

Cytotoxic Effects of the Atrazine Herbicide on Erythrocytes and Liver Damage in Lithobates spectabilis

 ,
,  , , , ,
, , , ,  and
and 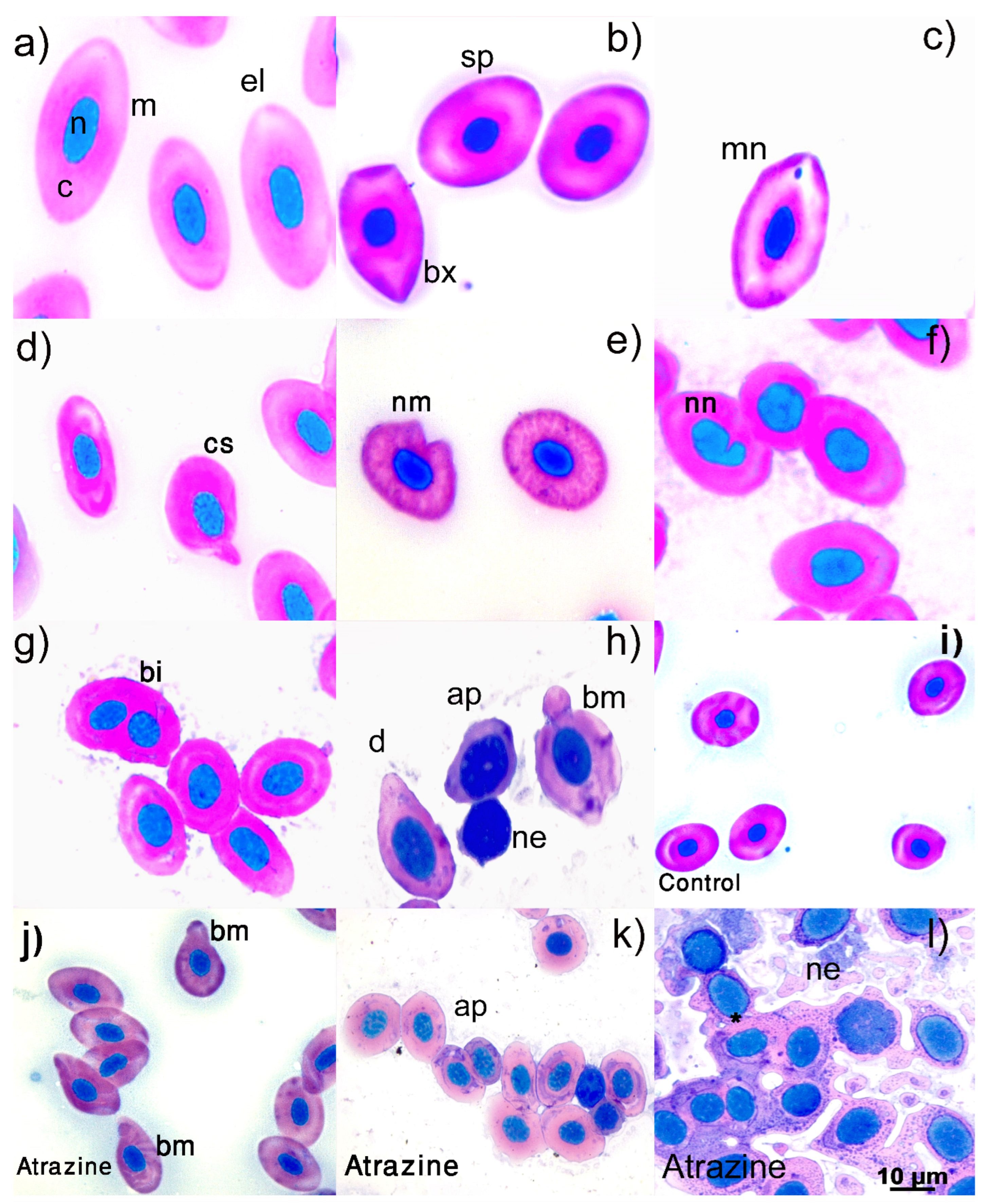{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Lithobates spectabilis
2.2. Capture Site
2.3. Experimental Design
Animal Housing Conditions
2.4. Phase 1: Acclimatization
2.5. Phase 2: Atrazine Exposure
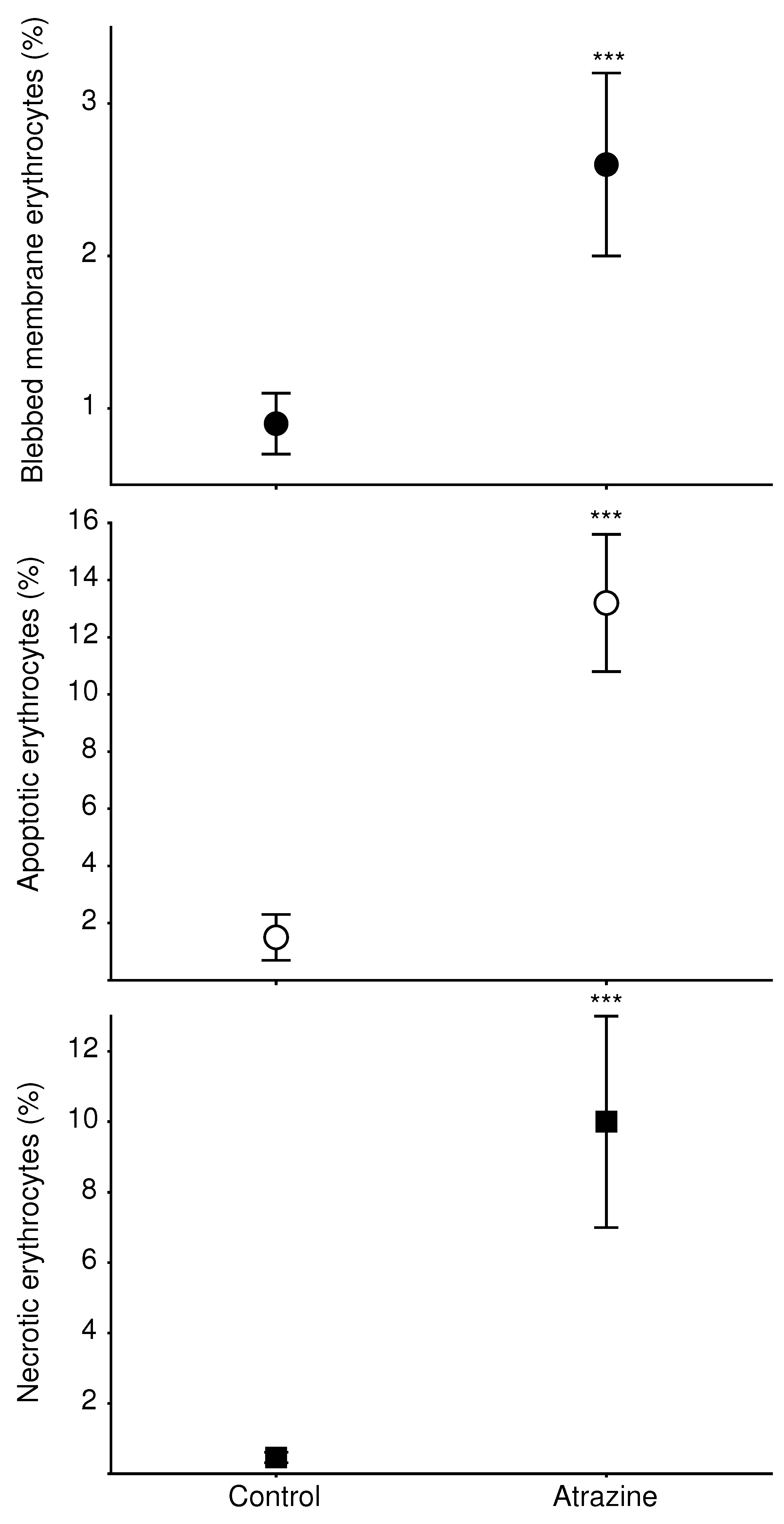
2.6. Erythrocyte Abnormalities
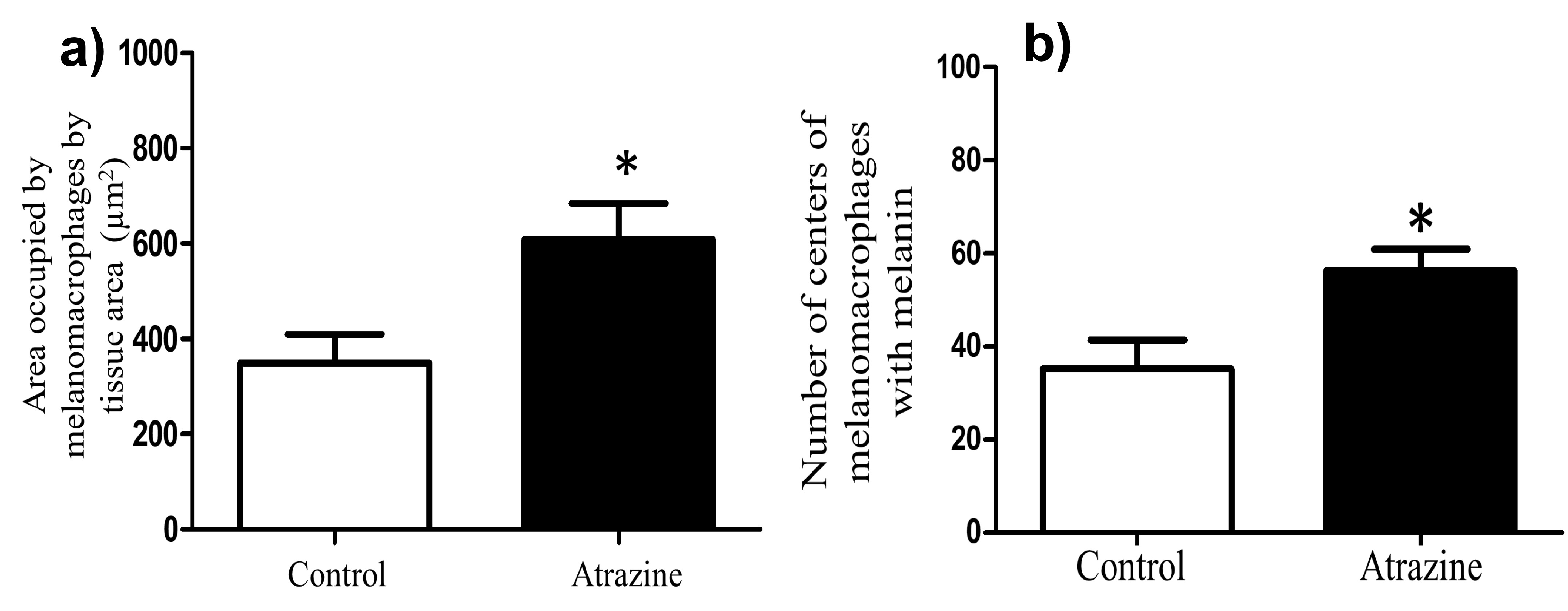
2.7. Melanomacrophages
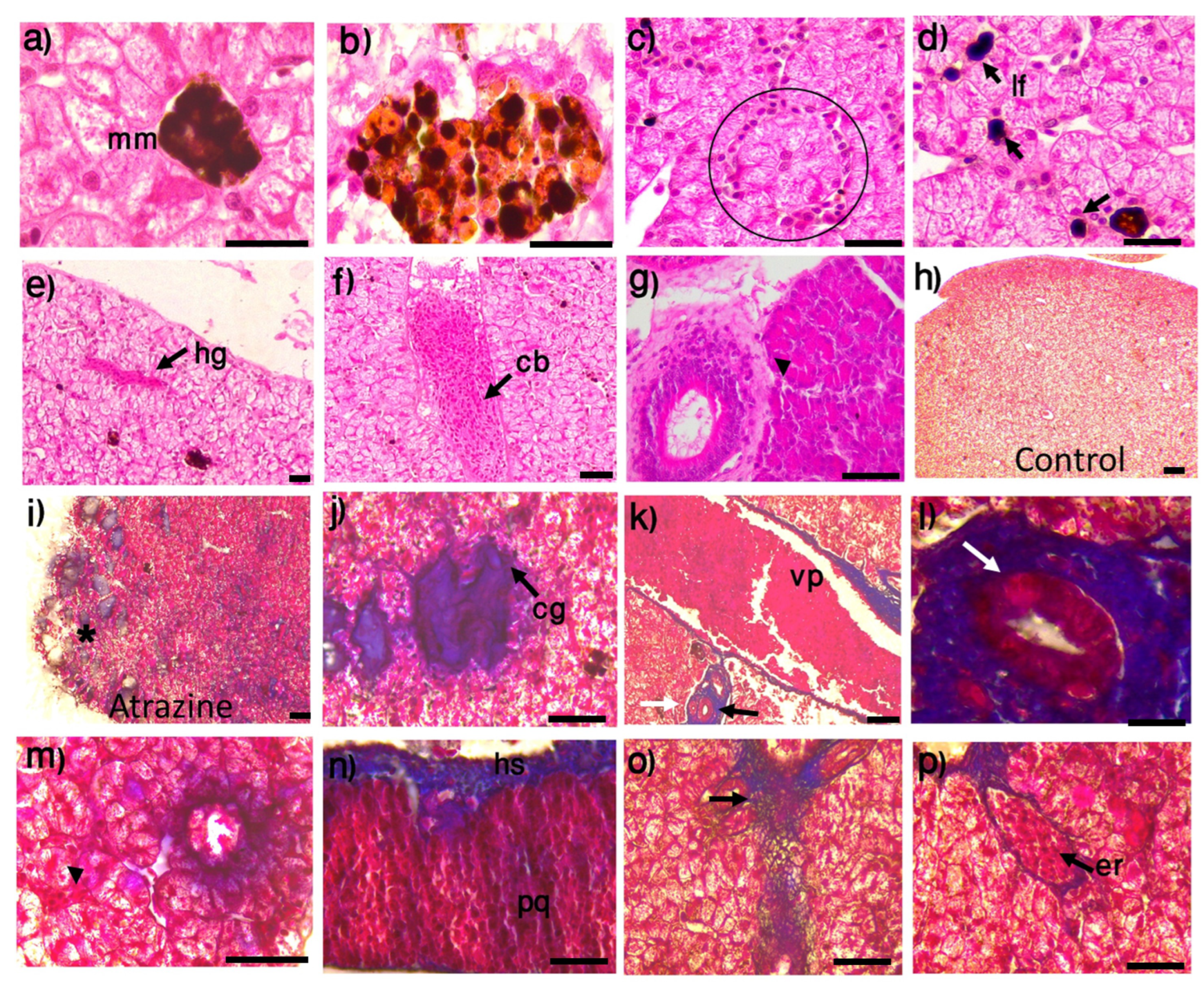
2.8. Histological Analysis of the Liver
2.9. Statistical Analysis
3. Results
3.1. Erythrocyte Morphology
3.2. Melanomacrophages and Melanine
3.3. Liver Histopathology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Egea-Serrano, A.; Relyea, R.A.; Tejedo, M.; Torralva, M. Understanding of the impact of chemicals on amphibians: A meta-analytic review. Ecol. Evol. 2012, 2, 1382–1397. [Google Scholar] [CrossRef]
- de Oliveira, J.S.P.; Gonçalves Vieira, L.; Fernandes Carvalho, W.; Benvindo de Souza, M.; de Lima Rodrigues, A.S.; Simões, K.; de Melo De Silva, D.; dos Santos Mendonça, J.; Hirano, L.Q.L.; Quagliatto Santos, A.L.; et al. Mutagenic, genotoxic and morphotoxic potential of different pesticides in the erythrocytes of Podocnemis expansa neonates. Sci. Total Environ. 2020, 737, 140304. [Google Scholar] [CrossRef]
