


Lettuce in Monoculture or in Intercropping with Tomato Changes the Antioxidant Enzyme Activities, Nutrients and Growth of Lettuce


 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
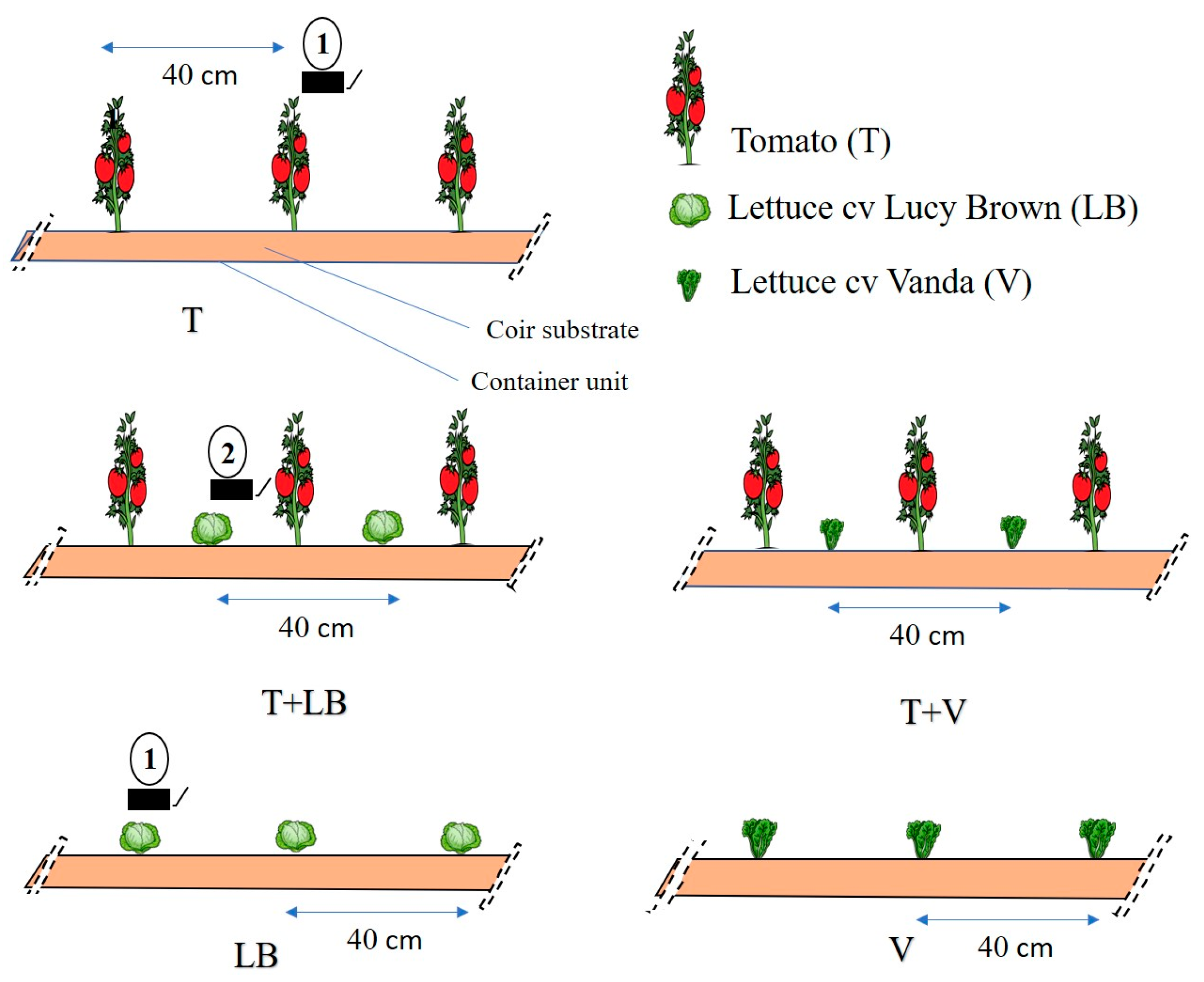
2.1. Treatments and Experimental Design
2.2. Plant Growing Conditions
2.3. Greenhouse Climate Measurements
2.4. Biochemical Evaluations of Lettuce
2.4.1. Lipid Peroxidation
2.4.2. Antioxidant Enzyme Activities
2.4.3. Catalase
2.4.4. Superoxide Dismutase
2.4.5. Peroxidase
2.5. Efficiency of Nutrient Absorption in Lettuce
2.6. Plant Growth Biometric Parameters
2.7. Economic Factors
2.8. Statistical Analysis
3. Results and Discussion
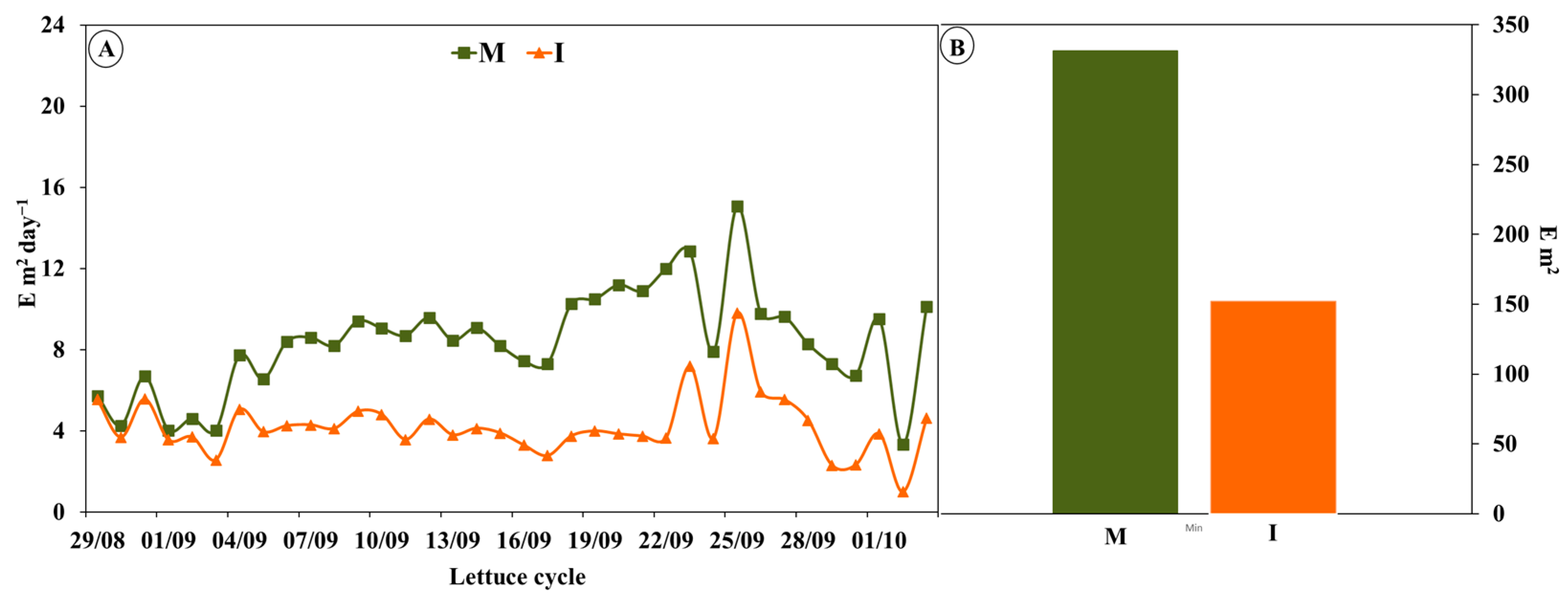
3.1. Incident Radiation
3.2. Biochemical Indicators in Plants
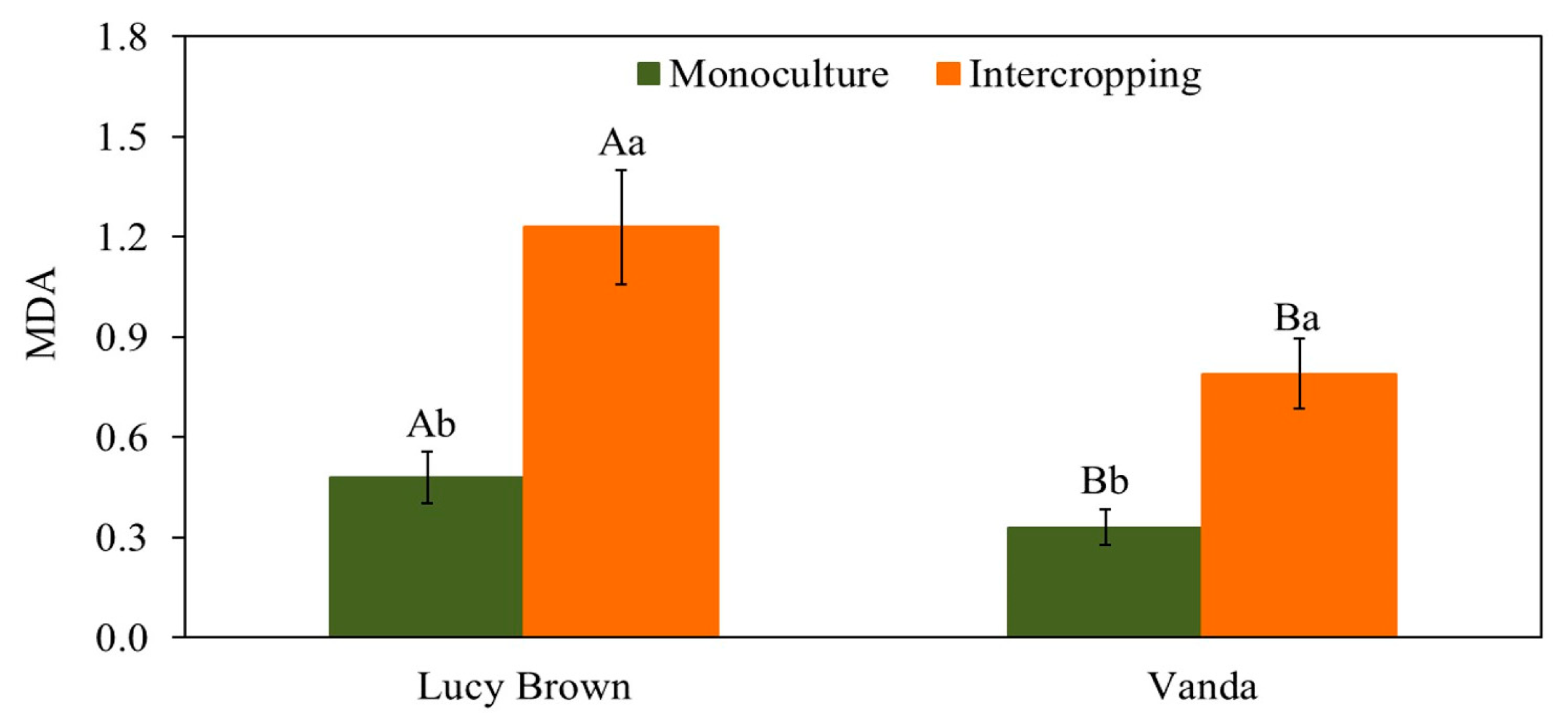
3.2.1. Lipid Peroxidation Measurement
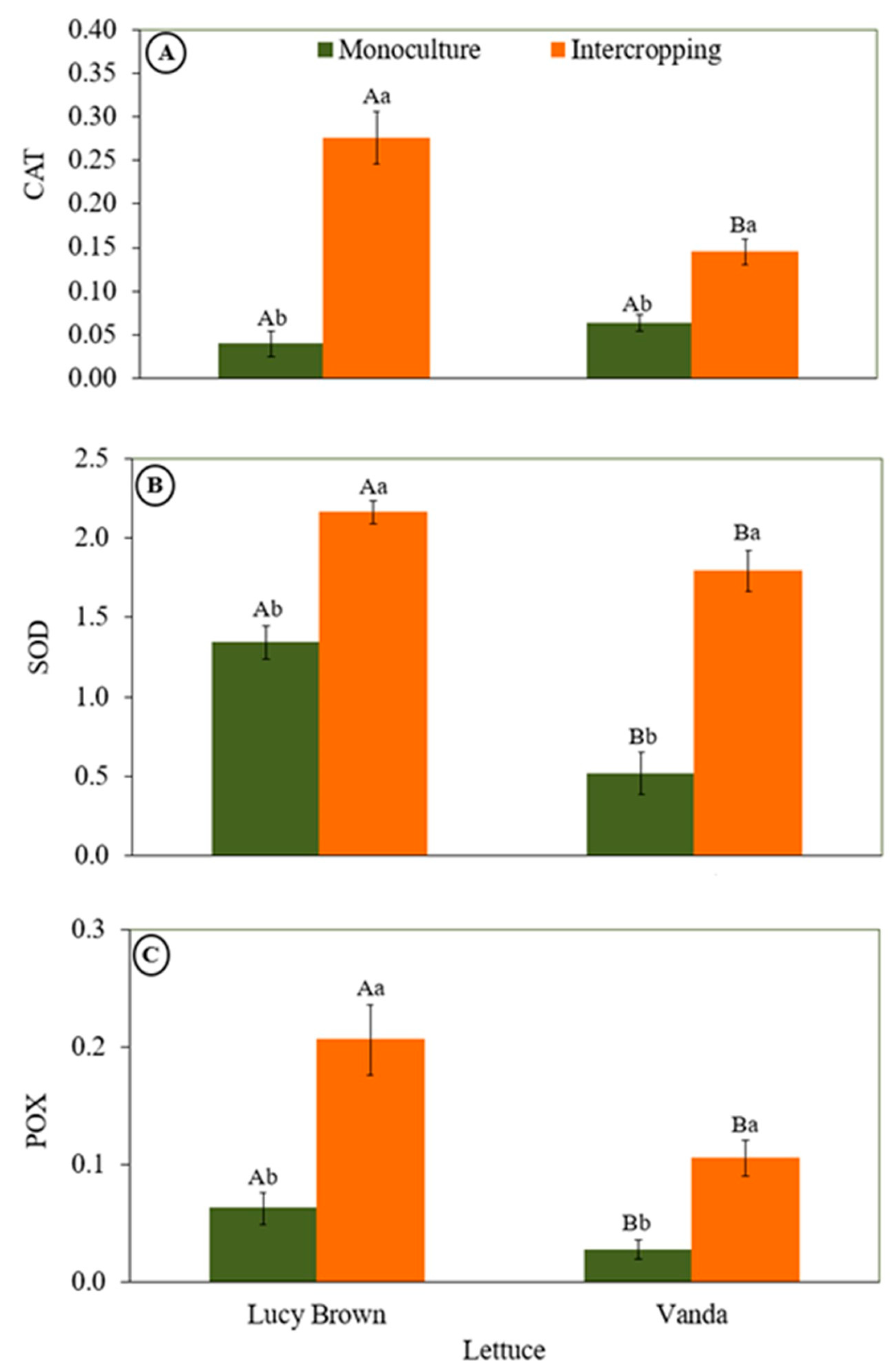
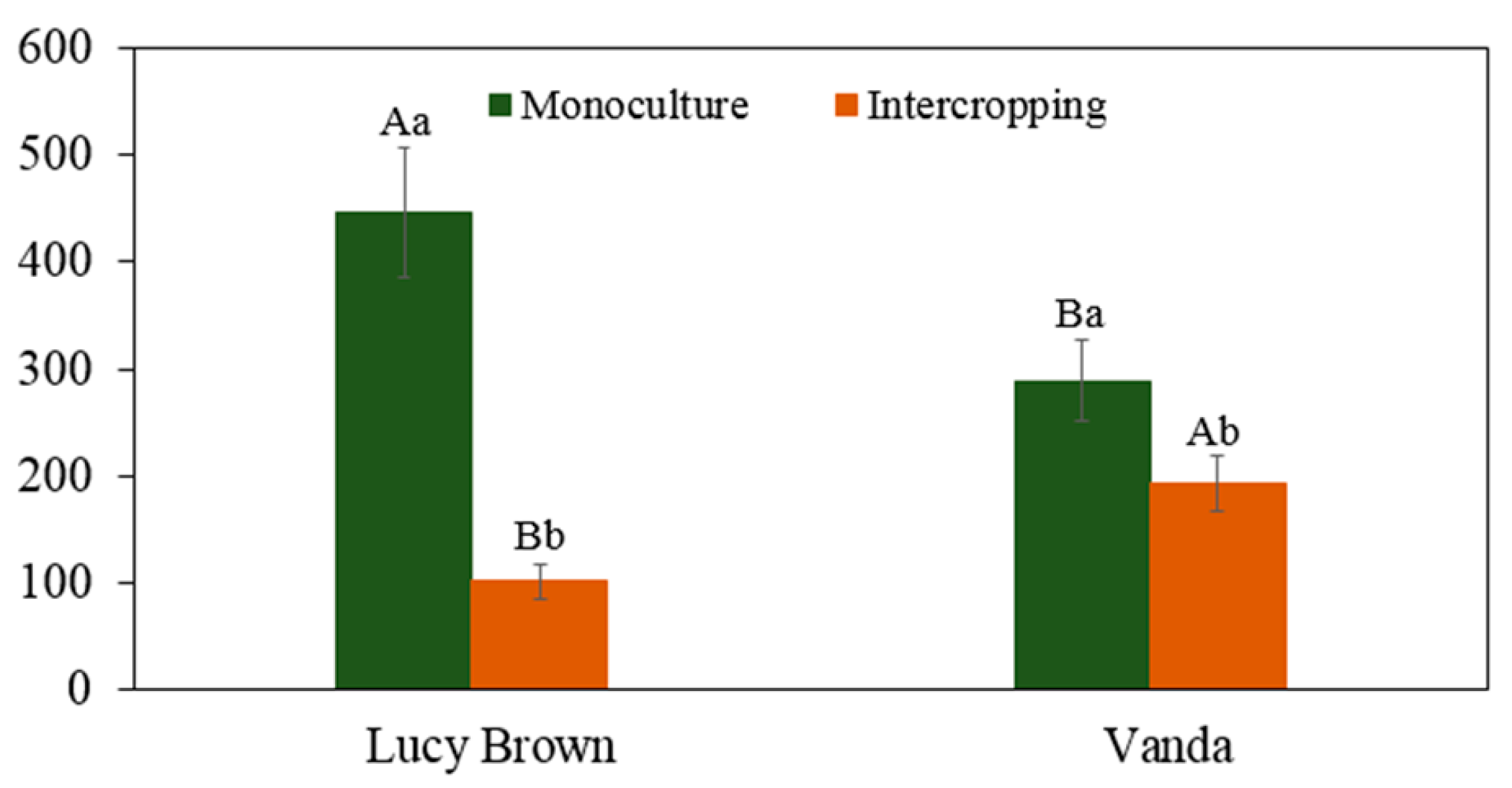
3.2.2. Antioxidant Enzyme Activities
3.3. Efficiency of Nutrient Absorption in Lettuce
3.4. Lettuce Growth
3.5. Tomato Growth
3.6. Economic Benefit
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wu, Y.; Gong, W.; Yang, F.; Wang, X.; Yong, T.; Yang, W. Responses to shade and subsequent recovery of soya bean in maize-soya bean relay strip intercropping. Plant Prod. Sci. 2016, 19, 206–214. [Google Scholar] [CrossRef]
- Francis, C.A. Biological efficiencies in multiple-cropping systems. Adv. Agron. 1989, 42, 1–42. [Google Scholar] [CrossRef]
- Willey, R.W. Intercropping: Its importance and research needs. I. Competition and yield advantages. Field Crop Abst. 1979, 32, 1–10. [Google Scholar]
- Rao, M.R.; Willey, R.W. Evaluation of yield stability in intercropping: Studies on sorghum/pigeonpea. Exp. Agric. 1980, 16, 105–116. [Google Scholar] [CrossRef]
- Sarrantonio, M.; Gallandt, E. The role of cover crops in North American cropping systems. J. Crop Prod. 2008, 8, 53–74. [Google Scholar] [CrossRef]
- Huang, C.; Liu, Q.; Gou, F.; Li, X.; Zhang, C.; Werf, W.V.D.; Zhang, F. Plant growth patterns in a tripartite strip relay intercrop are shaped by asymmetric aboveground competition. Field Crops Res. 2017, 201, 41–51. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, W.; Wu, Q. Effects of shading in maize/soybean relay-cropping system on the photosynthetic characteristics and yield of soybean. Acta Agron. Sin. 2007, 9, 1502–1507. [Google Scholar]
- Yang, F.; Wang, X.; Liao, D.; Lu, F.; Gao, R.; Liu, W.; Yong, T.; Wu, X.; Du, J.; Yang, W. Yield response to different planting geometries in maize–soybean relay strip intercropping systems. Agron. J. 2015, 107, 296–304. [Google Scholar] [CrossRef]
