Abstract
A prospection was carried out on Fuerteventura Island in order to explore the existing biodiversity of Vitis vinifera ssp. vinifera in almost desert conditions (<120 mm per year). For this purpose, 40 individuals were collected and genotyped with 20 SSRs. Nine known varieties and one unknown variety, named Majorera on the island, were identified. In addition, four new mutations were found in the varieties Listan negro and Listan prieto, respectively. Thirteen unknown individuals and five erratic accessions were identified. Seven new names are proposed for inclusion in the world databank (one main name (Majorera), one new synonym for Listan negro (Hoja moral), four new mutation names (Listan prieto de Antigua, Listan prieto de Vega, Hoja moral de El Rosario and Hoja moral de Betancuria) and a new synonymy for Muscat Hamburg (Moscatel), which is very widespread on the island). Finally, the possibility is raised that the new Majorera variety is not a pure vinifera but the product of an interspecific cross, as has happened with the variety found on the island of La Palma, Bienmesabe tinto. Once again, the Canary Archipelago shows itself to be a possible centre for the creation of biodiversity for the cultivated vine.
1. Introduction
One of the main problems affecting vine cultivation (one of the oldest and most economically important crops in the world) in the 21st century is climate change [1]. Vitis vinifera L. species, like all other living organisms, will suffer significant genetic erosion. However, this will not be the only erosion that vine has suffered throughout its history. Already in the 19th century, this plant species suffered the consequences of the Phylloxera plague, causing a high mortality of vines and the disappearance of most of its varieties worldwide (except for certain areas of the planet such as Crete, the Canary Islands, Chile…) [2]. During the 20th century, the problem lay in the excessive use of a small number of varieties, which came to be considered international (such as Cabernet sauvignon, Chardonnay, Pinot noir…) [1]. In the 21st century, the problem will be climate change in its main aspects: the increase in average temperatures during vegetative growth and in some regions, the frequency and severity of frosts during spring and autumn, very hot days during the summer, extreme rain or hail events and widespread spatial and temporal drought [3], so much so that, at the inauguration of the 43rd World Congress of Vine and Wine (Mexico, November 2022), its Director General stated: “…this year’s harvest was characterised by extreme heat and a record drought that accelerated ripening in vineyards all over the world… this was the worst drought in the last 500 years” [4]. What is most striking is that these seasonal (punctual) and devastating phenomena due to large-scale climatology, which has been defined as climatic variability, are centred in narrow areas of our planet, which happen to correspond mostly to wine-growing areas. Hannah et al.’s [5] predictions are not encouraging at all. They predict a gradual disappearance of the current wine-growing areas towards more northern areas of the planet, substantially changing the global landscape by 2050 and beyond. The effects of climate change on vines have already been observed for 50 years. Above all, what is being observed is an earlier phenological stage (budbreak, flowering, veraison and harvest) and shorter intervals between these stages, with the latter being shortened by 6 to 17 days, depending on the variety and location [1]. Thus, for the most widely planted varieties on our planet (known as international varieties), the production of a quality product from their harvest will be endangered and jeopardised.
Focusing on this problem analysis, it can be stated that adaptation responses to climate change in the world of wine can vary from those implemented at a winery level, which can be relatively easy, cheap and immediate, but which provide a low adaptation potential, or those implemented at a vineyard level, which can be more expensive and can lead to more difficulties, but which have a higher adaptation potential. Strategies that could be implemented at a vineyard level, in turn, are divided into two. There are those with medium-term effects and medium adaptation potential, based on vineyard management and aimed at mitigating radiation, leaf and bunch temperature and water deficit. There are also those with a high potential for adaptation to climate change effects; therefore, long-term effects will be based on changing the location of vineyards and/or replacing old varieties (often international varieties) with others that are much better adapted to the new changing conditions [6]. Obviously, the implementation of the latter will require a high level of investment and a change in legislation (especially at the level of appellation d’origine contrôlée (AOC)). For many vine growers, a change in vineyard location will be an impossible task, but it will be possible to opt for a change to different grapevine varieties in the same location. This would be a matter of using a new varietal range that has a wide temperature niche for vegetative growth and ripening, adapted to the new circumstances, and is also efficient in water management [1,7]. To this end, and as advised by Wolkovich et al. [1], current efforts are being made to study both the intervarietal and intra-varietal variability of the current marketed crops and, among them, Vitis vinifera L.
This study’s main objective is to prospect areas that are much drier than normal for a wine-growing area, with very low rainfall, to locate high-temperature and drought-resistant vines. In Spain, there are territories where these conditions are found, as is the case of the Canary Islands (Figure 1). The Canary Islands are one of the five archipelagos of Macaronesia, a group of volcanic islands located in the Atlantic Ocean. The easternmost islands of this archipelago have desert conditions, as they are only a few kilometres from the Sahara Desert, as is the case of Fuerteventura, which is only 97 km away [8].
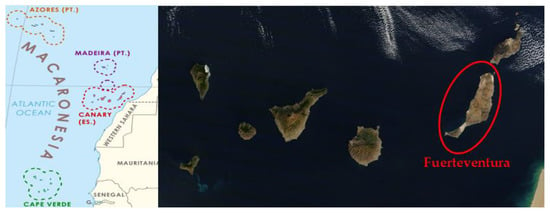
Figure 1.
(Left) Macaronesia map [9]. (Right) Canary Archipelago and Fuerteventura Island [10].
Fuerteventura is an island situated at the coordinates 28°24′ N 14°00′ W and is the archipelago’s second-largest island in terms of area [11]. According to the Koppen climate classification, the climate is hot desert (BWh) in most of Fuerteventura Island, with very low rainfall that is always below 120 mm per year. Due to the island’s low altitude (between 0 and 813 m), this island does not retain humid air masses, as is the case on other archipelago islands [12]. Fuerteventura is the Canary Island with the smallest surface area dedicated to vine cultivation (about 10 ha) and is the only island without an AOC [13]. Paradoxically, the first written document in which the first wine was produced in the Canary Islands was in Fuerteventura [14].
This scarcity of vine cultivation has been maintained throughout history by the presence of gavias (Figure 2). Gavias are structures built by Fuerteventura inhabitants to retain and make the most of the scarce precipitation water that falls on the island. Thus, the walls and surroundings of these structures are where most of the vines are found, planted with the aim of providing consistency to the structure’s containment dam [15].

Figure 2.