- Cavas, T. In vivo genotoxicity evaluation of atrazine and atrazine–based herbicide on fish Carassius auratus using the micronucleus test and the comet assay. Food Chem. Toxicol. 2011, 49, 1431–1435. [Google Scholar] [CrossRef]
- Battaglin, W.A.; Rice, K.C.; Focazio, M.J.; Salmons, S.; Barry, R.X. The occurrence of glyphosate, atrazine, and other pesticides in vernal pools and adjacent streams in Washington, DC, Maryland, Iowa, and Wyoming, 2005–2006. Environ. Monit. Assess. 2009, 155, 281–307. [Google Scholar] [CrossRef]
- Černoch, I.; Fránek, M.; Diblíková, I.; Hilscherová, K.; Randák, T.; Ocelka, T.; Bláha, L. Determination of atrazine in surface waters by combination of POCIS passive sampling and ELISA detection. J. Environ. Monit. 2011, 13, 2582–2587. [Google Scholar] [CrossRef]
- Snyder, M.N.; Henderson, W.M.; Glinski, D.A.; Purucker, S.T. Biomarker analysis of American toad (Anaxyrus americanus) and grey tree frog (Hyla versicolor) tadpoles following exposure to atrazine. Aquat. Toxicol. 2017, 182, 184–193. [Google Scholar] [CrossRef]
- Sparling, D.W.; Fellers, G.M. Toxicity of two insecticides to california, USA, anurans and its relevance to declining amphibian populations. Environ. Toxicol. Chem. 2009, 28, 1696–1703. [Google Scholar] [CrossRef]
- Graymore, M.; Stagnitti, F.; Allinson, G. Impacts of atrazine in aquatic ecosystems. Environ. Int. 2001, 26, 483–495. [Google Scholar] [CrossRef]
- Lenkowski, J.R.; Reed, J.M.; Deininger, L.; McLaughlin, K.A. Perturbation of Organogenesis by the Herbicide Atrazine in the Amphibian Xenopus laevis. Environ. Health Perspect. 2008, 116, 223–230. [Google Scholar] [CrossRef]
