is only known from its type locality, a stream tributary of Rio Uberaba, Rio Grande drainage, upper Rio Paraná basin, at about 665 m asl (
Figure 3B).
3.2.2. Subgenus Paracambeva Costa, 2021
Species of Paracambeva are easily distinguished from species of other subgenera of Trichomycterus by the presence of a continuous, broad dark grey to black stripe between the snout and the caudal-fin base. All species of Paracambeva occurring in the upper Rio Paraná basin are members of a clade named T. reinhardti species group Costa & Katz, 2021, which are readily diagnosable by their infraorbital canal not being attached to the antorbital (Costa, 2021). Species of this group are commonly found in small shallow streams, being collected under marginal plants or in gravel substrates.
Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov.
LSID: urn:lsid:zoobank.org:act:07AF9548-552D-4DD7-8964-2A611CD8A70C
Trichomycterus septemradiatus non
Trichomycterus septemradiatus Katz, Barbosa & Costa, 2013: [
5] (5) in part (misidentification); [
11] (337) (misidentification).
Holotype. UFRJ 13406, 57.1 mm SL; Brazil: Minas Gerais State: Carmo do Rio Claro Municipality: stream tributary of Rio Itaci, a tributary of Rio Sapucaí, Rio Grande drainage, upper Rio Paraná basin, 20°54′58′′ S 45°56′21′′ W, at about 840 m asl; A. M. Katz & V.M. Azevedo-Santos, 31 October 2021.
Paratypes. UFRJ 13407, 3 ex., 33.9–50.2 mm SL; collected with holotype. UFRJ 13408, 2 ex., 46.2–49.0 mm SL (C&S); CICCAA 07649, 2 ex., 37.9–46.9 mm SL; same locality as holotype; V.M. Azevedo-Santos, 6 July 2021. UFRJ 13409, 6 ex., 24.5–56.5 mm SL; same locality as holotype; V.M. Azevedo-Santos, 14 July 2021.
Diagnosis. Trichomycterus adautoleitei differs from all other congeners of Paracambeva by having a long pectoral-fin filament, with its length about 40% of the pectoral-fin length excluding the filament (vs. about 20% or less), and some aspects of the colour pattern, including the presence of round black spots on the dorsum (vs. absence in all other species), the absence of a longitudinal series of dark brown to black dots on the dorsal part of the flank (vs. presence in all other species), and a longitudinal midline stripe that is well delimited, black, and highly contrasting with the yellow ground of the flank (vs. with diffuse margins and not highly contrasting with the colour ground of the flank in all other species, except for T. reinhardti (Eigenmann, 1917)). Trichomycterus adautoleitei is also distinguished from all other species of Paracambeva, except for Trichomycterus funebris Katz & Costa, 2021, Trichomycterus ingaiensis Katz & Costa, 2021, and Trichomycterus septemradiatus Katz, Barbosa & Costa, 2013, by having seven pectoral-fin rays (vs. six). Trichomycterus adautoleitei differs from T. ingaiensis and T. septemradiatus by having more ventral procurrent caudal-fin rays (13 vs. 8–11), and from T. funebris by having fewer vertebrae (35 or 36 vs. 37 or 38), the dorsal-fin origin at vertical through the centrum of the 19th or 20th vertebra (vs. 22nd), and the anal-fin origin at vertical through the centrum of the 22nd or 23rd vertebra (vs. 25th or 26th).
Figure 13.
Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov. Holotype, UFRJ 13406, Carmo do Rio Claro, 57.1 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Figure 13.
Trichomycterus (Paracambeva) adautoleitei Costa, Azevedo-Santos & Katz sp. nov. Holotype, UFRJ 13406, Carmo do Rio Claro, 57.1 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Description. General morphology: Morphometric data presented in
Table 5. Body relatively slender, subcylindrical on anterior region, compressed on posterior region. Greatest body depth in area immediately anterior to pelvic-fin base. Dorsal and ventral profiles slightly convex between snout and dorsal-fin base end, nearly straight on caudal peduncle. Anus and urogenital papilla opening at vertical just posterior to dorsal-fin base. Head sub-trapezoidal, with anterior profile of snout slightly convex in dorsal view. Eye small, dorsally positioned on head, nearer snout tip than posterior margin of opercle. Posterior nostril slightly nearer anterior nostril than orbital rim. Tip of nasal barbel posteriorly reaching opercular patch of odontodes, tip of maxillary barbel reaching pectoral-fin base, and tip of rictal barbel reaching area just posterior to interopercular patch of odontodes. Mouth subterminal. Jaw teeth pointed, irregularly arranged, 39–41 on premaxilla, 40–43 on dentary. Minute skin papillae on dorsal and ventral surfaces of head. Branchial membrane attached to isthmus only at its anterior-most point, in ventral midline.
Table 5.
Morphometric data of Trichomycterus adautoleitei Costa, Azevedo-Santos & Katz sp. nov.
Table 5.
Morphometric data of Trichomycterus adautoleitei Costa, Azevedo-Santos & Katz sp. nov.
| | Holotype | Paratypes (n = 5) |
|---|
| | | |
| Standard length (SL) | 57.1 | 43.3–56.5 |
| | | |
Percentage of standard length | |
| Body depth | 15.3 | 11.5–17.1 |
| Caudal peduncle depth | 11.7 | 9.0–11.8 |
| Body width | 11.6 | 7.2–9.7 |
| Caudal peduncle width | 4.5 | 2.0–4.1 |
| Pre-dorsal length | 65.0 | 52.9–58.5 |
| Pre-pelvic length | 60.6 | 48.0–61.8 |
| Dorsal-fin base length | 10.1 | 8.8–10.0 |
| Anal-fin base length | 8.0 | 7.3–8.2 |
| Caudal-fin length | 15.6 | 14.6–16.8 |
| Pectoral-fin length | 11.9 | 10.4–13.1 |
| Pelvic-fin length | 8.0 | 6.0–8.3 |
| Head length | 18.4 | 15.2–18.1 |
| | | |
Percentage of head length | |
| Head depth | 49.5 | 37.7–49.5 |
| Head width | 84.8 | 65.2–84.8 |
| Snout length | 42.8 | 34.1–42.8 |
| Interorbital width | 28.3 | 22.5–28.3 |
| Preorbital length | 8.7 | 8.7–9.8 |
| Eye diameter | 10.8 | 8.3–11.5 |
Dorsal and anal fins subtriangular. Total dorsal-fin rays 10 or 11 (ii + I–II + 7), total anal-fin rays 9 (ii + II + 5). Anal-fin origin at vertical just posterior to middle of dorsal-fin base, between base of 3rd and 4th branched dorsal-fin rays. Dorsal-fin origin at vertical through centrum of 19th or 20th vertebra; anal-fin origin at vertical through centrum of 22nd or 23rd vertebra. Pectoral fin subtriangular in dorsal view, first pectoral-fin ray terminating in filament about 40% of pectoral-fin length excluding filament. Total pectoral-fin rays 7 (I + 6). Pelvic fin rounded, its posterior extremity at vertical slightly posterior to dorsal-fin origin. Pelvic-fin bases medially separated by minute interspace. Total pelvic-fin rays 5 (I + 4). Caudal fin subtruncate, posterior corners rounded. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 16 (xv + I), total ventral procurrent rays 13 (ix–x + I).
