Abstract
A total of 1177 Sthenoteuthis oualaniensis were randomly collected from the northwest Indian Ocean from February between May 2019 and 2020 by lighting falling-net vessels. The age, growth, and population structure of S. oualaniensis were studied based on the statolith microstructure. The results showed that the range of mantle length (ML) was 123–562 mm for females and 88–273 mm for males, and the range of body weight (BW) was 78–6268 g for females and 82–518 g for males in 2019 and 2020, respectively. The hatching date extended from May to December, with the 2019 samples mainly composed of the autumn population, while the 2020 samples were mostly composed of the summer population. The analysis of covariance (ANCOVA) showed that there were significant differences in the growth of ML–age and BW–age between sexes. In the growth model of the ML–age relationship, both females and males in 2019 were best described by s linear model, and 2020 was best described by a logarithmic model. The growth model of the BW–age relationship of females and males in 2019 was best described as linear and the growth model of the BW–age relationship of females and males in 2020 was described logarithmically and exponentially, respectively. The average absolute daily growth rate (AGR) and instantaneous growth rate (IGR) for ML of the 2019 samples were 0.85 mm/d and 0.40%/d, and the average AGR and IGR for ML of the 2020 samples were 0.65 mm/d and 0.18%, respectively. The growth of S. oualaniensis samples in 2019 was faster than that in 2020. This study provided basic information on the age, growth, and population of S. oualaniensis, which will supply a scientific basis for stock assessment and sustainable development.
1. Introduction
The Purpleback Flying Squid Sthenoteuthis oualaniensis is classified under Cephalopoda, Ommastrephidae, and Sthenoteuthis, which are widely distributed in equatorial and subtropical regions of the Indian and Pacific Oceans [1]. The Arabian Sea is located northwest of the Indian Ocean. The tropical Indian Ocean circulation dynamics and monsoon dynamics [2,3], provides an ideal temperature, salty, and nutrient-rich growth environment for S. oualaniensis [3,4] supporting a productive fishery. S. oualaniensis has a strong reproductive ability and its fisheries resources are abundant. Ommastrephidae are very common, but the northwest Indian Ocean has one of the high-densities [5,6]. With the development of global fisheries, cephalopods are targeted by various countries such as China, Indonesia, and the Philippines [7,8,9], and provide abundant high-quality protein [10]. The estimated total instantaneous biomass of S. oualaniensis in the world is 8 to 11.2 million tons [10]. The biomass of S. oualaniensis in the Indian Ocean is 3–4 million tons, and the resource density is the highest in the Arabian Sea [11]. Therefore, S. oualaniensis has high development and utilization value.
At present, several studies have described the life history of S. oualaniensis, including population structure, age and growth, diet and behavior, reproduction, and ecology. Due to the complex population structure and wide geographical distribution, different intraspecific forms were divided into groups by body size [12,13,14,15]. Presently, there is no clear boundary between the distribution of different populations of S. oualaniensis in the Indian Ocean. Based on the morphological analysis of squid in the northwest Indian Ocean, researchers found the region north of 18° N was dominated by large populations, the region from 12–18° N was dominated by medium populations, and the region south of 12° N was dominated by small populations [15,16]. S. oualaniensis are typically opportunistic species that prey on small fish and crustaceans, and even exhibit fairly pronounced cannibalistic behavior [14,17]. In addition, S. oualaniensis plays an important role in the oceanic food web, which is commonly found in the stomachs of various carnivores, such as seabirds [18], large fish [19], and marine mammals [20].
The statoliths of cephalopods are located in the balance capsule of the posterior abdomen of the head. They are calcified hard tissues that appear in pairs in the cephalopods. They not only play the role of balancing the mantle, but are also an important part of the acceleration sensor of cephalopods [21]. The statoliths are characterized by a stable structure, corrosion resistance, and irreversible deposition [22]. The growth rings deposited in the statoliths store a large amount of life information [23,24]. Population genetic information exchanges and major environmental events can lead to changes in the morphology of the statolith microstructures [25,26]. Therefore, the statoliths are often considered an ideal body part for studying life history [27,28]. In addition, the growth rate of S. oualaniensis is fast and the population structure is complex. The reproductive strategy of the S. oualaniensis is to lay eggs many times. After laying eggs once, the squid will continue to grow and lay eggs again when they reach sexual maturity [29]. There are different spawning groups, including spring, summer, and autumn [12,30]. The microstructure of the statoliths can be used for studying age growth, population division, life history, and migration path [25,31].
In recent years, based on the statoliths and beaks, many studies have been conducted on small squid populations distributed in the South China Sea [26,32,33]; however, there are few studies on the S. oualaniensis population in the northwest Indian Ocean, and there are no comprehensive articles on the S. oualaniensis population based on the statolith information. In this study, we studied the age, growth, and population structure of S. oualaniensis in the northwest Indian Ocean, using the statolith microstructure of S. oualaniensis samples collected from the northwest Indian Ocean over two consecutive years, to provide fishery biology and resource management information of S. oualaniensis in the northwest Indian Ocean.
2. Materials and Methods
2.1. Sampling
A total of 1177 S. oualaniensis samples were randomly collected from Chinese lighting falling-net vessels “Xinhai 1223” in the northwest Indian Ocean (13°05′–19°45′ N, 61°06–67°50′ E) from February through May 2019, and from February to May 2020 (Figure 1). About 15–30 samples were collected randomly based on the different investigation locations and times. Among them, 514 S. oualaniensis were collected from 20 sampling localities in 2019 and 663 S. oualaniensis were collected from 36 sampling localities in 2020. All squid samples were immediately frozen and later dissected in the laboratory.
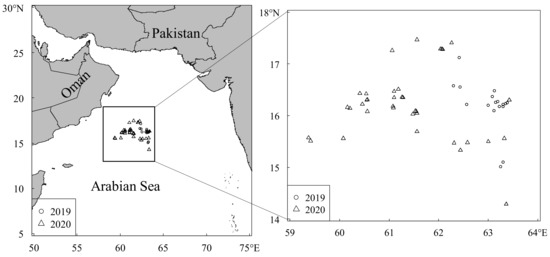
Figure 1.
Sample localities of S. oualaniensis in the northwest Indian Ocean.
2.2. Basic Biological Determination and Statolith Extraction
The squid were measured after thawing the samples; the mantle length (ML) and body weight (BW) were measured to accuracies of 1 mm and 1 g, respectively. The sex and maturity stages were identified through visual examinations, and five age stages were identified (immature: I and II; maturing: III; mature: IV, and V) [15,34]. There were 876 pairs of complete statolith samples from S. oualaniensis extracted, washed, and stored in a centrifugal tube with 75% alcohol for age determination [35].
According to the illuminator and morphological characteristics of the species, the squid can be divided into large, medium, and small populations [36]. A total of 588 age data were obtained, including 290 in 2019 (female: 154, male: 136) and 298 in 2020 (female: 168, male: 130).
2.3. Statolith Processing and Aging
Standard statolith aging methods were used under the assumption rings and were formed daily [32,37]. We chose the statoliths, as this region exhibits many dark, periodic longitudinal bands that can be easily used to interpret the age of the squid. After cleaning with water, the statoliths were placed flat in a plastic mold and hardened in a plastic mold with a gradually caking epoxy [32,37]. The focal plane sample was ground on a sander with coarse sandpapers of different (grit: 120, 600, 1200, and 2000) and then polished with an alumina solution on smooth sandpaper (2400) to remove scratches until the age increments were clearly visible.