Detail of a gavia on the Fuerteventura Island [15].
Thus, and given all the above, the aim of this research was to carry out a prospection of this arid island, which has never suffered from the Phylloxera plague (like the rest of the Canary Islands), in order to identify the different individuals currently forgotten in the gavias or in old abandoned plantations. At the same time, a study of the island’s vine varieties names was also carried out, as well as, whenever possible, a study of the population structure.
2. Materials and Methods
2.1. Plant Material
Forty vine samples from different locations on Fuerteventura Island were analysed (woody tissue). Prospection was carried out by vine growers and hunters by means of a mass selection process. Samples were collected either in the bush (barren land) or on the slopes of gavias. Once collected, they were stored at −20 °C until analysis. Detailed information about the accessions analysed is given in Table S1.
2.2. DNA Extraction and Purification
DNA extraction was carried out using a protocol developed by the Tecnología Enológica (TECNENOL) research group [16,17]. It is an adaptation of Fort et al.’s [18] protocol, specific for RNA procurement. It was also improved by introducing polyvinylpyrrolidone (PVP) into the extraction buffer and adding another chloroform wash. The NanoDrop TM 1000 Spectrophotometer (Thermo Fisher® Scientific, Waltham, MA, USA) was used to determine the extraction yield and purity of the extracted DNA.
2.3. Microsatellites
The 20 SSRs used for the genotyping of the samples from Fuerteventura Island were previously chosen for their discrimination capacity and polymorphism. Of all of them, 7 match (Table S2) with the 9 that were proposed by the international community as reference genetic markers (VVS2, VVMD5, VVMD7, VVMD25, VVMD27, VVMD28, VVMD32, VrZAG62, VrZAG79) [19]. SSR “kit”, with which the TECNENOL group has been working for several years, comprises: VVS2, VVS3, VVS29 [20]; VVMD5, VVMD6, VVMD7 [21]; VVMD27, VVMD28, VVMD36 [22]; VrZAG21, VrZAG47, VrZAG62, VrZAG64, VrZAG79, VrZAG83 [23]; scu06vv [24]; VvUCH11, VvUCH12, VvUCH19 [25]; VChr19a [26]. Microsatellites VrZAG47 and VVMD27 are not independent loci, i.e., they amplify the same area of the genome. The difference between the two is the design of their primers [27].
2.4. Polymerase Chain Reaction (PCR)
In order to carry out the PCR, an Applied Biosystems 2720 Thermal Cycler (Foster City, CA, USA) was used. Amplification was carried out from 4 ng of DNA and 1 μM of each primer with a fluorescent dye attached to the primer Forward (Fw) (6-FAM: VVS3, VVMD7, VVMD28, VVMD36, VrZAG47, VrZAG62, VrZAG83, VvUCH11 y VvUCH19; HEX: VVS2, VVS29, VVMD6, VVMD27, VrZAG21, VrZAG79 y VChr19a; NED: VVMD5, VrZAG64, scu06vv, VvUCH12) by using AmpliTaq DNA Polymerase kit (Applied Biosystems, Foster City, CA, USA). Thermocycling conditions were: a first stage at 95 °C for 5 min, a second phase with 40 cycles at: 95 °C for 45 s; the annealing temperature for each SSR (48.1 °C, 50 °C, 52 °C, 54.4 °C, 56.1 °C, 56.6 °C and 58 °C (Table S3)) for 30 s; 72 °C for 1 min and a half; and a last stage at 72 °C for 7 min.
2.5. Measurement of Amplified Fragment Lengths
In addition to the PCR product, formamide and the internal marker GeneScan ROXTM 500 (Applied Biosystems, Foster City, CA, USA) was used to prepare the plates for measuring the length of the amplified fragments. Furthermore, these plates were subjected to thermal denaturation (3 min at 95 °C). Fragment separation was performed by capillary electrophoresis with an ABI PRISM 3730® genetic analyser (Applied Biosystems, Foster City, CA, USA). Finally, Peak Scanner Software (Applied Biosystems, NJ, USA) was used to interpret the electrophenograms.
2.6. Data Analysis
GenAlEx 6.5 software [28,29] was used to assess the reliability of the chosen SSR kit. For this purpose, the statistics Na (number of different alleles), Ne (number of effective alleles. These are the alleles that are transmitted to the next generation), Ho (observed heterozygosity. These are the heterozygotes computed), He (expected heterozygosity. Estimation of the heterozygotes that the population under study might have. Also known as diversity index), F (fixation index. Parameter measuring the goodness of homozygosity) and PI (probability of identity. Probability that two molecular profiles (MP-SSR) with the same SSR values are the same variety) were used. Furthermore, the same software was used to identify MP-SSRs identical to each other (data normalisation) and, in addition, the assignment test was performed. This last strategy is based on the allele frequency of each entry and calculates a log probability value of this entry for each subpopulation using the allele frequencies of the respective subpopulations. Thus, an individual is assigned to the subpopulation with the highest log likelihood value [30]. To examine the genetic relationships between the populations found, two-dimensional principal coordinate analysis (PCoA) was used, both for populations and for populations of individuals based on the standardised covariance of the genetic distances calculated for the co-dominant markers. Finally, the value of Fst, the coefficient of genetic differentiation between populations, was calculated assuming the infinite allele model.
For the three-dimensional representation of PCoA, the Python Data programme [31] was used, applying the Matplotlib strategy. The programme MEGA version 7 [32] was used for the phylogenetic trees and circular dendrograms (neighbour-joining method [33]).
The assessment of the population structure, which entailed the best combination of populations and the identification of admixed individuals, was carried out using the Structure 2.3. [34,35] programme. This model-based software uses a Bayesian clustering method in which several ancestral populations (K) are assumed to be present, each characterised by a set of allele frequencies at each locus. Individuals in the sample are assigned to populations (clusters), or conjointly to more populations if their genotypes indicate that they are admixed. All loci are assumed to be independent and each population K is assumed to follow Hardy–Weinberg equilibrium. Posterior probabilities were estimated using the Markov chain Monte Carlo (MCMC) method. MCMC chains were run with a 100,000 burn-in period followed by 1,000,000 iterations using a model allowing for admixture and correlated allele frequencies. Structure was run at least ten times by setting K from 1 to 7 (1 to 9 for global varieties), and an average likelihood value, L (K), was calculated across all runs for each K. The mean log probability of the data for each K was calculated to determine the most appropriate number of clusters, and the value of K for which this probability was highest was selected. The ∆K was then calculated using the method proposed by Evanno et al. [36]. ΔK is a quantity based on the rate of change in the log probability of the data between successive K values.
