How Mitochondrial DNA Can Write Pre-History: Kinship and Culture in Duero Basin (Spain) during Chalcolithic and Bronze Age

 ,
, 
 ,
,
Abstract
1. Introduction
2. Materials and Methods
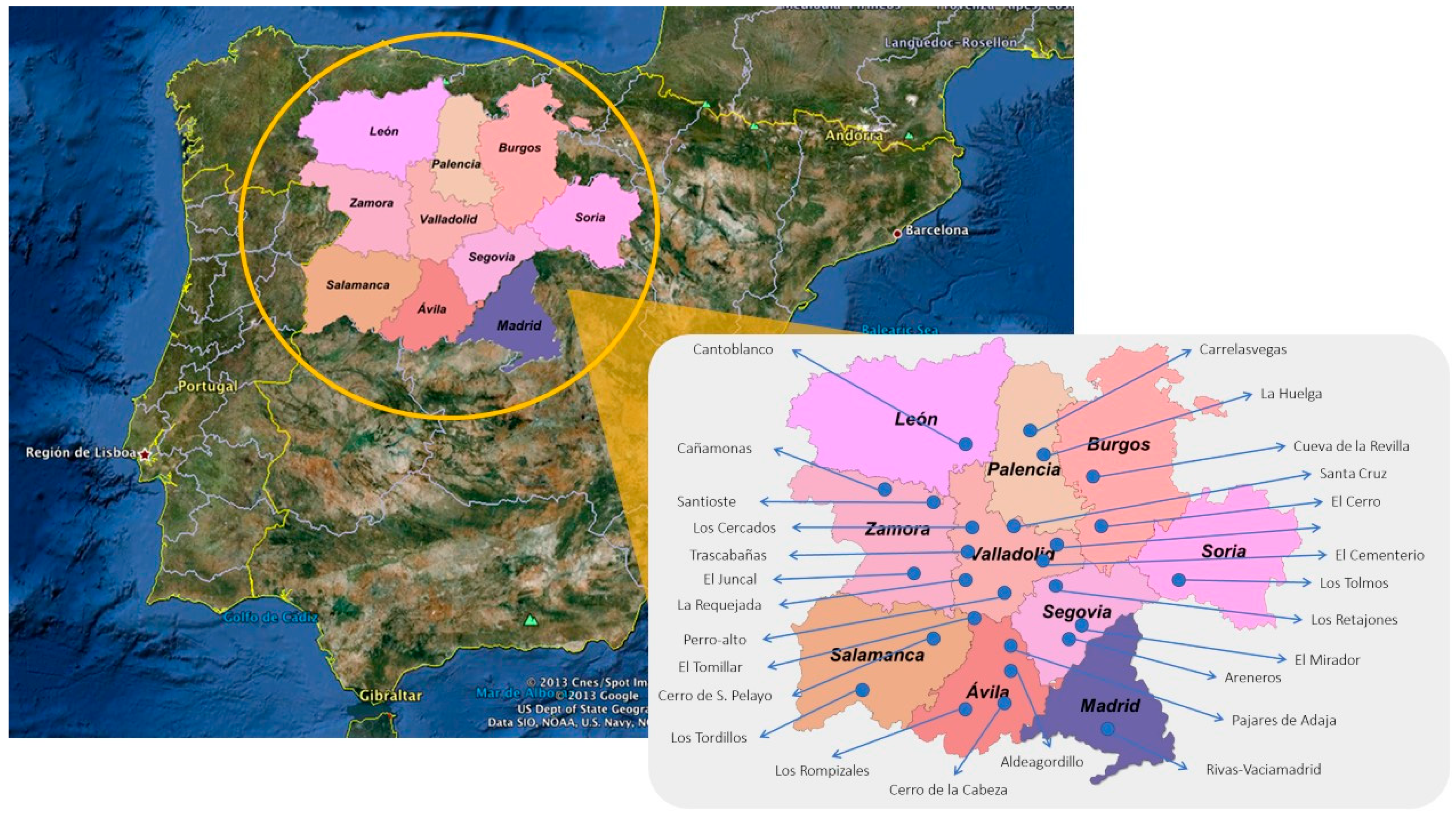
2.1. Materials: Selected Samples from Each Chronological Period Studied
2.2. Methodology
2.2.1. Experimental Process and Authenticity Criteria
- Pretreatment of samples and ancient DNA extraction in Laboratory 1:
- Samples pretreatment, ancient DNA extraction, and DNA quantification in Laboratory 2:
- DNA amplification by PCR, results analysis, and cloning.
2.2.2. Analytical Procedure
- Data organisation
- Analysis of Molecular Variance (AMOVA) and FST distances
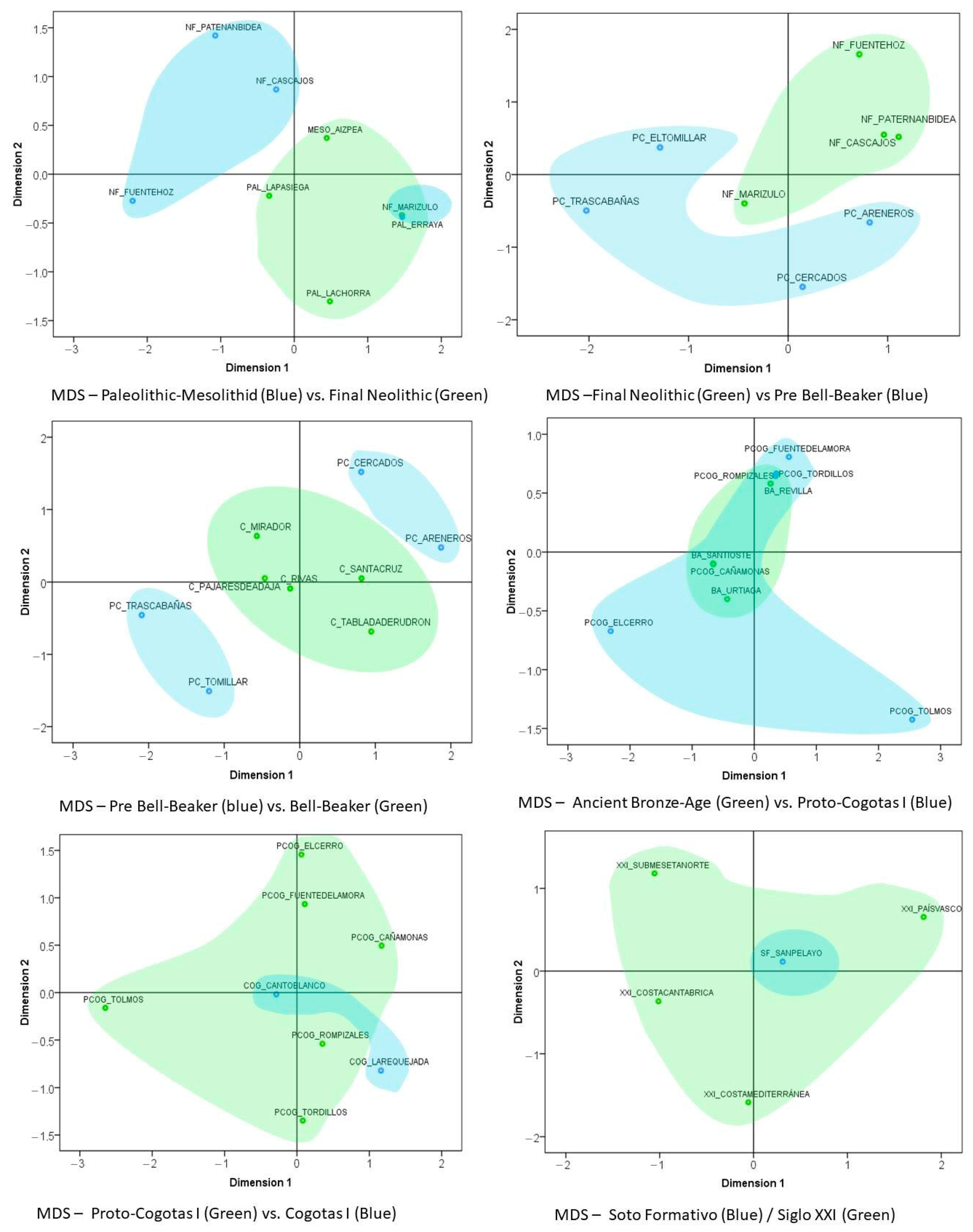
- Multidimensional Scaling Analysis (MDS)
- Correspondence Analysis: Performed by SPSS Statistics 24 software.