Different parts of the statoliths were observed using an Olympus microscope and photographed by a charge-coupled device (CCD) at different magnifications (10×, 20×, 40×, 100×, 200×, 400×). Then, an intact picture was consolidated using Photoshop CS 24.0 for the subsequent age estimation. The age information of each sample was extracted by two observers and each was read three times. To decrease the bias in age estimation, measurements were considered at the effective age when differences in the mean age determined by two observers were less than 10% [37,38], and the mean of six readings were taken as the true age of the squid. The age and growth patterns of cephalopods can be reliably assessed by comparing the statolith increments observed under artificial feeding conditions [39,40]. Based on the statolith daily increments, we chose to back-calculate the hatching date and explore the growth patterns of the S. oualaniensis specimens in the northwest Indian Ocean.
2.4. Population Structure Identification
Previous studies have shown that the age and growth patterns of Ommastrephidae squid can be reliably determined by the daily deposition of the statolith increments [37]; thus, the hatching date could be back-calculated from the capture date and age information to infer the hatching group.
2.5. Data Analysis
Based on the biological data, the following models were used to quantify the relationships between ML and BW and between ML and age. Analysis of covariance (ANCOVA) was used to evaluate whether differences existed in the ML and BW growth patterns between females and males.
Linear function [41]:
L = a x + b
Exponential function [42]:
L = a e b x
Power function [43]:
L = a x b
Logarithmic function [44]:
where L is the ML (in mm)/BW (in g) at age x; x is the squid age (in days), and a and b are the parameters to be estimated.
L = a ln(x) + b
For the above four models, the likelihood and the Akaike information criterion (AIC) of each model were calculated using the following equation [32,45,46]:
where L(data/θ) is the likelihood of observing the data given parameters θ; vector θ denotes parameters to be estimated; y and f(t) are the observed values for the featured parameters and the estimated values from the model, respectively; n is the number of observations; m is the number of model parameters; and Lmax is the maximum value of L(data/θ) [45]. The model which yields the smallest AIC value was selected as the best model [32,35]. The absolute daily growth rates (AGRs, in mm/d or g/d) and instantaneous growth rates (IGRs, in %/d) of the ML and BW were estimated at each 20-day interval. The IGRs and AGRs were calculated using the following functions [46]:
where R1 and R2 were the estimated ML or BW values at the start (t1) and end (t2) of the time interval, respectively.
AIC = −2 ln (Lmax) + 2 m
AGR = (R2 − R1)/(t2 − t1)
IGR = ln(R2) − ln(R1)/(t2 − t1) × 100%
All statistical analyses were conducted in RStudio 4.0.3 and Excel 2019, and the pictures were analyzed in Photoshop 2020.3.
3. Results
3.1. Individual Composition and Shape Features
The mantle of the Indian Ocean S. oualaniensis was conical and the ML was about four times the mantle width. The body surface had the illuminator apparatus of different sizes, and the central part of the back of the mantle was purplish brown. The fin length was about 1/3 of the ML and the two fins were rhomboid in shape (Figure 2).

Figure 2.
Appearance of S. oualaniensis.
The ANCOVA showed that in 2019 (F = 14.54, p = 0.00 < 0.05) and 2020 (F = 2562.29, p = 0.00 < 0.05), significant differences were found in the ML between the sexes; thus, the ML was divided by sex. Based on the mantle length, the interval of 50 mm can be used to describe the class of the ML suitably.
In 2019, the female MLs ranged from 123–356 mm (mean ML: 218.94 mm), with the majority (83.32%) of samples ranging from 121–220 mm. The dominant group ranged from 171–220 mm, accounting for 42.61% (Figure 3a). The male MLs ranged from 98–273 mm (mean ML: 194.83 mm), with the majority (91.89%) of samples ranging from 121–220 mm. The dominant ML group ranged from 471–520 mm, accounting for 35.36% (Figure 3b). In 2020, the female MLs ranged from 279–562 mm (mean ML: 448.36 mm), with the majority (79.74%) of samples ranging from 371–520 mm. The dominant group ranged from 471–520 mm, accounting for 35.36% (Figure 3a). The male MLs ranged from 88–272 mm (mean ML: 188.96 mm), with the majority (86.94%) of samples ranging from 121–270 mm. The dominant ML group ranged from 171–220 mm, accounting for 35.36% (Figure 3b).
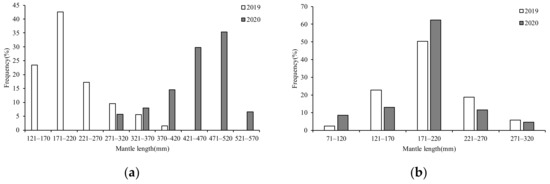
Figure 3.
Composition of ML for S. oualaniensis in different years: (a) female; (b) male.
The ANCOVA showed that in 2019 (F = 23.26, p = 0.00 < 0.05) and 2020 (F = 834.02, p = 0.00 < 0.05), there were significant BW differences between the sexes; thus, the BW composition in S. oualaniensis was divided by sex. Considering the different distributions of BW in 2019 and 2020, we chose a different but appropriate BW interval for each year (200 g for females in 2019, 1000 g for females in 2020, and 60 g for males in 2019 and 2020).
In 2019, the female BWs ranged from 78–1246 g (mean BW: 386.17 g), with the majority (77.51%) of samples ranging from 71–670 g. The dominant group ranged from 71–270 g, accounting for 40.40% (Figure 4a). The male BWs ranged from 82–518 g (mean BW: 229.14 g), with the majority (72.07%) of samples ranging from 101–280 mm. The dominant BW group ranged from 161–220 mm, accounting for 30.96% (Figure 4b). In 2020, the female BWs ranged from 673–6268 g (mean BW: 3375.26 g), with the majority (68.66%) of samples ranging from 2271–5270 g. The dominant group ranged from 3271–4270 g, accounting for 27.54% (Figure 4b). The male BWs ranged from 105–437 g (mean BW: 192.72 g), with the majority (85.94%) of samples ranging from 101–280 g. The dominant BW group ranged from 161–220 g, accounting for 39.63% (Figure 4c).
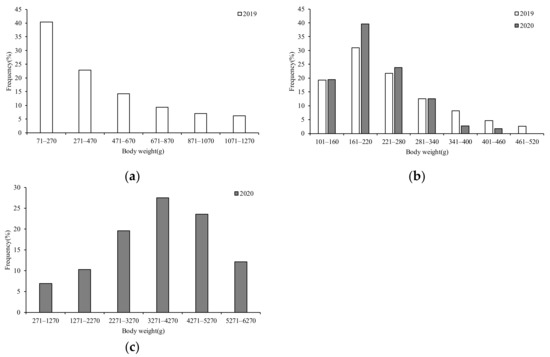
Figure 4.
Composition of BW for S. oualaniensis in different years: (a,c) female; (b) male.
