1. Introduction
Bio-brain exhibits remarkable information processing ability [
1]. Neural information coding is crucial for facilitating this processing. Various responses are formed in response to various stimuli, which is essential for the brain to differentiate between various stimuli to achieve advanced brain functions [
2,
3]. The performance of SNC is extremely dependent on brain-inspired models. However, the bio-rationality of a brain-inspired model remains inadequate as they do not fully capture the structure and function of real neural systems. This research gap limits the ability of current models to replicate the complexities of biological neural coding mechanisms. Spiking neural network (SNN), inspired by the brain [
4,
5], can drive advancements in artificial intelligence through research on their specific neural coding. An SNN architecture consists of a neuron model, a synaptic plasticity model, and a network topology.
Bio-neurons act as units for processing neural information within the brain, while neuron models are mathematical representations that replicate electrophysiological characteristics of bio-neurons. Many types of neuron models have been developed by researchers, including the Hodgkin–Huxley (H-H) neuron model, the leaky integrate-and-fire (LIF) neuron model, the Izhikevich neuron model, and so on. Since the H-H neuron model incurs significant computational expense [
6], the LIF neuron model reduces computational expense but fails to accurately replicate the firing behavior observed in bio-neurons [
7]. The Izhikevich neuron model can reflect the firing activity of bio-neurons well while maintaining a reduced computing expense [
8]. Therefore, it is extensively utilized in the construction of SNNs [
9,
10].
Bio-synapses serve as the fundamental components for transmitting neural information among bio-neurons [
11]. Excitatory synapses (ESs) have the capacity to enhance the effectiveness of neural information transfer [
12], whereas inhibitory synapses (ISs) have the capacity to decrease the speed and sensitivity of neural information transfer [
13]. Biological studies have shown that ISs, in conjunction with ESs, play a crucial role in the regulation of neural activity in the bio-brain. For instance, Xue et al. [
14] found that the proportion of ISs rose alongside heightened neuronal excitability by measuring how neurons of young mice responded to optical stimuli, which reflects excitation–inhibition balance in anatomy. Inspired by bio-synapses, synaptic plasticity models co-regulated by ESs and ISs are utilized to construct SNNs. Zhao et al. [
15] constructed an SNN with both ES and IS models. They discovered that this SNN outperforms an SNN without an inhibitory synapse model on MNIST datasets. For bio-synapses, the neurotransmitter dispersion induces a synaptic time delay (STD), which is stochastically distributed within the interval [0.1, 40]
[
16]. Hence, synaptic plasticity models with STD have been introduced in SNNs. Zhang et al. [
17] developed an SNN with STD, which demonstrated the ability to accurately reproduce the expected spike sequence and achieved better accuracy than an SNN without an STD on the TIDIGITS speech recognition task. However, in this study, the STD was set to a fixed value that did not conform to the bio-STD interval [0.1, 40]
. Hence, on the basis of co-regulation of ESs and ISs, a synaptic plasticity model with STD conforming to the bio-STD can improve the performance of SNNs.
The topology decides the forms of connection between neurons in a brain network. According to principles of complex network theory, networks are classified according to their topological structure into regular, random, and complex networks. Among these, complex networks encompass small-world (SW) networks and scale-free (SF) networks [
18]. The SW networks exhibit both a high mean clustering coefficient (CC) and a low mean shortest path length (SPL) [
19]. The SF network exhibits the degree distribution of nodes adheres to a power-law distribution, characterized by significant heterogeneity, which endows it with robust fault tolerance [
20]. Biological research has shown that bio-functional brain networks (FBN) are complex networks with SW and/or SF properties. Van et al. [
21] generated FBNs of 28 healthy subjects, finding that these networks exhibited SW and SF properties. Liu et al. [
22] examined the SW property of FBNs in 31 schizophrenia patients and 31 healthy individuals, revealing that SW property was absent in schizophrenia patients. Stylianou et al. [
23] generated FBNs for 15 Parkinson’s disease patients to investigate the effect of treatment of dopaminergic on SF property and found that SF property was normal by the treatment. Based on results for FBNs, researchers have constructed SNNs with the topology of SW and/or SF properties. Tsakalos et al. [
24] developed a small-world spiking neural network (SWSNN) by utilizing Watts and Strogatz (WS) algorithm for topology generation. They discovered that this SWSNN achieved superior recognition accuracy compared to a two-layered SNN on the image dataset. Reis et al. [
25] constructed an SFSNN utilizing Barabási–Albert (BA) algorithm and investigated synchronization. They discovered that the SFSNN could inhibit burst synchronization under the exterior disturbance of light pulses while still maintaining low synchronization after the disturbance had stopped for an extended period. In a previous study [
26], we constructed SWSNN and SFSNN to explore anti-interference capacity under pulse noise. Our findings revealed that SWSNN was more robust than SFSNN, which indicates that anti-interference capacity of these two SNNs is affected by topology. Based on the topological characteristics of bio-FBN, the complex spiking neural network (CSNN) exhibiting SW property and SF property can enhance the bio-rationality of brain-inspired models.