As has been usual in previous studies carried out by the same research group, the 40 MP-SSR obtained were compared with their extensive database [37,38,39,40,41] in order to incorporate new individuals and new mutations, if any. In addition, the existing literature on Canary Island varieties was reviewed in depth [39,40,41,42,43], as well as the main database worldwide, Vitis International Variety Catalog (VIVC) [44]). This exhaustive search for information allowed us not only to cross-reference the MP-SSRs but also to verify the accuracy of the names registered as names of origin. Thus, the original information provided by the winegrowers, the conclusive information of each accession, the percentage of similarity with a certain sample from the TECNENOL database and the SSRs in which variability has been detected, among other characteristics, are shown in Table S1. Additionally, information on the unique MP-SSRs for this population and the values found for 7 reference SSRs can be found in Table S2.
3. Results
Only 40 individuals were collected from the prospection on the island of Fuerteventura, which has no viticultural tradition, and these were analysed with TECNENOL’s own 20 SSRs.
3.1. SSR Polymorphism
The first objective was to find the accessions whose MP-SSRs were identical for all SSRs. We found 25 MP-SSRs identical to other profiles in the population or to each other. Thus, the population of unique profiles in Fuerteventura Island was 15 individuals (Table S2).
Table S4 presents the values for the main statistics that allowed us to affirm whether the chosen SSR kit was effective and efficient for characterising and identifying the Fuerteventura Island grapevine population. The total number of alleles found for this population was 134 (Na). The mean was 6.7 alleles per SSR, with SSRs VVS3, UCH19 and VVS29 showing the fewest alleles (2, 2 and 3, respectively) and SSRs VVMD28 and ZAG79 showing the most alleles (10 each). The mean number of alleles passing to the next generation (Ne) was 4.1, with SSRs UCH19 and VVS29 showing the lowest values (1.14 and 1.23, respectively), and SSRs VVMD36 and ZAG47 showing the highest values (6.62 and 6.82, respectively). The mean value of observed heterozygosity (Ho = 0.807) was significantly higher than the mean value of expected heterozygosity, also known as the genetic diversity index (He = 0.689). The lower values corresponded to SSRs UCH19 and VVS29 (Ho = 0.13 and 0.2, respectively; He = 0.12 and 0.18, respectively) and the higher values corresponded, for Ho, to SSRs VVS2, ZAG62, ZAG79 and VChr19a (Ho = 1) and, for He, to SSRs ZAG47 and VVMD36 (with a value of 0.85). Only two SSRs showed positive values for the F index. These were SSR ZAG83, with a value very close to 0 (0.015), and VVMD6, with a larger value (0.11). Finally, it should be noted that the accumulative identity probability for the population of 15 vines on the island of Fuerteventura was 7.5 × 10−20, with SSR VVMD36 having the lowest value (3.9 × 10−2) and SSR UCH19 having the highest value (7.7 × 10−1).
3.2. Grapevine Variety Analysis
Table S1 shows that the 40 accessions analysed corresponded to nine identified varieties and 1 corresponded to a new variety. All of them had a Spanish origin except for two: the Greek variety Muscat from Alexandria and the English variety Muscat Hamburg. Furthermore, within the group of Spanish varieties, it was found that Listan negro and Torrontes volcanico varieties are local varieties of the Canary Archipelago.
The grapevine variety with the most representatives found in the prospection of Fuerteventura Island was Listan prieto, with 18 accessions. In this group, three different MP-SSRs were described: the MP-SSR coinciding with the most extended in the TECNENOL database, which corresponded to four individuals; a mutated MP-SSR in the first allele of the SSR VVMD28 (VVMD28-1), which included 11 components (with the representative that entered with the name Listan prieto de Antigua (the new name for this mutation)); and four samples with two mutations (VVS3-2 and VVMD28-1) whose representative was entered under the name Listan prieto de Vega (the new name for this mutation).
The local Canary Islands variety Listan negro group contained seven entries, and four different MP-SSRs were identified in this grouping. In Fuerteventura Island, this variety is known as Hoja moral (new synonymy). Of these four different MP-SSRs, four accessions corresponded to the most widespread MP-SSR and were registered under the name of the new synonymy of Hoja moral: one MP-SSR was registered under the name of Hoja moral de El Rosario, which mutated in VVUCH12-2; one sample was registered under the name of Hoja moral de Betancuria, which presented variation in VVZAG21-1; and, finally, another sample was entered under the name of Hoja moral, which presented a mutation in VVS3-2.
The MP-SSR of the Greek variety Muscat of Alexandria was also identified in five accessions from Fuerteventura Island. All the components were found to be identical to the most widespread MP-SSR. Three of them were registered under the name Moscatel, the synonymy registered in the VIVC for this variety, and the other two individuals were entered as unknown.
The Spanish white grapevine variety, Airén, was represented by three accessions registered with names such as unknown and erroneous names Moscatel and Listan prieto, and presented a single MP-SSR identical to the most widespread. The Andalusian variety Palomino fino was represented in this collection by two accessions mutated in VVS3-1 and entries with the erroneous name Listan prieto. The MP-SSRs with only one representative were the Andalusian-Extremeña (SW Spain) grapevine variety Beba, which entered under the term Burra blanca, the Andalusian Mollar cano, which was registered as unknown, the English Muscat Hamburg, which entered under the name Moscatel, the Canarian Torrontes volcanico, entered as unknown and the new variety, registered under the name Majorera, which corresponded to a new MP-SSR.
3.3. Fuerteventura Grapevine Population Genetic Structure
From the 15 unique profiles found in Fuerteventura Island (Table S2), another data normalisation was performed in order to initiate a population structure study. Mutant individuals for a given variety were eliminated from this collection. Thus, the population under study had 10 members, corresponding to the 10 varieties described above (Tables S1 and S2). All of them belonged to the most extended profiles, except for the Palomino fino variety, whose two representatives had the same mutation (97.5% similarity), so a mutated individual had to be taken as a representative.
The use of the Structure 2.3. programme to find the best distribution of this 10-member collection in different populations was impossible due to the low number of individuals that made up the collection. For this reason, a principal coordinate analysis (PCoA) representation was performed directly to observe the position taken by these individuals according to their genetic distances (Figure 3).