- Creation of Maps of Genetic Distances and Haplogroup Frequencies
3. Results
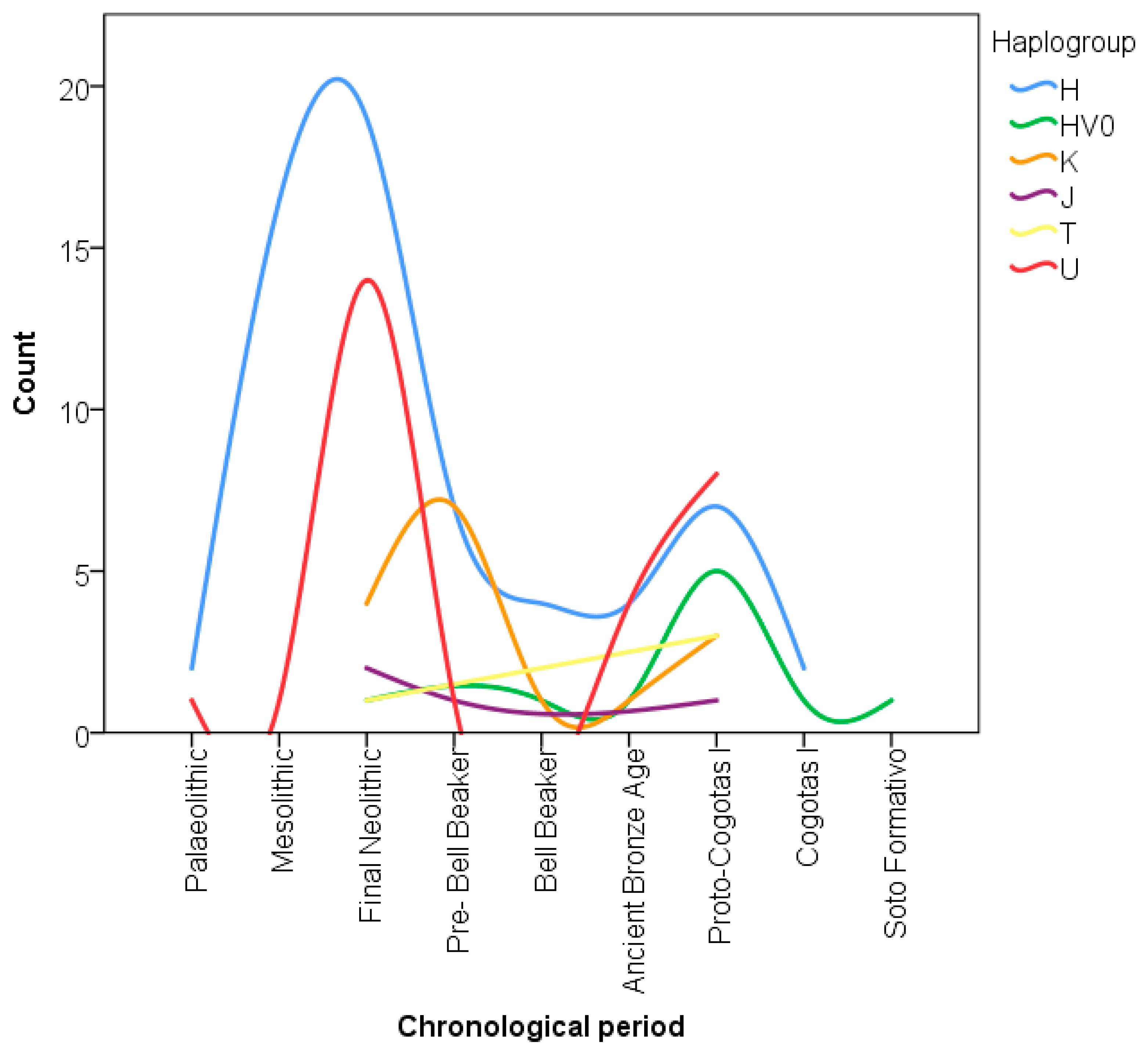
3.1. mtDNA Haplotypes and Haplogroups Determined
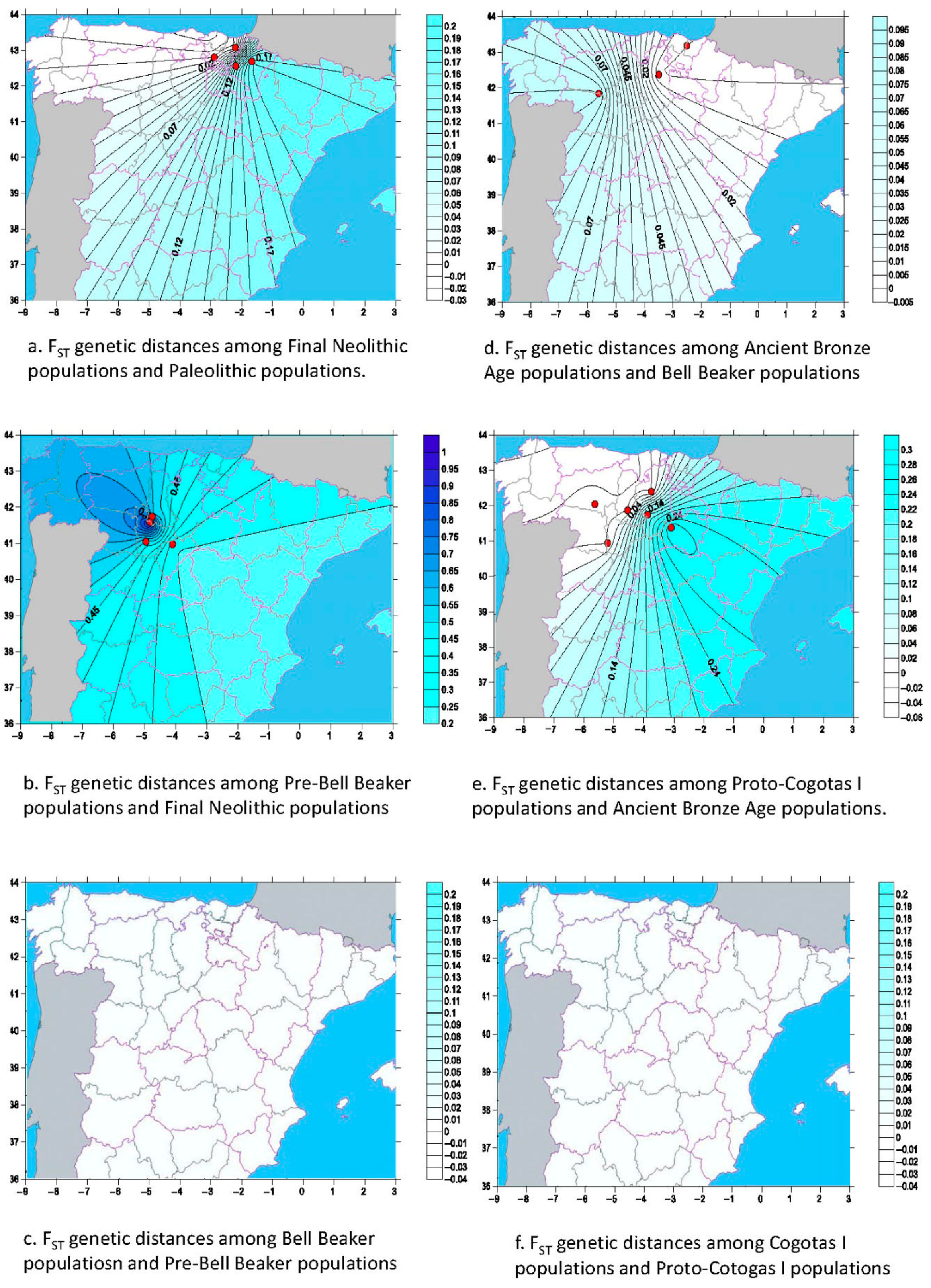
3.2. Analysis of Variations among the Chronological Groups: AMOVA and Genetic Distances FST
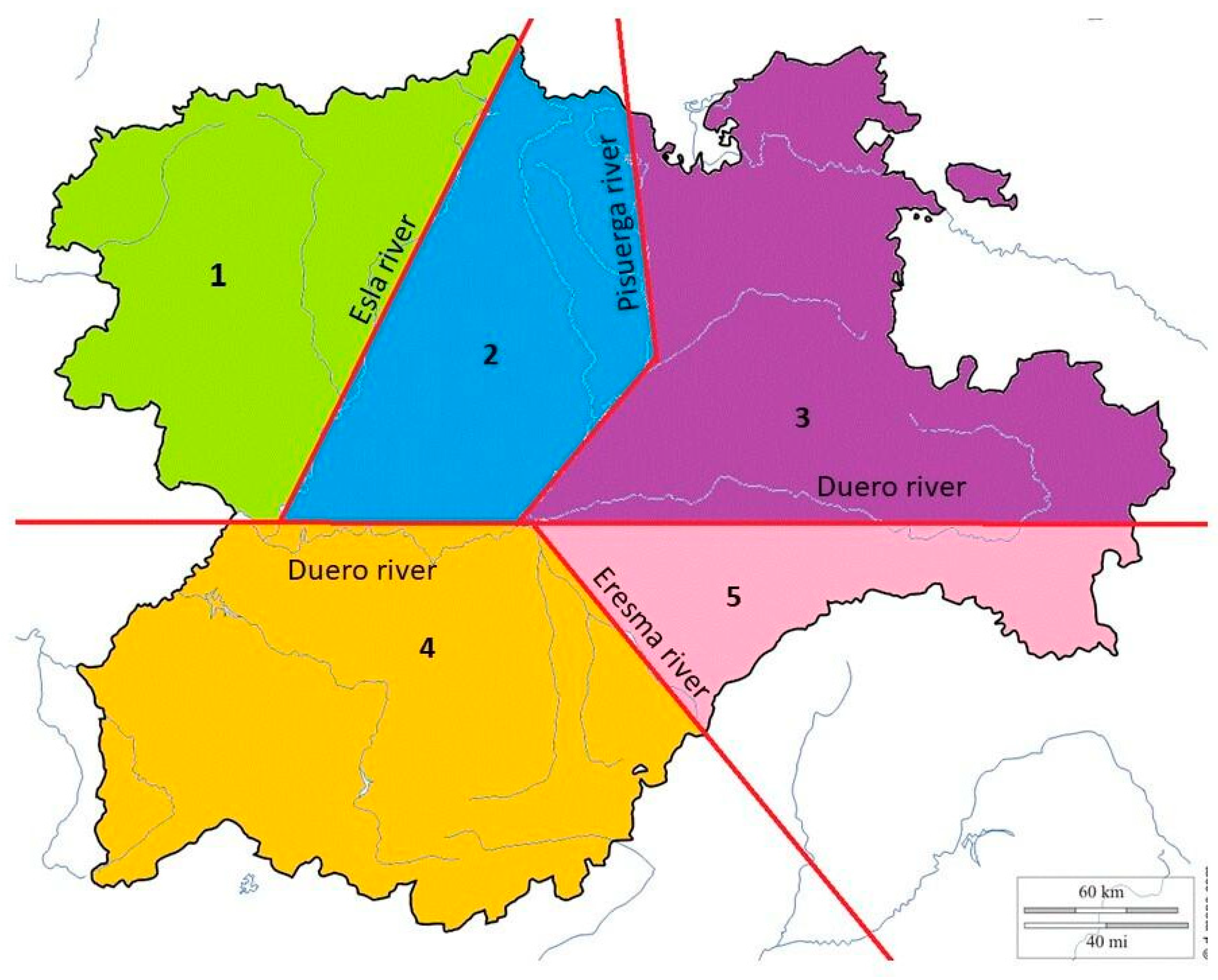
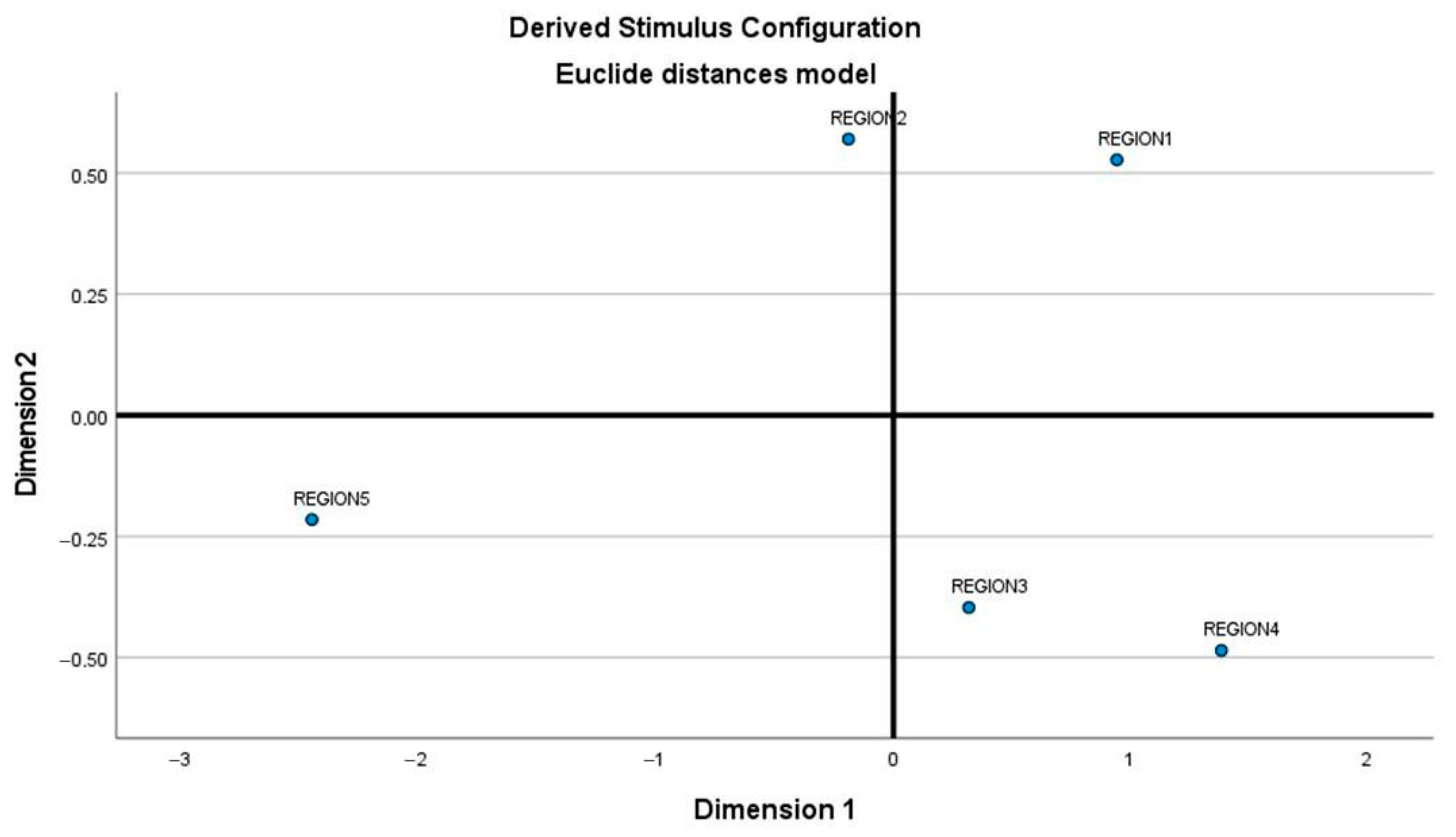
3.3. Analysis of Variations among the Geographical Groups: AMOVA and Genetic Distances FST
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abarquero Moras, Francisco Javier. 2005. Cogotas I. La Difusión de un Tipo Cerámico Durante la Edad del Bronce. Monografías Arqueología en Castilla y León—4. Valladolid: Junta de Castilla y León and Consejería de Cultura y Turismo. ISBN 84-9718-293-6. [Google Scholar]
- Aliaga, Raquel Almela. 2008. El mundo funerario calcolítico de la Región de Madrid. Cuadrenos de Prehistoria y Arqueología 34: 23–39. [Google Scholar] [CrossRef][Green Version]
- Alves-Cardoso, Francisca, Sara Palomo-Díez, Alejandro Alonso Conde, Cláudia Gomes, Silvia Casimiro, Rodrigo Banha da Silva, and Eduardo Arroyo-Pardo. 2022. Metagenomic Research of Infectious Diseases in Archaeological Contexts: Evidence from the Hospital Real de Todos-os-Santos (Portugal). Applied Sciences 12: 6096. [Google Scholar] [CrossRef]
- Anderson, Sharon, Alan T. Bankier, Bart G. Barrell, Maarten H. L. de Bruijn, Alan R. Coulson, Jacques Drouin, Ian C. Eperon, Donald P. Nierlich, Bruce A. Roe, Frederick Sanger, and et al. 1981. Sequence and organization of the human mitochondrial gnome. Nature 290: 457–65. [Google Scholar] [CrossRef]
- Andrews, Richard M., Iwona Kubacka, Patrick F. Chinnery, Robert N. Lightowlers, Douglass M. Turnbull, and Neil Howell. 1999. Reanalysis and revisión of the Cambridge reference sequence for human mitochondrial DNA. Nature Genetics 23: 147. [Google Scholar] [CrossRef] [PubMed]
- Blanco González, Antonio, and Ángel Esparza Arroyo. 2019. Conectividad en la Edad del Bronce del occidente peninsular. Examinando la relación entre sitios y vías pecuarias mediante SIG. Trabajos de Prehistoria 76: 67–83. [Google Scholar] [CrossRef]
- Blasco-Bosqued, Concepción. 1997. La Edad del Bronce en el interior Peninsular. Una aproximación al II Milenio A.C. en las cuencas de los ríos Duero y Tajo. CuPAUAM 24: 59–100. [Google Scholar] [CrossRef][Green Version]
- Delibes de Castro, Germán. 1977. El Vaso Campaniforme en la Meseta Norte Española. Valladolid: Universidad de Valladolid. [Google Scholar]
- Delibes de Castro, Germán. 1987. Sobre los enterramientos del grupo campaniforme de Ciempozuelos: Diversidad y tradición. In El Origen de la Metalurgia en la Península Ibérica. II. Oviedo: Seminario de la Fundación Ortega y Gasset, pp. 37–51. [Google Scholar]
- Delibes de Castro, Germán, and Julio Fernández Manzano. 2000. La Trayectoria Cultural de la Prehistoria Reciente (6400–2500 BP) en la Submeseta Norte Principales Hitos de un Proceso. In 3º Congresso de Arqueología Peninsular. Coordinated by Vítor Oliveira Jorge. Pré-História Recente da Península Ibérica. Porto: ADECAP, vol. 4, pp. 95–122. ISBN 972-97613-5-3. [Google Scholar]
- Delibes de Castro, Germán, and Fernando Romero Carnicero. 2011. La plena colonización agraria del Valle Medio del Duero. Complutum 22: 49–94. [Google Scholar] [CrossRef]
- Esparza Arroyo, Ángel, Javier Velasco Vázquez, and Germán Delibes De Castro. 2012. Exposición de cadáveres en el yacimiento de Tordillos (Aldeaseca de la Frontera, Salamanca). Perspectiva bioarqueológica y posibles implicaciones para el estudio del ritual funerario de Cogotas I. Zephyrus 69: 95–128. [Google Scholar]