The bio-brain responds to exterior stimuli through neural coding [
27]. Callier et al. [
28] investigated the reaction of populations of neurons in monkey’s brain when exposed to skin pressure marks. Their study revealed that these neurons can encode the timing, location, and magnitude of skin indentation quickly and consistently, indicating that population coding can effectively encode features of contact events. Inspired by these biological findings, researchers have examined the coding of brain-inspired models. Zhu et al. [
29] examined energy consumption of an SNN. They discovered that the distribution of network energy is positively associated with the strength of coupling between neurons, implying that energy coding is a useful tool for assessing cost-effectiveness of the brain-inspired model. Du et al. [
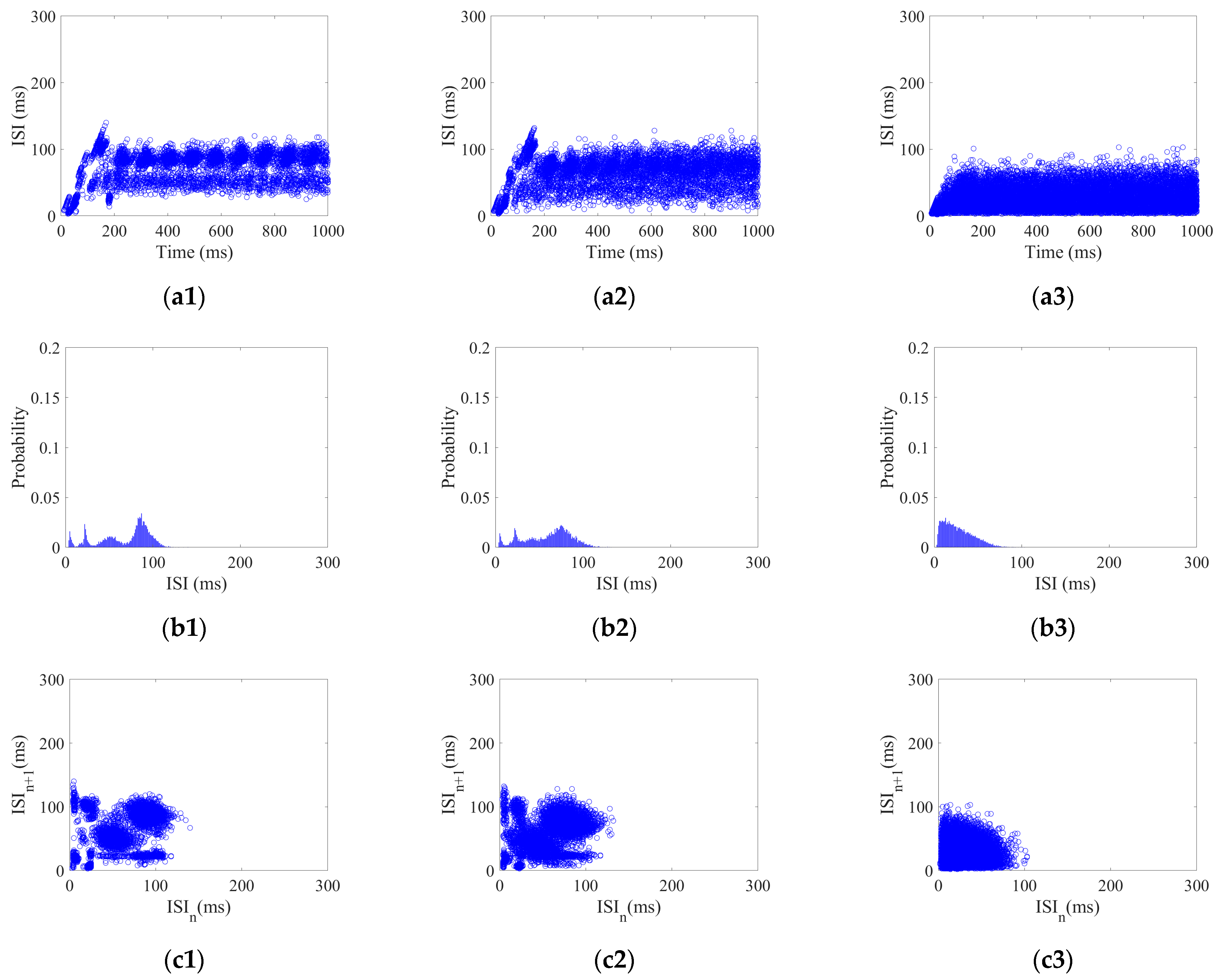
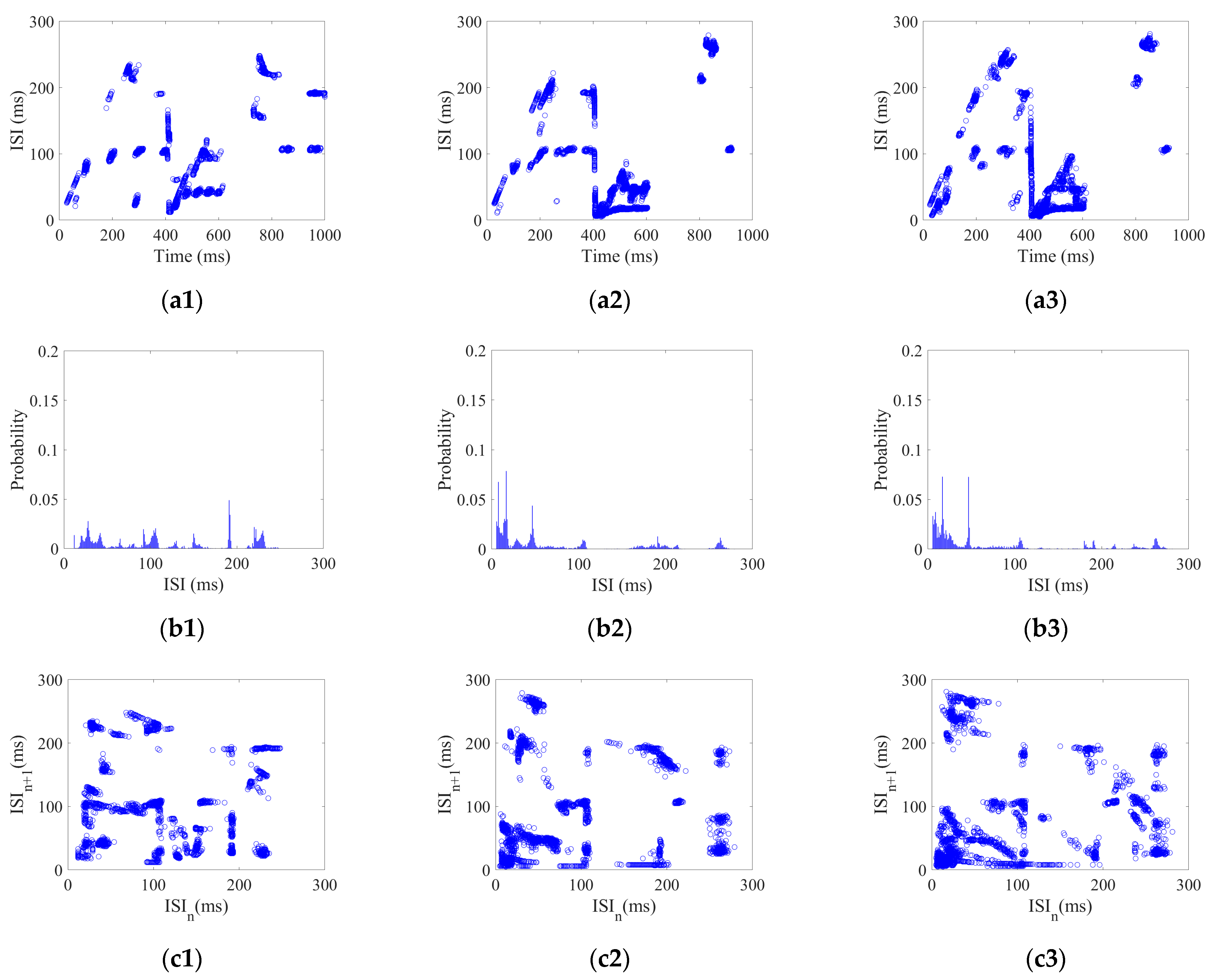
30] studied the firing activity of an SNN under a sinusoidal induced electric field (IEF) using the inter-spike intervals (ISI). They found that the ISI gradually became an