- Chen, Y. Development of agricultural recycle economy in arid areas of Hexi Corridor. J. Anhui Agric. Sci. 2011, 6, 3726–3728. [Google Scholar] [CrossRef]
- Rahman, T.; Liu, X.; Hussain, S.; Ahmed, S.; Chen, G.; Yang, F.; Chen, L.; Du, J.; Liu, W.; Yang, W. Water use efficiency and evapotranspiration in maize-soybean relay strip intercrop systems as affected by planting geometries. PLoS ONE 2017, 12, e0178332. [Google Scholar] [CrossRef]
- Nascimento, C.S.; Cecílio Filho, A.B.; Mendoza-Cortez, J.W.; Nascimento, C.S.; Bezerra Neto, F.; Grangeiro, L.C. Effect of population density of lettuce intercropped with rocket on productivity and land-use efficiency. PLoS ONE 2018, 13, e0194756. [Google Scholar] [CrossRef]
- Bezerra Neto, F.; Andrade, F.V.; Negreiros, M.Z.; Santos Júnior, J.J. Desempenho agroeconômico do consórcio cenoura x alface lisa em dois sistemas de cultivo em faixa. Hortic. Bras. 2003, 21, 635–641. [Google Scholar] [CrossRef]
- Cecílio Filho, A.B.; Rezende, B.L.A.; Barbosa, J.C.; Feltrim, A.L.; Silva, G.S.; Grangeiro, L.C. Interaction between lettuce and tomato plants, in intercropping cultivation, established at different times, under protected cultivation. Hortic. Bras. 2008, 26, 158–164. [Google Scholar] [CrossRef]
- Tringovska, I.; Yankova, V.; Markova, D.; Mihov, M. Effect of companion plants on tomato greenhouse production. Sci. Hortic. 2015, 186, 31–37. [Google Scholar] [CrossRef]
- Cunha-Chiamolera, T.P.L.; Urrestarazu, M.; Cecílio-Filho, A.B.; Morales, I. Agronomic and economic feasibility of tomato and lettuce intercropping in a soilless system as a function of the electrical conductivity of the nutrient solution. HortScience 2017, 52, 1195–1200. [Google Scholar] [CrossRef]
- Malézieux, E.; Crozat, Y.; Dupraz, C.; Laurans, M.; Makowski, D.; Ozier-Lafontaine, H.; Rapidel, B.; Tourdonnet, S.; Valantin-Morison, M. Mixing plant species in cropping systems: Concepts, tools and models. A review. Agron. Sustain. Dev. 2009, 29, 43–62. [Google Scholar] [CrossRef]
- Liu, X.; Rahman, T.; Song, C.; Su, B.; Yang, F.; Yong, T.; Wu, Y.; Zhang, C.; Yang, W. Changes in light environment, morphology, growth and yield of soybean in maize-soybean intercropping systems. Field Crops Res. 2017, 200, 38–46. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Rizhsky, L.; Hegie, A.; Koussevitzky, S.; Mittler, R. Double mutants deficient in cytosolic and thylakoid ascorbate e peroxidase reveal a complex mode of interaction between reactive oxygen species, plant development, and response to abiotic stresses. Plant Physiol. 2007, 144, 1777–1785. [Google Scholar] [CrossRef]