Laterosensory system: Supraorbital, posterior section of infraorbital canal and postorbital canal continuous. Supraorbital sensory canal pores 3: s1, adjacent to medial margin of anterior nostril; s3, adjacent and just posterior to medial margin of posterior nostril; s6, in transverse line through posterior half of orbit; pore s6 nearer orbit than its homologous pore. Infraorbital sensory canal arranged in 2 segments; anterior section isolated, with two pores: i1, at transverse line through anterior nostril, i3, at transverse line just anterior to posterior nostril; posterior segment posteriorly connected to supraorbital and postorbital canal, with 2 pores: i10, adjacent to ventral margin of orbit, i11, posterior to orbit. Postorbital canal pores 2: po1, at vertical through posterior portion of interopercular patch of odontodes, and po2, at vertical through posterior portion of opercular patch of odontodes. Lateral line pores 2; posterior-most pore at vertical just posterior to pectoral-fin base.
Osteology: Mesethmoid slender, anterior margin approximately straight, main axis gradually widening posteriorly. Mesethmoid cornu narrow, tip rounded. Postero-lateral margin of lateral ethmoid without projections. Antorbital slender, thin, drop-shaped, separated from sesamoid supraorbital by interspace larger than antorbital length. Sesamoid supraorbital slender, without lateral projections, its length about two and half times antorbital length. Premaxilla sub-trapezoidal in dorsal view, laterally tapering, slightly longer than maxilla. Maxilla slightly curved. Autopalatine sub-rectangular in dorsal view when excluding its postero-lateral process, its largest width about half its length including anterior cartilage; medial margin weakly concave. Autopalatine posterolateral process well-developed, its length about half autopalatine length excluding anterior cartilage. Metapterygoid subtrapezoidal, about so deep as long, dorsal extremity truncate, anterior margin slightly convex, posterior margin slightly concave, posterior portion with small posterior projection. Quadrate compact, dorsal margin with weak projection posterior to articulation to metapterygoid, posterodorsal margin separated from hyomandibula outgrowth. Hyomandibula long, with well-developed anterior outgrowth; dorsal margin of hyomandibula outgrowth with deep concavity. Opercle long and moderately slender, longer than interopercle. Opercular odontode patch slender, its width about three fifths of dorsal hyomandibula articular facet. Opercular odontodes 13–15, narrow, about straight, irregularly arranged. Dorsal process of opercle short, slightly curved, dorsally terminating in stick-like tip. Opercular articular facet for hyomandibula with rounded lateral shield, articular facet for preopercle small, rounded. Interopercle moderate in length, about three fourths longitudinal length of hyomandibula, with convex anterior portion; dorsal process placed near anterior margin of interopercle. Interopercular odontodes 33–35, nearly straight, tip pointed to slightly rounded, arranged in irregular longitudinal rows. Preopercle compact, with minute ventral projection. Parurohyal robust, lateral process pointed, nearly straight. Parurohyal head well-developed, with pronounced anterolateral paired process. Middle parurohyal foramen minute. Posterior process of parurohyal short, about half distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 8. Vertebrae 35 or 36. Ribs 15. Two dorsal hypural plates corresponding to hypurals 3 + 4 + 5; single ventral hypural plate corresponding to hypurals 1 + 2 + parhypural.
Colouration in alcohol: Flank, dorsum and head side dark yellow, to slightly lighter on ventral part of flank, with black dots on dorsum, black stripe along longitudinal midline of flank, and sometimes few black dots on posterior portion of caudal peduncle. Dark chromatophores concentrated on area between maxillary barbel and longitudinal flank stripe, on area between middle portion of dorsal surface of head and nape, and in oblique zone anterior to opercular and interopercular patches of odontodes; black dots on snout and barbels. Venter and ventral surface of head light yellow. Fins hyaline, with minute black dots on whole dorsal and caudal fins, and on first pectoral-fin ray, and diffuse grey stripe on caudal fin longitudinal midline.
Etymology. It is named in honour of Antônio Adauto Leite (1927–2020), the founder of the Museum of Indigenous Archaeology of Carmo do Rio Claro (MUARI), which has a rich collection of archaeological pieces between 2000 and 12,000 years old, mainly found in the region of the type locality of the new species.
Distribution.
Trichomycterus adautoleitei is only known from its type locality, a stream tributary of Rio Itaci, a tributary of Rio Sapucaí, Rio Grande drainage, upper Rio Paraná basin, at about 840 m asl (
Figure 3C).
Trichomycterus (Paracambeva) coelhorum Costa, Azevedo-Santos & Katz sp. nov.
LSID: urn:lsid:zoobank.org:act:6BCFCCC8-60AE-423D-8EF2-189469BFBE0E
Holotype. UFRJ 13410, 44.8 mm SL; Brazil: Minas Gerais State: Veríssimo Municipality: stream tributary of Rio Uberaba, Rio Grande drainage, upper Rio Paraná basin, 19°39′21′′ S 48°15′21′′ W, at about 665 m asl; V. M. de Azevedo Santos et al., 15 April 2022.
Paratypes. UFRJ 12916, 5 ex., 30.7–39.3 mm SL; UFRJ 12961, 3 ex., 34.9–38.8 mm SL (C&S); CICCAA 07650, 4 ex., 32.2–36.8 mm SL; collected with holotype.