3.2. Statolith Microstructure
The growth rings were composed of alternating light and dark rings. According to the changes in the brightness and width of growth rings, the statoliths were divided into three main growth zones found in the dorsal dome from the nucleus (N, Figure 5a) toward the edge of the dorsal dome: the postnuclear zone (P, Figure 5b), dark zone (DZ, Figure 5b,c), and peripheral zone (PZ, Figure 5c) for all individuals examined. The N of the statolith is the starting point of growth, and each wheel grain is centered on it and grows in a ring outward. The N is dark and almost no grains can be seen. The P area is bright and the ring is uniform. The distance between the two rings is moderate. Because it is close to the statolith nucleus, the ring in the P area is nearly round. The DZ is located outside the P area, where the brightness of the wheel stripes is the darkest and the distance between the two wheel stripes is the widest, which is most conducive to the counting and reading of the growing stripes (Figure 5b). Outward from the DZ is the PZ, where the light is brightest and the distance between the rings is narrowest, and the shape of each ring is similar to a drop shape with a sharper end and a flatter section (Figure 5c). In general, from the N of the statolith to PZ, the brightness of the statolith sections changed from bright to dark and then to the brightest, and the width of the wheel grain changed from narrow to wide and then to the narrowest.
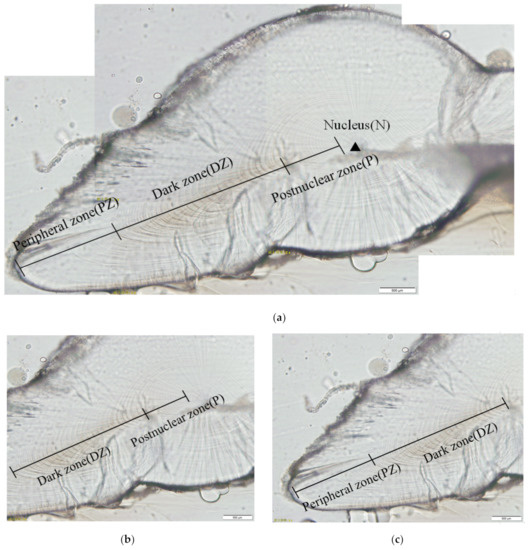
Figure 5.
Statolith microstructure of S. oualaniensis: (a) showing three distinct growth zones with different incremental width; (b) showing the postnuclear zone (P) and dark zone (DZ); (c) showing the dark zone (DZ) and peripheral zone (PZ).
3.3. Age Structure and Maturity
The age composition of the two years of S. oualaniensis was statistically different between the sexes and years. According to the age range of all samples, the 30-d interval described the age classification better. In 2019, the age of females ranged from 113–263 days, with the majority (82.31%) of samples being 141–230 days. The dominant age group ranged from 171–200 days, accounting for 31.13% (Figure 6a). The age range of males ranged from 136–253 days, with the majority (68.59%) of samples being 171–230 days. The dominant age group ranged from 171–200 days, accounting for 42.82% (Figure 6b). In 2020, the age of females ranged from 148–267 days, with the majority (82.31%) of samples being 201–260 days. The dominant age group ranged from 231–260 days, accounting for 36.86% (Figure 6a). The age range of males ranged from 133–229 days, with the majority (68.20%) of samples being 171–230 days. The dominant age group ranged from 171–200 days, accounting for 38.86% (Figure 6b).
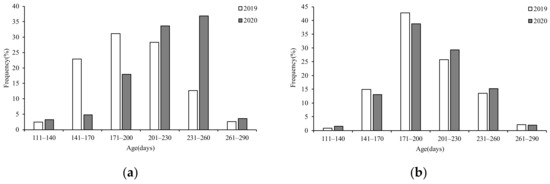
Figure 6.
Composition of age for S. oualaniensis in different years: (a) female (b) male.
Maturity status differed between sexes were observed in two years of S. oualaniensis samples (Figure 7). In 2019, the majority of females (87.46%) were in stages I and II, while the majority of males (90.72%) were in stages II–IV (Figure 7a,b). In 2020, the majority (77.25%) of female samples were in stages III and IV (Figure 6a), and the majority (71.93%) of male samples were in stages III and IV (Figure 7a,b).
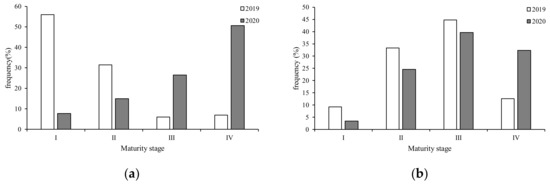
Figure 7.
Maturity stage compositions of the two S. oualaniensis sexes in the northwest Indian Ocean in the two years of study: (a) female; (b) male.
3.4. Hatching Date and Group
The hatching dates were estimated by combining the age data and sample date. In 2019, the hatching date of females was from May to December 2018, and the peak hatching time was from August to November (Figure 8a). The hatching date of males was from May to December 2018, and the hatching peak was from August to November (Figure 8b). In 2020, the hatching dates of females and males were distributed from May to December 2019, and the hatching peaks were from July to August and June to August, respectively. In general, the hatching date in 2019 was mostly distributed from August to November 2018. The hatching date in 2020 was mostly distributed from June to August 2019. The 2019 samples were mainly composed of the autumn population, while the 2020 samples were mostly composed of the summer population.
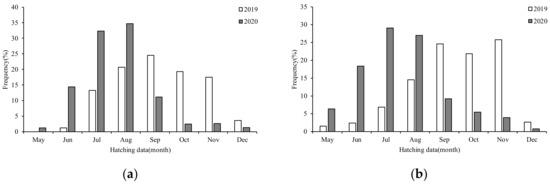
Figure 8.
Hatching date frequency distribution of S. oualaniensis of different sexes in different years: (a) female; (b) male.
3.5. Growth Patterns
The ANCOVA showed there were significant differences in the ML–age between the sexes of the S. oualaniensis in 2019 (F = 51.94, p = 0.00 < 0.05), and the same result was found in the S. oualaniensis samples in 2020 (F = 4251.99, p = 0.00 < 0.05). Based on the lowest derived AIC value (Table 1), the ML–age relationship of females and males for the autumn population (2019) was best described by a linear growth model (Figure 9a,b), while the ML–age relationship of females and males for the summer population (2020) was best described by a logarithmic growth model (Figure 9a,b).

Table 1.
Parameters of the linear, power, exponential, and logarithmic growth models fitted to the ML–age and BW–age relationships of the S. oualaniensis samples.
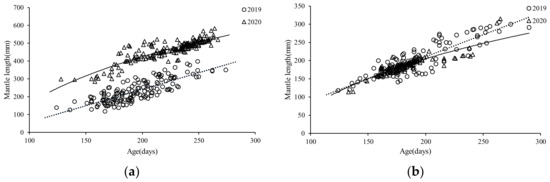
Figure 9.
The relationship between ML–age for S. oualaniensis in the northwest Indian Ocean in different years: (a) female; (b) male.
The ANCOVA showed there were significant differences in the BW–age between the sexes in 2019 (F = 47.51, p = 0.00 < 0.05), and the same result was found in the S. oualaniensis samples in 2020 (F = 870.30, p = 0.00 < 0.05). Based on the lowest derived AIC value (Table 1), the BW–age relationship of females and males for the autumn population was best described by a linear growth model (Figure 10a,b), and the BW–age relationship of females and males for the summer population was best described by a logarithmic and an exponential growth model, respectively (Figure 10a,b).
