
Longevity of System Functions in Biology and Biomimetics: A Matter of Robustness and Resilience



Abstract
1. Introduction
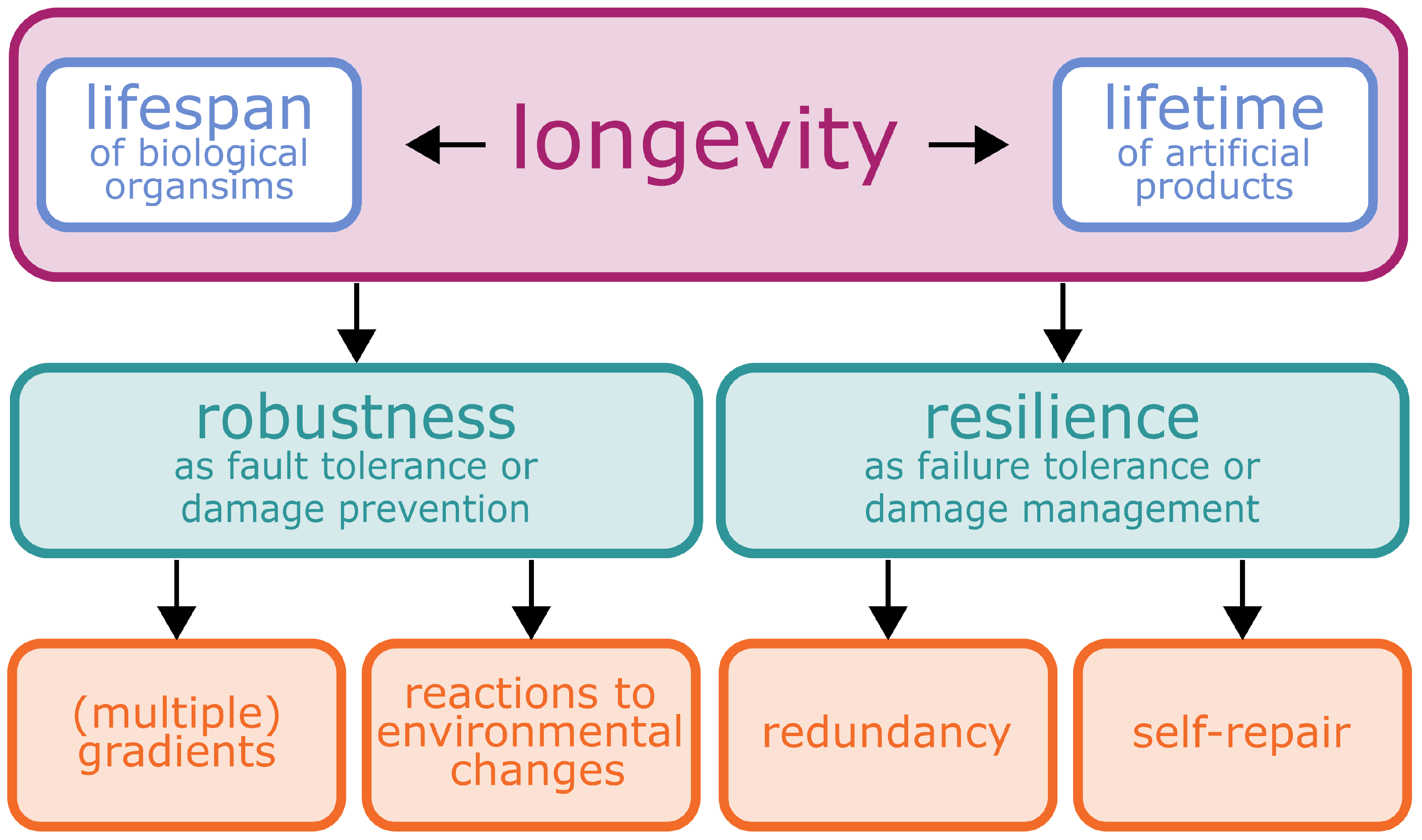
1.1. Longevity—More than a Prominent Catchphrase
1.1.1. Lifespan as a Longevity Measure in Plants
1.1.2. Product Lifetime as a Longevity Measure in Artificial Material Systems
1.2. Natural Material Cycles as Inspiration
1.3. Biological Material Systems as Inspiration
1.4. Material Systems Operate over Time
1.5. Aim of the Work
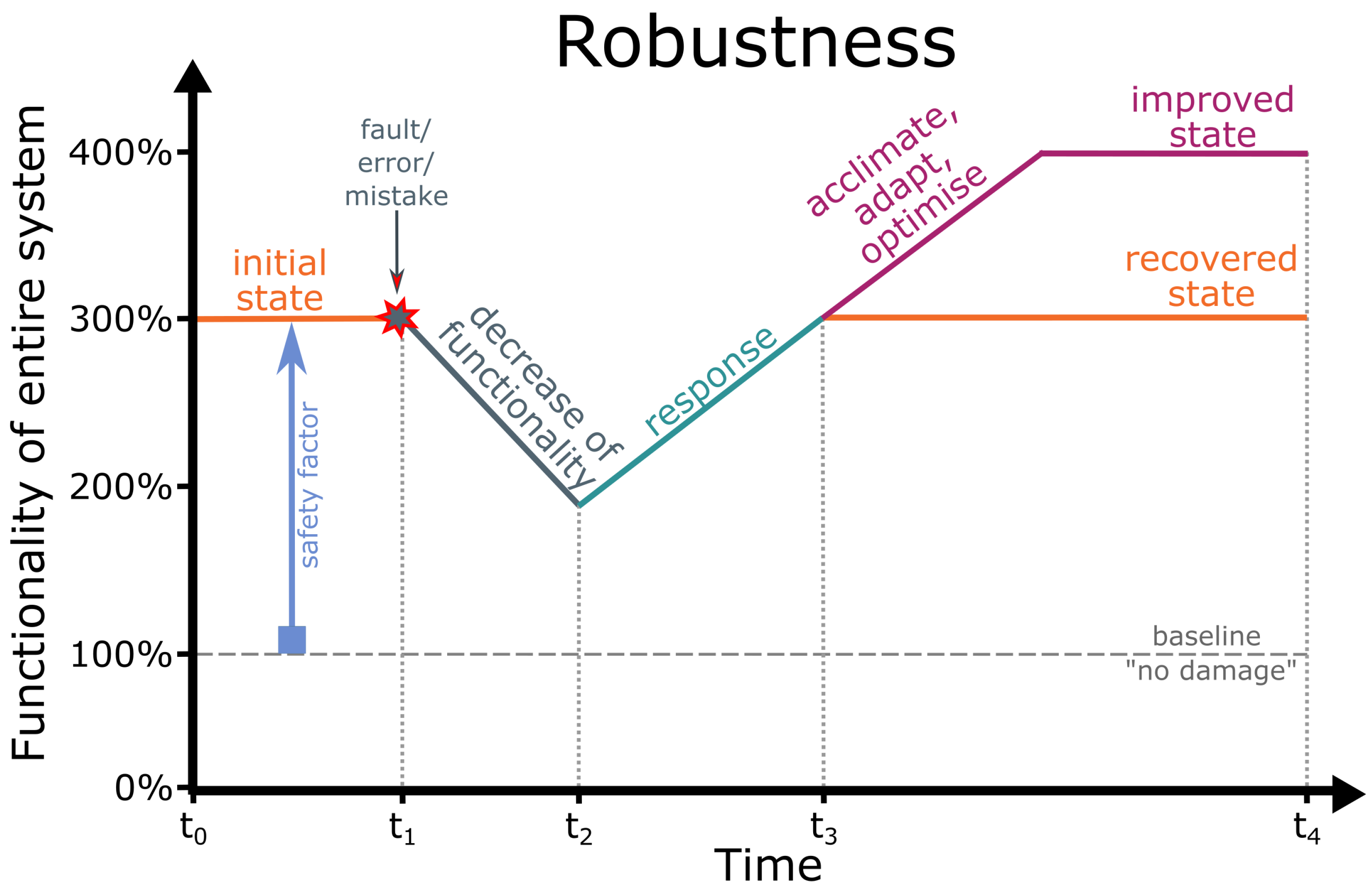
2. Robustness
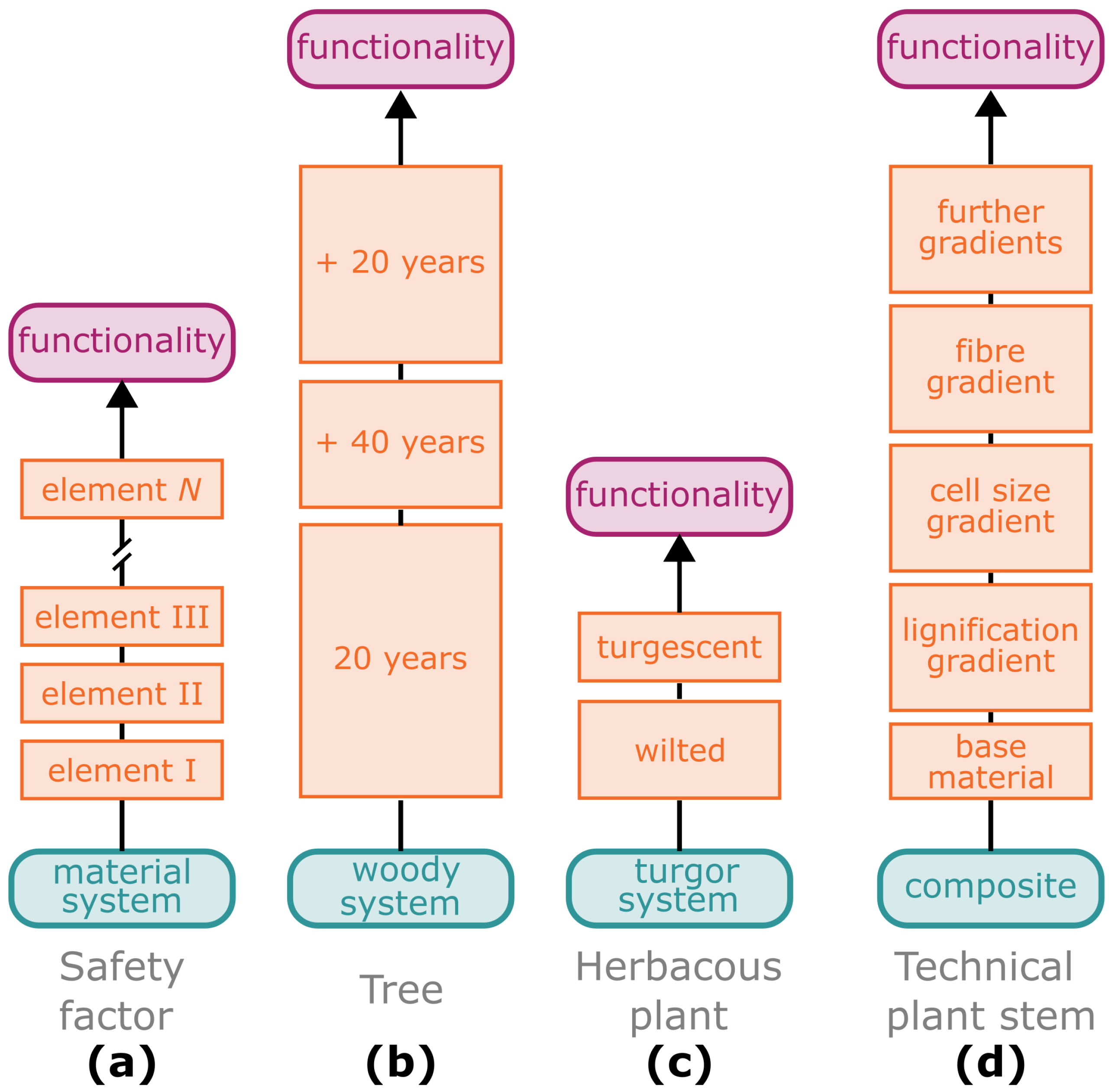
2.1. Safety Factor—The Sum of Individual Elements
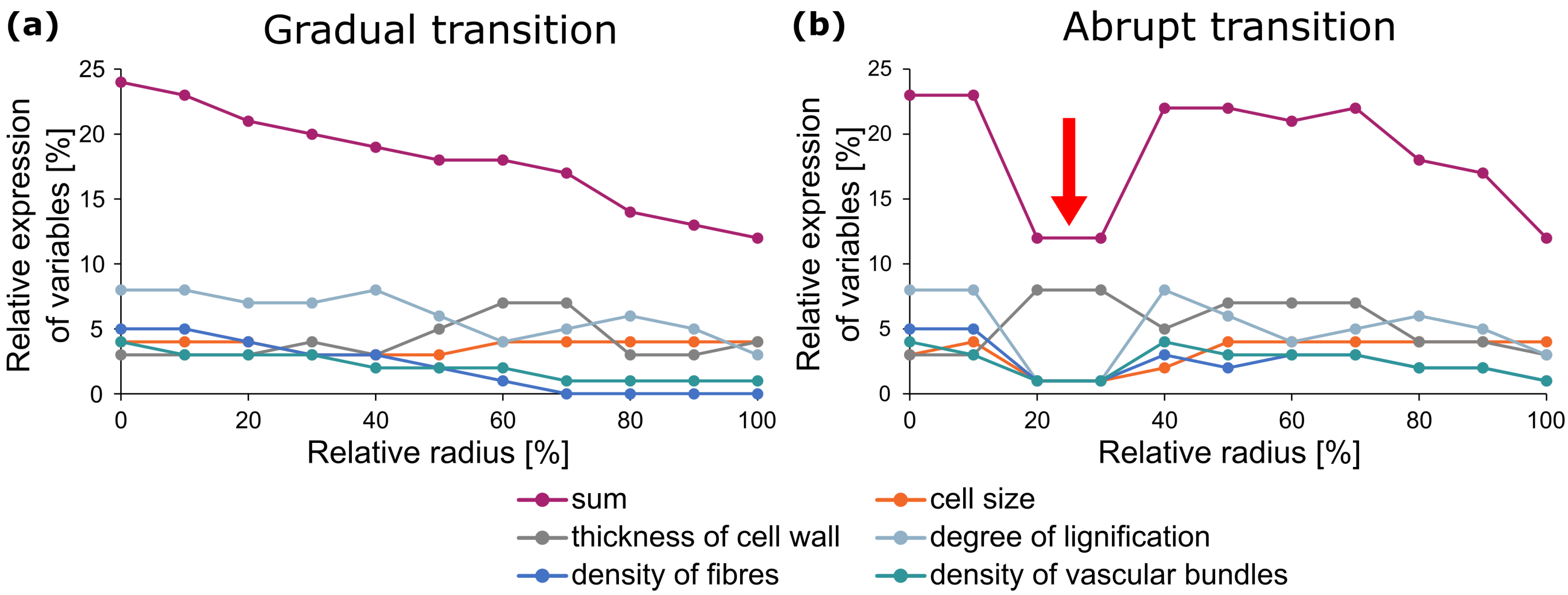
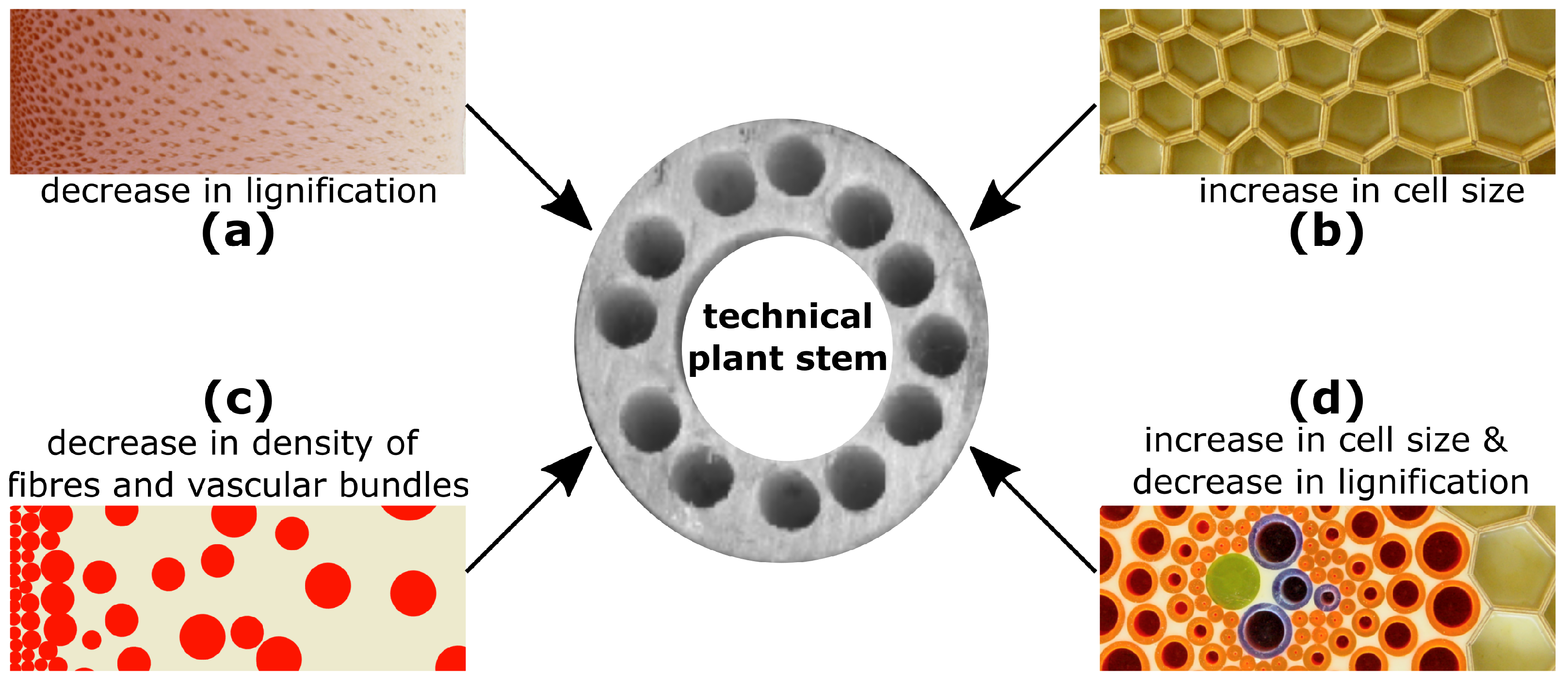
2.2. Damage Resistance through Multiple Gradients
2.3. Damage Resistance through Reactions to Environmental Changes
2.3.1. Wind-Induced Response of Plants and Plant Organs
2.3.2. Stimulus-Responsive Biomimetic Applications
2.3.3. Mechano-Stimulated Acclimation of Plants
2.3.4. “Trained” Plant Material Systems
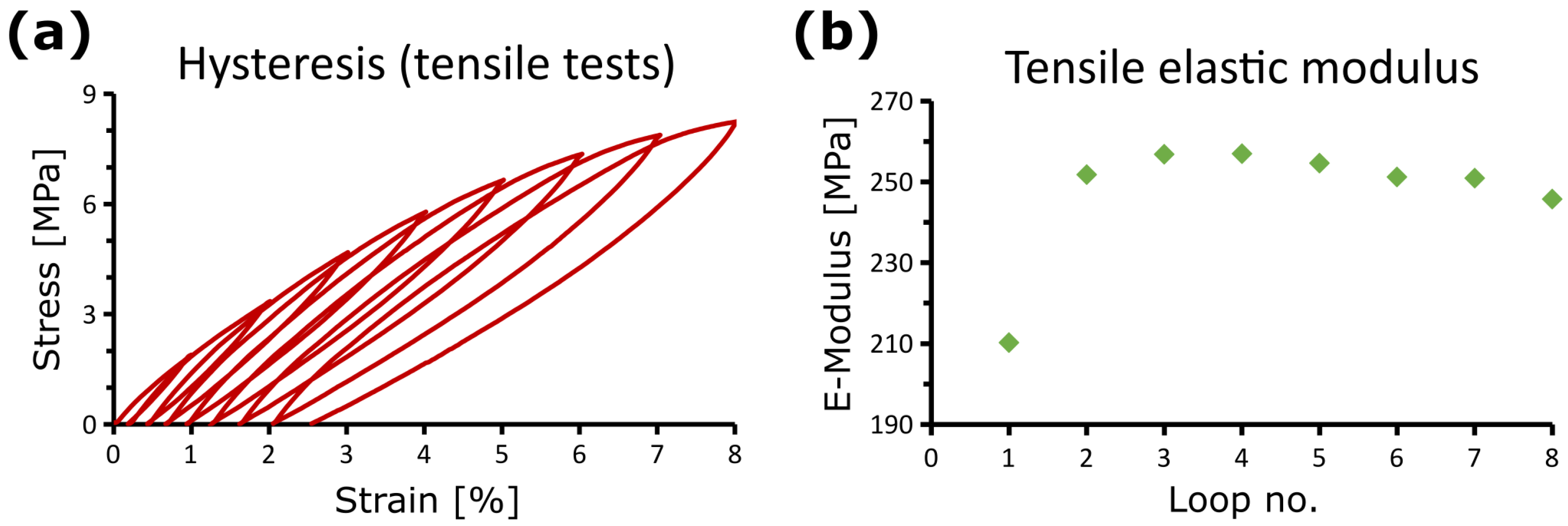
2.3.5. “Trained” Artificial Materials
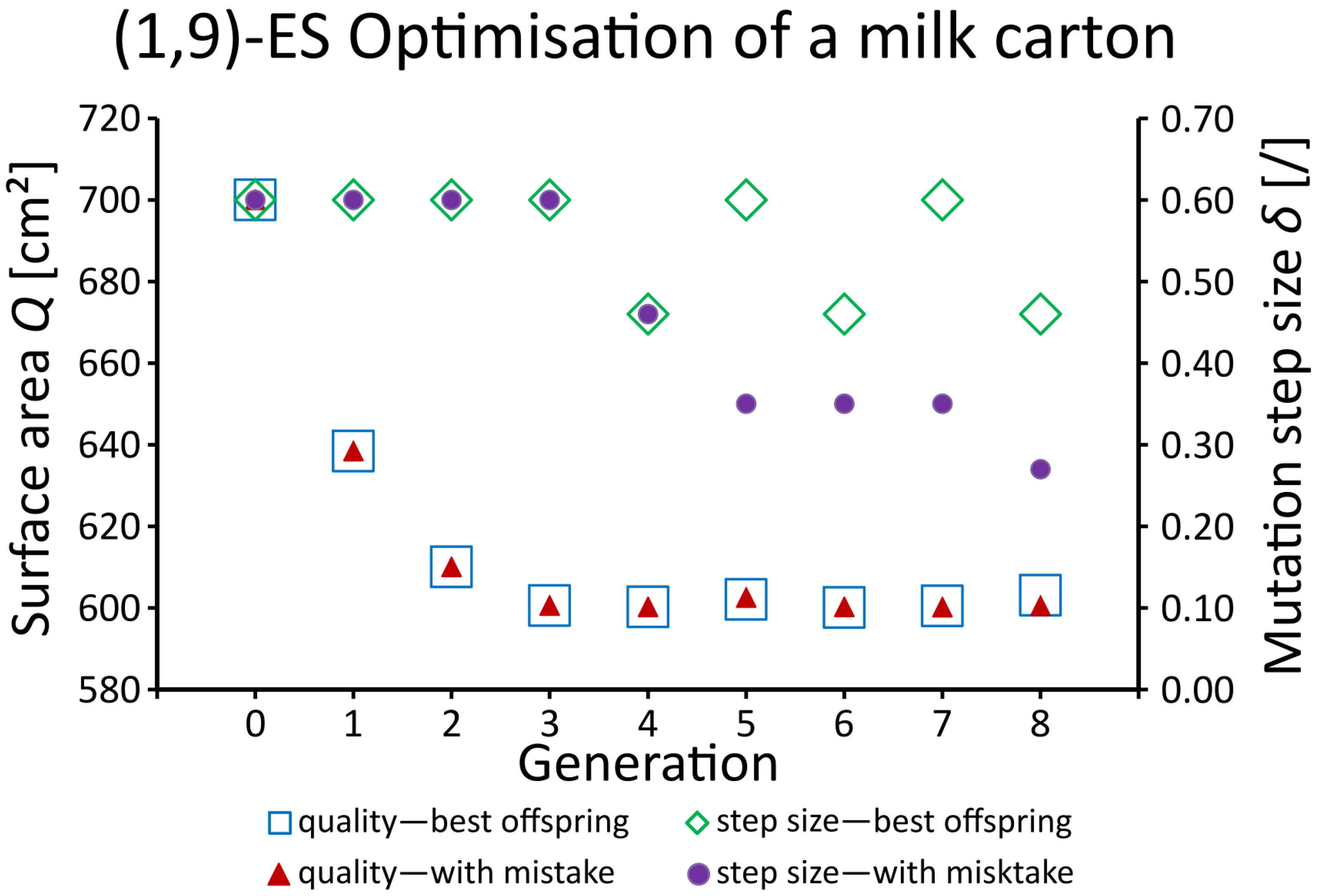
2.3.6. Adaptation and Optimisation
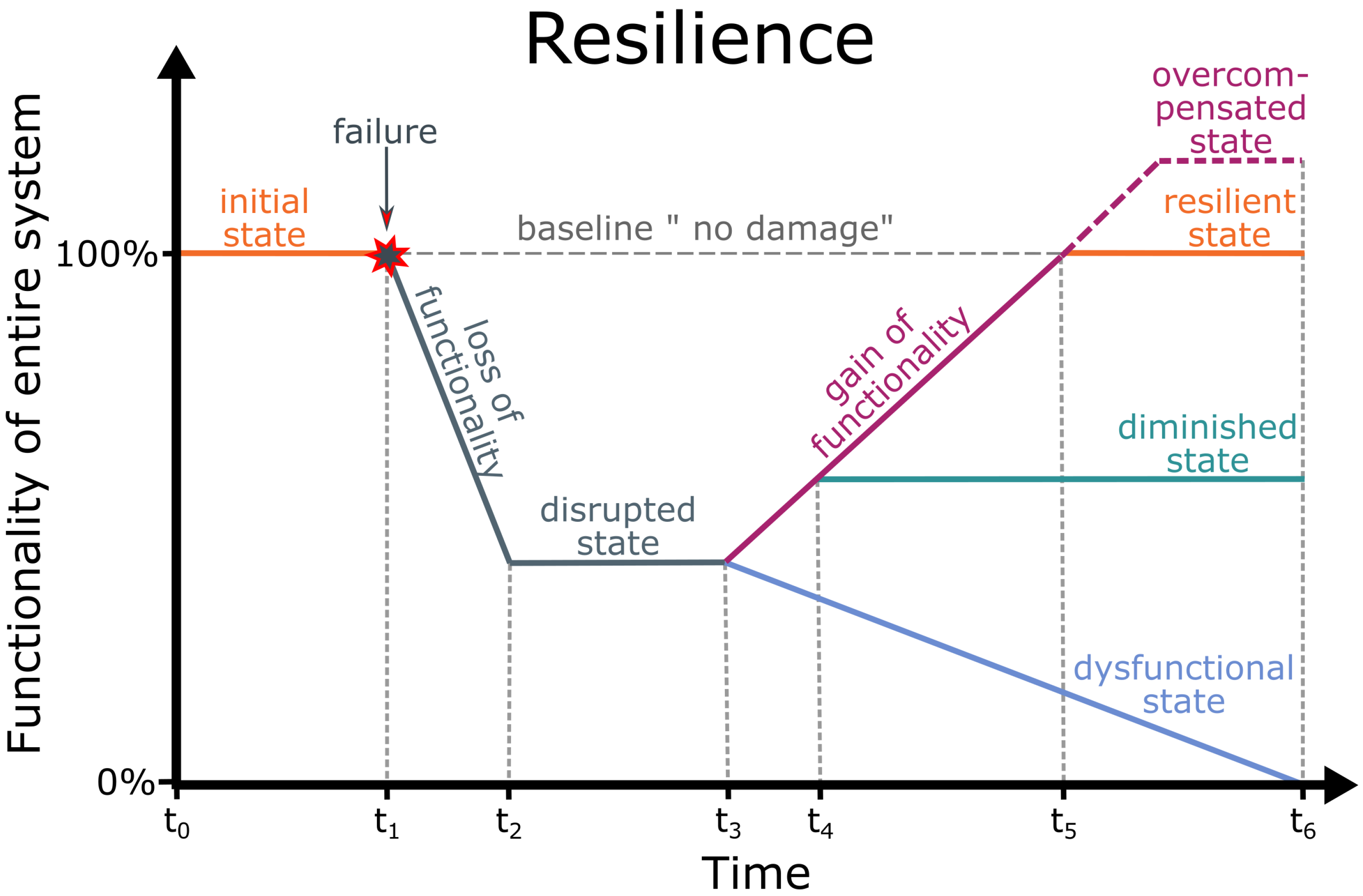
3. Resilience
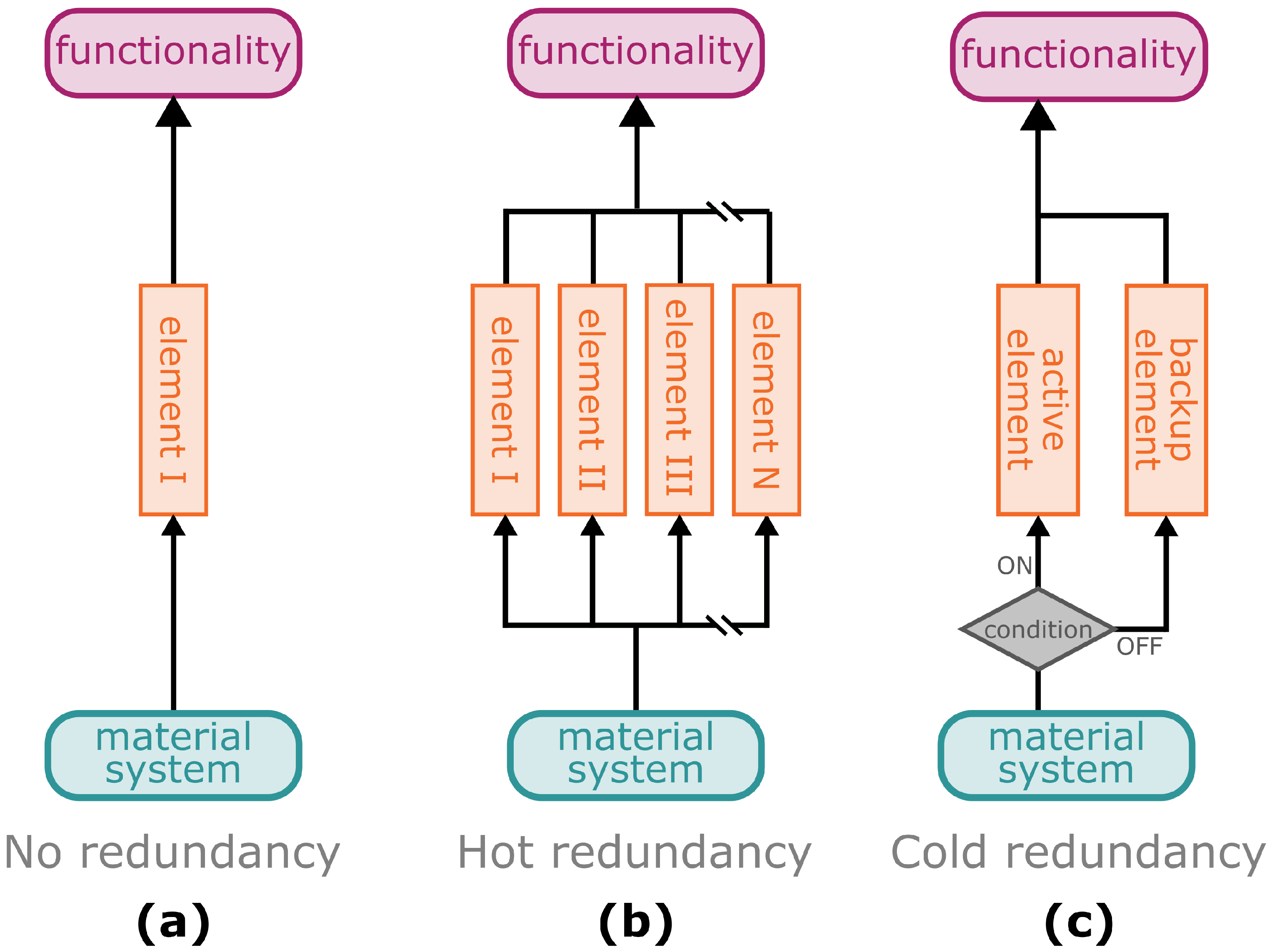
3.1. Resilience through Redundancy
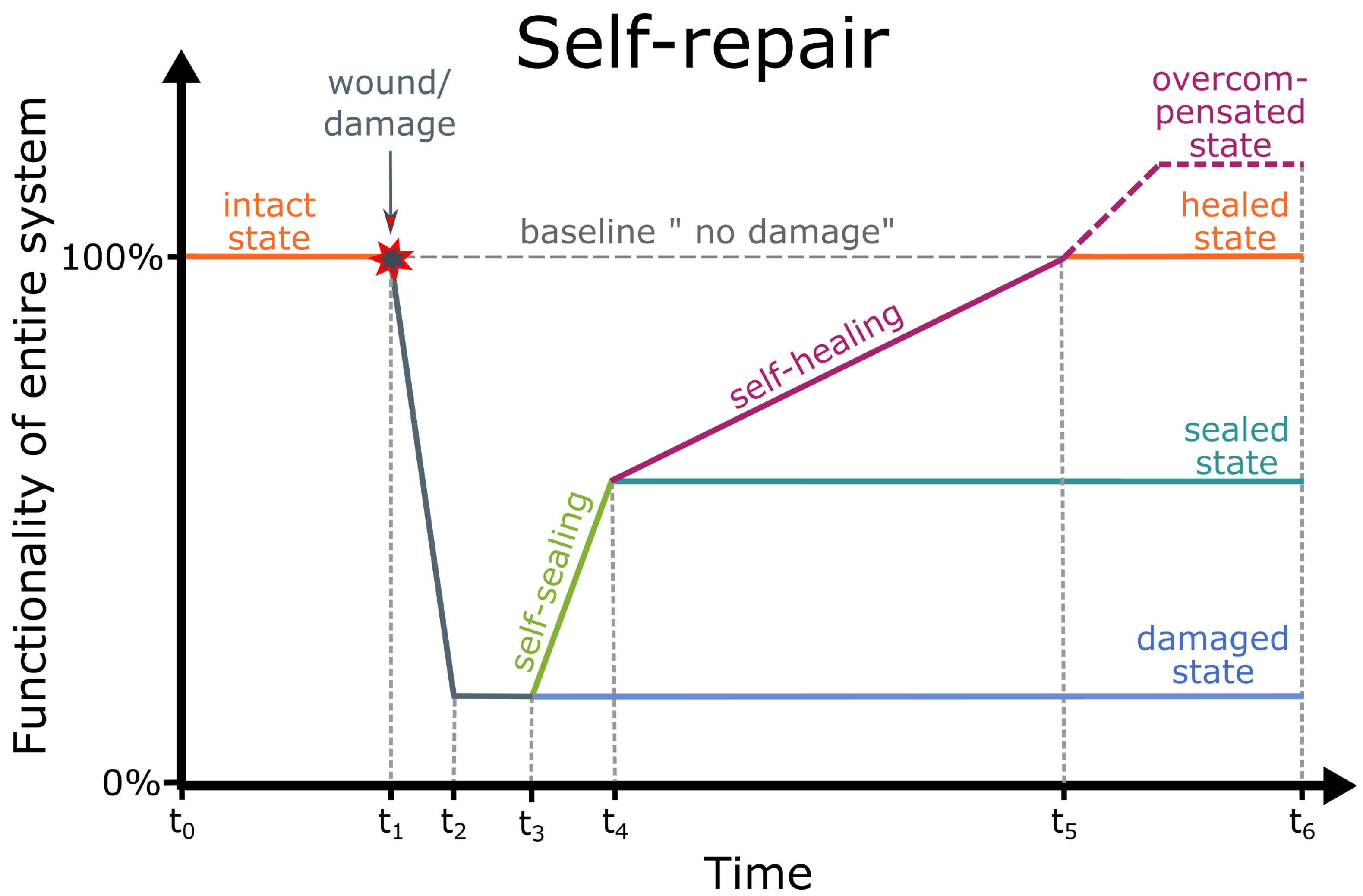
3.2. Resilience through Self-Repair of Damage
4. Conclusions
5. Glossary
- Acclimation/acclimatisation: Acclimation of individual plants to environmental constraints takes place over a time period of days and weeks, resulting in changes of gene expression and thus altered morphological, anatomical and mechanical properties [89], leading to “trained” plants. In the materials science context, this is usually referred to as adaptivity.