integer multiple of the firing period over time, forming a stable neural coding for IEF with random phase noise. Biological results have demonstrated that the bio-brain can form specific patterns of neural coding in response to various forms of exterior stimulus. For example, Stephanie et al. [
31] observed neural activity in the brains of mice in response to different forms of taste stimulus and found that there was obvious specificity in the population coding in the gustatory cortex, indicating that different taste stimuli allow mice to produce different coding patterns. Therefore, the SNC can generate distinctive coding patterns that correspond to specific exterior stimuli. The performance of SNC is extremely dependent on brain-inspired models. However, the bio-rationality of the topology in current brain-inspired models remains deficient. This gap hinders the potential of neural coding in applications.
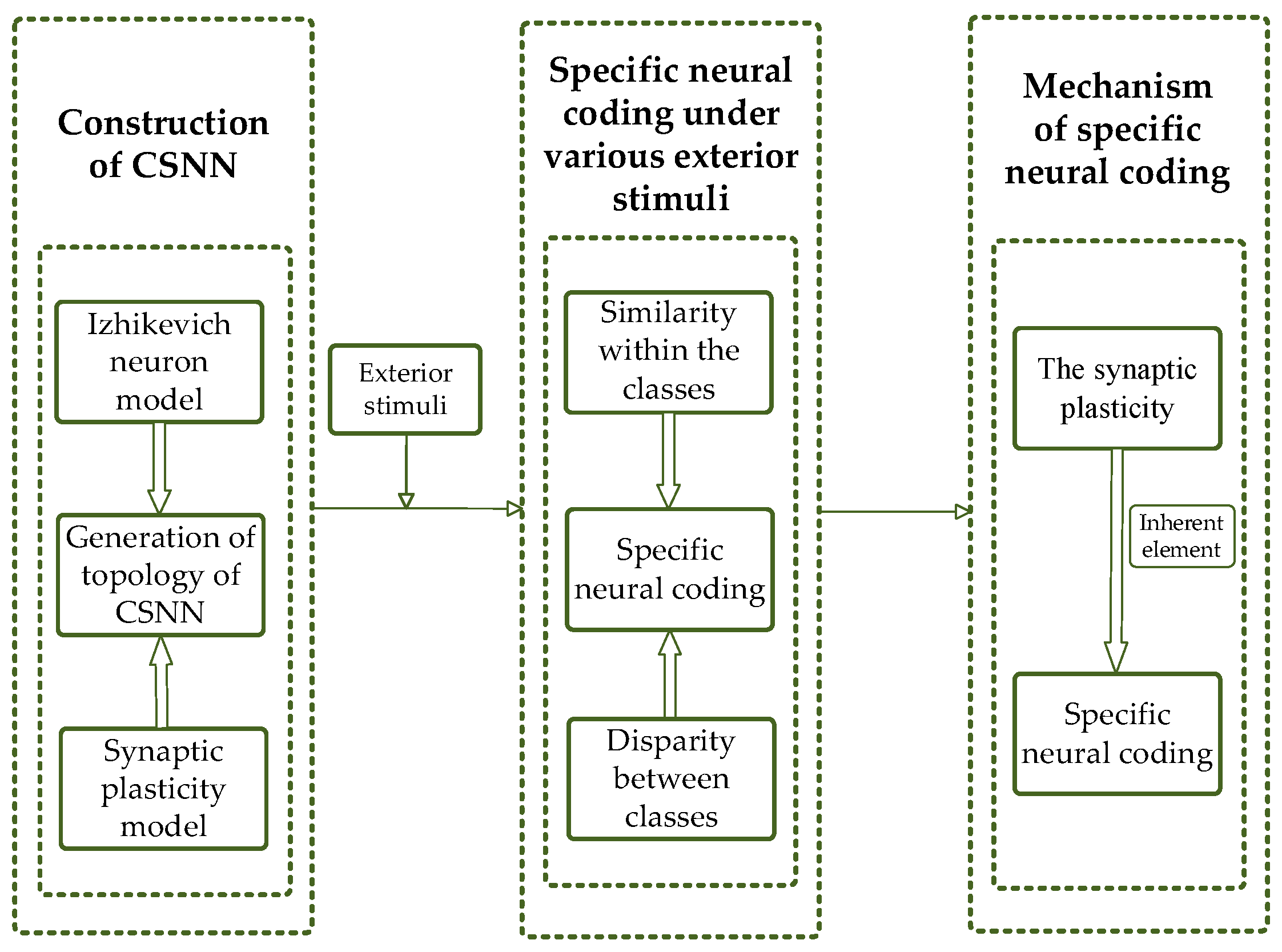