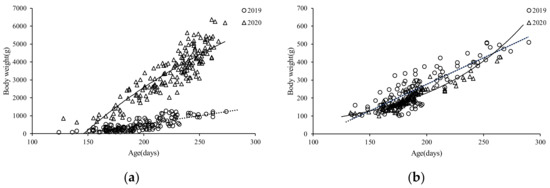
Figure 10.
The relationship between BW–age for S. oualaniensis in the northwest Indian Ocean in different years: (a) female; (b) male.
ML–age growth model for the autumn population(females):
ML = 1.8537 Age − 129.01 (R2 = 0.6607, n = 154)
ML–age growth model for the autumn population(males):
ML = 1.2067 Age − 30.866 (R2 = 0.7749, n = 136)
BW–age growth model for the autumn population(females):
BW= 9.4971 Age − 1379.1 (R2 = 0.6407, n = 154)
BW–age growth model for the autumn population(males):
BW = 2.9509 Age − 313.81 (R2 = 0.7715, n = 136)
ML–age growth model for the summer population(females):
ML = 373.33 ln (Age) − 1554.3 (R2 = 0.7617, n = 168)
ML–age growth model for the summer population(males):
ML = 189.52 ln (Age) − 799 (R2 = 0.7469, n = 130)
BW–age growth model for the summer population(females):
BW = 8246.80 ln (Age) − 41122 (R2 = 0.7919, n = 168)
BW–age growth model for the summer population(males):
BW = 22.8370 e 0.0115Age (R2 = 0.8147, n = 130)
3.6. Growth Rates
The results showed that both the summer and autumn populations grew rapidly. The maximum AGR (Figure 11a) and IGR (Figure 11b) values for ML of the autumn population (2019) were 1. 73 mm/d and 0.18%/d, respectively, within 191–210 days of age. The maximum AGR (Figure 11a) and IGR (Figure 11b) values for ML of the summer population (2020) were 1.09 mm/d and 0.24%/d, respectively, within 191–210 days of age. The average AGR and IGR for ML of the 2019 samples were 0.85 mm/d and 0.40%/d, respectively, and the average AGR and IGR of the 2020 samples were 0.65 mm/d and 0.18%/d, respectively. The growth of S. oualaniensis samples in 2019 was faster than that in 2020.
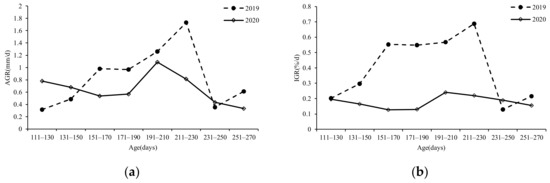
Figure 11.
Relationship between ML growth rate and age of S. oualaniensis: (a) AGR; (b) IGR.
4. Discussion
4.1. ML and BW Distributions
Our results showed females were generally larger than males, and the complex differences among different body sizes were the typical characteristics. The body size of the squid at maturity and the presence or absence of photophores on the back are important characteristics of its population division [12,47]. Yatsu [30] divided S. oualaniensis into three body size groups: large, medium, and small populations, according to differences in ML and age. Studies have shown that large populations of S. oualaniensis are found in the Red Sea, Gulf of Aden, and Arabian waters in the northwest Indian Ocean [12,15,48]. The medium group of S. oualaniensis can be found throughout the species range, which has a wide distribution [10]. Small groups are found mainly in the equatorial waters of the Indian and Pacific Oceans (15° N–15° S) and are characterized by their small body size and early maturity [16,49]. According to our study, the age of S. oualaniensis samples in 2019 and 2020 was approximately 200 days. The body size groups of the S. oualaniensis samples collected this time mainly included the large and medium groups. Compared with the jigging vessel, the light falling-net was less selective on individual size and more random [50,51]. Therefore, it represented the population structure and composition of S. oualaniensis in the northwest Indian Ocean much better.
4.2. Statolith Microstructure
Arkhipkin [52] found the statoliths of all squid in the family Ommastrephidae had similar growth patterns. The statoliths were characterized by one ring a day and the life cycle of S. oualaniensis was approximately one year [26,53]. Compared with fish, cephalopods have much smaller statoliths, and the size of the statoliths of the S. oualaniensis is only about 1 mm [50]. In this study, the N of the statolith is the starting point of growth, and each wheel grain is centered on it and grows in a ring outward. Clear growth increments comprised of light and dark bands were distributed on N to PZ. These growth increments exhibited different widths and patterns in each zone. When the statolith growth rings were counted, it was found that the growth rings in P were not clear and not spaced evenly, which made the rings difficult to count. However, the growth rings in DZ were clear and uniform, which were easy to count, while the statolith growth rings spaced in PZ were narrow and difficult to count. The growth of the statolith microstructure of S. oualaniensis starts from the N, so the growth rings are counted uniformly from N to PZ [54].
Studies have shown the growth of squid statolith is closely related to the development of the individual [31]. The P of the statolith is formed in the larval stage of squid. When the larval squid are in the endogenous nutrient stage and grow slowly, the rings of the statolith are relatively dense. The DZ is formed in the juvenile stage. At this time, the squid has the ability to prey, its growth rate is faster, and the DZ in the statolith microstructure shows a wider growth ring [55]. The PZ is formed at the subadult and adult stages [31]. The growth of the squid is also closely related to gonadal development. Nutrients in the juvenile stage are mainly used for individual growth [56]. After growth to a certain stage, nutrients obtained are mainly used for gonadal development, and the growth of the body size slows down [57]. Therefore, the growth rings in the PZ of the statolith microstructure are dense. In addition, the arrangement of the statolith ring patterns may also be related to the food and habitat environment [58]. The transition between DZ and PZ in the statolith microstructure may be related to individual feeding habits. In the juvenile stage, the individual squid is just formed and mainly feeds on crustaceans [10,15]. When they develop to the subadult and adult stages, they become strong swimmers, and their diet changes to fish and small cephalopods [59]. In addition, it has been suggested that the transition between the DZ and PZ of the statolith microstructure may be related to the vertical movement of the population.
4.3. Age Structure
The age composition of male and female samples of two years was consistent with the normal distribution. The life span of the S. oualaniensis varies in different seas, and the results of this study were similar to those of previous studies (less than 10 months) in the northwest Indian Ocean [60] and the South China Sea [13]. However, the life span of S. oualaniensis was different (less than 6 months) from those in equatorial waters [61], the North Pacific Ocean [62], and the Bay of Bengal [63]. The differences in the age of S. oualaniensis are not only related to various biological factors, such as population, sex, and prey [31,58], but also may be related to the marine environment, such as temperature, salinity, and chlorophyll-a concentration [64]. During major marine environmental events, SST increases or decreases [27] and has an effect on the growth and development of S. oualaniensis. In addition, the age structure varies according to geographical region, sex, and population [25]. Some studies have shown that female S. oualaniensis live longer than males [60], and this phenomenon was also found in this study. In this study, no S. oualaniensis samples were found in stage V of sexual maturity. Most of the female samples in 2019 were in the immature stage The male samples were in the preparatory stage, maturing stage, and mature stage, while most of the squid samples in 2020 were in the maturing stage and mature stage. The results of the 2019 study are consistent with previous studies in the northwest Indian Ocean [15]. In addition, the results further indicate that the males of this species mature faster than females [10].