- Adaptation: In a biological context, adaptation is the result of genetic change in populations over evolutionary time [80]. In engineering, optimisation of systems can be achieved via evolutionary algorithms or machine learning within several generations or epochs.
- Adaptive system: “An adaptive system is a set of elements which interact with each other and has at least one process which controls the system’s adaptation, that is, the correlation between structure, function or behaviour and its environment, to increase its efficiency to achieve its goals”. ([136], p. 760)
- Bio-inspiration: Transfer of an idea derived from living organisms into a technical application [128].
- Failure: Loss of load carrying capacity in materials or structural elements (with brittle, ductile or intermediate behaviour) [137].
- Fault: A material system or structural element that can no longer satisfy the user requirements [138]. In technology, faults mean a break or a defect of the hardware; in computer science, errors occur in the software or calculations, whereas a mistake is caused by a human being.
- Functionality: The quality or status of being functional [141].
- Functional principle: The underlying principle for executing a function. Sometimes also referred to as operating principle.
- Lifespan: The period between germination and death of a plant.
- Lifetime: The period between the production of a product and its being discarded [14].
- Longevity: The attainment of the species-specific lifespan of biological organisms by ensuring the survival of the plant species through the release of spores or seeds. In an engineering context, the attainment of the consumer-expected lifespan of material systems without loss of functionality.
- Material system: Through the structural and functional interaction of various materials, a material system emerges with different chemical and physical properties, new functions and other functionalities compared with the individual materials.
- Obsolescence: The ageing of a product or artificial material system, causing it to lose functionality and usability. An extreme case is planned obsolescence, in which a shortening of the lifetime of a product is accepted or actively implemented by the manufacturer [20].
- Property: Properties or characteristics of materials, technical components or plant organs are measured via standardised or established test methods. Mechanical properties characterise the reaction to applied loads; geometric properties include cross-sectional and volumetric geometry, size and shape.
- Resilience: Ability of a system to manage damage by returning to its original functionality after (partial) failures have occurred (modified after [142]).
- Resistance: The capability of a system to withstand harmful events without significant damage and to remain unharmed or unaffected (modified after [143]).
- Redundancy: A system in which the functional failure of one (or more) contributing elements can be compensated by other elements and thus the functionality of the overall system can be maintained.
- -
- Cold redundancy: Redundant system in which one (or more) elements are passive and take over the function of one (or more) elements in case of their failure [112]. Also known as standby redundancy.
- -
- Hot redundancy: Redundant system, with all involved elements being active at the same time and taking over the failure of one (or more) elements [108]. Also known as active redundancy.
- Response: Immediate response of plants, such as wind-induced reconfiguration of leaves, branches or entire plants occurs within seconds to minutes [88].
- Robustness: The capability of a system to prevent damage and to maintain its functionality even if unforeseen faults (e.g., in hardware) or errors (e.g., in software) occur or a mistake is made by a human being [144].
- Safety factor: A dimensionless factor or percentage that describes the multiple by which the structure can carry more than their actual static load.
- Self-repair: Generic term that encompasses self-sealing and self-healing in biological and technical systems [41,145].
- -
- -
- Senescence: Senescence is a degeneration process during the ageing of plants, marking the last phase of a developmental program. It occurs in a signal-controlled and time-coordinated manner [146].
- Structural materials: Structural materials are structured in complex hierarchical architectures at various scales, making it possible to combine lightweight construction with good strength and toughness properties. A wide variety of these material systems can be found in nature (e.g., bamboo or bones); however, implementation in synthetic structures is currently only possible at high expense [27].