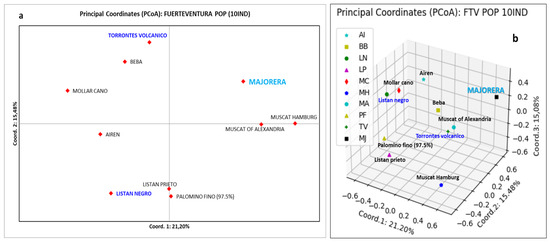
Figure 3.
Fuerteventura Island population (unique and non-redundant molecular profiles). (a) Two-dimensional representation using PCoA. (b) Three-dimensional representation using PCoA. The names in navy blue correspond to local varieties of the Canary Islands. The name in light blue corresponds to the new local variety. The names in black correspond to foreign varieties of the archipelago.
Figure 3a shows how coordinate 1 with a 21.2% goodness of fit separates the Majorera (Canary Islands), Muscat of Alexandria (Greek) and Muscat Hamburg (English) varieties from the rest, with Palomino fino and Listan prieto being very close to the axis or above it, respectively. Coordinate 2 (15.4% goodness of fit), on the other hand, places in the upper quadrants the Spanish varieties Mollar cano, Beba, Torrontes volcánico (from Lanzarote Island) and the new variety Majorera (from Fuerteventura Island), while the two Muscat varieties are above the axis. The new Majorera grapevine variety is practically located alone in the upper right quadrant. Figure 3b shows a three-dimensional representation using PCoA, with a distribution very similar to the previous one. Once again, the Majorera variety stands out from the rest of the individuals from the Fuerteventura Island collection. In this image, it can also be seen how the English grapevine variety Muscat Hamburg is significantly different from both the Majorera variety and the rest.
3.4. Majorera Variety Relation with Respect to Canary Archipelago Grapevine Population
As a strategy to confirm the uniqueness of the Majorera variety MP-SSR, a study in which this variety was related to the rest of the varieties of the Canary Islands was carried out.
The Structure 2.3. programme was used to find the best ancestral population distribution from 1 to 7. Figure S1 shows the results of this study, with K = 6 being the best distribution, followed at a very short distance by K = 4. The graphs generated by the Structure 2.3. programme for these distributions are presented in Figure S2. It can be seen in this figure, as for K = 6, that there is no clear distinction between the ancestral populations POP1 and POP2. On the other hand, the definition of the four ancestral populations for K = 4 is clear and sharp. The result observed for K = 6 in Figure S2 is confirmed numerically in Table S5. This table shows in detail the membership of each of the 32 individuals of the Canary Archipelago population to one of the six ancestral populations defined in Structure 2.3. The results show that POP1 and POP2 are not well defined, their individuals all being admixed (q < 85%), but, in addition, POP3 also has all its individuals admixed. This fact makes it impossible to continue the study using the six-population distribution. For this reason, the K = 4 distribution was used, as it had given a result very slightly different from the previous one (Figure S1), and, in addition, Figure S2 shows the graph of this distribution with its four well-defined ancestral populations. The numerical results supporting Figure S2 are presented in Figure S3 and graphically represented in Figure 4. For K = 4, the 32 Canarian varieties are grouped into four populations, each with pure individuals (q ≥ 85%) and admixed (q < 85%). In POP1, 34% of individuals (11 varieties), 4 pure individuals (36%) and 7 admixed (64%), are assigned. It is a very diverse population. The first pure member of POP1 is the red Bienmesabe tinto variety (rest of the Canary Islands (IC)), followed by the Majorera variety (Fuerteventura Island (FTV)) and the Lanzarote Island (LNZ) varieties Blanca de la Granja del Cabildo and Breval negro. The seven crossbred varieties are from all the islands. POP2, which has 16% of the total population (five varieties), with three pure individuals (60%) and two admixed individuals (40%), is a pure population from El Hierro island (HI). With one more individual than POP2 (19%), the third population (POP3) is presented with five pure individuals (83%) and one admixed (17%). This population is made up of individuals from HI and individuals from LNZ. POP4 is another large population with 10 members (31%), of which 3 will be pure (30%) and the remaining 7 admixed (70%). This population will also be characterised by being very diverse but with a large presence of pure IC individuals.
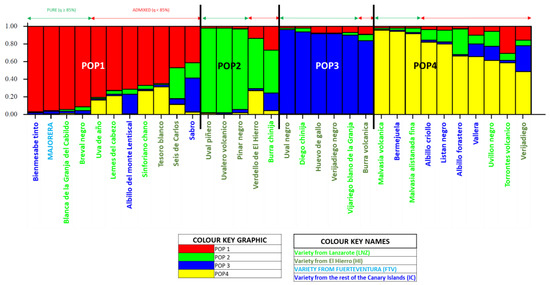
Figure 4.
Population of Canary Islands varieties (unique and non-redundant molecular profiles). Diagram Structure 3.2. 4K distribution for pure and admixed individuals.
Normalising the data, i.e., leaving out the admixed varieties (17 varieties), the two- and three-dimensional PCoA graphical representations of the 15 pure individuals were carried out, as well as a phylogenetic tree representation (Figure 5).
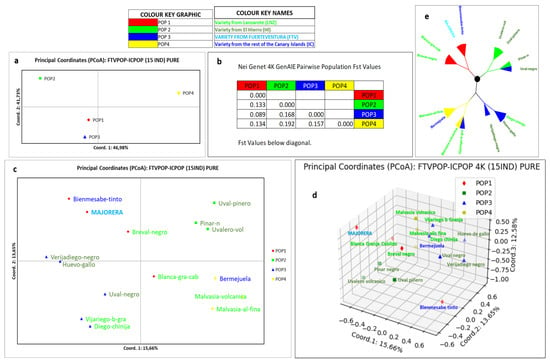
Figure 5.
Population of Canary Islands grapevine varieties (15 pure varieties). (a) Two-dimensional representation of the 4 Canarian populations by population. (b) Values of the Fst statistic for each population. (c) Two-dimensional representation of the 4 populations by individuals. (d) Three-dimensional representation of the 4 populations by individuals. (e) Phylogenetic tree of the 4 populations by individuals. The names in navy blue correspond to Canary Islands varieties, the name in light blue corresponds to the Fuerteventura Island variety, the names in acid green correspond to Lanzarote Island varieties and the names in dark green correspond to El Hierro island varieties.