- Möller, I.M.; Sweetlove, L.J. ROS signalling—Specificity is required. Trends Plant Sci. 2010, 15, 370–374. [Google Scholar] [CrossRef]
- Wrzaczek, M.; Brosché, M.; Kangasjärvi, J. ROS signaling loops—Production, perception, regulation. Curr. Opin. Plant Biol. 2013, 16, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.M.; Huang, L.F.; Zhang, X.; Wang, M.L.; Xu, G.Y.; Xia, X.J. OsCML4 improves drought tolerance through scavenging of reactive oxygen species in rice. J. Plant Biol. 2015, 58, 68–73. [Google Scholar] [CrossRef]
- Cordovilla, M.P.; Bueno, M.; Aparicio, C.; Urrestarazu, M. Effects of salinity and the interaction between Thymus vulgaris and Lavandula angustifolia on growth, ethylene production and essential oil contents. J. Plant Nutr. 2014, 37, 875–888. [Google Scholar] [CrossRef]
- Davey, M.; Stals, E.; Panis, B.; Keulemans, J.; Swennen, R. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem. 2005, 347, 201–207. [Google Scholar] [CrossRef]
- Rio, D.D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovas. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Tang, K.; Zhan, J.C.; Yang, H.R.; Huang, W.D. Changes o fresveratrol and antioxidant enzymes during UV-induced plant defense response in peanut seedlings. J. Plant Physiol. 2010, 167, 95–102. [Google Scholar] [CrossRef]
- Rai, A.C.; Singh, M.; Shah, K. Effect of water withdrawal on formation of free radical, proline accumulation and activities of antioxidant enzymes in ZAT12-transformed transgenic tomato plants. Plant Physiol. Biochem. 2012, 61, 108–114. [Google Scholar] [CrossRef]
- Gratão, P.L.; Monteiro, C.C.; Tezotto, T.; Carvalho, R.F.; Alves, L.R.; Peters, L.J.; Azevedo, R.A. Cadmium stress antioxidante responses and root-to-shoot communication in grafted tomato plants. Biometals 2015, 28, 803–816. [Google Scholar] [CrossRef]
- Abouzari, A.; Fakheri, B.A. Reactive oxygen species: Generation, Oxidative Damage, and Signal Transduction. Int. J. Life Sci. 2015, 9, 3–17. [Google Scholar] [CrossRef]
- Alves, R.C.; Oliveira, K.R.; Lúcio, J.C.B.; Silva, J.S.; Carrega, W.C.; Queiroz, S.F.; Gratão, P.L. Exogenous foliar ascorbic acid applications enhance salt-stress tolerance in peanut plants throughout an increase in the activity of major antioxidant enzymes. S. Afr. J. Bot. 2022, 150, 759–767. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosys. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in theentire antioxidant defence grid. O.M. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Pozo, J.; Alvaro, J.E.; Morales, I.; Requena, J.; La Malfa, T.; Mazuela, P.; Urrestarazu, M. A new local sustainable inorganic material for soilless culture in Spain: Granulated volcanic rock. HortScience 2014, 49, 1537–1541. [Google Scholar] [CrossRef]