4.4. Hatching Group
The growth rings of the statolith were deposited after hatching, so the hatching season can be backward calculated according to the count of catching date and age. The distribution of the spawning peak was different in different seasons, which may be due to the asynchronous maturation and continuous spawning of S. oualaniensis. The spawning population of S. oualaniensis was divided into spring, summer, and autumn [48]. The results of this study showed that the hatching date of S. oualaniensis in both 2019 and 2020 was from May to December, and the peak hatching date of the 2019 samples was from August to November 2018, which was mainly the autumn population. The hatching date peak of the 2020 samples was from June to August 2019, which was mainly the summer population. The spawning of S. oualaniensis in the northwest Indian Ocean is almost year-round, but the peak hatching date is different [48]. Previous studies on S. oualaniensis in the northwest Indian Ocean found spring and autumn populations [60], which are partially identical to the results of this study. In addition, the spring population of S. oualaniensis are found in the Arabian Sea [45]. In the northwest Indian Ocean, the spawning groups of S. oualaniensis are not distributed independently, but superimposed on each other, and the population structure is complex, which may be related to the cluster living in warmer waters [16]. In this study, the two years of hatching peak were concentrated from August to November and from June to August, and the spawning cycle was long, which further proved that there were multiple spawning groups, and the reproduction was characterized by multiple batches and seasonal spawning [15,65].
4.5. Growth Patterns
The results of this study showed that there were significant differences in the growth relationship between sexes in both ML–age and BW–age of two years of S. oualaniensis samples. Among them, the growth models of ML–age and BW–age of the autumn population were best described by a linear function. The growth models of ML–age of the summer population females and males S. oualaniensis samples were best described by a logarithmic function, while the growth models of females and males BW–age were best described by logarithmic and exponential functions, respectively. This may be related to the SST, habitat environment, and feeding level of the growing area [12,14,43]. Linear, power, exponential, and logarithmic functions were used to fit the growth relationship of ML–age and BW–age of different populations in different sea areas [13,66,67]. For example, the growth models of ML–age and BW–age for the medium-sized group of S. oualaniensis in the South China Sea are best described by a logarithmic and a linear model, respectively [66], which are partially similar to those in this study. The growth model of S. oualaniensis ML–age in the equatorial of the Pacific Ocean is described by a linear model [60], similar to the relationships in the spring population of Dosidicus gigas [35,68], winter population of Illex argentinus [54] and O. bartramii [37], which were the same as the results of the autumn population. Studies have shown that the growth model of BW–age of S. oualaniensis in the South China Sea is best described by an exponential model [13], similar to the relationships in the spring–autumn of D. gigas [68], which were partly the same as the results of this study.
The growth of squid is affected by many factors, especially the ML–age and BW–age relationships, which are obviously affected by the change in environmental factors, geographical distribution, and complex population structures [43,44,69]. The growth models of ML–age and BW–age for 2019 and 2020 S. oualaniensis samples were different, which may be largely related to the marine environment in different years. Squid is very sensitive to regional environmental conditions, especially SST. When SST is higher or lower than the appropriate temperature, the growth and development of squid may be directly affected [70,71]. In addition, in the year of the La Niña event, the SST in the South China Sea increased abnormally, the individual size of S. oualaniensis was smaller than that in the normal year, and the ML of initial sexual maturity decreased [72]. Previous studies have shown that most of the squid in the Indian Ocean area south of 16° N were in the immature stage, and most of the squid in the area north of 16° N were in the mature stage [15]. The maturation of females has a clear spatial variation along the latitude, which indirectly indicates the effect of sea surface temperature on the growth and development of squid. The early life history of the S. oualaniensis samples collected in 2020 was affected by a strong positive dipole phenomenon, and the SST exceeded the suitable range for S. oualaniensis growth during this period. However, the samples collected in 2019 lived at a normal SST, which may be one of the important reasons for the existence of growth models in different years. There are also differences in the growth models of S. oualaniensis with different body size groups and spawning groups. Therefore, the best growth model of S. oualaniensis needs to be determined by comparing the fitting effects of various models. Due to the limitation of the number of samples, this study studied and analyzed part stages of growth and development of the northwest Indian squid and fitted its best growth model. In the follow-up study, we will continue to sample and expand the number of samples to further study and analyze the age and growth of squid.
4.6. Growth Rates
Growth rate includes AGR and IGR, which is of great significance to the study of cephalopod growth. The results of this study showed there was a certain difference in the ML growth rate with the age of both summer and autumn populations. The AGR of the summer population ML showed a trend of decreasing, increasing, and decreasing with time, with a peak value of 1.09 mm/d between 171–210 days. The AGR of the autumn population ML increased, decreased, and then increased, with two peaks at 151–170 days and 211–230 days, which were 0.98 mm/d and 1.73 mm/d, respectively. The water temperature of the habitat is an important factor affecting the growth of cephalopods, especially at the early stage of their life [73]. The 2020 samples may be influenced by the strong positive dipole in the Indian Ocean. The water temperature in the habitat area exceeds the appropriate temperature range for their growth and development, leading to the reduction of their growth rate and the delay of their first sexual maturation. In contrast, the growth trend of the 2019 samples was more similar to the studies of other scholars [60]. In the subadult stage, the energy intake of S. oualaniensis was mainly used for individual growth, so the AGR continued to increase [56,57]. When the ML reached a certain length, S. oualaniensis consumed more energy for reproduction, and the AGR decreased [57]. The results of this study showed that the autumn population was in the subadult stage from 151–170 d (5–6 months). S. oualaniensis has the characteristics of intermittent and multibatch spawning, and females can lay eggs for 1–3 months for the first time [10]. To increase the reproductive rate of spawning, S. oualaniensis continued to feed and grow for 171–210 days (6–7 months) after the first sexual maturity to store energy for the next spawning, which is also found in the population of S. oualaniensis in the South China Sea [13]. The age and growth of S. oualaniensis is a complex biological problem.
Authors should discuss the results and how they can be interpreted from the perspective of previous studies and of the working hypotheses. The findings and their implications should be discussed in the broadest context possible. Future research directions may also be highlighted.
5. Conclusions
In this study, we studied the growth, age composition, and population division of S. oualaniensis based on the statolith microstructure, which proved the feasibility of using the statolith microstructure to determine the age. The results showed that the majority of the squid samples in 2019 were autumn populations, the majority in 2020 were summer populations, and the growth of S. oualaniensis samples in 2019 was faster than that in 2020. The ages of the squid samples were mostly 140–260 days, and 150–230 days (5–8 months) was the subadult stage in the life history of the Indian Ocean S. oualaniensis. The population structure of S. oualaniensis is complex, and the best growth model of S. oualaniensis of different sexes in the same sea may be different. The best growth model of S. oualaniensis of the same sex in different years in the same sea may also be different. Therefore, in subsequent research, we will extend the sampling time and increase the number of samples, to provide a scientific basis for the rational exploitation of S. oualaniensis resources in the Indian Ocean.
Author Contributions
Conceptualization, H.-J.L.; methodology, H.-J.L.; software, M.-L.Z.; validation, H.-J.L., Y.-Z.O. and H.-H.W.; formal analysis, Y.-Z.O.; investigation, H.-H.W.; resources, H.-J.L.; data curation, H.-J.L.; writing—original draft preparation, Y.-Z.O.; visualization, M.-L.Z.; supervision, Z.-Y.C.; project administration, H.-J.L.; funding acquisition, H.-J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of China (NSFC 41506184) and the National Key Research and Development Program of China 489 (2019YFD0901402).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Code of Ethics of the University Department of Marine Studies; we only used specimens obtained from the surveys that were already dead.