Figure 5a shows the two-dimensional distribution of the four ancestral populations that have generated the Canary Islands population, with a 93% goodness of fit (assignment test). Coordinate 1 (46.98%) separates POP4 from the rest, while coordinate 2 (41.7%) places POP4 and POP2 in the upper quadrants. Overall, POP2, POP3 and POP4 are distributed far apart and equidistant while POP1 is positioned close to POP3. The Fst parameter confirms this arrangement numerically (Figure 5b). Figure 5c presents the two-dimensional representation of PCoA, but now by individuals. It can be seen how the location of the populations broken down by individuals has redistributed the four populations, causing POP2 to move to the right quadrants. It can be seen that the Majorera variety is positioned next to the red Bienmesabe tinto variety. These two varieties, together with the Breval negro variety, are located alone in the upper left quadrant. In fact, in this representation, each population occupies a different quadrant. Figure 5d shows the three-dimensional plot of the 15 Canarian individuals using PCoA. In this representation, with one more dimension, the Majorera variety behaviour is again significantly different from the rest of the collection. Another variety that is also very distant from the rest is Bienmesabe tinto. Both belong to POP1 together with Breval negro and the variety Blanca de la Granja del Cabildo. Finally, the global Canary Islands population is shown in Figure 5e, represented by a phylogenetic tree. It shows the phylogenetic relationship between the four populations, with three main branches from which POP1, POP2 and, from the third branch, POP3 and POP4 derive.
3.5. Majorera Variety Relation with Respect to World Population
In this section, the Majorera variety was compared with 308 vinifera varieties from 22 countries around the world (TECNENOL database [37,38,39,40,41]). To perform this, two strategies were carried out: one purely genetic, and a second in which a geographical parameter was introduced.
As far as the first strategy is concerned, the genetic structure was studied using Structure 2.3. The population now consists of 309 individuals to be distributed according to the best value of K (from 1 to 9). In this case, the best distribution was for K = 2 (Figure S4) followed by K = 7. The vinifera world population was divided into two ancestral populations, POP1 and POP2, with 182 (59%) and 127 (41%) members, respectively (Figure 6a and Figure S5). Specifically, in Figure S5, the q values for each variety for a given population are presented. Thus, POP1 has 88% pure individuals (161 individuals out of a total population of 309) and 12% admixed (21/309). This is a population with an abundance of representatives from Italy (51 pure individuals and 4 admixed out of a total of 72 Italians), France (39 pure and 5 admixed out of 49), Spain (24 pure and 4 admixed out of 106), Portugal (12 pure and 4 admixed out of 22) and Greece (11 pure out of 14), among others. On the other hand, POP2, with 98 pure individuals (77%) and 29 admixed (23%), hosts individuals from Spain (67 pure and 11 admixed out of 106 individuals), Italy (10 pure and 7 admixed out of 72), France (4 pure and 1 admixed out of 49), Portugal (2 pure and 4 admixed out of 22) and Greece (3 pure out of 14), among others. The IC population (with 32 varieties) is entirely positioned in POP2, with 30 pure and 2 admixed individuals (Albillo criollo and Malvasia alistanada fina) (Figure S5). From these results, we eliminated 50 varieties with a q value lower than 85%.
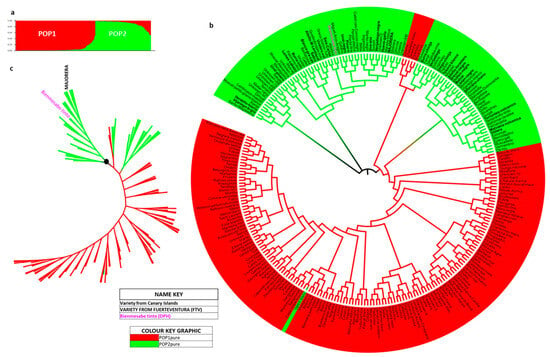
Figure 6.
The world population (259 individuals) distributed into 2 populations. (a) Graphical representation of K = 2 according to Structure 2.3. (with pure and admixed individuals); (b) neighbour-joining circular dendrogram of the 259 world population pure individuals, highlighting the location of Bienmesabe tinto (pink and bold), Majorera (capital and bold) and IC varieties (bold). (c) Phylogenetic tree corresponding to the population of 259 pure individuals, highlighting the Bienmesabe tinto and Majorera varieties.
Figure 6b shows the circular dendrogram of this K = 2 distribution for the 259 pure varieties after the application of an assignment test that gave a goodness of fit equal to 100%. Interestingly, four varieties assigned to POP1 were incorporated into POP2 (Chasselas blanc (FRA), Tenerone (ITA), Parellada (ESP) and Rojal tinta (ESP)), and the POP2 variety Valent blanc (ESP) was incorporated into POP1. Otherwise, the distribution is as expected according to the Structure 2.3. programme and the GenAlEx 6.5 assignment test. The varieties Bienmesabe tinto and Majorera are located in the second major branch from the origin (Figure 6b,c) and in different clusters. The IC grapevine varieties are distributed over almost all branches and arms of POP2.
The 259 individuals distribution by PCoA representation is shown in Figure 7. To highlight the role of the IC population and new Majorera variety, a new population (POP3) corresponding to IC was constructed and extracted from POP2. Applying the assignment test, it gave a goodness of fit of 93% (data not shown). Figure 7(a.2,a.3) show the distribution of these three populations by population. It can be seen how coordinate 1 (76.65%) separates the populations of Spanish origin (POP2 and POP3 (IC)) from POP1, which corresponds to the clustering of most of the varieties from the rest of the world. In turn, coordinate 2 (23.35%) isolates POP1 and POP3 (IC) in the upper quadrants. The statistical parameter (Fst) supporting this positioning is shown in Figure 7(a.3). The two-dimensional distribution by individuals (Figure 7(a.1)) shows how coordinate 1 (5.68%) again separates POP2 and POP3 (IC) from POP1, but now coordinate 2 (3.68%) leaving the individuals of all the populations distributed in the upper and lower quadrants indistinctly. Bienmesabe tinto and Majorera variety locations overlap with the populations of POP2 and POP3 (IC). The three-dimensional representation (Figure 7b) shows broadly the distribution described for Figure 7(a.1) except that the varieties Bienmesabe tinto and Majorera are separated from the rest of the varieties of both POP2 and POP3 (IC).