- Sonneveld, C.; Straver, N.B. Nutrient solution for vegetables and flowers grown in water or substrates. Voedingspolossingen glastijnbouw 1994, 8, 1–33. [Google Scholar]
- Shimizu, N.; Hosogi-Park, P.J. Reactive oxygen species (ROS) generation ROS-induced lipid peroxidation are associated with plasma membrane modifications in host cells in response to AK-toxin I from Alternaria alternate Japonese pear pathotype. J. Gen. Plant Pathol. 2006, 72, 6–15. [Google Scholar] [CrossRef]
- Gratão, P.L.; Monteiro, C.C.; Carvalho, R.F.; Tezotto, T.; Piotto, F.A.; Peres, L.E.P.; Azevedo, R.A. Biochemical dissection of diageotropica and never ripe tomato mutants to Cd stressful conditions. Plant Physiol. Biochem. 2012, 56, 79–96. [Google Scholar] [CrossRef]
- Cia, M.C.; Guimarães, A.C.R.; Medici, L.O.; Chabregas, S.M.; Azevedo, R.A. Antioxidant response to water deficit by drought-tolerant and -sensitive sugarcane varieties. Ann. Appl. Biol. 2012, 161, 313–324. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutase: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Teisseire, H.; Guy, V. Copper-induced changes in antioxidant enzymes activities in fronds of duckweed (Lemna minor). Plant Sci. 2000, 153, 65–72. [Google Scholar] [CrossRef]
- Malavolta, E.; Vitti, G.C.; Oliveira, S.A. Avaliação do Estado Nutricional da Plantas: Princípios e Aplicações; Potafós Publ.: Piracicaba, Brazil, 1997; p. 319. [Google Scholar]
- Commission Implementing Regulation (EU) No 543/2011 of 7 June 2011 Laying Down Detailed Rules for the Application of Council Regulation (EC) No 1234/2007 in Respect of the Fruit and Vegetables and Processed Fruit and Vegetables Sectors. January 2023. Available online: https://eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX%3A32011R0543 (accessed on 15 March 2023).
- CEAGESP. Cotações: Legumes. Available online: http://www.ceagesp.com.br/ (accessed on 17 February 2023).
- Barbosa, J.C.; Maldonado, J.W. AgroEstat—Sistema para Análises Estatísticas de Ensaios Agronômicos; FCAV/UNESP: Jaboticabal, Brazil, 2015. [Google Scholar]
- Cunha-Chiamolera, T.P.L.; Morales, I.; Cecílio-Filho, A.B.; Urrestarazu, M. Viabilidad de los cultivos asociados de hortalizas en los sistemas sin suelo. Agrícola Vergel 2015, 34, 148–152. [Google Scholar]
- Gong, W.Z.; Jiang, C.D.; Wu, Y.S.; Chen, H.H.; Liu, W.Y.; Yang, W.Y. Tolerance vs. avoidance: Two strategies of soybean (Glycine max) seedlings in response to shade in intercropping. Photosynthetica 2015, 53, 259–268. [Google Scholar] [CrossRef]