- Clements, C.; Ralph, S.; Petras, M. Genotoxicity of select herbicides in Rana catesbeiana tadpoles using the alkaline single-cell gel DNA electrophoresis (comet) assay. Environ. Mol. Mutagen. 1997, 29, 277–288. [Google Scholar] [CrossRef]
- Gonçalves, M.W.; Marins de Campos, C.B.; Guerra Batista, V.; da Cruz, A.D.; de Marco Junior, P.; Pereira Bastos, R.; de Melo e Silva, D. Genotoxic and mutagenic effects of Atrazine Atanor 50 SC on Dendropsophus minutus Peters, 1872 (Anura: Hylidae) developmental larval stages. Chemosphere 2017, 182, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Iglesias, J.M.; Franco-Belussi, L.; Natale, G.S.; de Oliveira, C. Biomarkers at different levels of organisation after atrazine formulation (SIPTRAN 500SC®) exposure in Rhinella schineideri (Anura: Bufonidae) Neotropical tadpoles. Environ. Pollut. 2019, 244, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.; Rayburn, A. In vivo genotoxicity of atrazine to anuran larvae. Mutat. Res. Toxicol. Environ. Mutagen. 2004, 560, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.E.; Gillis, R.; Mowbray, R.C. Effect of chemical synergy and larval stage on the toxicity of atrazine and alachlor to amphibian larvae. Environ. Toxicol. Chem. 1998, 17, 519–525. [Google Scholar] [CrossRef]
- Sena, L.; Asouzu Johnson, J.; Nkomozepi, P.; Mbajiorgu, E.F. Atrazine-Induced Hepato-Renal Toxicity in Adult Male Xenopus laevis Frogs. Appl. Sci. 2021, 11, 1776. [Google Scholar] [CrossRef]