Diagnosis. Trichomycterus coelhorum is distinguished from all other congeners of Paracambeva by having fewer dorsal procurrent caudal-fin rays (10–12 vs. 13–25). Trichomycterus coelhorum is also distinguished from all other congeners of Paracambeva, except for T. humboldti Costa & Katz, 2021, Trichomycterus pauciradiatus Alencar & Costa, 2006, Trichomycterus piratymbara Katz, Barbosa & Costa, 2013, T. reinhardti, and T. sainthilairei, by having six pectoral-fin rays. Trichomycterus coelhorum differs from T. humboldti, T. pauciradiatus, T. piratymbara, T. reinhardti, and T. sainthilairei by having fewer ventral procurrent caudal-fin rays (9 or 10 vs. 11–18); from T. piratymbara, T. reinhardti, and T. sainthilairei by having fewer vertebrae (34–36 vs, 37–40); from T. piratymbara, T. humboldti, and T. pauciradiatus by having fewer ribs (12–14 vs. 15–19); and from T. humboldti and T. pauciradiatus by having five pelvic-fin rays (vs. four).
Table 6.
Morphometric data of Trichomycterus coelhorum Costa, Azevedo-Santos & Katz sp. nov.
Table 6.
Morphometric data of Trichomycterus coelhorum Costa, Azevedo-Santos & Katz sp. nov.
| | Holotype | Paratypes (n = 10) |
|---|
| | | |
| Standard length (SL) | 60.8 | 34.4–38.8 |
| | | |
Percentage of standard length | |
| Body depth | 15.2 | 11.1–15.1 |
| Caudal peduncle depth | 10.6 | 8.7–10.2 |
| Body width | 9.4 | 7.6–9.9 |
| Caudal peduncle width | 3.6 | 2.5–3.5 |
| Pre-dorsal length | 62.6 | 51.5–59.7 |
| Pre-pelvic length | 61.7 | 47.5–54.3 |
| Dorsal-fin base length | 11.7 | 8.5–11.2 |
| Anal-fin base length | 7.8 | 6.7–8.3 |
| Caudal-fin length | 17.3 | 13.4–19.9 |
| Pectoral-fin length | 11.4 | 9.0–12.2 |
| Pelvic-fin length | 8.4 | 6.7–8.0 |
| Head length | 18.8 | 15.0–17.0 |
| | | |
Percentage of head length | |
| Head depth | 52.3 | 42.8–51.1 |
| Head width | 81.2 | 66.5–76.0 |
| Snout length | 39.2 | 30.9–38.0 |
| Interorbital width | 27.6 | 22.6–24.9 |
| Preorbital length | 8.3 | 7.1–10.7 |
| Eye diameter | 11.9 | 8.3–10.7 |
Description. General morphology: Morphometric data are presented in
Table 6. Body relatively deep, subcylindrical anteriorly, compressed posteriorly. Greatest body depth in area immediately anterior to pelvic-fin base. Dorsal and ventral profiles slightly convex between snout and anterior limit of caudal peduncle, nearly straight on caudal peduncle. Anus and urogenital papilla opening at vertical through anterior third of dorsal-fin base. Head sub-trapezoidal in dorsal view, with anterior profile of snout convex. Eye minute, dorsally positioned on head, nearer snout tip than posterior margin of opercle. Posterior nostril nearer anterior nostril than orbit. Tip of nasal barbel posteriorly reaching opercular patch of odontodes or area just anterior to it; tip of maxillary barbel reaching between posterior portion of interopercular patch of odontodes and pectoral-fin base; rictal barbel reaching between middle and posterior portions of interopercular patch of odontodes. Mouth subterminal. Jaw teeth pointed, irregularly arranged, 30–34 on premaxilla, 29–31 on dentary. Minute skin papillae on dorsal and ventral surfaces of head. Branchial membrane attached to isthmus only at its anterior-most point, in ventral midline.
Dorsal and anal fins subtriangular. Total dorsal-fin rays 11 (ii + II–III + 5–6 + I), total anal-fin rays 9 (ii + II + 4 + I). Anal-fin origin at vertical through middle portion of dorsal-fin base, at vertical through base of 5th or 6th segmented dorsal-fin ray. Dorsal-fin origin at vertical through centrum of 19th or 20th vertebra; anal-fin origin at vertical through centrum of 19th or 20th vertebra. Pectoral fin subtriangular in dorsal view, first pectoral-fin ray terminating in minute filament about 5–10 % of pectoral-fin length excluding filament. Total pectoral-fin rays 6 (I + 5). Pelvic fin rounded, posteriorly overlapping anus but not reaching urogenital papilla, its posterior extremity at vertical through base of first bifid dorsal-fin ray. Pelvic-fin bases medially separated by minute interspace. Total pelvic-fin rays 5 (I + 4). Caudal fin subtruncate, posterior corners rounded. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 10 – 12 (ix–xi + I), total ventral procurrent rays 9 or 10 (viii–ix + I).
Laterosensory system: Supraorbital, posterior section of infraorbital canal and postorbital canal continuous. Supraorbital sensory canal pores 3: s1, adjacent to medial margin of anterior nostril; s3, adjacent and just posterior to medial margin of posterior nostril; s6, in transverse line through posterior half of orbit; pore s6 nearer orbit than its homologous pore. Infraorbital sensory canal arranged in 2 segments; anterior section isolated, with two pores: i1, at transverse line through anterior nostril, i3, at transverse line just anterior to posterior nostril; posterior segment posteriorly connected to supraorbital and postorbital canal, with 2 pores: i10, adjacent to ventral margin of orbit, i11, posterior to orbit. Postorbital canal pores 2: po1, at vertical through posterior portion of interopercular patch of odontodes, and po2, at vertical through posterior portion of opercular patch of odontodes. Lateral line pores 2; posterior-most pore at vertical just posterior to pectoral-fin base.