- Correia, B.; Hancock, R.; Amaral, J.; Gómez-Cadenas, A.; Valledor, L.; Pinto, G. Combined drought and heat activities protective responses in Eucalyptus globulus that are not activated when subjected to drought and heat stress alone. Front. Plant Sci. 2018, 9, 819. [Google Scholar] [CrossRef]
- Nascimento, C.S.; Nascimento, C.S.; Lopes, G.; Carrasco, G.; Gratão, P.L.; Cecílio Filho, A.B. Biofortified rocket (Eruca sativa) with selenium by using the nutrient film technique. Horticulturae 2022, 8, 1088. [Google Scholar] [CrossRef]
- Roy, P.R.; Ul-Arif, M.T.; Akter, T.; Ray, S.R.; Sayed, M.A. Exogenous ascorbic acid and hydrogen peroxide alleviates salt-induced oxidative stress in rice (Oryza sativa L.) by enhancing antioxidant enzyme activities and proline content. Adv. Reinar. Biol. 2016, 10, 148–154. [Google Scholar]
- Niu, L.; Liao, W. Hydrogen peroxide signaling in plant development and abiotic responses: Crosstalk with nitric oxide and calcium. Front. Plant Sci. 2016, 7, 230. [Google Scholar] [CrossRef]
- Alves, R.C.; Nicolau, M.C.M.; Checchio, M.V.; Sousa-Junior, G.S.; Oliveira, F.A.; Prado, M.R.; Gratão, P.L. Salt stress alleviation by seed priming with silicon in lettuce seedlings: An approach based on enhancing antioxidant responses. Bragantia 2020, 79, 19–29. [Google Scholar] [CrossRef]
- Wang, M.; Wu, C.; Cheng, Z.; Meng, H. Growth and physiological changes in continuously cropped eggplant (Solanum melongena L.) upon relay intercropping with garlic (Allium sativum L.). Front. Plant Sci. 2015, 6, 262. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, Y.; Xu, Z.; Chen, Z.; Duan, L. Combining physiological and metabolomic analysis to unravel the regulations of coronatine alleviating water stress in tobacco (Nicotiana tabacum L.). Biomolecules 2020, 10, 835. [Google Scholar] [CrossRef]
- Alves, R.C.; Rossatto, D.R.; Silva, J.S.; Checchio, M.V.; Oliveira, K.R.; Oliveira, F.A.; Queiroz, S.F.; Cruz, M.C.P.; Gratão, P.L. Seed priming with ascorbic acid enhances salt tolerance in micro-tom tomato plants by modifying the antioxidant defense system components. Biocatal. Agric. Biotechnol. 2021, 31, 101927. [Google Scholar] [CrossRef]
- Kolahi, M.; Mohajel-Kazemi, E.; Yazdi, M.; Goldson-Barnaby, A. Oxidative stress induced by cadmium in lettuce (Lactuca sativa Linn.): Oxidative stress indicators and prediction of their genes. Plant Physiol Biochem. 2020, 146, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Trani, P.E.; van Raij, B. Hortaliças. In Recomendações de Adubação e Calagem para o Estado de São Paulo (Boletim Técnico, 100), 2nd ed.; van Raij, B., Cantarella, H., Quaggio, J.A., Furlani, A.M.C., Eds.; IAC: Campinas, Brazil, 1997; pp. 155–164. [Google Scholar]