- Esparza Arroyo, Ángel, Javier Velasco Vázquez, and Celis Sánchez. 2016. Notas sobre la fase Soto Formativo en el poblado de Los Cuestos de La Estación (Benavente, Zamora). BSAA Arqueología LXXXII: 63–85. [Google Scholar]
- Esparza-Arroyo, Ángel, Sara Palomo-Díez, Javier Velasco-Vázquez, Germán Delibes, Eduardo Arroyo-Pardo, and Domingo C. Salazar-García. 2017. Familiar Kinship? Palaeogenetic and Isotopic Evidence from a Triple Burial of the Cogotas I Archaeological Culture (Bronze Age, Iberian Peninsula). Oxford Journal of Archaeology 36: 223–42. [Google Scholar] [CrossRef]
- Excoffier, Laurent, and Heidi El Lischer. 2010. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Molecular Ecology Resources 10: 564–67. [Google Scholar] [CrossRef]
- Excoffier, Laurent, Peter E. Smouse, and Joseph M. Quattro. 1992. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 131: 479–91. [Google Scholar] [CrossRef]
- Fernández, Eva. 2005. Polimorfismos de ADN Mitocondrial en Poblaciones Antiguas de la Cuenca Mediterránea. Ph.D. Thesis, Universidad de Barcelona, Barcelona, Spain. [Google Scholar]
- Fernández, Eva, Alejandro Pérez-Pérez, Cristina Gamba, Eva Prats, Pedro Cuesta, Josep Anfruns, Miquel Molist, Eduardo Arroyo-Pardo, and Daniel Turbón. 2014. Ancient DNA analysis of 8000 B.C. near eastern farmers supports an early neolithic pioneer maritime colonization of Mainland Europe through Cyprus and the Aegean Islands. PLoS Genetics 10: e1004401. [Google Scholar] [CrossRef]
- Fulton, Tara L. 2012. Setting up an ancient DNA laboratory. Methods in Molecular Biology 840: 1–11. [Google Scholar] [CrossRef]
- García Barrios, Ana S. 2007. Un enfoque de género en la Arqueología de la Prehistoria Reciente del VAlle Medio del Duero: Los cráneos femeninos calcolíticos de Los Cercados (Mucientes, Valladolid). In Nuevos Enfoques para la Enseñanza de la Historia: Mujer y Género Ante el Espacio Europeo de Educación Superior. Madrid: Asociación Cultural Al-Mudayna, pp. 41–56. [Google Scholar]
- Garrido-Pena, Rafael. 1999. El Campaniforme en la meseta: Análisis de su contexto social, económico y ritual. Ph.D. Thesis, Universidad Complutense de Madrid, Departamento de Prehistoria, Facultad de Geografía e Historia, Madrid, Spain. [Google Scholar]
- Garrido-Pena, Rafael. 2005. El fenómeno campaniforme: Un Siglo de debates sobre un enigma sin resolver. Prehistoria en 4: 1–16. [Google Scholar]
- Gomes, Cláudia, Sara Palomo-Díez, Jordi Foig, Ana María López-Parra, Carlos Baeza-Richer, Ángel Esparza-Arroyo, Juan Gibaja, and Eduardo Arroyo-Pardo. 2015. Nondestructive extraction DNA methods from bones or teeth, true or false? Forensic Science International: Genetics Supplement Series 5: e279–e282. [Google Scholar] [CrossRef]
- Gomes, Cláudia, Sara Palomo-Díez, Ana María López-Parra, and Eduardo Arroyo-Pardo. 2021. Genealogy: The Tree Where History Meets Genetics. Genealogy 5: 98. [Google Scholar] [CrossRef]
- Hagelberg, Erika, Bryan Sykes, and Robert Hedges. 1989. Ancient bone DNA amplified. Nature 342: 485. [Google Scholar] [CrossRef]
- Hervella, Montserrat, Neskuts Izagirre, Santos Alonso, Rosa Fregel, Antonio Alonso, M. Vicente Cabrera, and Concepción De La Rúa. 2012. Ancient DNa from hunter-gatherer and farmer groups from northern Spain supports a random dispersion model for the Neolithic expansion into Europe. PLoS ONE 7: e34417. [Google Scholar] [CrossRef]
- Higuchi, Russell, Barbara Bowman, Mary Freiberger, Oliver A. Ryder, and Allan C. Wilson. 1984. DNA sequences from the quagga, an extinct member of the horse family. Nature 312: 282–84. [Google Scholar] [CrossRef]
- Huber, Nicole, Walther Parson, and Arne Dür. 2018. Next generation database search algorithm for forensic mitogenome analyses. Forensic Science International: Genetics 37: 204–14. [Google Scholar] [CrossRef]