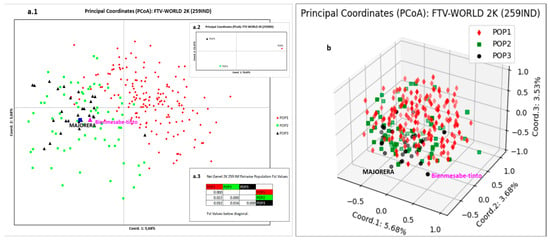
Figure 7.
PCoA representation of Canary Islands population and the wold varieties normalised for K = 2. (a.1) Two-dimensional representation of the 3 populations by individuals, (a.2) two-dimensional representation of the 3 populations by population, (a.3) Fst statistic values for each population. (b) Three-dimensional representation of the 3 populations by individuals. POP3 corresponds to IC.
As for the distribution using a geographical component, in this case, the 309 varieties were grouped geographically, 307 belonging to Vitis vinifera ssp. vinifera, the Bienmesabe tinto variety and the Majorera variety. Five populations were formed: EASTMED-CAU (Algeria, Cyprus, Georgia, Israel, Lebanon, Tunisia and Turkey), BP (Bosnia and Herzegovina, Bulgaria, Croatia, Greece, Serbia, Slovenia and Montenegro), ITA (Italy), FRA-CEU (Austria, France, Germany, Hungary and Switzerland) and IP (Spain and Portugal). The Canary Islands population was extracted from the latter (IC).
Using the GenAlEx 6.5 programme, an assignment test was carried out in order to find out which individuals were poorly placed in each group (admixed), and to be able to eliminate them (data normalisation). The 309 individuals’ goodness of distribution was 61% (data not shown). The final population of well-placed individuals with a goodness of fit of 91% (data not shown) and spread over six geographic areas dropped to 188 varieties.
Figure S6 presents the characteristics of each geographical population. Thus, EASTMED-CAU, with 4% of the population (12 individuals), presented four pure individuals (33%), with a Turkish variety and the rest of the unknown ones that were previously assigned to this grouping, as well as eight admixed (67%) corresponding to the rest of this cluster nationalities. BP, with 9% of the population (28 varieties), showed 50% of pure individuals, with Greek and Croatian varieties dominating, and 50% of admixed varieties coming mostly from Greece. The Italian cluster (ITA), with 24% of the world population (73 samples), was also almost equally distributed, with 51% of pure varieties (37 individuals) and 49% of admixed (36 individuals). The French-European cluster, with 60 varieties, represented 24% of the world population, with 72% pure (43 varieties) and 28% mixed (17 individuals), with French varieties dominating in both cases. The most numerous grouping corresponded to IP, with 34% of the components of the world population (104 individuals), divided between 63% of pure individuals (66 varieties) and 37% of admixed (38 samples). In this case, it should be noted that, with Spanish varieties dominating in both cases (pure and mixed), a proportionally greater crossbreeding was observed in Portuguese varieties. And, lastly, IC, corresponded to 10% of the total, with 24 pure varieties (75%) and 8 mixed varieties (25%).
Figure 8 shows the circular dendrogram and the phylogenetic tree of this population of pure varieties. Both representations show the 188 varieties distributed in three main branches. The first one corresponds to the EASTMED-CAU, BP and ITA populations, with two small independent IP arms. The second corresponds to the IP population itself, with the Canary Islands population and, diluted in this population, Bienmesabe tinto and Majorera varieties. Finally, in the third large branch, the FRA-CEU population is present, and, in the same way as in the first large branch, there are two small independent branches with IP varieties. Bienmesabe tinto and Majorera varieties are found in the same branch as the IC varieties but in different clusters.
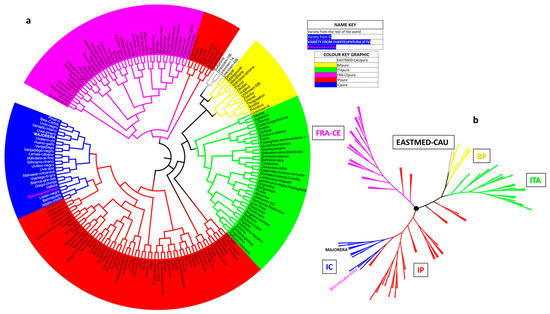
Figure 8.
World population (188 individuals) distributed in populations corresponding to six geographical areas. (a) Circular neighbour-joining dendrogram of the 188 pure individuals of the world population, highlighting the location of the Bienmesabe tinto and Majorera varieties; (b) phylogenetic tree of the distribution of these 6 populations with all their individuals.
Figure 9 corresponds to the graphical representations using two-dimensional PCoA with populations and with individuals (Figure 9a), and three-dimensional PCoA with individuals (Figure 9b). In the two-dimensional representation by populations (Figure 9(a.2)), it can be seen how coordinate 1 (42.02%) separates the IP and IC populations from the rest, with these being located in the right quadrants. The IC population is significantly differentiated from the rest, occupying a solitary quadrant, in the same way as the ITA and IP populations. Coordinate 2 (18.09%) separates the most singular populations in the upper quadrants, these being the most distant, ITA and IC. The Fst statistic confirms the arrangement of each population in the corresponding quadrants (Figure 9(a.3)). The two-dimensional individual distribution looks somewhat similar to that of populations (Figure 9(a.1)). Coordinate 1 (6.03%) clearly separates the Spanish populations (IP and IC) from those of ITA and FRA-CE, while the populations of EASTMED-CAU and BP are distributed along the central part of the two coordinate axes. Coordinate 2 (3.98%) has no effect on these populations. Bienmesabe tinto and Majorera varieties are located overlapping with POP2 and IC. Again, the three-dimensional representation shows a significant differentiation of the Bienmesabe tinto and Majorera varieties with respect to the rest of the varieties worldwide (Figure 9b).
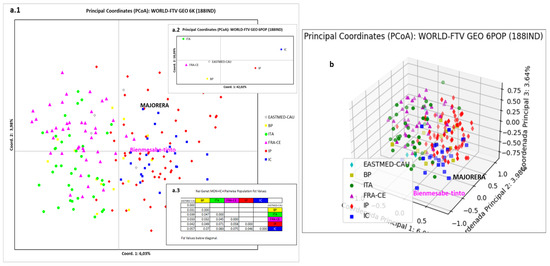
Figure 9.
PCoA representation of the Canary Islands population and of the world varieties by geographical criterion. (a.1) Two-dimensional representation of the 6 populations by individuals, (a.2) two-dimensional representation of the 6 populations by population, (a.3) values of the Fst statistic for each population. (b) Three-dimensional representation of the 6 populations by individuals.