- Dornelles, M.F.; Oliveira, G.T. Effect of Atrazine, Glyphosate and Quinclorac on Biochemical Parameters, Lipid Peroxidation and Survival in Bullfrog Tadpoles (Lithobates catesbeianus). Arch. Environ. Contam. Toxicol. 2014, 66, 415–429. [Google Scholar] [CrossRef]
- Fenoglio, C.; Boncompagni, E.; Fasola, M.; Gandini, C.; Comizzoli, S.; Milanesi, G.; Barni, S. Effects of environmental pollution on the liver parenchymal cells and Kupffer-melanomacrophagic cells of the frog Rana esculenta. Ecotoxicol. Environ. Saf. 2005, 60, 259–268. [Google Scholar] [CrossRef]
- Steinel, N.C.; Bolnick, D.I. Melanomacrophage Centers As a Histological Indicator of Immune Function in Fish and Other Poikilotherms. Front. Immunol. 2017, 8, 827. [Google Scholar] [CrossRef]
- Zaya, R.M.; Amini, Z.; Whitaker, A.S.; Kohler, S.L.; Ide, C.F. Atrazine exposure affects growth, body condition and liver health in Xenopus laevis tadpoles. Aquat. Toxicol. 2011, 104, 243–253. [Google Scholar] [CrossRef]
- Mahmoud, F.A.b.R.; Gadel-Rab, A.G.; Said, R.E.S.; Saber, S.A.L.; ElSalkh, B.A.A.; Said, A.S.; Atia, M.M. Impact of atrazine and nitrate on liver and kidney of egyptian toad Sclerophrys regularis: Bioindicator alarming on ecosystem. Acta Sci. Biol. Sci. 2022, 44, e56386. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Lithobates Spectabilis. The IUCN Red List of Threatened Species 2020: e.T58722A53971736. 2020. Available online: https://www.iucnredlist.org/species/58722/53971736 (accessed on 19 September 2022).