- Hummel, Susane. 2003. Ancient DNA Typing: Methods, Strategies, and Applications. Heidelberg and Berlin: Springer. [Google Scholar]
- Martínez-Labarga, Cristina, and Olga Rickards. 1999. La utilización del DNA antiguo en la investigación de la historia evolutiva humana. Revista Española de Antropología Biológica 20: 195–213. [Google Scholar]
- Meyer, Matthias, Qiaomei Fu, Ayinuer Aximu-Petri, Isabelle Glocke, Birgit Nickel, Juan-Luis Arsuaga, Ignacio Martínez, Ana Gracia, José María Bermúdez de Castro, Eudald Carbonell, and et al. 2014. A mitochondrial genome sequence of a hominin from Sima de los Huesos. Nature 505: 403–6. [Google Scholar] [CrossRef]
- Morris, Alan G., Anja Heinze, Eva KF Chan, Andrew B. Smith, and Vanessa M. Hayes. 2014. First ancient Mitochondrial Human genome from Prepastoralist Sothern African. Genome Biology and Evolution 6: 2647–53. [Google Scholar] [CrossRef]
- Mullis, Kary B., and Fred A. Faloona. 1987. Specific Synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods in Enzymology 155: 335–50. [Google Scholar] [CrossRef]
- Olalde, Íñigo, Hannes Schroeder, Marcela Sandoval-Velasco, Lasse Vinner, Irene Lobón, Oscar Ramirez, Sergi Civit, Pablo García Borja, Domingo C. Salazar-García, Sahra Talamo, and et al. 2015. A Common genetic origin for Early farmers from Mediterranean Cardial and Central European LBK Cultures. Molecular Biology and Evolution 32: 3132–42. [Google Scholar] [CrossRef]
- Pääbo, Svante. 1985. Preservation of DNA in ancient Egyptian mummies. Journal of Archaeological Science 12: 411–17. [Google Scholar] [CrossRef]
- Pääbo, Svante. 1989. Ancient DNA: Extraction, characterization, molecular cloning, and enzymatic amplification. Proceedings of the National Academy of Sciences 86: 1939–43. [Google Scholar] [CrossRef]
- Pääbo, Svante, and Allan C. Wilson. 1988. Polymerase chain reaction reveals cloning artifacts. Nature 334: 387–88. [Google Scholar] [CrossRef]
- Pääbo, Svante, Hendrik Poinar, David Serre, Viviane Jaenicke-Despres, Juliane Hebler, Nadin Rohland, Melanie Kuch, Johannes Krause, Linda Vigilant, and Michael Hofreiter. 2004. Genetic analyses from ancient DNA. Annual Review of Genetics 38: 645–79. [Google Scholar] [CrossRef]
- Palomo-Díez, Sara. 2015. Caracterízación Genética de las poblaciones de las Edades de Cobre y Bronce en la submeseta norte de la Península Ibérica. Ph.D. Thesis, Universidad Complutense de Madrid, Madrid, Spain. [Google Scholar]
- Palomo-Díez, Sara, and Ana María López-Parra. 2022. Utility and application of lineage markers: Mitochondrial DNA and Y Chromosome. In Handbook of DNA Profiling. Edited by Hirak Ranjan Dash, PanKaj Shrivastava and José Antonio Lorente. Heidelberg and Berlin: Springer, pp. 423–54. [Google Scholar] [CrossRef]
- Palomo-Díez, Sara, Ángel Esparza-Arroyo, Cláudia Gomes, Olga Rickards, Javier Velasco-Vázquez, Ana María López-Parra, Carlos Baeza Ribher, Cristina Martínez-Labarga, and Eduardo Arroyo-Pardo. 2017. Genetic characterization and determinatio of the number of individuals by molecular análisis in a prehistoric finding. Forensic Science International: Genetics 6: E487–E489. [Google Scholar] [CrossRef][Green Version]
- Palomo-Díez, Sara, Ángel Esparza-Arroyo, Mirian Tirado-Vizcaíno, Javier Velasco Vázquez, Ana María López Parra, Cláudia Gomes, Carlos Baeza-Richer, and Eduardo Arroyo-Pardo. 2018. Kinship analysis and allelic dropout: A Forensic approach on an archaeological case. Annals of Human Biology 45: 365–68. [Google Scholar] [CrossRef]
- Palomo-Díez, Sara, Cláudia Gomes, Ana María López-Parra, Carlos Baeza-Richer, Ángel Esparza-Arroyo, and Eduardo Arroyo-Pardo. 2016. Evaluación De Resultados Del Análisis De ADN Antiguo En Función De Las Características Macroscópicas De Las Muestras. In Poblaciones Humanas, Genética, Ambiente y Alimentación, 1st ed. Madrid: UAM Ediciones. ISBN 978-84-617-4098-7. [Google Scholar]