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roser, M.; Ortiz-Ospina, E.; Ritchie, H. Life Expectancy. Our World in Data. 2013. Available online: https://ourworldindata.org/life-expectancy (accessed on 13 March 2023).
- ISO 18458:2015; Biomimetics—Terminology, Concepts and Methodology. International Organization for Standardization: Geneva, Switzerland; Beuth: Berlin, Germany, 2015.
- Niklas, K.J.; Kutschera, U. The evolution of the land plant life cycle. New Phytol. 2010, 185, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Noodén, L.D.; Guiamét, J.J.; John, I. Whole plant senescence. In Plant Cell Death Processes; Elsevier: Amsterdam, The Netherlands, 2004; pp. 227–244. [Google Scholar] [CrossRef]
- May, M.R.; Provance, M.C.; Sanders, A.C.; Ellstrand, N.C.; Ross-Ibarra, J. A Pleistocene clone of Palmer’s oak persisting in southern California. PLoS ONE 2009, 4, e8346. [Google Scholar] [CrossRef] [PubMed]
- Koshy, K.; Pushpangadan, P. Bambusa vulgaris blooms, a leap towards extinction? Curr. Sci. 1997, 72, 622–624. [Google Scholar]
- Thomas, H.; Ougham, H.J.; Wagstaff, C.; Stead, A.D. Defining senescence and death. J. Exp. Bot. 2003, 54, 1127–1132. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H. Senescence, ageing and death of the whole plant. New Phytol. 2013, 197, 696–711. [Google Scholar] [CrossRef]
- De Witte, L.C.; Stöcklin, J. Longevity of clonal plants: Why it matters and how to measure it. Ann. Bot. 2010, 106, 859–870. [Google Scholar] [CrossRef]
- Bobich, E.G.; Nobel, P.S. Vegetative reproduction as related to biomechanics, morphology and anatomy of four cholla cactus species in the Sonoran Desert. Ann. Bot. 2001, 87, 485–493. [Google Scholar] [CrossRef][Green Version]
- Mylo, M.D.; Hoppe, A.; Pastewka, L.; Speck, T.; Speck, O. Elastic property and fracture mechanics of lateral branch-branch junctions in cacti: A case study of Opuntia ficus-indica and Cylindropuntia bigelovii. Front. Plant Sci. 2022, 13, 950860. [Google Scholar] [CrossRef]
- Sallon, S.; Solowey, E.; Cohen, Y.; Korchinsky, R.; Egli, M.; Woodhatch, I.; Simchoni, O.; Kislev, M. Germination, genetics and growth of an ancient date seed. Science 2008, 320, 1464. [Google Scholar] [CrossRef]
- Poppinga, S.; Nestle, N.; Šandor, A.; Reible, B.; Masselter, T.; Bruchmann, B.; Speck, T. Hygroscopic motions of fossil conifer cones. Sci. Rep. 2017, 7, 40302. [Google Scholar] [CrossRef]
- Murakami, S.; Oguchi, M.; Tasaki, T.; Daigo, I.; Hashimoto, S. Lifespan of commodities, part I: The creation of a database and its review. J. Ind. Ecol. 2010, 14, 598–612. [Google Scholar] [CrossRef]
- Cooper, T. Inadequate life? Evidence of consumer attitudes to product obsolescence. J. Consum. Policy 2004, 27, 421–449. [Google Scholar] [CrossRef]
- Stiftung Warentest. Gerade Gekauft und Schon Wieder Hin? 2013. Available online: https://www.test.de/Geplante-Obsoleszenz-Gerade-gekauft-und-schon-wieder-hin-4596260-0/ (accessed on 12 January 2023). (In German).
- Van Nes, N.; Cramer, J. Influencing product lifetime through product design. Bus. Strategy Environ. 2005, 14, 286–299. [Google Scholar] [CrossRef]
- Khan, M.A.; Mittal, S.; West, S.; Wuest, T. Review on upgradability—A product lifetime extension strategy in the context of product service systems. J. Clean. Prod. 2018, 204, 1154–1168. [Google Scholar] [CrossRef]
- Gagg, C.R.; Lewis, P.R. Wear as a product failure mechanism–overview and case studies. Eng. Fail. Anal. 2007, 14, 1618–1640. [Google Scholar] [CrossRef]
- Proske, M.; Winzer, J.; Marwede, M.; Nissen, N.F.; Lang, K.D. Obsolescence of electronics–The example of smartphones. In Proceedings of the 2016 Electronics Goes Green 2016+(EGG), Berlin, Germany, 6–9 September 2016; pp. 1–8. [Google Scholar] [CrossRef]
- Rivera, J.L.; Lallmahomed, A. Environmental implications of planned obsolescence and product lifetime: A literature review. Int. J. Sustain. Eng. 2016, 9, 119–129. [Google Scholar] [CrossRef]
- Maitre-Ekern, E.; Dalhammar, C. Regulating planned obsolescence: A review of legal approaches to increase product durability and reparability in Europe. Rev. Eur. Comp. Int. Environ. Law 2016, 25, 378–394. [Google Scholar] [CrossRef]
- Kara, S.; Hauschild, M.; Sutherland, J.; McAloone, T. Closed-loop systems to circular economy: A pathway to environmental sustainability? CIRP Ann. 2022, 71, 505–528. [Google Scholar] [CrossRef]
- Kral, U.; Kellner, K.; Brunner, P.H. Sustainable resource use requires “clean cycles”’ and safe “final sinks”. Sci. Total Environ. 2013, 461, 819–822. [Google Scholar] [CrossRef]
- De Schoenmakere, M.; Gillabel, J. Circular by Design: Products in the Circular Economy; European Environment Agency: Copenhagen, Denmark, 2017; Volume 6. Available online: https://www.eea.europa.eu/publications/circular-by-design (accessed on 7 December 2022).
- United Nations. Report of the Inter-Agency and Expert Group on Sustainable Development Goal Indicators. Statistical Commission Forty-Seventh Session. 8–11 March 2016. Available online: https://digitallibrary.un.org/record/821651 (accessed on 7 December 2022).
- Wegst, U.G.; Bai, H.; Saiz, E.; Tomsia, A.P.; Ritchie, R.O. Bioinspired structural materials. Nat. Mater. 2015, 14, 23–36. [Google Scholar] [CrossRef]
- Ashby, M.F.; Shercliff, H.; Cebon, D. Materials: Engineering, Science, Processing and Design; Butterworth-Heinemann: Oxford, UK, 2018. [Google Scholar]
- Lloyd, G.E.R.; Lloyd, G.E.R. Adversaries and Authorities: Investigations Into Ancient Greek and Chinese Science; Cambridge University Press: Cambridge, UK, 1996; Volume 42. [Google Scholar]
- Krohs, U. Der Funktionsbegriff in der Biologie. In Wissenschaftstheorie; Brill Mentis: Paderborn, Germany, 2009; pp. 287–306. (In German) [Google Scholar]
- Johnson, R.; Cureton, A. Kant’s Moral Philosophy. In The Stanford Encyclopedia of Philosophy, Fall 2022 ed.; Zalta, E.N., Nodelman, U., Eds.; Metaphysics Research Lab, Stanford University: Stanford, CA, USA, 2022. [Google Scholar]
- Mayr, E. Cause and effect in biology: Kinds of causes, predictability and teleology are viewed by a practicing biologist. Science 1961, 134, 1501–1506. [Google Scholar] [CrossRef]
- Huiskes, R. If bone is the answer, then what is the question? J. Anat. 2000, 197, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Diesendruck, C.E.; Sottos, N.R.; Moore, J.S.; White, S.R. Biomimetic self-healing. Angew. Chem. Int. Ed. 2015, 54, 10428–10447. [Google Scholar] [CrossRef] [PubMed]
- Cohades, A.; Branfoot, C.; Rae, S.; Bond, I.; Michaud, V. Progress in self-healing fiber-reinforced polymer composites. Adv. Mater. Interfaces 2018, 5, 1800177. [Google Scholar] [CrossRef]
- Bauer, G.; Nellesen, A.; Speck, T. Biological lattices in fast self-repair mechanisms in plants and the development of bio-inspired self-healing polymers. WIT Trans. Ecol. Environ. 2010, 138, 453–459. [Google Scholar] [CrossRef]
- Mylo, M.D.; Krüger, F.; Speck, T.; Speck, O. Self-repair in cacti branches: Comparative analyses of their morphology, anatomy and biomechanics. Int. J. Mol. Sci. 2020, 21, 4630. [Google Scholar] [CrossRef]
- Rampf, M.; Speck, O.; Speck, T.; Luchsinger, R. Investigation of a fast mechanical self-repair mechanism for inflatable structures. Int. J. Eng. Sci. 2013, 63, 61–70. [Google Scholar] [CrossRef]
- Wu, X.F.; Rahman, A.; Zhou, Z.; Pelot, D.D.; Sinha-Ray, S.; Chen, B.; Payne, S.; Yarin, A.L. Electrospinning core-shell nanofibers for interfacial toughening and self-healing of carbon-fiber/epoxy composites. J. Appl. Polym. Sci. 2013, 129, 1383–1393. [Google Scholar] [CrossRef]
- Bekas, D.; Tsirka, K.; Baltzis, D.; Paipetis, A.S. Self-healing materials: A review of advances in materials, evaluation, characterization and monitoring techniques. Compos. Part B Eng. 2016, 87, 92–119. [Google Scholar] [CrossRef]
- Speck, O.; Schmauder, K.; Speck, T.; Paul-Victor, C. Wound reactions in stems of Leonurus cardiaca: A morphological, anatomical and biomechanical study. Botany 2020, 98, 81–89. [Google Scholar] [CrossRef]
- Yang, Y.; Davydovich, D.; Hornat, C.C.; Liu, X.; Urban, M.W. Leaf-inspired self-healing polymers. Chem 2018, 4, 1928–1936. [Google Scholar] [CrossRef]
- Drost, H.G.; Janitza, P.; Grosse, I.; Quint, M. Cross-kingdom comparison of the developmental hourglass. Curr. Opin. Genet. Dev. 2017, 45, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Anandan, S.; Rudolph, A.; Speck, T.; Speck, O. Comparative morphological and anatomical study of self-repair in succulent cylindrical plant organs. Flora 2018, 241, 1–7. [Google Scholar] [CrossRef]
- Bauer, G.; Speck, T. Restoration of tensile strength in bark samples of Ficus benjamina due to coagulation of latex during fast self-healing of fissures. Ann. Bot. 2012, 109, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Speck, O.; Schlechtendahl, M.; Borm, F.; Kampowski, T.; Speck, T. Humidity-dependent wound sealing in succulent leaves of Delosperma cooperi—An adaptation to seasonal drought stress. Beilstein J. Nanotechnol. 2018, 9, 175–186. [Google Scholar] [CrossRef]
- Hesse, L.; Kampowski, T.; Leupold, J.; Caliaro, S.; Speck, T.; Speck, O. Comparative analyses of the self-sealing mechanisms in leaves of Delosperma cooperi and Delosperma ecklonis (Aizoaceae). Int. J. Mol. Sci. 2020, 21, 5768. [Google Scholar] [CrossRef]
- Konrad, W.; Flues, F.; Schmich, F.; Speck, T.; Speck, O. An analytic model of the self-sealing mechanism of the succulent plant Delosperma cooperi. J. Theor. Biol. 2013, 336, 96–109. [Google Scholar] [CrossRef]