Informed Consent Statement
Not applicable.
Acknowledgments
We would like to express our thanks for the support for two scientific surveys made by “Xinhai 1223”, which are gratefully acknowledged. Thanks to the Natural Science Foundation of China (NSFC 41506184) and the National Key Research and Development Program of China 489 (2019YFD0901402) for the partial support. Finally, we thank the editor and the anonymous reviewers whose comments greatly improved the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, Y.G.; Chen, X.J. Marine Economic Ommastrephids Resources and Fishery in the World; China Ocean Press: Beijing, China, 2005; pp. 78–93. [Google Scholar]
- Zheng, J.Y.; Xu, K.; Chen, G.X.; Hu, K.M.; Chen, J.P.; Yang, L.; Wang, Q.; Wang, X.; Wang, W.Q.; Wang, D.X. Research progress on the interaction between tropical Indian Ocean circulation dynamics and monsoon. J. Nanjing Univ. Inf. Sci. Technol. 2018, 10, 275–281. [Google Scholar]
- Qian, W.G.; Chen, X.J.; Liu, B.L.; Tian, S.Q.; Ye, X.C. The relationship between fishing ground distribution of Sthenoteuthis oualaniensisand zooplankton in the northwestern Indian Ocean in autumn. Mar. Fish. 2006, 28, 265–271. [Google Scholar]
- Xiao, G.; Xu, B.; Zhang, H.; Tang, F.H.; Chen, F.; Zhu, W.B. A study on spatial-temporal distribution and marine environmental elements of Symplectoteuthis oualaniensis fishing grounds in outer sea of Arabian Sea. S. China Fish. Sci. 2022, 18, 10–19. [Google Scholar]
- Chesalin, M.V.; Zuyev, G.V. Pelagic cephalopods of the Arabian Sea with an emphasis on Sthenoteuthis oualaniensis. Bull. Mar. Sci. 2002, 71, 209–221. [Google Scholar]
- Zuyev, G.V.; Nigmatullin, C.; Chesalin, M.; Nesis, K.N. Main results of long-term worldwide studies on tropical nektonic oceanic squid genus Sthenoteuthis: An overview of the Soviet investigations. Bull. Mar. Sci. 2002, 71, 1019–1060. [Google Scholar]
- Chen, X.J.; Lu, H.J.; Liu, B.L.; Tian, S.Q. Current exploitation and some scientific issues in the sustainable utilization of Ommastrephidae. J. Shanghai Ocean Univ. 2012, 21, 831–840. [Google Scholar]
- Zhang, J.; Chen, G.B.; Zhang, P.; Chen, Z.Z.; Fan, J.T. Estimation of purpleback flying squid (Sthenoteuthis oualaniensis) resource in the central and southern South China Sea based on fisheries acoustics and light-falling net. J. Fish. Sci. China 2014, 10, 822–831. [Google Scholar]
- Siriraksophon, S.; Sukramongkol, N.; Nakamura, Y. Exploration of oceanic squid, Sthenoteuthis oualaniensis resources in the South China Sea, Vietnamese waters. In Proceedings of the SEAFDEC Seminar on Fishery Resources in the South China Sea, Area IV: Vietnamese Waters, Bangkok, Thailand, 18–20 September 2000; pp. 181–197. [Google Scholar]
- Jereb, P.; Roper, C.F.E. Cephalopods of the World: An Annotated and Illustrated Catalog of Cephalopod Species Known to Date: Myopsid and Oegopsid Squids; FAO: Rome, Italy, 2010; Volume 2, pp. 315–318. [Google Scholar]
- Trotsenko, B.G.; Pinchukov, M.A. Mesoscale distribution features of the purpleblack squid Sthenoteuthis oualaniensis with reference to the structure of the upper quasi-homogeneous layer in the West India Ocean. Oceanology 1994, 34, 380–385. [Google Scholar]
- Nesis, K.N. Population structure of oceanic Ommastrephehids, with particular reference to Sthenoteuthis oualaniensis: A review. In Recent Advances in Fisheries Biology; Okutani, K., O’Dor, R.K., Kubodera, T., Eds.; Tokai University Press: Tokyo, Japan, 1993; pp. 375–383. [Google Scholar]
- Chen, Z.Y.; Lu, H.J.; Liu, W.; Liu, K.; Chen, X.J. Beak Microstructure Estimates of the Age, Growth, and Population Structure of Purpleback Flying Squid (Sthenoteuthis oualaniensis) in the Xisha Islands Waters of the South China Sea. Fishes 2022, 7, 187. [Google Scholar] [CrossRef]
- Clarke, M. A review of the systematics and ecology of oceanic squids. Adv. Mar. Biol. 1966, 4, 91–300. [Google Scholar]
- Chen, X.J.; Liu, B.L.; Tian, S.Q.; Qian, W.G.; Zhao, X.H. Fishery biology of purpleback squid, Sthenoteuthis oualaniensis, in the northwest Indian Ocean. Fish. Res. 2007, 83, 98–104. [Google Scholar]
- Ye, X.C.; Chen, X.J. Study of biological characteristics of Sthenoteuthis oualaniensis in the northwestern Indian Ocean. J. Shanghai Ocean. Univ. 2004, 13, 316–322. [Google Scholar]
- Shchetinnikov, A.S. Feeding spectrum of squid Sthenoteuthis oualaniensis (Oegopsida) in the Eastern Pacific. J. Mar. Biol. Assoc. U. K. 1992, 72, 849–860. [Google Scholar] [CrossRef]
- Corre, M.L.; Cherel, Y.; Lagarde, F.; Lormee, H.; Jouventin, P. Seasonal and interannual variation in the feeding ecology of a tropical oceanic seabird, the red-tailed tropicbird Phaethon rubricauda. Mar. Ecol. Prog. Ser. 2003, 255, 289–301. [Google Scholar] [CrossRef][Green Version]
- Parry, M. Feeding behavior of two ommastrephid squids Ommastrephes bartramiiand Sthenoteuthis oualaniensis off Hawaii. Mar. Ecol. Prog. Ser. 2006, 318, 229–235. [Google Scholar] [CrossRef]
- Colombo, E.M.; Luis, C.F.; José, F.E.; Felipe, G.M.; Alejandro, S.L.; David, C.S.; Carlos, J.P. Feeding habits and trophic level of the smooth hammerhead shark, Sphyrna zygaena (Carcharhiniformes: Sphyrnidae), off Ecuador. J. Mar. Biolog. Assoc. 2018, 99, 673–680. [Google Scholar]
- Arkhipkin, A.I.; Bizikov, V.A. Role of the statolith in functioning of the acceleration receptor system in squids and sepioids. J. Zool. 2000, 250, 31–55. [Google Scholar] [CrossRef]
- Radtke, R.L. Chemical and structural characteristics of statoliths from the short-finned squid Illex illecebrosus. Mar. Biol. 1983, 76, 47–54. [Google Scholar] [CrossRef]
- Hurley, G.V.; Odense, P.O.; O’Dor, R.K.; Dawe, E.G. Strontium Labeling for Verifying Daily Growth Increments in the Statolith of the Short-Finned Squid (Illex illecebrosus). Can. J. Fish. Aquat. Sci. 1985, 42, 380–383. [Google Scholar] [CrossRef]