- Parson, Walther, and Arne Dür. 2007. EMPOP—A Forensic mtDNA database. Forensic Science International: Genetics 1: 88–92. [Google Scholar] [CrossRef]
- Parson, Walter, Leonor Gusmao, D.R. Hares, J.A. Irwin, W.R. Mayr, N. Morling, E. Pokorak, Meridit Prinz, Antonio Salas, Peter Schneider, and et al. 2014. DNA Commission of the International Society for Forensic Genetics: Revised and extended guidelines for mitochondrial DNA typing. Forensic Science International: Genetics 13: 134–42. [Google Scholar] [CrossRef]
- Prieto, Lourdes, Bettina Zimmermann, Ana Goios, A. Rodríguez-Monge, G.G. Paneto, Cintia Alves, Antonio Alonso, Cíntia Fridman, Sergio Cardoso, Gabriela Lima, and et al. 2011. The GHEP-EMPOP collaboration on mtDNA population data—A new resource for forensic casework. Forensic Science International: Genetics 5: 146–51. [Google Scholar] [CrossRef]
- Rodríguez Marcos, José Antonio. 2007. Estudio Secuencial de la Edad del Bronce en la Ribera del Duero (Provincia de Valladolid). Monografías. Arqueología en Castilla y León—7. Valladolid: Junta de Castilla y León. [Google Scholar]
- Rohland, Nadine, and Michael Hofreiter. 2007a. Ancient DNA extraction from bones an teeth. Nature Protocols 2: 1756–62. [Google Scholar] [CrossRef]
- Rohland, Nadine, and Michael Hofreiter. 2007b. Comparison and optimization of ancient DNa extraction. Bio Techniques 42: 343–52. [Google Scholar] [CrossRef]
- Rohland, Nadine, Heike Siedel, and Michael Hofreiter. 2009. A rapid column—Based ancient DNA extraction method for increased sample throughput. Molecular Ecology Resources 10: 677–83. [Google Scholar] [CrossRef]
- Rojo-Guerra, Manuel Kunst, Rafael Garrido-Pena, Íñigo García, and Martínez De Lagrán. 2006. La Neolitización de la Meseta Norte a la luz del C-14: Análisis de 47 Dataciones Absolutas Inéditas de dos Yacimientos Doméstico Sdle Valle de Ambrona, Soria, España. Valencia: Archivo de Prehistoria Levantina, vol. XXVI. [Google Scholar]
- Rovira Lloréns, Salvador. 2005. Metalurgia de crisol: La obtención de cobre en la prehistoria de la Península Ibérica. De re Metallica (Madrid): Revista de la Sociedad Española para la Defensa del Patrimonio Geológico y Minero, 78–94. [Google Scholar]
- Scorrano, Gabriele, Gabriele Scorrano, Federica Valentini, Cristina Martínez-Labarga, Mario Federico Rolfo, Antonella Fiammenghi, Domenico Lo Vetro, Fabio Martini, Antonella Casoli, Giovanni Ferraris, and et al. 2014. Methodological strategies to assess the degree of bone preservation for ancient DNA studies. Annals of Human Biology 42: 10–19. [Google Scholar] [CrossRef]
- Slatkin, Montgomery. 1995. A measure of population subdivision based on microsatellite allele frequencies. Genetics 139: 457–62. [Google Scholar] [CrossRef]
- van Oven, Mannis, and Manfred Kayser. 2008. Updated Comprehensive Phylogenetic Tree of Global Human Mitochondrial DNA Variation. Human Mutation. Mutation in Brief 30: E386–E394. [Google Scholar] [CrossRef]
- Villalba-Mouco, Vanessa, Marieke S. van de Loosdrecht, Cosimo Posth, Rafael Mora, Jorge Martínez-Moreno, Manuel Rojo-Guerra, Domingo C. Salazar-García, José I Royo-Guillén, Michael Kunst, Hélène Rougier, and et al. 2019. Survival of Late Pleistocene Hunter-Gatherer Ancestry in the Iberian Peninsula. Current Biology 29: 1169–77. [Google Scholar] [CrossRef]
- Whallon, Robert. 1968. A new approach to pottery typology. American Antiquity 37: 13–33. [Google Scholar] [CrossRef]