4. Discussion
After showing the results obtained in the Fuerteventura Island prospection, in this section, the singularity and possible origin of the new MP-SSR found is going to be justified, among other aspects.
4.1. SSR Polymorphism
As usual in TECNENOL studies, the 20 SSRs used to genotype the samples in question proved to be efficient and effective. In this sample population from Fuerteventura Island, the SSRs with the best performance were the ZAG79 (Na: 10; Ho: 1; He: 0.833; F: −0.2; PI: 4.8 × 10−2), VVMD27 (Na: 9; Ho: 0.933; He: 0.836; F: −0.117; PI: 4.7 × 10−2), ZAG47 (Na: 9; Ho: 0.933; He: 0.849; F: −0.099; PI: 4.0 × 10−2) and VVMD36 (Na: 8; Ho: 0.933; He: 0.853; F: −0.094; PI: 3.9 × 10−2). But, there were also SSRs that did not meet the desired expectations, such as UCH19 (Na: 2; Ho: 0.133; He: 0.124; F: −0.071; PI: 7.7 × 10−1), VVS29 (Na: 3; Ho: 0.2; He: 0.184; F: −0.084; PI: 6.7 × 10−1) and VVS3 (Na: 2; Ho: 0.733; He: 0.491; F: −0.493; PI: 3.8 × 10−1). However, all SSR joint actions did provide very satisfactory results.
Before pointing out the statistical parameters that give confidence in the chosen SSRs, it should be borne in mind that the values found for each population depend on three conditioning factors: (a) the number of SSRs used, (b) the number of samples analysed and (c) the proximity of the population samples to be analysed, which is why it is difficult to compare with other authors [45,46,47,48,49,50,51,52]. It should be noted that the sample under study is a population belonging to an almost desert island where viticulture has been lost over the years, which is why only 40 samples have been collected. This number is similar to the number of individuals analysed in the study on Andalusian grape varieties carried out by Jiménez-Cantizano et al. [52] (49 samples), which also used 20 SSRs, so it is a good basis for comparison. In this case, 15 MP-SSRs were found out of the 40 individuals prospected, and 30 different genotypes were obtained in the Andalusian varieties study. From these data, when contrasting the coincident statistical parameters to compare the goodness of the SSRs used (Table S4), it is observed that all the parameter values of this research show significantly lower results, with the exception of Ho. While the mean Ho value of the present work was 0.803 (80.3% of heterozygotes), in the Andalusian varieties study, it was 0.763 (76.3% of heterozygotes); thus, the Fuerteventura Island population had fewer homozygous individuals (almost 5% fewer). On the other hand, this study’s accumulative IP reached the value of 7,5 × 10−20, 44.4% lower than the value obtained in the work on Andalusian varieties (1.7 × 10−19) for 20 SSRs in both cases. With respect to the other works consulted [45,46,47,48,49,50,51], the values were always significantly lower (except for PI) and uneven, mainly due to the low number of samples analysed and not due to a loss of efficiency and efficacy of the SSRs used.
4.2. Grapevine Variety Analysis
From the 40 accessions registered for the Fuerteventura Island population, once the first data normalisation was carried out, the Fuerteventura population was reduced to 15 individuals with a unique MP-SSR. These corresponded to 10 varieties, of which 9 were known and registered in the databases (Airén, Beba, Listan negro, Listan prieto, Mollar cano, Muscat Hamburg, Muscat of Alexandria, Palomino fino and Torrontes volcánico) and 1 was unknown (Majorera). The difference in five individuals was due to the existence of intra-varietal variability, i.e., four new mutations and one mutation already described in the Lanzarote Island study (whose name is Listan negro santanero) [40] had been described for known varieties in this collection. Apart from this, it must be said that the Palomino fino variety (in this work) was also represented by a mutant described in two previous studies since the population of Fuerteventura Island did not have the most widespread profile for this variety. This mutant genetic profile was found on the island of Lanzarote, where it received the name Listan blanca chicharrera (published name) [40], which has also been found in el Hierro island [41]. Another case of mutation described previously occurred in the Andalusian variety Mollar cano. This variety was represented by a mutation found on the Island of Lanzarote [40] known as Mollar bonilla.
Overall, in the Fuerteventura population characterisation and identification study, 13 samples were identified that were entered with the name unknown and 5 accessions with erratic names were detected, which have been perfectly identified. With regard to lexicography, it is suggested that the term Majorera be included in the VIVC database as the main name of the new variety in this collection. It is also proposed to include the new synonymy Hoja moral, with which the Listan negro variety is known throughout the island, as well as the names of the mutations found for this variety and also for the Listan prieto variety (Listán prieto de Antigua, Listan prieto de Vega, Hoja moral de El Rosario and Hoja moral de Betancuria), since, as has been pointed out in previous research articles [40,41], the VIVC database gives proper names to mutations of the Pinot varieties noir, Chasselas blanc and Trousseau noir, among others. The term Moscatel (synonymy registered to designate the Muscat of Alexandria variety and well used by the Fuertevetura island winegrowers) is detected to name the red variety Muscat Hamburg and is proposed as a new synonymy for this English red Muscat designed by Mr. Seward Snow [44]. A very unusual case occurs with the name of the accession Beba. It entered under the name of Burra blanca, a synonymy registered for the Airen variety and often used in the Canary archipelago to designate the Beba variety (a case also found in the study of El Hierro island [41]). For this reason, it is considered an erratic name and not a synonymy, since, if it were considered a synonymy, we would be creating a case of homonymy.
4.3. Fuerteventura Grapevine Population Genetic Structure
The aim of this section is basically to find out about the Fuerteventura varieties’ collection behaviour, paying special attention to the new local variety.