In order to deal with the above challenges, we propose a brain-inspired model with bio-rationality and investigate the SNC of CSNN under various strengths of various stimuli. In this study, we construct a CSNN with a topology that exhibits SW and SF properties, where the nodes are represented by Izhikevich neuron models, and the edges consist of synaptic plasticity models incorporating bio-STD, co-regulated by ESs and ISs. This design enhances the bio-rationality of the model, making it more aligned with the bio-brain and its information-processing capabilities. To investigate the SNC of CSNN under various strengths of different stimuli, K-means clustering algorithm and cosine similarity algorithms are employed. K-means clustering algorithm is well-suited for this task, efficiently grouping coding patterns into clusters and enabling the analysis of coding differences across stimuli, which helps to identify different responses across varying stimuli and allows for a clear analysis of coding differences. Meanwhile, the cosine similarity algorithm quantifies the similarity between coding patterns, and thus enabling a deeper understanding of the similarity within the classes. Since neural coding primarily relies on time coding patterns rather than absolute spike counts, cosine similarity provides an effective measure for evaluating the consistency of SNC under different stimulus. These two strategies complement each other and offer a way to analyze the time neural coding in CSNNs. Furthermore, we discuss the mechanism of the SNC by analyzing the correlation between the SNC and the synaptic plasticity.
The main contributions of this paper are as follows:
To enhance the bio-rationality of brain-inspired models, a CSNN is proposed with a topology that incorporates both SW and SF properties. The nodes are Izhikevich neuron models, and the edges are represented by synaptic plasticity models with a bio-STD co-regulated by ESs and ISs.
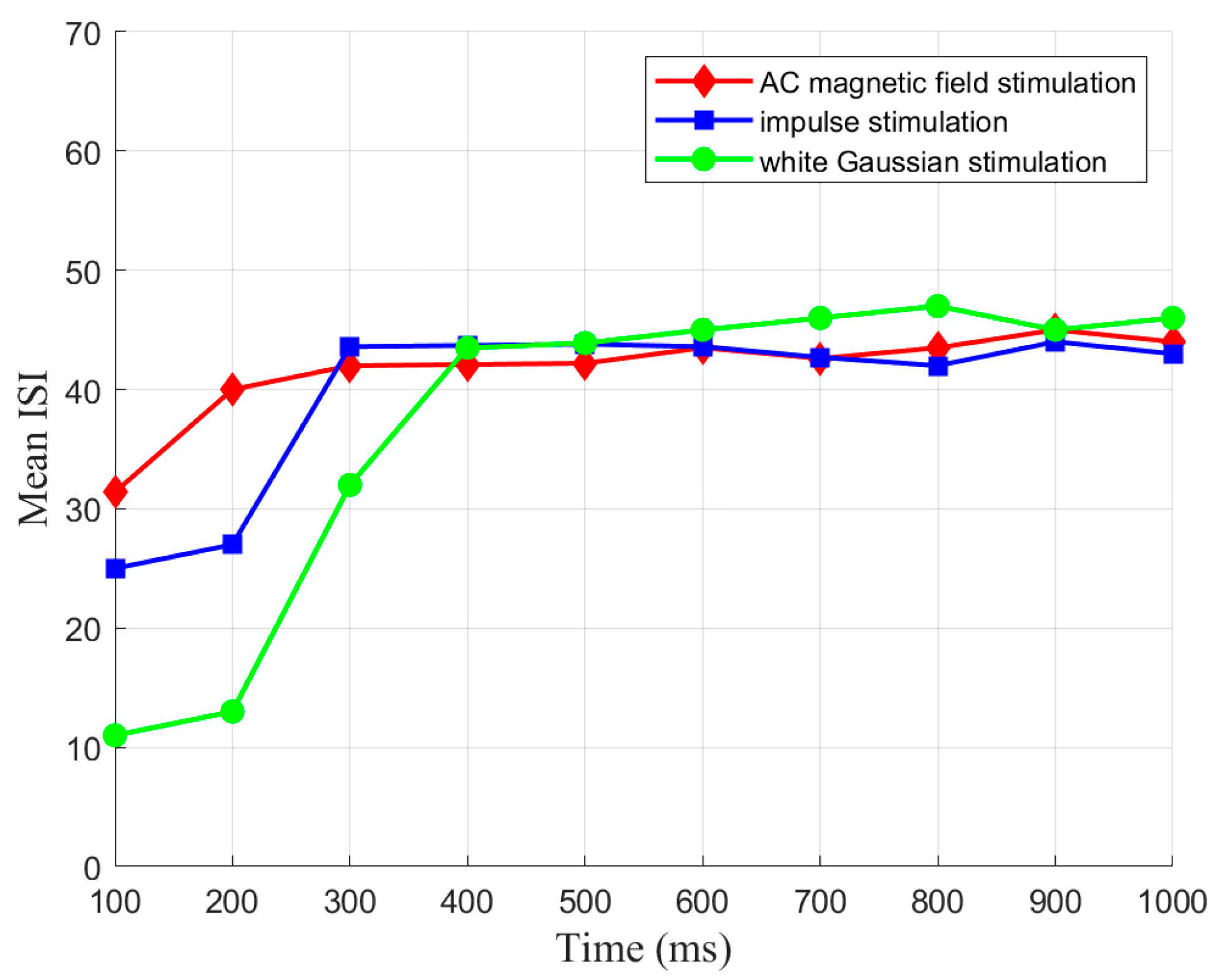
To investigate the SNC of CSNN under various strengths of various stimuli, the K-means clustering algorithm and cosine similarity algorithm are used. Our results indicate that CSNN exhibits marked time coding similarity under various strengths of same stimulus; the CSNN exhibits marked SNC under various stimuli.