- Zimmermann, Bettina, Alexander Röck, Gabriela Huber, Tanja Krämer, Peter M. Schneider, and Walther Parson. 2011. Application of a west Eurasian-specific filter for quasi-median network analysis: Sharpening the blade for mtDNA error detection. Forensic Science International: Genetics 5: 133–37. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Archaeological Site | Lab. Name | Estimated Antiquity | Location in Spain | Individuals | Period |
|---|---|---|---|---|---|
| El Tomillar | ATOM | 4000 BP | Bercial de Zapardiel, Ávila | 8 | Pre-Bell Beaker |
| El Tomillar | ATOM | 3780 ± 100–3830 ± 95 BP | Bercial de Zapardiel, Ávila | 11 | Pre-Bell Beaker |
| Los Areneros | ARE | 4240 ± 35–4125 ± 25 BP. | La Lastrilla, Segovia | 8 | Pre-Bell Beaker |
| Los Cercados | CER | 3970 ± 60 BP. | Mucientes, Valladolid | 3 | Pre-Bell Beaker |
| Camino de Trascabañas | TC | 4180 ± 35 BP. | Simancas y Ciguñuela | 1 | Pre-Bell Beaker |
| Aldeagordillo | ALG | 3690 ± 50 BP | Ávila | 4 | Bell Beaker |
| El Mirador | MIR | 3650 ± 35 BP | La Lastrilla, Segovia | 1 | Bell Beaker |
| Perro Alto | PA | 3730 ± 65 BP | Fuente Olmedo, Valladolid | 1 | Bell Beaker |
| Santa Cruz de Cabezón II | CAB | Not available | Cabezón de Pisuerga, Valladolid | 1 | Bell Beaker |
| Pago de Valhondo | PAD | 3970 ± 50 BP | Pajares de Adaja, Ávila | 1 | Bell Beaker |
| Terrazas del Manzanares | RIV | 3050 ± 100 BP | Rivas Vaciamadrid, Madrid | 1 | Bell Beaker |
| Tablada del Rudrón | TR | Not available | Tubilla del Agua, Burgos | 4 | Bell Beaker |
| Santioste | SAN | 3780 ± 50 BP | Otero de Sariegos, Zamora | 1 | Bronze Age |
| Cueva de la Revilla | REV | 3550 ± 40–3325 ± 35 BP | Atapuerca, Burgos | 8 | Bronze Age |
| Carrelasvegas | CV | 3230 ± 80 BP | Santillana de Campos, Palencia | 1 | Proto-Cogotas I |
| El Cementerio | QUO | 480 ± 35 BP | Quintanilla de Onésimo, Valladolid | 1 | Proto-Cogotas I |
| El Cerro de la Horra | HOR | 3180 ± 50–3225 ± 30 BP | La Horra, Burgos | 3 | Proto-Cogotas I |
| El Juncal | JUN | 3.335 ± 35 BP | Villaralbo, Zamora | 1 | Proto-Cogotas I |
| Fuente de la Mora | FM | Not available | Valladolid | 3 | Proto-Cogotas I |
| La Huelga | HU | 3290 ± 35 BP | Dueñas, Palencia | 1 | Proto-Cogotas I |
| Los Rompizales | RPZ | 3165 ± 30–3250 ± 35 BP | Quintanadueñas, Burgos | 4 | Proto-Cogotas I |
| Las Cañamonas | CÑ | 3205 BP | San Cristobal de Entreviñas, Zamora | 1 | Proto-Cogotas I |
| Los Tolmos | TOL | 3600–3.200 BP | Caracena, Soria | 4 | Proto-Cogotas I |
| Tordillos | TOR | 3205 ± 35 BP | Aldeasca de la Forentera, Salamanca | 12 | Proto-Cogotas I—Cogotas I |
| La Requejada | LR | 3020 ± 35–3120 ± 30 BP | San Román de Hornija, Valladolid | 3 | Cogotas I |
| Cerro de la Cabeza | CC | 2160 ± 50 BP | Ávila | 2 | Cogotas I |
| Canto Blanco | CB | 3123 ± 30 BP | Sahagún, León | 1 | Cogotas I |
| El Cerro de San Pelayo | SP | 2715 ± 30 BP | Salamanca | 1 | Soto formativo |
| Total Number of Individuals | 91 | ||||
| Group 1: Paleolithic-Mesolithic N = 4 (Hervella et al. 2012) | Group 2: Final Neolithic N = 42 (Hervella et al. 2012) | Group 3 Pre-Bell-Beaker N = 17 | Group 4 Bell-Beaker N = 9 | Group 5 Ancient Bronze N = 11 | Group 6 Proto-Cogotas I N = 25 | Group 7 Cogotas I N = 3 | Group 8 XXI Century N = 335 (Prieto et al. 2011) |
|---|---|---|---|---|---|---|---|
| Erraya | Cascajos | Los Areneros | El Mirador | La Revilla Cave | Tordillos | Cantoblanco | XXI Century Northern Iberian Subplateau |
| La Pasiega | Fuente Hoz | Los Cercados | Rivas-Vaciamadrid | Santioste | Las Cañamonas | La Requejada | XXI Century Cantabric Coast |
| La Chora | Paternabidea | Trascabañas | Santa Cruz de Cabezón | Urtiaga (Hervella et al. 2012) | El Cerro de la Horra | XXI Century Mediterranean Coast | |
| Aizpea | Cueva de Marizulo | El Tomillar | Tablada de Rudrón | Fuente de la Mora | XXI Century Basque Country | ||
| Pajares de Adaja | El Cementerio | ||||||