Osteology: Mesethmoid slender, anterior margin approximately straight, main axis gradually widening posteriorly. Mesethmoid cornu narrow, tip rounded. Postero-lateral margin of lateral ethmoid without projections. Antorbital thin, drop-shaped, separated from sesamoid supraorbital by interspace larger than antorbital length. Sesamoid supraorbital slender, without lateral projections, short, its length about two times antorbital length. Premaxilla sub-trapezoidal in dorsal view, slightly longer than maxilla. Maxilla slightly curved. Autopalatine sub-rectangular in dorsal view when excluding its postero-lateral process, its largest width about three fourths of its length including anterior cartilage; medial margin weakly concave. Autopalatine posterolateral process well-developed, its length about equal autopalatine largest width. Metapterygoid subtriangular, deeper than long, dorsal extremity pointed, anterior convex, posterior margin concave, posterior portion with small posterior projection. Quadrate compact, dorsal margin with weak projection posterior to articulation to metapterygoid, posterodorsal margin in contact with hyomandibula outgrowth. Hyomandibula long, with well-developed anterior outgrowth; dorsal margin of hyomandibula gently concave. Opercle long and moderately slender, longer than interopercle. Opercular odontode patch slender, its width about three fifths of dorsal hyomandibula articular facet. Opercular odontodes 18–20, narrow, about straight, irregularly arranged. Dorsal process of opercle short, subtriangular. Opercular articular facet for hyomandibula with rounded lateral shield, articular facet for preopercle small, rounded. Interopercle short, about half longitudinal length of hyomandibula, with convex anterior portion; dorsal process placed nearly continuous with anterior margin of interopercle. Interopercular odontodes 26–28, nearly straight, tip pointed, arranged in irregular longitudinal rows. Preopercle compact, with minute ventral projection. Parurohyal robust, lateral process pointed, nearly straight to slightly curved. Parurohyal head well-developed, with pronounced anterolateral paired process. Middle parurohyal foramen large, elliptical. Posterior process of parurohyal short, about half distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 8. Vertebrae 35 or 36. Ribs 15. Two dorsal hypural plates corresponding to hypurals 3 + 4 + 5; single ventral hypural plate corresponding to hypurals 1 + 2 + parhypural.
Colouration in alcohol: Flank pale yellow, with dark brown to black stripe along longitudinal midline of flank and black dots above stripe, more concentrated on caudal peduncle; often faint minute brown dots on ventral portion of caudal peduncle side and above venter. Dorsum pale brown, with two longitudinal dark brown to black stripes and minute black dots between stripes. Lateral and dorsal surfaces of head pale yellow with brown dots. Venter and ventral surface of head yellowish white. Barbels brown. Fins hyaline, with minute black dots on whole dorsal and caudal fins, and on first pectoral-fin ray; diffuse grey stripe on caudal fin longitudinal midline.
Etymology. The name coelhorum is in honour of the Coelho family from Frutal Municipality in Minas Gerais State. This is in recognition of the help of the zoologist Paula Nunes Coelho with the type series and her family for the logistical support during studies in the region.
Distribution.
Trichomycterus coelhorum is only known from its type locality, a stream tributary of the Rio Uberaba, Rio Grande drainage, upper Rio Paraná basin, at about 665 m asl (
Figure 3C).
Trichomycterus (Paracambeva) sainthilairei Katz & Costa, 2021
Trichomycterus piratymbara non
Trichomycterus piratymbara Katz & Costa, 2013: [
42] (238) (misidentification).
Trichomycterus sainthilairei Katz & Costa, 2021: [
1] (17) (original description; type locality: Brazil: Estado de Minas Gerais: Município de Capitólio: Córrego Tamborete, Rio Grande drainage, Rio Paraná basin, 20°38′53′′ S 46°09′55′′ W, at an altitude about 875 m asl; holotype: UFRJ 10215).
Diagnosis. Trichomycterus sainthilairei is distinguished from all other species of Paracambeva, except for T. coelhorum, T. humboldti, T. pauciradiatus, T. piratymbara, and T. reinhardti, by having six pectoral-fin rays (vs. seven). Trichomycterus sainthilairei is distinguished from T. coelhorum by having more dorsal (17–20 vs. 10–12) and ventral (13–16 vs. 9 or 10) procurrent caudal-fin rays, more vertebrae (37 or 38 vs. 34–36), and more ribs (16 or 17 vs. 12–14); from T. humboldti and T. pauciradiatus by having five pelvic-fin rays (vs. four); from T. piratymbara by having the anal-fin origin at vertical through the posterior half of the dorsal-fin base (vs. anterior half); and from T. reinhardti by having fewer dorsal procurrent caudal-fin rays (17–20 vs. 22–25). For a full description and anatomical illustrations, see Costa and Katz (2021).
Distribution. Trichomycterus sainthilairei is known only from the type locality, in the Córrego do Tamborete, Rio Grande drainage, upper Rio Paraná basin, at about 875 m asl.
Material examined. All materials were from Brazil: Estado de Minas Gerais: Município de Capitólio: Córrego Tamborete, Rio Grande drainage, Rio Paraná basin. UFRJ 10215, holotype; UFRJ 9886, six paratypes; UFRJ 9911, six paratypes; UFRJ 9971, five paratypes (C&S); CICCAA 04085, three paratypes; Brazil: Minas Gerais State: Capitólio Municipality: 20°38′53′′ S 46°09′55′′ W, at about 875 m asl; A.M. Katz & P.H.N. Bragança, 14 February 2014. MZUSP 87188, 15 paratypes; same locality as holotype; C. Oliveira, 15 April 2005. MNRJ 28660, 33 paratypes; Km 300 of the road MG-050; P.A. Buckup et al., 6 November 2004.
Trichomycterus (Paracambeva) septemradiatus Katz, Barbosa & Costa, 2013
Trichomycterus septemradiatus Katz, Barbosa & Costa, 2013: [
8] (360) (original description; type locality: stream tributary of Cuiabá river, tributary of Grande river [correctly tributary of Rio Sapucaí], Paraná river basin, between villages Conceição da Aparecida and Nova Resende [correctly road Fazenda Cuiabá], Município de Conceição da Aparecida, Estado de Minas Gerais, Brazil, 21°04′24′′ S 46°12′04′′ W, at an altitude of 856 m; holotype: UFRJ 8576, 45.4 mm SL).
Figure 15.
Trichomycterus (Paracambeva) sainthilairei. Holotype, UFRJ 10215, Capitólio, 65.4 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Figure 15.
Trichomycterus (Paracambeva) sainthilairei. Holotype, UFRJ 10215, Capitólio, 65.4 mm SL: (A) lateral, (B) dorsal, and (C) ventral views.