- Klein, H.; Hesse, L.; Boljen, M.; Kampowski, T.; Butschek, I.; Speck, T.; Speck, O. Finite element modelling of complex movements during self-sealing of ring incisions in leaves of Delosperma cooperi. J. Theor. Biol. 2018, 458, 184–206. [Google Scholar] [CrossRef]
- Speck, O.; Langer, M.; Mylo, M.D. Plant-inspired damage control—An inspiration for sustainable solutions in the Anthropocene. Anthr. Rev. 2022, 9, 220–236. [Google Scholar] [CrossRef]
- Carlson, J.M.; Doyle, J. Complexity and robustness. Proc. Natl. Acad. Sci. USA 2002, 99, 2538–2545. [Google Scholar] [CrossRef]
- Kitano, H. Biological robustness. Nat. Rev. Genet. 2004, 5, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Kitano, H. Towards a theory of biological robustness. Mol. Syst. Biol. 2007, 3, 137. [Google Scholar] [CrossRef]
- Lesne, A. Robustness: Confronting lessons from physics and biology. Biol. Rev. 2008, 83, 509–532. [Google Scholar] [CrossRef] [PubMed]
- Kiakojouri, F.; De Biagi, V.; Abbracciavento, L. Design for robustness: Bio-Inspired perspectives in structural engineering. Biomimetics 2023, 8, 95. [Google Scholar] [CrossRef] [PubMed]
- Gere, J.M.; Goodno, B.J. Mechanics of Materials, 8th ed.; Cengage Learning: Stanford, CA, USA, 2013. [Google Scholar]
- Randall, F.A. The safety factor of structures in history. Prof. Saf. 1976, 21, 12–18. [Google Scholar]
- Möller, N.; Hansson, S.O. Principles of engineering safety: Risk and uncertainty reduction. Reliab. Eng. Syst. Saf. 2008, 93, 798–805. [Google Scholar] [CrossRef]
- Stamatelatos, M. Probabilistic Risk Assessment: What Is It and Why Is It Worth Performing It? 2000. Available online: https://copswiki.org/w/pub/Common/M1922/pra-Probabilistic%20Risk%20Assessment-What%20is%20it%20and%20why%20is%20it%20work%20performing.pdf (accessed on 12 January 2022).
- Doorn, N.; Hansson, S.O. Should probabilistic design replace safety factors? Philos. Technol. 2011, 24, 151–168. [Google Scholar] [CrossRef]
- Duncan, J.M. Factors of safety and reliability in geotechnical engineering. J. Geotech. Eng. 2000, 126, 307–316. [Google Scholar] [CrossRef]
- Niklas, K.J. Determinate growth of Allium sativum peduncles: Evidence of determinate growth as a design factor for biomechanical safety. Am. J. Bot. 1990, 77, 762–771. [Google Scholar] [CrossRef]
- Langer, M.; Kelbel, M.; Müller, C.; Speck, T.; Speck, O. Twist-to-bend ratios and safety factors of petioles having various geometries, shapes and sizes. Front. Plant Sci. 2021, 12, 2586. [Google Scholar] [CrossRef]
- King, D.; Loucks, O.L. The theory of tree bole and branch form. Radiat. Environ. Biophys. 1978, 15, 141–165. [Google Scholar] [CrossRef]
- Lehmann, L.S.; Kampowski, T.; Caliaro, M.; Speck, T.; Speck, O. Drooping of Gerbera flower heads: Mechanical and structural studies of a well known phenomenon. Biol. Lett. 2019, 15, 20190254. [Google Scholar] [CrossRef]
- Liu, Z.; Meyers, M.A.; Zhang, Z.; Ritchie, R.O. Functional gradients and heterogeneities in biological materials: Design principles, functions and bioinspired applications. Prog. Mater. Sci. 2017, 88, 467–498. [Google Scholar] [CrossRef]
- Langer, M.; Speck, T.; Speck, O. The transition zone between petiole and lamina: A functionally crucial but often overlooked leaf trait. Plants 2021, 10, 774. [Google Scholar] [CrossRef] [PubMed]
- Sacher, M.; Lautenschläger, T.; Kempe, A.; Neinhuis, C. Umbrella leaves—Biomechanics of transition zone from lamina to petiole of peltate leaves. Bioinspir. Biomimetics 2019, 14, 046011. [Google Scholar] [CrossRef] [PubMed]
- Rjosk, A.; Neinhuis, C.; Lautenschläger, T. Anatomy and biomechanics of peltate Begonia leaves—Comparative case studies. Plants 2022, 11, 3297. [Google Scholar] [CrossRef] [PubMed]
- Addicott, F.T. Abscission; University of California Press: Berkeley, CA, USA, 1982. [Google Scholar]
- Primka, E.J.; Smith, W.K. Synchrony in fall leaf drop: Chlorophyll degradation, color change, and abscission layer formation in three temperate deciduous tree species. Am. J. Bot. 2019, 106, 377–388. [Google Scholar] [CrossRef]
- Mylo, M.D.; Hesse, L.; Masselter, T.; Leupold, J.; Drozella, K.; Speck, T.; Speck, O. Morphology and anatomy of branch–branch junctions in Opuntia ficus-indica and Cylindropuntia bigelovii: A comparative study supported by mechanical tissue quantification. Plants 2021, 10, 2313. [Google Scholar] [CrossRef]
- Shtein, I.; Koyfman, A.; Eshel, A.; Bar-On, B. Autotomy in plants: Organ sacrifice in Oxalis leaves. J. R. Soc. Interface 2019, 16, 20180737. [Google Scholar] [CrossRef]
- Bateman, P.; Fleming, P. To cut a long tail short: A review of lizard caudal autotomy studies carried out over the last 20 years. J. Zool. 2009, 277, 1–14. [Google Scholar] [CrossRef]
- Spatz, H.C.; Beismann, H.; Brüchert, F.; Emanns, A.; Speck, T. Biomechanics of the giant reed Arundo donax. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1997, 352, 1–10. [Google Scholar] [CrossRef]
- Milwich, M.; Speck, T.; Speck, O.; Stegmaier, T.; Planck, H. Biomimetics and technical textiles: Solving engineering problems with the help of nature’s wisdom. Am. J. Bot. 2006, 93, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Speck, O.; Spatz, H.C. Damped oscillations of the giant reed Arundo donax (Poaceae). Am. J. Bot. 2004, 91, 789–796. [Google Scholar] [CrossRef]
- Rüggeberg, M.; Burgert, I.; Speck, T. Structural and mechanical design of tissue interfaces in the giant reed Arundo donax. J. R. Soc. Interface 2010, 7, 499–506. [Google Scholar] [CrossRef]
- Milwich, M.; Planck, H.; Speck, T.; Speck, O. The technical plant stem: A biomimetically inspired narrow fabric. Band Und Flecht Ind. 2007, 44, 34–38. [Google Scholar]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: Cham, Switzerland, 2008; Volume 2, pp. 4–6. [Google Scholar]
- Gardiner, B.; Berry, P.; Moulia, B. Wind impacts on plant growth, mechanics and damage. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- De Langre, E.; Gutierrez, A.; Cossé, J. On the scaling of drag reduction by reconfiguration in plants. Comptes Rendus Mec. 2012, 340, 35–40. [Google Scholar] [CrossRef]
- Harder, D.L.; Speck, O.; Hurd, C.L.; Speck, T. Reconfiguration as a prerequisite for survival in highly unstable flow-dominated habitats. J. Plant Growth Regul. 2004, 23, 98–107. [Google Scholar] [CrossRef]
- Speck, O. Field measurements of wind speed and reconfiguration in Arundo donax (Poaceae) with estimates of drag forces. Am. J. Bot. 2003, 90, 1253–1256. [Google Scholar] [CrossRef]
- Moulia, B.; Coutand, C.; Julien, J.L. Mechanosensitive control of plant growth: Bearing the load, sensing, transducing and responding. Front. Plant Sci. 2015, 6, 52. [Google Scholar] [CrossRef]
- Knight, T.A. XI. Account of some experiments on the descent of the sap in trees. In a letter from Thomas Andrew Knight, Esq. to the Right Hon. Sir Joseph Banks, Bart. K. B. P. R. S. Philos. Trans. R. Soc. Lond. 1803, 93, 277–289. [Google Scholar] [CrossRef]
- Jaffe, M.J. Thigmomorphogenesis: The response of plant growth and development to mechanical stimulation. Planta 1973, 114, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Telewski, F.W. Mechanosensing and plant growth regulators elicited during the thigmomorphogenetic response. Front. For. Glob. Chang. 2021, 3, 147. [Google Scholar] [CrossRef]
- Badel, E.; Ewers, F.W.; Cochard, H.; Telewski, F.W. Acclimation of mechanical and hydraulic functions in trees: Impact of the thigmomorphogenetic process. Front. Plant Sci. 2015, 6, 266. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, H. Genetic variability in Plantago species in relation to their ecology: 4. Ecotypic differentiation in P. major. Theor. Appl. Genet. 1989, 77, 749–759. [Google Scholar] [CrossRef]
- Ding, F.; Kareem, A. Tall buildings with dynamic facade under winds. Engineering 2020, 6, 1443–1453. [Google Scholar] [CrossRef]
- Li, S.; Wang, K. Plant-inspired adaptive structures and materials for morphing and actuation: A review. Bioinspir. Biomimetics 2016, 12, 011001. [Google Scholar] [CrossRef]
- Lienhard, J.; Schleicher, S.; Poppinga, S.; Masselter, T.; Milwich, M.; Speck, T.; Knippers, J. Flectofin: A hingeless flapping mechanism inspired by nature. Bioinspir. Biomimetics 2011, 6, 045001. [Google Scholar] [CrossRef]
- Körner, A.; Born, L.; Mader, A.; Sachse, R.; Saffarian, S.; Westermeier, A.; Poppinga, S.; Bischoff, M.; Gresser, G.; Milwich, M.; et al. Flectofold—A biomimetic compliant shading device for complex free form facades. Smart Mater. Struct. 2017, 27, 017001. [Google Scholar] [CrossRef]
- Bechert, S.; Aldinger, L.; Wood, D.; Knippers, J.; Menges, A. Urbach Tower: Integrative structural design of a lightweight structure made of self-shaped curved cross-laminated timber. Structures 2021, 33, 3667–3681. [Google Scholar] [CrossRef]
- Menges, A.; Reichert, S. Material capacity: Embedded responsiveness. Archit. Des. 2012, 82, 52–59. [Google Scholar] [CrossRef]
- Walther, A. From responsive to adaptive and interactive materials and materials systems: A roadmap. Adv. Mater. 2020, 32, 1905111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zeng, H.; Eklund, A.; Guo, H.; Priimagi, A.; Ikkala, O. Feedback-controlled hydrogels with homeostatic oscillations and dissipative signal transduction. Nat. Nanotechnol. 2022, 17, 1303–1310. [Google Scholar] [CrossRef]
- Schwefel, H.P. Evolution and Optimum Seeking: The Sixth Generation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1993. [Google Scholar]
- Rechenberg, I. Cybernetic solution path of an experimental problem. R. Aircr. Establ. Libr. Transl. 1965, 1122. [Google Scholar]
- Standard VDI:6224; Part 1 Biomimetic Optimization—Application of Evolutionary Algorithms. Verein Deutscher Ingenieure: Berlin, Gremany; Beuth: Berlin, Gremany, 2012.