- Hernández-Antonio, A.; Hansen, A.M. Uso de plaguicidas en dos zonas agrícolas de México y evaluación de la contaminación de agua y sedimentos. Rev. Int. Contam. Ambient 2011, 27, 115–127. [Google Scholar]
- Villada Canela, M. Estimation of Contaminating Risk Groundwater Tables by Infiltration of Atrazine Herbicide in Irrigation Districts in Mexico. Master’s Thesis, Universidad Nacional Autónoma de México, Mexico City, Mexico, 2006. (In Spanish). [Google Scholar]
- Hillis, D.M.; Frost, J.S. Three new species of leopard frogs (Rana pipiens complex) from the Mexican Plateau. In Occasional Papers of the Museum of Natural History; University of Kansas: Lawrence, KS, USA, 1985; Volume 117, pp. 1–14. [Google Scholar]
- Fenech, M. The in vitro micronucleus technique. Mutat. Res. Mol. Mech. Mutagen. 2000, 455, 81–95. [Google Scholar] [CrossRef]
- Alimba, C.G.; Aladeyelu, A.M.; Nwabisi, I.A.; Bakare, A.A. Micronucleus cytome assay in the differential assessment of cytotoxicity and genotoxicity of cadmium and lead in Amietophrynus regularis. EXCLI J. 2018, 17, 89–101. [Google Scholar] [CrossRef]
- Manrique, W.G.; da Silva Claudiano, G.; Petrillo, T.R.; de Castro, M.P.; Pereira Figueiredo, M.A.; de Andrade Belo, M.A.; de Moraes, J.R.E.; de Moraes, F.R. Response of splenic melanomacrophage centers of Oreochromis niloticus (Linnaeus, 1758) to inflammatory stimuli by BCG and foreign bodies. J. Appl. Ichthyol. 2014, 30, 1001–1006. [Google Scholar] [CrossRef]
- Lefkowitch, J.H. Special stains in diagnostic liver pathology. Semin. Diagn. Pathol. 2006, 23, 190–198. [Google Scholar] [CrossRef]
- Botelho, R.G.; Monteiro, S.H.; Christofoletti, C.A.; Moura-Andrade, G.C.R.; Tornisielo, V.L. Environmentally Relevant Concentrations of Atrazine and Ametrine Induce Micronuclei Formation and Nuclear Abnormalities in Erythrocytes of Fish. Arch. Environ. Contam. Toxicol. 2015, 69, 577–585. [Google Scholar] [CrossRef]
- Bruno-Franco, M.; Mazzei, C. The red blood cell membrane: Structure and functions. Blood Transf. 2004, 2, 160–180. [Google Scholar]
- de Campos Ventura, B.; de Angelis, D.d.F.; Marin-Morales, M.A. Mutagenic and genotoxic effects of the Atrazine herbicide in Oreochromis niloticus (Perciformes, Cichlidae) detected by the micronuclei test and the comet assay. Pestic. Biochem. Physiol. 2008, 90, 42–51. [Google Scholar] [CrossRef]
- Betz, T.; Lenz, M.; Joanny, J.F.; Sykes, C. ATP-dependent mechanics of red blood cells. Proc. Natl. Acad. Sci. USA 2009, 106, 15320–15325. [Google Scholar] [CrossRef]
- Uçar, A.; Parlak, V.; Yeltekin, A.Ç.; Özgeriş, F.B.; Çağlar, O.; Türkez, H.; Alak, G.; Atamanalp, M. Assesment of hematotoxic, oxidative and genotoxic damage potentials of fipronil in rainbow trout Oncorhynchus mykiss, Walbaum. Toxicol. Mech. Methods 2021, 31, 73–80. [Google Scholar] [CrossRef]
- Weed, R.I.; Reed, C.F. Membrane alterations leading to red cell destruction. Am. J. Med. 1966, 41, 681–698. [Google Scholar] [CrossRef] [PubMed]
- Bratosin, D.; Estaquier, J.; Slomianny, C.; Tissier, J.P.; Quatannens, B.; Bulai, T.; Mitrofan, L.; Marinescu, A.; Trandaburu, I.; Ameisen, J.C.; et al. On the evolution of erythrocyte programmed cell death: Apoptosis of Rana esculenta nucleated red blood cells involves cysteine proteinase activation and mitochondrion permeabilization. Biochimie 2004, 86, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Martínez, M.A.; Wu, Q.; Ares, I.; Martínez-Larrañaga, M.R.; Anadón, A.; Yuan, Z. Fipronil insecticide toxicology: Oxidative stress and metabolism. Crit. Rev. Toxicol. 2016, 46, 876–899. [Google Scholar] [CrossRef] [PubMed]
- Officioso, A.; Manna, C.; Alzoubi, K.; Lang, F. Bromfenvinphos induced suicidal death of human erythrocytes. Pestic. Biochem. Physiol. 2016, 126, 58–63. [Google Scholar] [CrossRef]