| Perro Alto | La Huelga | ||||||
| Los Rompizales | |||||||
| Carrelasvegas |
| Group 1: Region 1 N = 2 | Group 2: Region 2 N = 12 | Group 3: Region 3 N = 19 | Group 4: Region 4 N = 16 | Group 5: Region 5 N = 14 |
|---|---|---|---|---|
| Las Cañamonas | Santa Cruz de Cabezón | El Cerro de la Horra | El Tomillar | Los Areneros |
| Santioste | Fuente de la Mora | Tablada de Rudrón | Pajares de Adaja | El Mirador |
| Los Cercados | Cueva de la Revilla | Perro Alto | El Cementerio | |
| Trascabañas | Los Rompizales | Tordillos | Los Tolmos | |
| La Huelga | Cerro de San Pelayo | |||
| Cantoblanco | ||||
| La Requejada |
| Period | Individual | Range | mtDNA Haplotype |
|---|---|---|---|
| Pre-Bell Beaker | 7BTOM | (16,105–16,399) (29–309) | 16,183C 16,189C 16,234A 16,299G 263G 291.1A |
| 9BTOM | (16,105–16,399) (29–309) | 16,183C 16,189C 16,234A 16,299G 263G 291.1A | |
| Pre-Bell Beaker | 1ARE | (16,106–16,399) (29–309) | 16,224C 16,311C 73G 263G |
| 2ARE | (16,105–16,399) (29–389) | 16,224C 16,311C 16,319A 73G 152C 263G 277T 315.1C | |
| 5ARE | (16,105–16,399) (29–389) | 16,224C 16,311C 16,319A 73G 152C 263G 309.1C 315.1C | |
| 6ARE | (16,105–16,399) (29–389) | 16,224C 16,311C 73G 150T 239C 263G 309.1C 315.1C | |
| 20ARE | (16,105–16,399) (29–80) (240–390) | 16,224C 16311C 73G 263G 309.1C 315.1C | |
| Proto-Cogotas I | 1ELC | (16,105–16,399) (50–190) | 16,298C 64T 72C |
| 2ELC | (16,105–16,399) (50–190) | 16,298C 64T 72C | |
| 3ELC | (16,105–16,399) (50–190) | 16,298C 64T 72C | |
| LTB3 | (16,105–16,399) (29–120) | 16,224C 16,311C 16,362C 73G | |
| LTB2 | (16,105–16,399) | 16,224C 16,311C | |
| LTB1 | (16,105–16,399) (29–120) | 16,224C 16,311C 16,362C 73G | |
| 2TOR | (16,105–16,399) (29–310) | 16,362C 73G 150T 263G 291.1A | |
| 6TOR | (16,105–16,399) (29–390) | 263G 315.1C | |
| 7TOR | (16,105–16,399) (29–310) | 16,362C 73G 150T 263G 291.1A | |
| 8TOR | (16,105–16,399) (29–390) | 263G 315.1C | |
| 1RPZ | (16,105–16,399) (29–128) | 16,192T 16,270T 16,304C 73G | |
| 3RPZ | (16,105–16,399) | 16,192T 16,270T |
| Source of Variation | d.f. | Sum of Squares | Components Variation | % of Variation |
|---|---|---|---|---|
| Among groups | 4 | 8.035 | 0.024 Va | 1.74 |
| Among populations | 18 | 27.414 | 0.113 Vb | 8.14 |
| Among individuals | 40 | 50.392 | 1.259 Vc | 90.12 |
| % Variation among Groups | Pre-Bell Beaker | Bell Beaker | Ancient Bronze Age | Proto-Cogotas I |
|---|---|---|---|---|
| Pre-Bell Beaker (n = 20) | ||||
| Bell Beaker (n = 9) | 5.37 (FST fixation Index = 0.05745; p = 0.33920) | |||
| Ancient Bronze Age (n = 9) | 2.91 (FST fixation index = 0.13076; p = 0.06061) | 16.06 (FST fixation index = −0.25644; p = 0.94917) | ||
| Proto-Cogotas I (n = 26) | 5.99 (FST fixation index = 0.25409; p = 0) | 4.78 (FST fixation index = 0.11769; p = 0.09873) | 9.39 (FST fixation index = 0.08724; p = 0.06549) | |
| Cogotas I (n = 4) | 5.28 (FST fixation index = 0.15807; p = 0.13490) | 0.77 (FST fixation index = −0.42670; p = 0.87292) | 16.87 (FST fixation index = 0.29316; p = 0.89345) | 12.47 (FST fixation index = 0.13386; p = 0.02639) |
| Source of Variation | d.f. | Sum of Squares | Variance Components | % of Variation |
|---|---|---|---|---|
| Among groups | 8 | 17.337 | 0.02850 Va | 1.75 |
| Among populations | 24 | 36.951 | −0.00756 Vb | −0.46 |
| Among individuals | 276 | 443.557 | 1.60709 Vc | 98.71 |
| % Variation among Groups | Paleolithic–Mesolithic | Final Neolithic | Pre-Bell Beaker | Bell Beaker | Ancient Bronze Age | Proto-Cogotas I | Cogotas I |
|---|---|---|---|---|---|---|---|
| Paleolithic–Mesolithic | |||||||
| Final Neolithic | 10.28 (FST fixation index = −0.13857; p = 0.23656) | ||||||