- Lipinski, M. Methods for the validation of squid Age from statoliths. J. Mar. Biol. Assoc. U. K. 1986, 66, 505–526. [Google Scholar] [CrossRef]
- Nesis, K.N. Population structure of the squid Sthenoteuthis oualaniensis (Lesson, 1830) in the tropical West Pacific. Trudy IO AN SSSR 1977, 107, 15–29. [Google Scholar]
- Lu, H.J.; Zhang, X.; Tong, Y.H.; Tang, Y.; Liu, K.; Liu, W.; Chen, X.J. Statolith microstructure and growth characteristics of Sthenoeuthis oualaniensis in the Xisha Islands waters of the South China Sea. J. Fish. China 2020, 44, 767–776. [Google Scholar]
- Bettencourt, V.; Guerra, A. Growth increments and biomineralization process in cephalopod statoliths. J. Exp. Mar. Biol. Ecol. 2000, 248, 191–205. [Google Scholar] [CrossRef]
- Semmens, J.M.; Moltschaniwskyj, N.A. An examination of variable growth in the loliginid squid Sepioteuthis lessoniana: A whole animal and reductionist approach. Mar. Ecol. Prog. Ser. 2000, 193, 135–141. [Google Scholar] [CrossRef]
- Harman, R.F.; Young, R.E.; Reid, S.B.; Mangold, K.M.; Suzuki, T.; Hixon, R.F. Evidence for multiple spawning in the tropical oceanic squid Stenoteuthis oualaniensis (Teuthoidea: Ommastrephidae). Mar. Biol. 1989, 101, 513–519. [Google Scholar] [CrossRef]
- Yatsu, A.; Watanabe, T.; Mori, J.; Kazuya, N.; Yukimasa, I.; Toshimi, M.; Yoshihiko, K.; Yasunori, S. Interannual variability in stock abundance of the neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean during 1979–1998: Impact of driftnet fishing and oceanographic conditions. Fish. Oceanogr. 2000, 9, 163–170. [Google Scholar] [CrossRef]
- Arkhipkin, A.I. Statoliths as ‘black boxes’ (life recorders) in squid. Mar. Freshw. Res. 2005, 56, 573–583. [Google Scholar] [CrossRef]
- Lu, H.J.; Chen, X.J. Age, growth and population structure of Illex argentinus based on statolith microstructure in Southwest Atlantic Ocean. J. Fish. China 2012, 36, 1049–1056. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Lu, H.J.; Tong, Y.H.; Liu, W.; Zhang, X.; Chen, X.J. Effects of difference of individual size on beak morphology of Sthenoteuthis oualaniensis in the Xisha Islands of South China Sea. J. Fish. China 2019, 43, 2501–2510. [Google Scholar]
- Lu, H.J.; Wang, C.J.; Chen, X.J. Study on the Biological Characteristics of Jumbo Flying Squid Dosidicus gigas in the High Seas Nearby the Equator of Eastern Pacific. J. Guangdong Ocean Univ. 2014, 34, 1–8. [Google Scholar]
- Chen, X.J.; Lu, H.J.; Liu, B.L.; Chen, Y. Age, growth and population structure of jumbo flying squid, Dosidicus gigas, based on statolith microstructure off the Exclusive Economic Zone of Chilean waters. J. Mar. Biolog. Assoc. U. K. 2011, 91, 229–230. [Google Scholar] [CrossRef]
- Chen, X.J.; Liu, J.L. Morphological analysis on population structure of Sthenoteuthis oualaniensis in the north estern Indian Ocean. J. Shanghai Ocean Univ. 2007, 16, 174–179. [Google Scholar]
- Yatsu, A.; Midorikawa, S.; Shimada, T.; Uozumi, Y.J. Age and growth of neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean. Fish. Res. 1997, 29, 257–270. [Google Scholar] [CrossRef]
- Fang, Z.; Li, J.H.; Thompson, K. Age, growth, and population structure of the red flying squid (Ommastrephes bartramii) in the North Pacific Ocean, determined from beak microstructure. Fish. Bull. 2016, 114, 34–44. [Google Scholar] [CrossRef]
- Perales, R.C.; Bartolomé, A.; García-Santamaría, M.T.; Teresa, G.S.; Pedro, P.A.; Eduardo, A. Age estimation obtained from analysis of octopus (Octopus vulgaris Cuvier, 1797) beaks: Improvements and comparisons. Fish. Res. 2010, 106, 171–176. [Google Scholar] [CrossRef]
- Hu, G.Y.; Fang, Z.; Liu, B.L.; Yang, D.; Chen, X.J.; Chen, Y. Age, growth and population structure of jumbo flying squid Dosidicus gigas off the Peruvian Exclusive Economic Zone based on beak microstructure. Fish Sci. 2016, 82, 597–604. [Google Scholar] [CrossRef]
- Rodhouse, P.G.; Hatfield, E.M.C. Dynamics of Growth and Maturation in the Cephalopod Illex argentinus de Castellanos, 1960 (Teuthoidea: Ommastrephidae). Philos. Trans. Biol. Sci. 1990, 329, 1254. [Google Scholar]
- Froese, R.; Thorson, J.T.; Reyes, R.B. A Bayesian approach for estimating length-weight relationships in fishes. J. Appl. Ichthyol. 2014, 30, 78–85. [Google Scholar] [CrossRef]
- Jackson, G.D. Application and Future Potential of Statolith Increment Analysis in Squids and Sepioids. Can. J. Fish. Aquat. 1994, 51, 2612–2625. [Google Scholar] [CrossRef]
- Arkhipkin, A.I. Age and growth of the mesopelagic squid Ancistrocheirus lesueurii (Oegopsida: Ancistrocheiridae) from the central-east Atlantic based on statolith microstructure. Mar. Biol. 1997, 129, 103–111. [Google Scholar] [CrossRef]
- Malcolm, H. Modeling and Quantitative Methods in Fisheries; Chapman and Hall; CRC Press: New York, NY, USA, 2001; pp. 227–232. [Google Scholar]
- Parker, R.A.; Ricker, W.E. Handbook of Computations for Biological Statistics of Fish Populations. J. Wildl. Manag. 1960, 24, 1105–1108. [Google Scholar] [CrossRef]
- Dunning, M. A review of the systematics, distribution and biology of the arrow squid genera Ommastrephes Orbigny, 1835, Sthenoteuthis Verrill, 1880, and Ornithoteuthis Okada, 1927 (Cephalopoda, Ommastrephidae). In Systematics and Biogeography of Cephalopods; Voss, N.A., Vecchione, M., Toll, R.B., Sweeney, M.J., Eds.; Smithsonian Contributions to Zoology 586; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 425–433. [Google Scholar]
- Okutani, T.; Tung, I.H. Reviews of biology of commercially important squids in Japanese and adjacent waters, I: Symplectoteuthis oualaniensis (Lesson). Veliger 1978, 21, 87–94. [Google Scholar]