- Mohanty, J.; Nagababu, E.; Rifkind, J. Red blood cell oxidative stress impairs oxygen delivery and induces red blood cell aging. Front. Physiol. 2014, 5, 84. [Google Scholar] [CrossRef]
- Islas-Flores, L.; Novoa-Luna, K.A.; Islas-Flores, H.; San Juan-Reyes, N.; Gómez-Oliván, L.M. Evaluation of the Toxicity of Municipal Effluents from a Locality in the State of Mexico Using Hyalella azteca as a Bioindicator. In Pollution of Water Bodies in Latin America: Impact of Contaminants on Species of Ecological Interest; Gómez-Oliván, L.M., Ed.; Springer: Cham, Switzerland, 2019; pp. 97–111. [Google Scholar] [CrossRef]
- Bayero, U. Toxicity of Atrazine (Herbicide) to Juveniles of the African Catfish, Clarias gariepinus (Bürchell, 1822). Master’s Thesis, Ahmadu Bello University, Zaria, Nigeria, 2017. [Google Scholar]
- Ratn, A.; Awasthi, Y.; Kumar, M.; Singh, S.K.; Tripathi, R.; Trivedi, S.P. Phorate induced oxidative stress, DNA damage and differential expression of p53, apaf-1 and cat genes in fish, Channa punctatus (Bloch, 1793). Chemosphere 2017, 182, 382–391. [Google Scholar] [CrossRef]
- Farag, M.R.; Alagawany, M. Erythrocytes as a biological model for screening of xenobiotics toxicity. Chem.-Biol. Interact. 2018, 279, 73–83. [Google Scholar] [CrossRef]
- Akhtar, N.; Fiaz Khan, M.; Tabassum, S.; Zahran, E. Adverse effects of atrazine on blood parameters, biochemical profile and genotoxicity of snow trout (Schizothorax plagiostomus). Saudi J. Biol. Sci. 2021, 28, 1999–2003. [Google Scholar] [CrossRef]
- Rodilla, V. Origin and evolution of binucleated cells and binucleated cells with micronuclei in cisplatin-treated CHO cultures. Mutat. Res. Toxicol. 1993, 300, 281–291. [Google Scholar] [CrossRef]
- Jindal, R.; Batoye, S. SEM Studies on Erythrocyte Alterations in Ctenopharyngodon idellus (Cuvier and Valenciennes) induced by Fenvalerate. Res. J. Anim. Vet. Fish. Sci. 2015, 3, 1–5. [Google Scholar]
- do Nascimento Monteiro, J.A.; Araújo da Cunha, L.; Helen, P.M.; Souza dos Reis, H.; da Silva Aguiar, A.C.; Lobato de Oliveira-Bahia, V.R.; Rodríguez Burbano, R.M.; Machado da Rocha, C.A. Mutagenic and histopathological effects of hexavalent chromium in tadpoles of Lithobates catesbeianus (Shaw, 1802) (Anura, Ranidae). Ecotoxicol. Environ. Saf. 2018, 163, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Zeljezic, D.; Garaj-Vrhovac, V.; Perkovic, P. Evaluation of DNA damage induced by atrazine and atrazine-based herbicide in human lymphocytes in vitro using a comet and DNA diffusion assay. Toxicol. Vitr. 2006, 20, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Edginton, A.N.; Rouleau, C. Toxicokinetics of 14C-Atrazine and Its Metabolites in Stage-66 Xenopus laevis. Environ. Sci. Technol. 2005, 39, 8083–8089. [Google Scholar] [CrossRef] [PubMed]
- Solomon, K.R.; Carr, J.A.; Du Preez, L.H.; Giesy, J.P.; Kendall, R.J.; Smith, E.E.; Van Der Kraak, G.J. Effects of Atrazine on Fish, Amphibians, and Aquatic Reptiles: A Critical Review. Crit. Rev. Toxicol. 2008, 38, 721–772. [Google Scholar] [CrossRef]
- Agius, C. On the failure to detect haemosiderin in the melano-macrophages of dogfish Scyliorhinus canicula (L.) after prolonged starvation. Experientia 1983, 39, 64–66. [Google Scholar] [CrossRef]
- Brown, C.L.; George, C. Age-dependent accumulation of macrophage aggregates in the yellow perch, Perca flavescens (Mitchill). J. Fish Dis. 1985, 8, 135–138. [Google Scholar] [CrossRef]
- Santos, A.T.; Valverde, B.S.L.; De Oliveira, C.; Franco-Belussi, L. Genotoxic and melanic alterations in Lithobates catesbeianus (anura) tadpoles exposed to fipronil insecticide. Environ. Sci. Pollut. Res. 2021, 28, 20072–20081. [Google Scholar] [CrossRef]