Diagnosis. Trichomycterus septemradiatus is distinguished from all other species of Paracambeva, except for T. adautoleitei, T. anaisae, T. funebris, T. ingaiensis, and T. luetkeni by having seven pectoral-fin rays (vs. six in all other species of the T. reinhardti group). Trichomycterus septemradiatus differs from T. adautoleitei by having fewer ventral procurrent caudal-fin rays (10 vs. 13), a longitudinal body stripe with diffuse margins, which is weakly contrasting with the pale yellow ground of the flank (vs. well delimited black stripe, strongly contrasting with the yellow ground of the flank), the absence of round black spots on the dorsum body (vs. presence), and the presence of a longitudinal row of small black spots on the dorsal portion of the flank (vs. absence); from T. anaisae and T. luetkeni by having fewer dorsal (15 vs. 21–23) and ventral (10 vs. 15–17) procurrent caudal-fin rays; from T. anaisae by the absence of two rows of black spots overlapping the longitudinal body stripe (vs. presence); from T. funebris by having a shorter latero-posterior process of the autopalatine, with its length being about half of the autopalatine length (vs. about two-thirds), and the anal-fin origin at vertical through the centrum of the 23rd vertebra (vs. 25th or 26th); and from T. ingaiensis by having the anal-fin origin at vertical between the base of the fifth dorsal-fin ray (vs. base of the seventh and ninth dorsal-fin rays). For a full description and anatomical illustrations, see Costa and Katz (2021).