- Ferreira, R.L.C.; Prado, R.M.; Souza Junior, J.P.; Gratão, P.L.; Tezotto, T.; Cruz, F.J.R. Oxidative stress, nutritional disorders, and gas exchange in lettuce plants subjected to two selenium sources. J. Soil Sci. Plant Nutr. 2020, 20, 1215–1228. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Dean, R.T.; Davies, M.J. Radical chemistry of epigallocatechin gallate and its relevance to protein damage. Arch. Biochem. Biophys. 2003, 414, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; VanBreusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. BioEssays 2006, 28, 1091–1101. [Google Scholar] [CrossRef]
- Singh, O.P.; Usha, K.; Saboki, E.; Srivastav, M.; Dahuja, A.; Singh, B. Enzymatic reactive oxygen species (ROS) scavenging system in mango varieties resistant and susceptible to malformation. Sci. Hortic. 2012, 38, 81–89. [Google Scholar] [CrossRef]
- Oukarroum, A.; Bussotti, F.; Goltsev, V.; Kalaji, H.M. Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba L. plants under salt stress. Environ. Exp. Bot. 2015, 109, 80–88. [Google Scholar] [CrossRef]
- Ríos, J.J.; Blasco, B.; Leyva, R.; Sanchez-Rodriguez, E.; Rubio-Wilhelmi, M.M.; Romero, L.; Ruiz, J.M. Nutritional balance changes in lettuce plant grown under different doses and forms of Se. J. Plant Nutr. 2013, 36, 1344–1354. [Google Scholar] [CrossRef]
- Barros Júnior, A.P.; Cecílio Filho, A.B.; Rezende, B.L.; Pôrto, D.R.Q.; Prado, R.M. 2011. Nitrogen fertilization on intercropping of lettuce and rocket. Hortic. Bras. 2011, 29, 398–403. [Google Scholar] [CrossRef]
- Cecílio Filho, A.B.; Alves, A.U.; Galati, V.C.; Bezerra Neto, F.; Barbosa, J.C.; Machado, B.Q.V. Intercropping of eggplant and tomato as function of times of transplant and cropping season. Rev. Caatinga. 2022, 35, 276–287. [Google Scholar] [CrossRef]
- Barbosa, A.P.; Arieira, J.O.; Hora, R.C.; Silva, A.F.M.; Kondo, P.N.Y.; Silva, M.R.; Severino, J.J.; Silva, T.R.B. An agronomic and economic evaluation of lettuce cultivars intercropped with rocket over two cultivation seasons. Afr. J. Agric. Res. 2015, 1, 1083–1090. [Google Scholar] [CrossRef]
- Bezerra Neto, F.; Gomes, E.G.; Araújo, R.R.; Oliveira, E.Q.; Nunes, G.H.S.; Grangeiro, L.C.; Azevedo, C.M.S.B. Evaluation of yield advantage indexes in carrot-lettuce intercropping systems. Interciencia 2010, 35, 59–64. [Google Scholar]
- Yan, Y.; Gong, W.; Yang, W.; Wan, Y.; Chen, X.; Chen, Z.; Wang, L. Seed treatment with uniconazole powder improves soybean seedling growth under shading by corn in relay strip intercropping system. Plant Prod. Sci. 2010, 13, 367–374. [Google Scholar] [CrossRef]