- Pérez-Iglesias, J.M.; Fanali, L.Z.; Franco-Belussi, L.; Natale, G.S.; De oliveira, C.; Brodeur, J.C.; Larramendy, M.L. Multiple Level Effects of Imazethapyr on Leptodactylus latinasus (Anura) Adult Frogs. Arch. Environ. Contam. Toxicol. 2021, 81, 492–506. [Google Scholar] [CrossRef]
- Mela, M.; Guiloski, I.; Doria, H.; Randi, M.; de Oliveira Ribeiro, C.; Pereira, L.; Maraschi, A.; Prodocimo, V.; Freire, C.; Silva de Assis, H. Effects of the herbicide atrazine in neotropical catfish (Rhamdia quelen). Ecotoxicol. Environ. Saf. 2013, 93, 13–21. [Google Scholar] [CrossRef]
- Barni, S.; Vaccarone, R.; Bertone, V.; Fraschini, A.; Bernini, F.; Fenoglio, C. Mechanisms of changes to the liver pigmentary component during the annual cycle (activity and hibernation) of Rana esculenta L. J. Anat. 2002, 200, 185–194. [Google Scholar] [CrossRef]
- Różanowska, M.; Sarna, T.; Land, E.J.; Truscott, T. Free radical scavenging properties of melanin: Interaction of eu- and pheo-melanin models with reducing and oxidising radicals. Free. Radic. Biol. Med. 1999, 26, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Saad, E.M.; Elassy, N.M.; Salah-Eldein, A.M. Effect of induced sublethal intoxication with neonicotinoid insecticides on Egyptian toads (Sclerophrys regularis). Environ. Sci. Pollut. Res. 2022, 29, 5762–5770. [Google Scholar] [CrossRef] [PubMed]
- Agius, C.; Roberts, R.J. Melano-macrophage centres and their role in fish pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.E.; Melo Costa, P.; Batista Nascimento, S.; Castro, W.V.; de Azambuja Ribeiro, R.I.M.; Batista Santos, H.; Thomé, R.G. Atrazine promotes immunomodulation by melanomacrophage centre alterations in spleen and vascular disorders in gills from Oreochromis niloticus. Aquat. Toxicol. 2018, 202, 57–64. [Google Scholar] [CrossRef]
- Chen, J.Y.; Song, Y.; Zhang, L.S. Immunotoxicity of atrazine in Balb/c mice. J. Environ. Sci. Heal. Part B 2013, 48, 637–645. [Google Scholar] [CrossRef]
- Pérez-Iglesias, J.M.; Franco-Belussi, L.; Moreno, L.; Tripole, S.; de Oliveira, C.; Natale, G.S. Effects of glyphosate on hepatic tissue evaluating melanomacrophages and erythrocytes responses in neotropical anuran Leptodactylus latinasus. Environ. Sci. Pollut. Res. 2016, 23, 9852–9861. [Google Scholar] [CrossRef]
- Ribeiro, H.J.; Procópio, M.S.; Matsumura Gomes, J.M.; Oliveira Vieira, F.; Castro Russo, R.; Balzuweit, K.; Chiarini-Garcia, H.; Santana Castro, A.C.; Rizzo, E.; Dias Corrêa, J. Functional dissimilarity of melanomacrophage centres in the liver and spleen from females of the teleost fish Prochilodus argenteus. Cell Tissue Res. 2011, 346, 417–425. [Google Scholar] [CrossRef]
- Santos, R.M.B.; Monteiro, S.M.V.; Cortes, R.M.V.; Pacheco, F.A.L.; Fernandes, L.F.S. Seasonal Differences in Water Pollution and Liver Histopathology of Iberian Barbel (Luciobarbus bocagei) and Douro Nase (Pseudochondrostoma duriense) in an Agricultural Watershed. Water 2022, 14, 444. [Google Scholar] [CrossRef]
- Kelly-Reay, K.; Weeks-Perkins, B.A. Determination of the macrophage chemiluminescent response in Fundulus heteroclitus as a function of pollution stress. Fish Shellfish Immunol. 1994, 4, 95–105. [Google Scholar] [CrossRef]
- Opute, P.A.; Oboh, I.P. Hepatotoxic Effects of Atrazine on Clarias gariepinus (Burchell, 1822): Biochemical and Histopathological Studies. Arch. Environ. Contam. Toxicol. 2021, 80, 414–425. [Google Scholar] [CrossRef]
- MacFarlane, M.; Merrison, W.; Dinsdale, D.; Cohen, G.M. Active Caspases and Cleaved Cytokeratins Are Sequestered into Cytoplasmic Inclusions in Trail-Induced Apoptosis. J. Cell Biol. 2000, 148, 1239–1254. [Google Scholar] [CrossRef] [PubMed]
- Akulenko, N. Changes in Liver Parenchyma of Green Frogs (Pelophylax Esculentus Complex) Under Conditions of Anthropogenic Pollution and Their Use in Monitoring of Water Bodies. Vestn. Zool. 2015, 49, 453–458. [Google Scholar] [CrossRef]