- Rechenberg, I. Case studies in evolutionary experimentation and computation. Comput. Methods Appl. Mech. Eng. 2000, 186, 125–140. [Google Scholar] [CrossRef]
- Herdy, M. Evolution strategies with subjective selection. In Proceedings of the Parallel Problem Solving from Nature—PPSN IV: International Conference on Evolutionary Computation—The 4th International Conference on Parallel Problem Solving from Nature, Berlin, Germany, 22–26 September 1996; Springer: Cham, Switzerland, 1996; pp. 22–31. [Google Scholar]
- Sauer, S.; Herdy, M.; Speck, T.; Speck, O. Evolutionsstrategie: Optimieren nach dem Vorbild der Natur–Interdisziplinäre Arbeitsweise der Biomechanik und Bionik. Prax. Nat. Biol. 2010, 59, 34–41. (In German) [Google Scholar]
- Spatz, H.; Kohler, L.; Niklas, K. Mechanical behaviour of plant tissues: Composite materials or structures? J. Exp. Biol. 1999, 202, 3269–3272. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Plant cell wall extensibility: Connecting plant cell growth with cell wall structure, mechanics and the action of wall-modifying enzymes. J. Exp. Bot. 2016, 67, 463–476. [Google Scholar] [CrossRef]
- Madni, A.M.; Erwin, D.; Sievers, M. Constructing models for systems resilience: Challenges, concepts and formal methods. Systems 2020, 8, 3. [Google Scholar] [CrossRef]
- Baburin, S.; Zyrin, V.; Kovalchuk, M. Dependence of power supply systems reliability on the type of redundancy. In Proceedings of the IOP Conference Series: Materials Science and Engineering, Saint Petersburg, Russia, 23–24 May 2019; IOP Publishing: Bristol, UK, 2019; Volume 643, p. 012134. [Google Scholar]
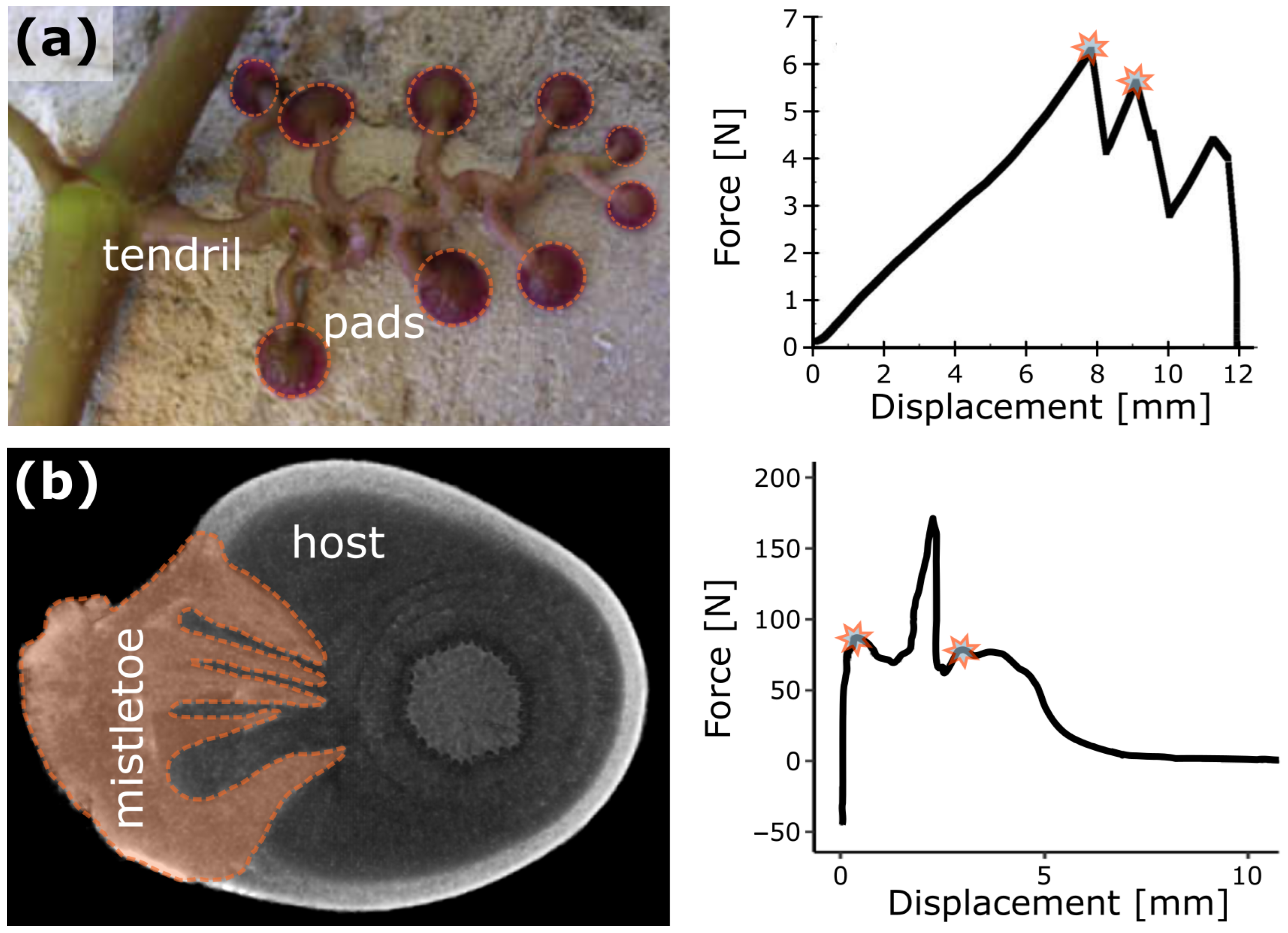
- Steinbrecher, T.; Danninger, E.; Harder, D.; Speck, T.; Kraft, O.; Schwaiger, R. Quantifying the attachment strength of climbing plants: A new approach. Acta Biomater. 2010, 6, 1497–1504. [Google Scholar] [CrossRef]
- Mylo, M.D.; Hofmann, M.; Delp, A.; Scholz, R.; Walther, F.; Speck, T.; Speck, O. Advances on the visualization of the internal structures of the European mistletoe: 3D reconstruction using microtomography. Front. Plant Sci. 2021, 12, 715711. [Google Scholar] [CrossRef] [PubMed]
- Mylo, M.D.; Hofmann, M.; Balle, F.; Beisel, S.; Speck, T.; Speck, O. Biomechanics of the parasite–host interaction of the European mistletoe. J. Exp. Bot. 2022, 73, 1204–1221. [Google Scholar] [CrossRef] [PubMed]
- Coit, D.W. Cold-standby redundancy optimization for nonrepairable systems. IIE Trans. 2001, 33, 471–478. [Google Scholar] [CrossRef]
- Teixeira-Costa, L. A living bridge between two enemies: Haustorium structure and evolution across parasitic flowering plants. Braz. J. Bot. 2021, 44, 165–178. [Google Scholar] [CrossRef]
- Westwood, J.H.; Yoder, J.I.; Timko, M.P.; Depamphilis, C.W. The evolution of parasitism in plants. Trends Plant Sci. 2010, 15, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Heide-Jørgensen, H.S. Introduction: The parasitic syndrome in higher plants. In Parasitic Orobanchaceae; Springer: Cham, Switzerland, 2013; pp. 1–18. [Google Scholar] [CrossRef]
- Teixeira-Costa, L.; Davis, C.C. Life history, diversity and distribution in parasitic flowering plants. Plant Physiol. 2021, 187, 32–51. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, X.; Chen, Y.; Wainwright, D.K.; Kenaley, C.P.; Gong, Z.; Liu, Z.; Liu, H.; Guan, J.; Wang, T.; et al. A biorobotic adhesive disc for underwater hitchhiking inspired by the remora suckerfish. Sci. Robot. 2017, 2, eaan8072. [Google Scholar] [CrossRef]
- Li, L.; Wang, S.; Zhang, Y.; Song, S.; Wang, C.; Tan, S.; Zhao, W.; Wang, G.; Sun, W.; Yang, F.; et al. Aerial-aquatic robots capable of crossing the air-water boundary and hitchhiking on surfaces. Sci. Robot. 2022, 7, eabm6695. [Google Scholar] [CrossRef]
- Weihs, D.; Fish, F.E.; Nicastro, A.J. Mechanics of remora removal by dolphin spinning. Mar. Mammal Sci. 2007, 23, 707–714. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Kwon, C.T.; Tang, L.; Wang, X.; Gentile, I.; Hendelman, A.; Robitaille, G.; Van Eck, J.; Xu, C.; Lippman, Z.B. Dynamic evolution of small signalling peptide compensation in plant stem cell control. Nat. Plants 2022, 8, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Pillar, V.D.; Blanco, C.C.; Müller, S.C.; Sosinski, E.E.; Joner, F.; Duarte, L.D.S. Functional redundancy and stability in plant communities. J. Veg. Sci. 2013, 24, 963–974. [Google Scholar] [CrossRef]
- Malinskii, Y.M.; Prokopenko, V.; Ivanova, N.; Kargin, V. Investigation of self-healing of cracks in polymers. Mech. Compos. Mater. 1970, 6, 382–384. [Google Scholar] [CrossRef]
- White, S.R.; Sottos, N.R.; Geubelle, P.H.; Moore, J.S.; Kessler, M.R.; Sriram, S.; Brown, E.N.; Viswanathan, S. Autonomic healing of polymer composites. Nature 2001, 409, 794–797. [Google Scholar] [CrossRef] [PubMed]
- Busch, S.; Seidel, R.; Speck, O.; Speck, T. Morphological aspects of self-repair of lesions caused by internal growth stresses in stems of Aristolochia macrophylla and Aristolochia ringens. Proc. R. Soc. B Biol. Sci. 2010, 277, 2113–2120. [Google Scholar] [CrossRef]
- Paul-Victor, C.; Dalle Vacche, S.; Sordo, F.; Fink, S.; Speck, T.; Michaud, V.; Speck, O. Effect of mechanical damage and wound healing on the viscoelastic properties of stems of flax cultivars (Linum usitatissimum L. cv. Eden and cv. Drakkar). PLoS ONE 2017, 12, e0185958. [Google Scholar] [CrossRef]
- Evert, R.F. Esau’s Plant Anatomy: Meristems, Cells and Tissues of the Plant Body: Their Structure, Function and Development; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Speck, O.; Speck, D.; Horn, R.; Gantner, J.; Sedlbauer, K.P. Biomimetic bio-inspired biomorph sustainable? An attempt to classify and clarify biology-derived technical developments. Bioinspir. Biomimetics 2017, 12, 011004. [Google Scholar] [CrossRef]
- Pang, J.W.; Bond, I.P. A hollow fibre reinforced polymer composite encompassing self-healing and enhanced damage visibility. Compos. Sci. Technol. 2005, 65, 1791–1799. [Google Scholar] [CrossRef]
- Pang, J.; Bond, I. ‘Bleeding composites’—Damage detection and self-repair using a biomimetic approach. Compos. Part A Appl. Sci. Manuf. 2005, 36, 183–188. [Google Scholar] [CrossRef]
- Trask, R.; Bond, I. Biomimetic self-healing of advanced composite structures using hollow glass fibres. Smart Mater. Struct. 2006, 15, 704. [Google Scholar] [CrossRef]
- Norris, C.J.; Meadway, G.J.; O’Sullivan, M.J.; Bond, I.P.; Trask, R.S. Self-healing fibre reinforced composites via a bioinspired vasculature. Adv. Funct. Mater. 2011, 21, 3624–3633. [Google Scholar] [CrossRef]