- Cecílio Filho, A.B.; Rezende, B.L.A.; Barbosa, J.C.; Grangeiro, L.C. Agronomic efficiency of intercropping tomato and lettuce. An. Acad. Bras. Cienc. 2011, 83, 1109–1119. [Google Scholar] [CrossRef]
- Su, B.Y.; Song, Y.X.; Song, C.; Cui, L.; Yong, T.W.; Yang, W.Y. Growth and photosynthetic responses of soybean seedlings to maize shading in relay intercropping system in Southwest China. Photosynthetica 2014, 52, 332–340. [Google Scholar] [CrossRef]
- Carballo-Méndez, F.J.; Urrestarazu, M.; Rodríguez-Ortiz, J.C.; Morales, I. Electrical conductivity of the nutrient solution on the vegetative propagation of bell pepper and tomato. Cienc. Rural. 2023, 53, e20210730. [Google Scholar] [CrossRef]
- Sandhu, R.K.; Boyd, N.S.; Qiu, Q.; Guan, Z.; Licata, M. 2020. Optimization of planting dates of Jalapeno pepper (Capsicum annuum ‘Jalapeño’ L.) and cantaloupe (Cucumis melo var. cantalupensis Ser.) relay cropped with strawberry (Fragaria x ananassa Duchesne). PLoS ONE 2020, 15, e0236677. [Google Scholar] [CrossRef] [PubMed]
- Andrade Filho, F.C.; Oliveira, E.Q.; Lima, J.S.S.; Moreira, J.N.; Silva, I.N.; Lins, H.A.; Cecílio Filho, A.B.; Barras Júnior, A.P.; Bezerra Neto, F. Agro-economic viability from two croppings of broadleaf vegetables intercropped with beet fertilized with roostertree in different population densities. Revista Facultad Ciencias Agrarias UNCuyo 2019, 52, 210–222. [Google Scholar]
- Cecílio Filho, A.B.; Machado, B.Q.V.; Alves, A.U.; Pereira, B.J.; Guerra, N.M.; Bezerra Neto, F. 2022. Bio-agronomic efficiency indices of eggplant and tomato intercropping. Hortic. Bras. 2022, 40, 181–189. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC | Macronutrients (mM) | Micronutrients (µM) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dS m−1 | pH | NO3− | H2PO42− | SO42− | K+ | Ca2+ | Mg2+ | Fe | Mn | Cu | Zn | B | Mo |
| 2.00 a | 5.80 | 10.25 | 1.50 | 1.75 | 4.75 | 5.00 | 1.51 | 15 | 10 | 0.75 | 5 | 30 | 0.5 |
| N | P | K | Mg | Fe | |
|---|---|---|---|---|---|
| g kg−1 plant−1 | mg kg−1 plant−1 | ||||
| Monoculture (M) | |||||
| Lucy Brown | 36.5 a | 7.6 a | 97.3 a | 3.2 a | 600 a |
| Vanda | 38.4 a | 7.5 a | 101.0 a | 2.6 b | 433 b |
| p | 0.1835 | 0.6326 | 0.0927 | 0.0188 | 0.0377 |
| Intercropping (I) | |||||
| Lucy Brown | 33.3 b | 5.7 b | 102.3 b | 4.2 a | 666 a |
| Vanda | 39.5 a | 9.6 a | 109.6 a | 2.7 b | 333 b |
| p | 0.0091 | 0.0008 | 0.0315 | 0.0059 | 0.0171 |
| % ∆ I−M | |||||
| Lucy Brown | −9.49 b | −31.2 b | 4.8 a | 22.9 a | 7.75 a |
| Vanda | 4.3 a | 21.5 a | 7.9 a | 3.7 b | −30.9 b |
| p | 0.0183 | 0.0007 | 0.2528 | 0.0085 | 0.0210 |