- Lu, H.J.; Tong, Y.H.; Liu, W.; Liu, K.; Dong, Z.X.; Cheng, X.; Chen, X.J. Fisheries biological characteristics of Sthenoteuthis oualaniensis in the spring season in the El Niño year of 2016 in the Zhongsha Islands waters of South China Sea. J. Fish. China 2018, 42, 912–921. [Google Scholar]
- Liu, B.L.; Chen, X.J.; Zhong, J.S. Statolith morphology of the purpleback flying squid Sthenoeuthis oualaniensis in the northwest Indian Ocean. J. Shanghai Ocean Univ. 2008, 17, 604–609. [Google Scholar]
- Yan, Y.R.; Feng, B.; Lu, H.S.; Lai, J.Y.; Du, S.Q. Fishery biology of purpleback flying squid Sthenoeuthis oualaniensis in the northern Sea areas around Nansha Island in summer. Oceanol. Limnol. Sin. 2012, 43, 1177–1186. [Google Scholar]
- Arkhipkin, A.I. Toward identification of the ecological lifestyle in nektonic squid using statolith morphometry. J. Mollus. Stud. 2003, 69, 171–178. [Google Scholar] [CrossRef][Green Version]
- Takagi, K.; Kitahara, T.; Suzuki, N.; Mori, J.; Yatsu, A. The age and growth of Sthenoteuthis oualaniensis (Cephalopoda: Ommastrephidae) in the Pacific Ocean. Bull. Mar. Sci. 2002, 71, 1105–1108. [Google Scholar]
- Lu, H.J.; Chen, X.J.; Fang, Z.; Liu, B.L. Statolith microstructure and growth characteristics of Illex argentinus in the southwest Atlantic Ocean. Prog. Fish. Sci. 2012, 33, 15–25. [Google Scholar]
- Chen, X.J.; Liu, B.L. Biology of Fishery Resources; Science Press: Beijing, China, 2017; pp. 58–62. [Google Scholar]
- Wang, H.H.; Lu, H.J.; He, J.R.; Liu, K.; Chen, X.Y.; Chen, X.J. Microstructures and growth characteristics of statoliths from Sthenoeuthis oualaniensis in the northwest Indian Ocean. Chin. J. Appl. Ecol. 2022, 1–9. [Google Scholar] [CrossRef]
- Chen, X.J.; Ma, J.; Liu, B.L.; Lu, H.J.; Cao, J. Population structure, age and growth of neon flying squid (Ommastrephes bartramii) in the northwest Pacific Ocean based on statolith microstructure. J. Fish. China 2011, 35, 1191–1198. [Google Scholar]
- Ma, J.; Chen, X.J.; Liu, B.L.; Lu, H.J.; Li, S.L.; Cao, J. Review of the influence of environment factors on microstructure of statoliths of cephalopod. J. Shanghai Ocean Univ. 2009, 18, 616–622. [Google Scholar]
- Castro, B.G.; Garrido, J.L.; Sotelo, C.G. Changes in composition of digestive gland and mantle muscle of the cuttlefish Sepia officinalis during starvation. Mar. Biol. 1992, 114, 11–20. [Google Scholar] [CrossRef]
- Lu, H.J.; Ou, Y.Z.; He, J.R.; Zhao, M.L.; Chen, Z.Y.; Chen, X.J. Age, Growth and Population Structure Analyses of the Purpleback Flying Squid Sthenoteuthis oualaniensis in the Northwest Indian Ocean by Beak Microstructure. J. Mar. Sci. Eng. 2022, 10, 1094. [Google Scholar] [CrossRef]
- Liu, B.L.; Chen, X.J.; Li, J.H.; Chen, Y. Age, growth and maturation of Sthenoteuthis oualaniensis in the eastern tropical Pacific Ocean by statolith analysis. Mar. Freshw. Res. 2016, 67, 1973–1981. [Google Scholar] [CrossRef]
- Takagi, K.; Yatsu, A. Age determination using Statolith microstructure of the purpleback fling squid, Sthenoteuthis oualaniensis, in the North Pacific Ocean. Nippon. Suisan Gakkaishi 1996, 65, 98–113. [Google Scholar]
- Sukramongkol, N.; Promjinda, S.; Prommas, R. Age and reproduction of Sthenoteuthis oualaniensis in the Bay of Bengal. In The Ecosystem-Based Fishery Management in the Bay of Bengal; Department of Fisheries (DOF), Ministry of Agriculture and Cooperatives: Bangkok, Thailand, 2007; pp. 195–205. [Google Scholar]
- Zhao, C.X.; Shen, C.Y.; Bakun, A.; Yan, Y.R.; Kang, B. Purpleback Flying Squid Sthenoteuthis oualaniensis in the South China Sea: Growth, Resources and Association with the Environment. Water 2020, 13, 65. [Google Scholar] [CrossRef]
- Snyder, R. Aspects of the biology of the giant form of Sthenoteuthis oualaniensis (Cephalopoda: Ommastrephidae) from the Arabian Sea. J. Molluscan Stud. 1998, 64, 21–34. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.H.; Du, F.Y.; Liu, B.L.; Zhang, P.; Liu, M.N.; Qiu, Y.S. Age and growth of Sthenoteuthis oualaniensis based on statolith microstructure in the South China Sea. J. Trop. Oceanogr. 2019, 38, 62–73. [Google Scholar]
- Zhao, C.X.; Chen, Z.P.; He, X.B.; Deng, Y.S.; Feng, B.; Yan, Y.R. Age, growth and population structure of purple back flying squid, Sthenoteuthis oualaniensi in the South China Sea in spring based on statolith microstructure. Acta Hydrobiol. Sin. 2017, 41, 884–890. [Google Scholar]
- Chen, X.J.; Li, J.H.; Liu, B.; Chen, Y.; Li, G.; Fang, Z.; Tian, S. Age, growth and population structure of jumbo flying squid, Dosidicus gigas, off the Costa Rica Dome. J. Mar. Biol. Assoc. U. K. 2013, 93, 2. [Google Scholar] [CrossRef]
- Lu, H.; Ou, Y.; Teng, Y.; Chen, Z.; Chen, X. Age, Growth and Population Structure Analyses of the Berryteuthis magister shevtsovi in the Japan Sea by Statolith Microstructure. Fishes 2022, 7, 215. [Google Scholar] [CrossRef]
- Yu, W.; Wen, J.; Chen, X.J.; Gong, Y.; Liu, B.L. Trans-Pacific multidecadal changes of habitat patterns of two squid species. Fish. Res. 2021, 233, 105762. [Google Scholar] [CrossRef]
- Xu, J.; Chen, X.J.; Chen, Y.; Ding, Q.; Tian, S.Q. The effect of sea surface temperature increase on the potential habitat of Ommastrephes bartramii in the Northwest Pacific Ocean. Acta Oceanol. Sin. 2016, 35, 109–116. [Google Scholar] [CrossRef]
- Lu, H.J.; Ning, X.; Liu, W.; Zhang, Y.X.; Chen, Z.Y.; Chen, X.J. Comparison in fishery biology of Sthenoteuthis oualaniensis in different climate events in the South China Sea. Oceanol. Limnol. Sin. 2021, 52, 1029–1038. [Google Scholar]
- Jackson, G.D.; Wadley, V.A. Age, growth, and reproduction of the tropical squid Nototodarus hawaiiensis (Cephalopoda: Ommastrephidae) off the north west slope of Australia. Fish. Bull. 1998, 96, 779–787. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).