| Pre-Bell Beaker (n = 20) | 11.28 (FST fixation index = −0.22096; p = 0.07722) | 11.36 (FST fixation index = −0.23925; p = 0) | |||||
| Bell Beaker (n = 9) | 8.63 (FST fixation index = −0.28990; p = 0.68817) | 0.06 (FST fixation index = −0.02227; p = 0.56891) | 3.94 (FST fixation index = −0.09343; p = 0.20919) | ||||
| Ancient Bronze Age (n = 9) | 11.39 (FST fixation index = −0.57862; p = 0.88172) | 2.25 (FST fixation index = −0.01562; p = 0.637349 | 5.21 (FST fixation index = −0.13987; p = 0.07527) | 15.50 (FST fixation index = −0.32931; p = 0.97458) | |||
| Proto Cogotas I (n = 26) | 7.78 (FST fixation index = −0.13671; p = 0.01466) | 0.15 (FST fixation index = −0.14797; p = 0) | 7.79 (FST fixation index = −0.27062; p = 0) | 4.45 (FST fixation index = 0.08344; p = 0.10655) | 6.03 FST fixation index = 0.06619; p = 0.07136) | ||
| Cogotas I (n = 4) | 7.84 (FST fixation index = −0.01961; p = 0.42620) | 2.65 (FST fixation index = −0.004181; p = 0.55621) | 3.28 (FST fixation index = −0.18059; p = 0.07527) | 0.77 (FST fixation index = −0.42670; p = 0.87977) | 16.44 (FST fixation index = −0.39237; p = 0.95112) | 11.80 (FST fixation index = −0.09426; p = 0.01564) | |
| XXI Century | 3.31 (FST fixation index = −0.03832; p = 0.71750) | 0.88 (FST fixation index = −0.00487; p = 0.48387 | 8.58 (FST fixation index = −0.10048; p = 0) | 1.60 (FST fixation index = −0.03276; p = 0.95601) | 2.64 (FST fixation index = −0.03194; p = 0.85826) | 0.23 (FST fixation index = −0.01411; p = 0.06843) | −8.95 (FST fixation index = −0.09194; p = 0.76442) |
| Source of Variation | d.f. | Sum of Squares | Components Variation | % of Variation |
|---|---|---|---|---|
| Among groups | 4 | 10.308 | 0.087 Va | 6.11 |
| Among populations | 18 | 25.840 | 0.074 Vb | 5.19 |
| Among individuals | 40 | 50.392 | 1.260 Vc | 88.70 |
| % Variation | Region 1 | Region 2 | Region 3 | Region 4 | |
|---|---|---|---|---|---|
| Region 1 | Among groups | ||||
| Region 2 | Among groups | 3.65 (FST fixation index: −0.29677; p = 0.849469) | |||
| Region 3 | Among groups | 9.81 (FST fixation index: 0.05415; p = 0.11144) | 1.04 (FST fixation index: −0.03169; p = 0.63441) | ||
| Region 4 | Among groups | 9.68 (FST fixation index: 0.00790; p = 0.31085) | 4.87 (FST fixation index: 0.01899; p = 0.50733) | 1.81 (FST fixation index: −1.81; p = 0.12805) | |
| Region 5 | Among groups | 13.60 (FST fixation index: 0.34047; p = 0.02737) | 3.92 (FST fixation index: 0.11036; p = 0.19648) | 11.30 (FST fixation index: 0.22580; p = 0.00196) | 15.63 (FST fixation index: 0.30023; p = 0.00196) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomo-Díez, S.; Esparza-Arroyo, Á.; Rickards, O.; Martínez-Labarga, C.; Arroyo-Pardo, E. How Mitochondrial DNA Can Write Pre-History: Kinship and Culture in Duero Basin (Spain) during Chalcolithic and Bronze Age. Genealogy 2023, 7, 51. https://doi.org/10.3390/genealogy7030051
Palomo-Díez S, Esparza-Arroyo Á, Rickards O, Martínez-Labarga C, Arroyo-Pardo E. How Mitochondrial DNA Can Write Pre-History: Kinship and Culture in Duero Basin (Spain) during Chalcolithic and Bronze Age. Genealogy. 2023; 7(3):51. https://doi.org/10.3390/genealogy7030051
Chicago/Turabian StylePalomo-Díez, Sara, Ángel Esparza-Arroyo, Olga Rickards, Cristina Martínez-Labarga, and Eduardo Arroyo-Pardo. 2023. "How Mitochondrial DNA Can Write Pre-History: Kinship and Culture in Duero Basin (Spain) during Chalcolithic and Bronze Age" Genealogy 7, no. 3: 51. https://doi.org/10.3390/genealogy7030051
APA StylePalomo-Díez, S., Esparza-Arroyo, Á., Rickards, O., Martínez-Labarga, C., & Arroyo-Pardo, E. (2023). How Mitochondrial DNA Can Write Pre-History: Kinship and Culture in Duero Basin (Spain) during Chalcolithic and Bronze Age. Genealogy, 7(3), 51. https://doi.org/10.3390/genealogy7030051