Distribution. Trichomycterus septemradiatus occurs in the Rio Sapucaí subdrainage, Rio Grande drainage, Rio Paraná basin, at altitudes between about 855 and 1040 m asl (
Figure 3C).
Material examined. All specimens were from Brazil: Estado de Minas Gerais: Rio Grande drainage, Rio Paraná basin. Município de Conceição da Aparecida. UFRJ 8576, holotype, 45.4 mm SL; UFRJ 7278, 11 paratypes; UFRJ 8385, 3 paratypes (C&S); stream tributary to Rio Cuiabá, Rio Sapucaí subdrainage, road Fazenda Cuiabá, 21°04′24′′ S 46°12′04′′ O, about 855 m asl; J.P.B. Barata, R. Paiva & M.A. Barbosa, 24 November 2006. UFRJ 5088, 1; Córrego Cuiabá, road Fazenda Cuiabá, about 2 km from Conceição da Aparecida, 21°05′22′′ S 46°13′43′′ W, about 910 m asl; M.A. Barbosa et al., 16 February 2000. UFRJ 9888, 5; UFRJ 9913, 2; same locality as UFRJ 5088; A. M. Katz & P.H. Bragança, 14 February 2014. Município de Carmo do Rio Claro: UFRJ 8283, 6; UFRJ 8373, 4; UFRJ 9689, 2 (C&S); UFRJ 9690, 1 ex (C&S); Ribeirão Santa Quitéria, Cachoeira Pedra Molhada, 21°00′23′′ S 46°15′00′′ W, about 855 m asl; V.M. Azevedo-Santos, 3 Aug. 2011. UFRJ 12940,
Figure 16, 6 ex.; same locality as preceding; A.M. Katz & V.M. Azevedo-Santos, 30 October 2021.



 ,
, 




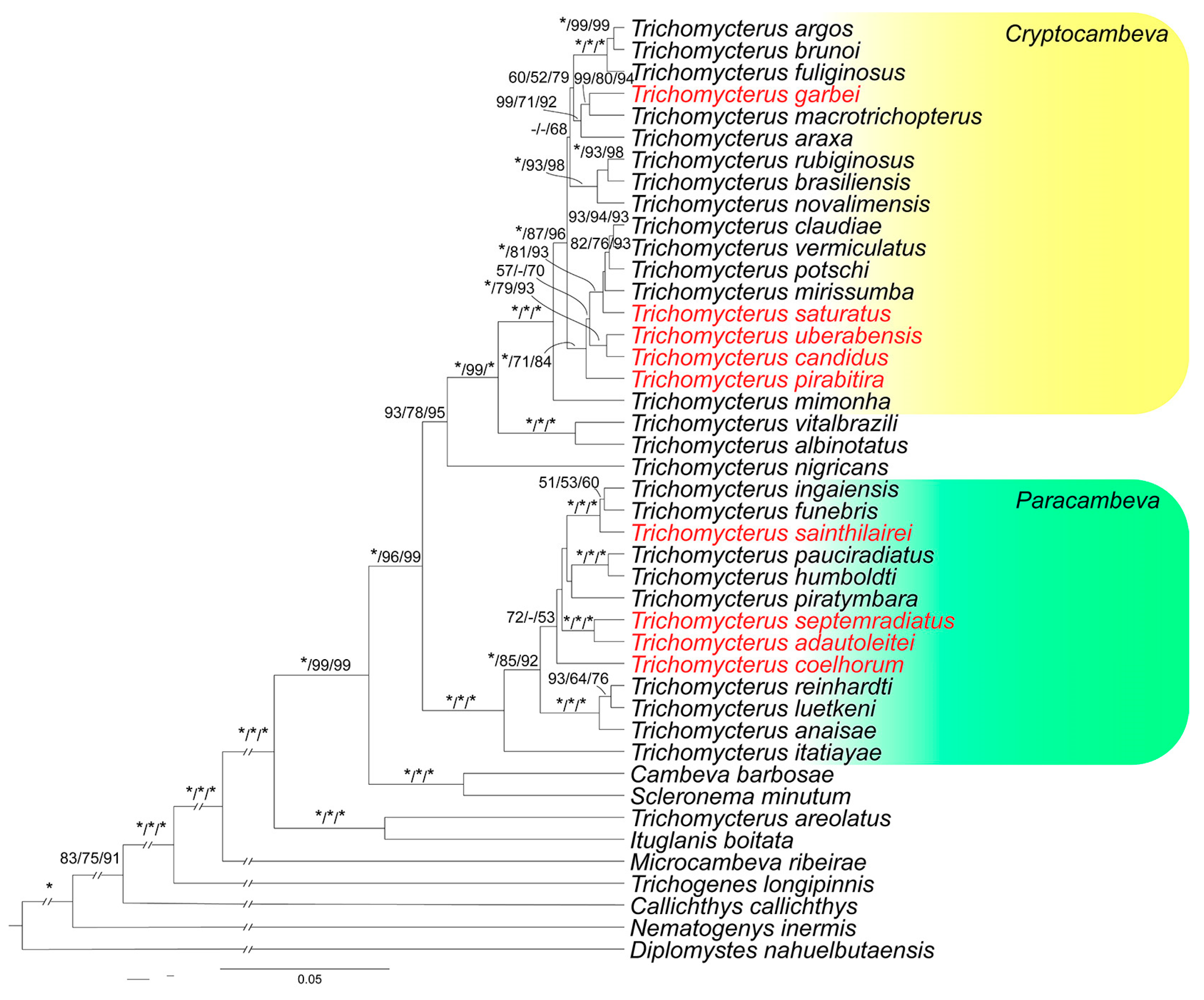{kind=link}
{kind=link}
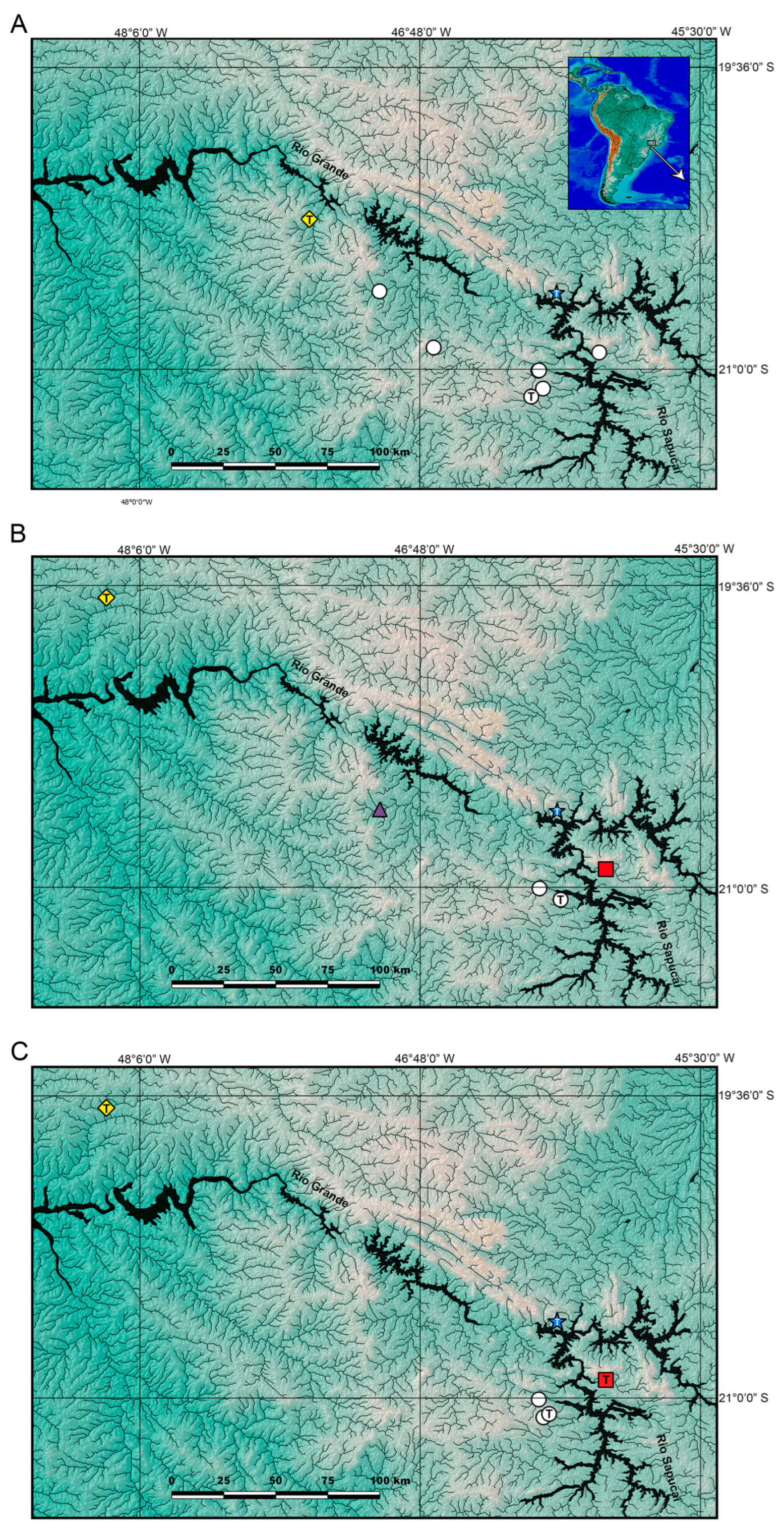{kind=link}
{kind=link}
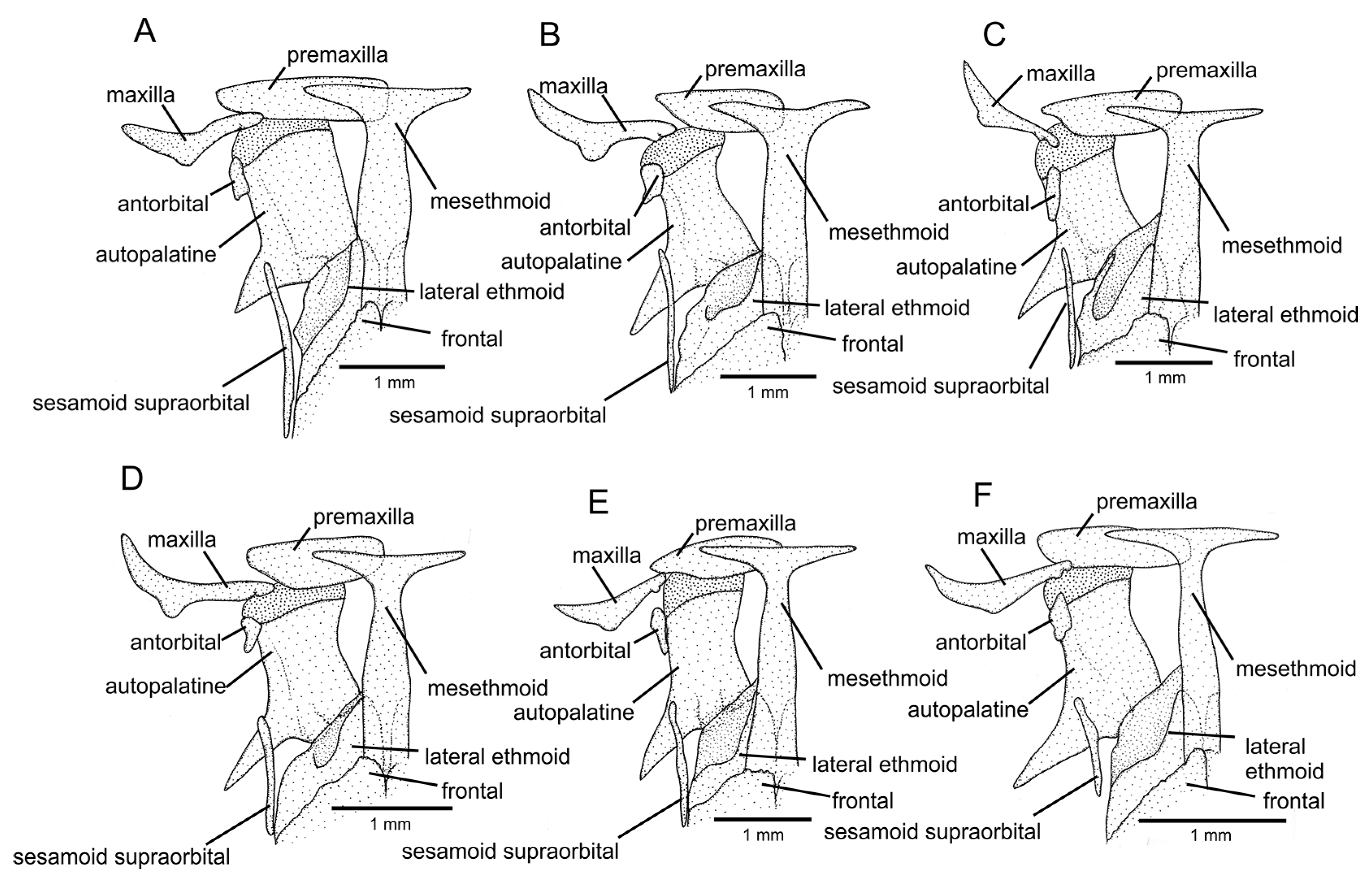{kind=link}
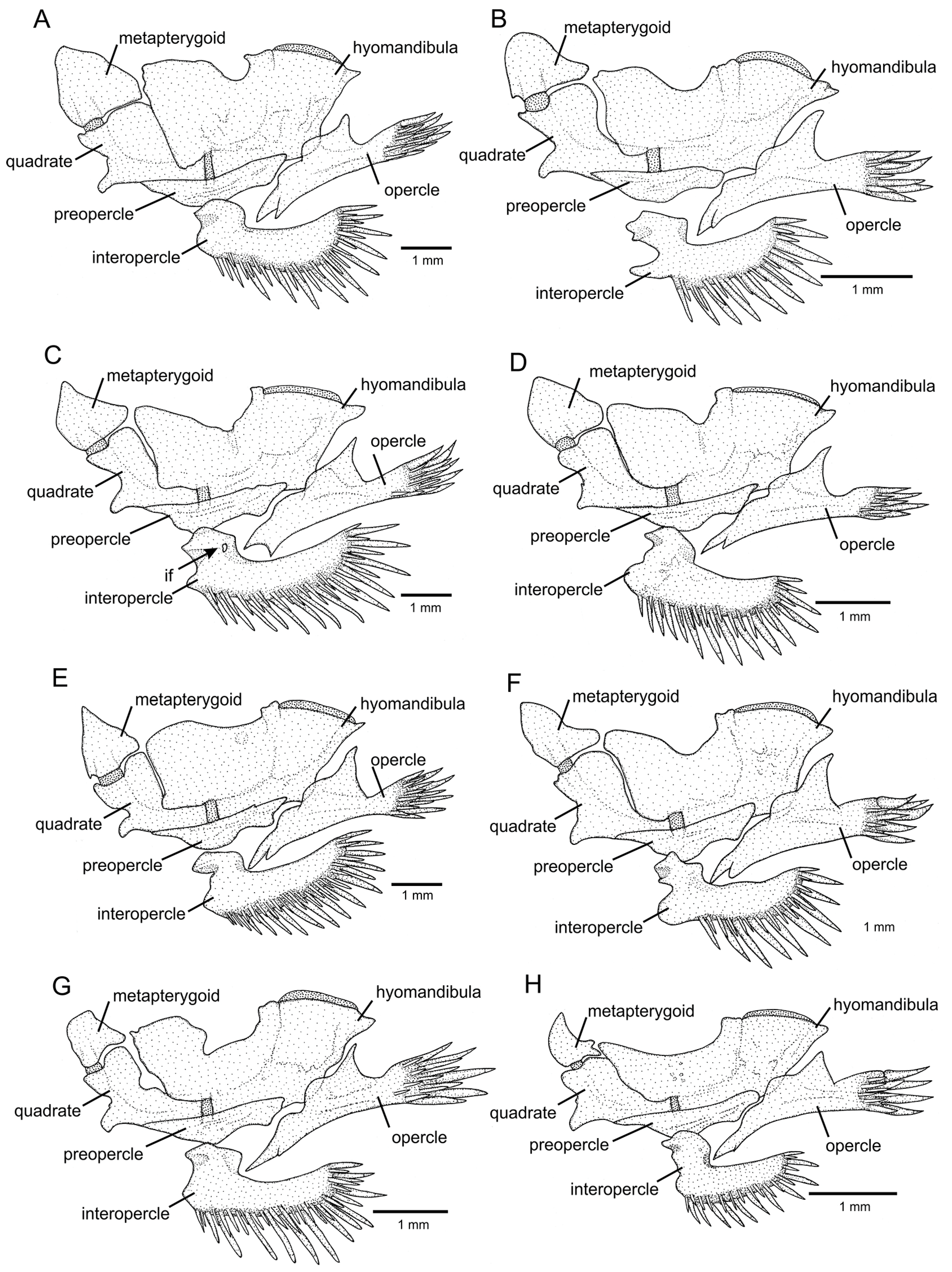{kind=link}
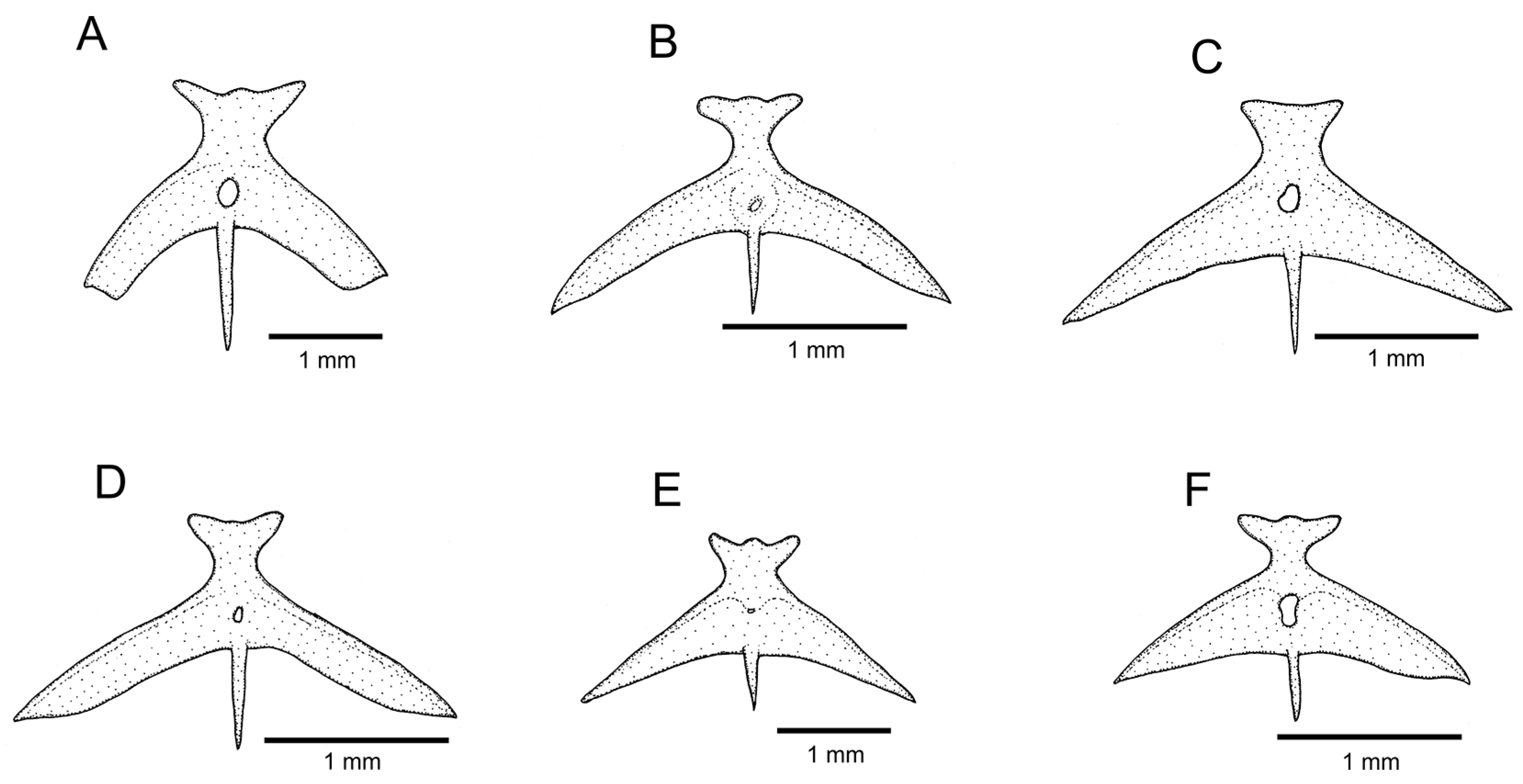{kind=link}
{kind=link}
{kind=link}
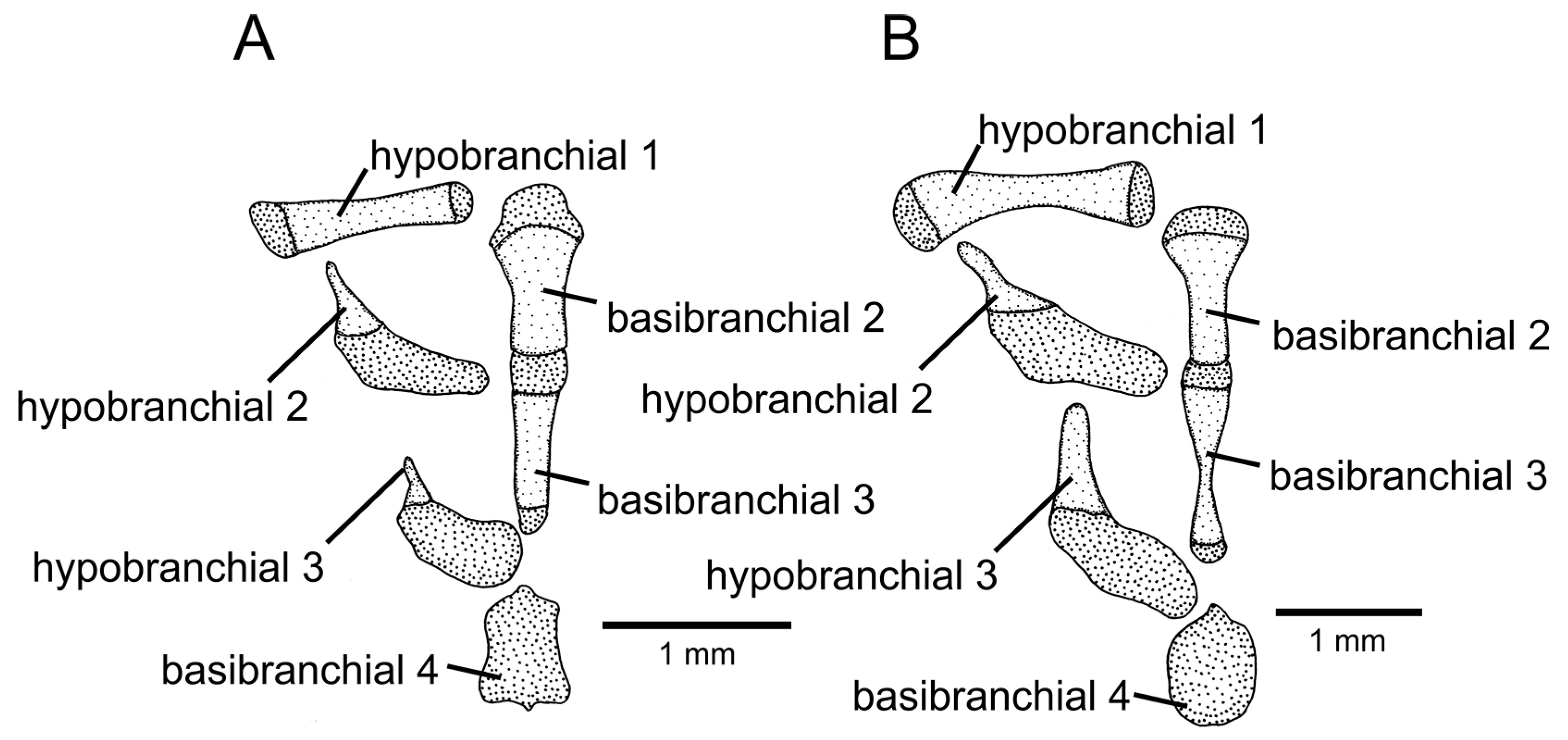{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