- Krumschnabel, G.; Manzl, C.; Berger, C.; Hofer, B. Oxidative stress, mitochondrial permeability transition, and cell death in Cu-exposed trout hepatocytes. Toxicol. Appl. Pharmacol. 2005, 209, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Groscurth, P. Morphological Features of Cell Death. Physiology 2004, 19, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, U.A.; Angunawela, P.; Wickramasinghe, D.D.; Ratnasooriya, W.D.; Udagama, P.V. Heavy metal–induced toxicity in the Indian green frog: Biochemical and histopathological alterations. Environ. Toxicol. Chem. 2017, 36, 2855–2867. [Google Scholar] [CrossRef]
- Mekkawy, I.A.; Mahmoud, U.M.; Sayed, A.E.D.H. Effects of 4-nonylphenol on blood cells of the African catfish Clarias gariepinus (Burchell, 1822). Tissue Cell 2011, 43, 223–229. [Google Scholar] [CrossRef]
- Ahmadivand, S.; Farahmand, H.; Mirvaghefi, A.; Eagderi, S.; Shokrpoor, S.; Rahmati-Holasoo, H. Histopathological and haematological response of male rainbow trout (Oncorhynchus mykiss) subjected to butachlor. Vet. Med. 2014, 59, 433–439. [Google Scholar] [CrossRef]
- Mohamed, F.A. Histopathological Studies on Tilapia zillii and Solea vulgaris from Lake Qarun, Egypt. World J. Fish Mar. Sci. 2009, 1, 29–39. [Google Scholar]
- Loumbourdis, N.S. Liver Histopathologic Alterations in the Frog Rana ridibunda from a Small River of Northern Greece. Arch. Environ. Contam. Toxicol. 2007, 53, 418–425. [Google Scholar] [CrossRef]
- Păunesco, A.; Ponepal, C.M.; Grigorean, V.T.; Popescu, M. Histopathological changes in the liver and kidney tissues of marsh frog (Pelophylax ridibundus) induced by the action of talstar 10EC insecticide. Analele Univ. Din Oradea -Fasc. Biol. 2012, 19, 5–10. [Google Scholar]
- Fanali, L.Z.; Freitas, J.S.; Franco-Belussi, L.; Taboga, S.R.; de Oliveira, C. Liver description in three neotropical anuran species: From anatomy to ultrastructure. Acta Zool. 2022, 103, 316–325. [Google Scholar] [CrossRef]
- Raymundo Raymundo, E.; Nikolskii Gavrilovç, I.; Duwig, C.; Prado Pano, B.L.; Moreno, C.I.; Gavi Reyes, F.; Figueroa Sandoval, B. Atrazine transport in an Andosol and a Vertisol of Mexico (In Spanish). Interciencia 2009, 34, 330–337. [Google Scholar]
- Hernández Vargas, J. Atrazine leaching in a vertisol from the Bajío de Guanajuato, Mexico. Master’s Thesis, Colegio de Posgraduados, Montecillo, Mexico, 2008. (In Spanish). [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez-Tepepa, M.; Hernández-Pérez, K.; Juárez-Santacruz, L.; Cruz-Lumbreras, S.R.; García-Nieto, E.; Anaya-Hernández, A.; Morales-Cruz, C. Cytotoxic Effects of the Atrazine Herbicide on Erythrocytes and Liver Damage in Lithobates spectabilis. Fishes 2023, 8, 207. https://doi.org/10.3390/fishes8040207
Méndez-Tepepa M, Hernández-Pérez K, Juárez-Santacruz L, Cruz-Lumbreras SR, García-Nieto E, Anaya-Hernández A, Morales-Cruz C. Cytotoxic Effects of the Atrazine Herbicide on Erythrocytes and Liver Damage in Lithobates spectabilis. Fishes. 2023; 8(4):207. https://doi.org/10.3390/fishes8040207
Chicago/Turabian StyleMéndez-Tepepa, Maribel, Karla Hernández-Pérez, Libertad Juárez-Santacruz, Senobia Rosalia Cruz-Lumbreras, Edelmira García-Nieto, Arely Anaya-Hernández, and Cuauhtémoc Morales-Cruz. 2023. "Cytotoxic Effects of the Atrazine Herbicide on Erythrocytes and Liver Damage in Lithobates spectabilis" Fishes 8, no. 4: 207. https://doi.org/10.3390/fishes8040207
APA StyleMéndez-Tepepa, M., Hernández-Pérez, K., Juárez-Santacruz, L., Cruz-Lumbreras, S. R., García-Nieto, E., Anaya-Hernández, A., & Morales-Cruz, C. (2023). Cytotoxic Effects of the Atrazine Herbicide on Erythrocytes and Liver Damage in Lithobates spectabilis. Fishes, 8(4), 207. https://doi.org/10.3390/fishes8040207