- Luterbacher, R.; Coope, T.; Trask, R.; Bond, I. Vascular self-healing within carbon fibre reinforced polymer stringer run-out configurations. Compos. Sci. Technol. 2016, 136, 67–75. [Google Scholar] [CrossRef]
- Sangadji, S.; Schlangen, E. Mimicking bone healing process to self repair concrete structure novel approach using porous network concrete. Procedia Eng. 2013, 54, 315–326. [Google Scholar] [CrossRef]
- Garcia, S.J. Effect of polymer architecture on the intrinsic self-healing character of polymers. Eur. Polym. J. 2014, 53, 118–125. [Google Scholar] [CrossRef]
- Martín H, J.A.; de Lope, J.; Maravall, D. Adaptation, anticipation and rationality in natural and artificial systems: Computational paradigms mimicking nature. Nat. Comput. 2009, 8, 757–775. [Google Scholar] [CrossRef]
- Altenbach, H.; Kolupaev, V.A. Classical and non-classical failure criteria. In Failure and Damage Analysis of Advanced Materials; Springer: Cham, Switzerland, 2015; pp. 1–66. [Google Scholar]
- Worden, K.; Dulieu-Barton, J.M. An overview of intelligent fault detection in systems and structures. Struct. Health Monit. 2004, 3, 85–98. [Google Scholar] [CrossRef]
- Walsh, D.M. Fitness and function. Br. J. Philos. Sci. 1996, 47, 553–574. [Google Scholar] [CrossRef]
- Achinstein, P. Function statements. Philos. Sci. 1977, 44, 341–367. [Google Scholar] [CrossRef]
- Merriam-Webster Online Dictionary. Definition of Functionality. 2022. Available online: https://www.merriam-webster.com/dictionary/functionality (accessed on 7 December 2022).
- Collins Dictionary. Definition of Resilience. 2022. Available online: https://www.collinsdictionary.com/de/worterbuch/englisch/resilience (accessed on 7 December 2022).
- Merriam-Webster Online Dictionary. Definition of Resistance. 2022. Available online: https://www.merriam-webster.com/dictionary/resistance (accessed on 7 January 2022).
- Khammash, M. An engineering viewpoint on biological robustness. BMC Biol. 2016, 14, 22. [Google Scholar] [CrossRef]
- Harrington, M.J.; Speck, O.; Speck, T.; Wagner, S.; Weinkamer, R. Biological archetypes for self-healing materials. In Self-Healing Materials; Springer: Cham, Switzerland, 2015; pp. 307–344. [Google Scholar] [CrossRef]
- Woo, H.R.; Masclaux-Daubresse, C.; Lim, P.O. Plant senescence: How plants know when and how to die. J. Exp. Bot. 2018, 69, 715–718. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BOTANY | Response | Acclimation | Adaptation |
|---|---|---|---|
| Subject | Individual plants | Individual plants | Populations of plants |
| Effect | Reconfiguration of plant organs | Change of gene expression | Change of genetic information |
| Time span | Seconds to minutes | Days to months | Evolutionary time |
| Result | “Re-oriented” plants | “Trained” plants | “Adapted” plants |
| Plant example | Reversible streamlining of plants and plant organs under wind loads [82,83,84] | Non-hereditary alteration of growth pattern (e.g., reduction in shoot elongation) [81,85,86,87,88,89] | Variation of hereditary traits (e.g., trampling tolerant plants with low growth and rosette formation) [90] |
| TECHNOLOGY | Response | Adaptivity | Optimisation |
|---|---|---|---|
| Subject | Individual components | Individual components | Populations of designs |
| Effect | Reconfiguration of component parts | Change of material properties | Optimisation of design parameters |
| Time span | Seconds to days | Number of cycles | Number of generations |
| Result | “Re-oriented” components | “Trained” components | “Optimised” design |
| Biomimetic example | Stimulus-responsive buildings (e.g., Urbach Tower) and facade-shading systems (e.g., Flectofin, Flectofold, Hygroskin) [91,92,93,94,95,96] | Persistent change of material properties through cyclic loading [97,98] | Optimised parameters through evolutionary algorithms (e.g., coffee blend, winglets for air planes, lightweight constructions) [99,100,101,102,103,104] |
| Evolution (Biology) | Evolutionary Strategy (Technology) | |
|---|---|---|
| Subject | Living being | Object to be optimised |
| Mutation | Random change of genetic information | Random change of input variables (= object parameters) |
| Recombination | Reshuffling of parental genetic material (e.g., meiosis) | New combination of parental object parameters |
| Selection | Selection of those individuals with the best fit to the natural environment | Selection of those individuals that best meet the optimisation criterion |
| Result | Adapted organism | Optimised object |
| Questions | Answers |
|---|---|
| What is being restored? | Geometric properties, mechanical properties, structural integrity, functionality of the entire system |
| What size of damage can be self-repaired? | Length, width and depth or radius (e.g., millimetres, centimetres) |
| How quickly should self-repair be carried out? | Seconds, days, weeks, month, years |
| How often can the damage self-repair? | Once, twice, several times |
| Should self-repair be initiated autonomously or by a specific trigger? | Autonomous self-repair without a trigger or non-autonomous self-repair initiated by a specific trigger (e.g., temperature, light, humidity, mechanical compression) |
| How can the structural integrity of the damaged and repaired status be measured? | Qualitative assessment by imaging techniques (e.g., light microscopy, SEM, X-ray tomography) |
| How can the mechanical integrity of the damaged and repaired status be measured? | Quantitative analyses by various equations [34,35,36,37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mylo, M.D.; Speck, O. Longevity of System Functions in Biology and Biomimetics: A Matter of Robustness and Resilience. Biomimetics 2023, 8, 173. https://doi.org/10.3390/biomimetics8020173
Mylo MD, Speck O. Longevity of System Functions in Biology and Biomimetics: A Matter of Robustness and Resilience. Biomimetics. 2023; 8(2):173. https://doi.org/10.3390/biomimetics8020173
Chicago/Turabian StyleMylo, Max D., and Olga Speck. 2023. "Longevity of System Functions in Biology and Biomimetics: A Matter of Robustness and Resilience" Biomimetics 8, no. 2: 173. https://doi.org/10.3390/biomimetics8020173
APA StyleMylo, M. D., & Speck, O. (2023). Longevity of System Functions in Biology and Biomimetics: A Matter of Robustness and Resilience. Biomimetics, 8(2), 173. https://doi.org/10.3390/biomimetics8020173