| Crop System | Leaf Area | Dry Mass | Total Soluble Solids |
|---|---|---|---|
| m2 plant−1 | g plant−1 | °Brix | |
| Tomato monoculture | 1.45 a | 143.3 a | 4.73 a |
| Tomato intercropping Lucy Brown | 1.44 a | 133.8 a | 4.66 a |
| Tomato intercropping Vanda | 1.46 a | 150.6 a | 4.61 a |
| Crop System | G (60–70 mm) | M (50–60 mm) | MMM (<50 mm) | Total |
|---|---|---|---|---|
| Tomato monoculture | 3.84 a | 6.47 a | 2.76 a | 13.07 a |
| Tomato intercropping Lucy Brown | 2.76 a | 6.20 a | 3.12 a | 12.08 a |
| Tomato intercropping Vanda | 3.17 a | 7.28 a | 2.76 a | 13.21 a |
| Crop System | Net Revenue | Cost of Plant | Profit Total |
|---|---|---|---|
| Tomato monoculture | 6.07 | 0.76 | 5.31 |
| Tomato intercropping Lucy Brown | 5.34 | 0.76 | 4.58 |
| Tomato intercropping Vanda | 6.04 | 0.76 | 5.28 |
| Lucy Brown monoculture | 1.34 | 0.40 | 0.94 |
| Lucy Brown intercropping | 0.30 | 0.25 | 0.05 |
| Vanda monoculture | 0.87 | 0.60 | 0.27 |
| Vanda intercropping | 0.58 | 0.25 | 0.33 |
| Tomato monoculture | 6.07 | 0.76 | 5.31 |
| Tomato intercropping Lucy Brown | 5.64 | 1.01 | 4.63 |
| Tomato intercropping Vanda | 6.62 | 1.01 | 5.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cunha-Chiamolera, T.P.L.; Cecílio Filho, A.B.; Santos, D.M.M.; Chiamolera, F.M.; Guevara-González, R.G.; Nicola, S.; Urrestarazu, M. Lettuce in Monoculture or in Intercropping with Tomato Changes the Antioxidant Enzyme Activities, Nutrients and Growth of Lettuce. Horticulturae 2023, 9, 783. https://doi.org/10.3390/horticulturae9070783
Cunha-Chiamolera TPL, Cecílio Filho AB, Santos DMM, Chiamolera FM, Guevara-González RG, Nicola S, Urrestarazu M. Lettuce in Monoculture or in Intercropping with Tomato Changes the Antioxidant Enzyme Activities, Nutrients and Growth of Lettuce. Horticulturae. 2023; 9(7):783. https://doi.org/10.3390/horticulturae9070783
Chicago/Turabian StyleCunha-Chiamolera, Tatiana P. L., Arthur Bernardes Cecílio Filho, Durvalina M. M. Santos, Fernando M. Chiamolera, Ramón G. Guevara-González, Silvana Nicola, and Miguel Urrestarazu. 2023. "Lettuce in Monoculture or in Intercropping with Tomato Changes the Antioxidant Enzyme Activities, Nutrients and Growth of Lettuce" Horticulturae 9, no. 7: 783. https://doi.org/10.3390/horticulturae9070783
APA StyleCunha-Chiamolera, T. P. L., Cecílio Filho, A. B., Santos, D. M. M., Chiamolera, F. M., Guevara-González, R. G., Nicola, S., & Urrestarazu, M. (2023). Lettuce in Monoculture or in Intercropping with Tomato Changes the Antioxidant Enzyme Activities, Nutrients and Growth of Lettuce. Horticulturae, 9(7), 783. https://doi.org/10.3390/horticulturae9070783