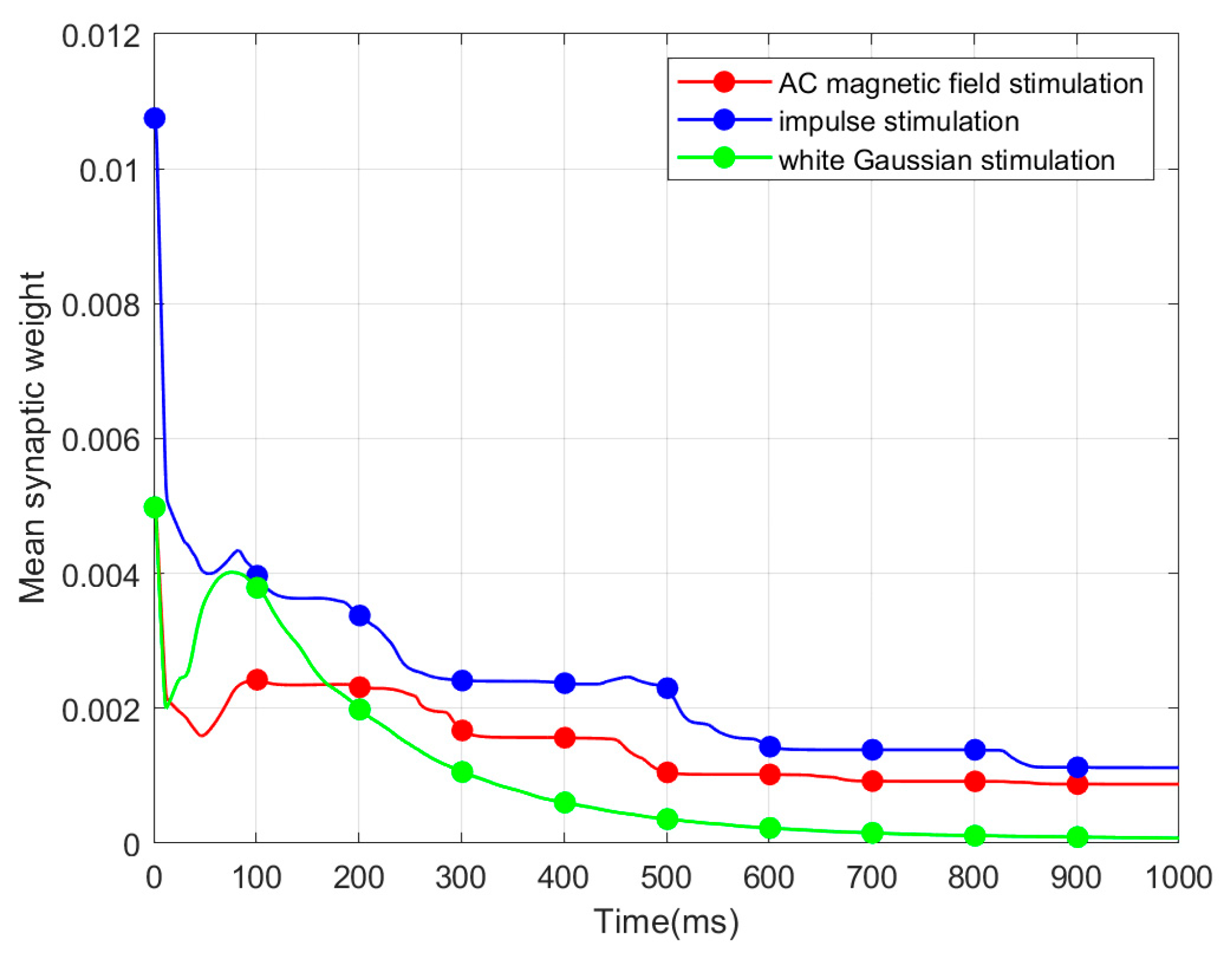
To elucidate mechanism of the SNC based on CSNN, we conduct a discussion between synaptic weight and SNC, which indicates that the inherent factor of the SNC is synaptic plasticity.
The following sections are organized as follows: The method of constructing the CSNN is described in
Section 2. The SNC of CSNN under various stimuli is proposed in
Section 3. Mechanism of the SNC is discussed in
Section 4. Finally, a conclusion is presented in
Section 5. Our flowchart is shown in
Figure 1.
5. Conclusions
This study proposed CSNN, a brain-inspired model characterized by bio-rationality. Its topology combines the SW property and SF property. To investigate the SNC of CSNN under various strengths of various stimuli, we used a K-means clustering algorithm and a cosine similarity algorithm. To elucidate its mechanism, we discuss the relationship between SNC and synaptic plasticity.
The following main conclusions are drawn from this work: (1) The CSNN exhibits marked time coding similarity under various strengths of the same stimulus; the CSNN exhibits marked SNC under various stimuli. (2) Our discussion indicates that the inherent factor of the SNC is synaptic plasticity. Investigating SNC and its underlying mechanisms can enhance our comprehension of intricate brain cognitive functions and the processes involved in information handling, while also fostering advancements in artificial intelligence.
For our future work, we aim to construct a more bio-rationality brain-inspired model, and will apply this brain-inspired model based on SNC to other pattern recognition tasks such as image recognition and speech recognition. Second, investigating how CSNN respond to time-varying exterior stimuli remains an essential avenue for future work, which could provide insights into their robustness and efficiency in processing real-world signals. Finally, while the present study identifies a significant correlation between MSW and ISI, future work should focus on uncovering the causal mechanisms underlying this relationship.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}