Figure 3a shows how coordinate 1 separates the varieties originating from or with more influence from the east of the Mediterranean Basin from the rest (of Spanish origin). On the other hand, coordinate 2 places in the upper quadrants those varieties known to have chlorotype A (characteristic of the Iberian Peninsula) [53]. It is also known that the female progenitor of these same varieties is the variety that originated in North Africa (according to Pierre Galet [54,55]), known by the names of Heben (main name) or Gibi (very widespread synonym). In addition, the Canary Islands variety Torrontes volcánico and the Majorera variety, of which neither their chlorotype nor their progenitors are known, are also found at this location. The exception is the variety Airen, which is located in the lower left quadrant, very close to the axis. Airen, which also has chlorotype A, has the Heben variety as its female progenitor. Its location so close to the previous varieties would justify the fact that it is considered to belong to the same group and, therefore, within the modus operandi error of all the graphical representations of this study and similar works. The location of each individual on a graph would be due to the confluence of 40 numerical data corresponding to the two alleles of each of the 20 SSRs. Under these conditions, only a 40-dimensional graphical representation would be error-free. It is for this reason that the reduction from 40 to 2 or at best 3 dimensions leads to these deviations. In the lower quadrants, and thus on the same axis or below the axis of coordinate 2, there are mostly varieties with chlorotype D. These are Listan prieto, Palomino fino, Muscat Hamburg and, with chlorotype B, Muscat of Alexandria. But, the latter variety is a cross between the Greek varieties Heptaliko (chlorotype B) and Muscat á petits grains blancs, also with chlorotype D, which means that the Muscat of Alexandria has Balkan Peninsula ancestry, where this chlorotype is mostly from. Muscat Hamburg is a cross between the Greek variety Muscat of Alexandria and the Italian variety Schiva grossa (chlorotype D), carried out by the English hybridiser Seward Snow [44]. Finally, we can also see in the lower quadrants the Canary Island variety Listan negro, of which the chlorotype is unknown but its cross is known. This variety’s progenitors are the Spanish varieties Palomino fino (chlorotype D) and Mollar cano (chlorotype A). As far as the new variety Majorera is concerned, as it is found practically only in the upper right quadrant, it can be concluded that it has a very characteristic MP-SSR and is influenced by the east of the Mediterranean Basin. Figure 3b broadly reproduces the same distribution as Figure 3a for the sample collection under study. Again, the Majorera variety is one of those moving away from the group, together with the surprising position of Muscat Hamburg, which, in the two-dimensional graph, appeared next to its parental, Muscat of Alexandria. Listan negro also suffers a shift between its parentals: whereas, in Figure 3a, it was located close to the Palomino fino, now it is closer to the Mollar cano. Evidently, the reliability of the varieties’ position in this study improves with a dimension increase. In this particular case, the accumulative reliability percentage in two dimensions is 36.68% (the sum of the percentages of coordinates 1 and 2 (Figure 3a)) but, for three dimensions, the goodness of representation increases to 51.76% (sum of the three coordinates in Figure 3b).
Finally, it can be concluded that the new local variety from Fuerteventura has a significantly different MP-SSR from the rest of the viniferas prospected on the island. The differences found in the “signature” variety Muscat Hamburg may be due to the Italian influence of its MP-SSR, thus differentiating it from the rest of the members of this population with Spanish influence.
4.4. Majorera Variety Relation with Respect to Canary Archipelago Grapevine Population
In order to obtain a better understanding of the Majorera variety behaviour and to check whether it continued to be significantly different, in this case from the varieties of the archipelago itself, a population genetics study was carried out. To this end, the 32 Canary Islands varieties were divided into six populations. This proposal was rejected because of the dysfunction of POP1 with respect to POP2 and because POP3, although well defined as such, did not have pure individuals. The proposal to divide the Canary Islands grapevine population into four ancestral populations was then taken. The membership of each individual to a given population was based on the q value, which is defined as the measure of the link of an individual with an ancestral population based on its genetic similarity (percentage of its genome inferred that belongs to the group [56]) and is ordered from the largest to the smallest value (Table S5). Seventeen admixed individuals were detected and eliminated, and the Canary Island population consisted of 15 varieties, which were divided into these four populations. POP1 held the varieties Bienmesabe tinto (Island of La Palma), Majorera (Island of Fuerteventura), Breval negro (Island of Lanzarote) and Blanca de la Granja del Cabildo (Island of Lanzarote) (Figure 4 and Figure S3). Bienmesabe tinto is a variety that was published in 2019 by TECNENOL [39] as a new variety of Vitis vinifera ssp. vinifera, and, after checking its MP-SSR in the VIVC database, did not give any match. Currently, consulting this database, it can be seen that it has been registered as the result of a cross between species of the genus Vitis, i.e., as a direct hybrid producer (DPH), having been previously described by other research groups as such [57,58]. In this sense, the fact that this MP-SSR, which shows the Bienmesabe tinto variety, is positioned so far away from the other grape varieties is of great value. Based on these data and given Majorera’s proximity to this variety and how far it is located from the rest of vinifera, it is possible that it is also a DPH. POP2 is presented as a cluster closely related to HI varieties and, in POP3, the HI and LNZ varieties share prominence. On the other hand, in POP4, we find the progeny of Malvasia Dubrovacka and Bermejuela, Malvasia volcanica and a new variety, also from LNZ, which is known as Malvasia alistanada fina. Malvasia Dubrovacka has a chlorotype A, which is very frequent in the Iberian Peninsula but rare in the Balkan Peninsula [49,53], although several studies group its MP-SSR with Malvasia profiles from the Balkans [59]. Palomino fino progeny, Listan negro (Palomino fino × Mollar cano), Albillo criollo and Albillo forastero (Palomino fino × Verdelho branco) are also located in this population. This is important because of the chlorotype D of the Palomino fino variety, which is closely related to the eastern Mediterranean basin [49,53]. Therefore, it can be concluded that this cluster groups varieties (pure or admixed) with Eastern Mediterranean influence.
Figure 5 shows contradictory information regarding the relationship that these populations may have. While, in Figure 5a–e, it can be seen that POP1 and POP2 are markedly differentiated in all images, in the case of POP3 and POP4, it depends on whether the graph is represented by individuals or populations. In the case of population representations (Figure 5a), POP3 and POP4 appear distant and, in the case of individual representations, they are related. Clearly, the reliability of this representation has decreased with respect to the representation by populations. Nevertheless, trends can be observed. It can also be seen how POP1 presents all its individuals as dispersed, while the rest of the populations maintain all their individuals as grouped together as a sample of the relationship between them, clearly differentiating one population from the other.
With regard to the variety under study, Majorera, it can be concluded in this section that it has a significantly different MP-SSR from the rest of the Canary Islands varieties, with the exception of the Bienmesabe tinto variety. This uniqueness may be due to the nature of its progenitors. There is a possibility that this variety is a DPH, i.e., an interspecific cross. However, the fact that, in the three-dimensional representation (Figure 5d), it is very distant from the Bienmesabe tinto variety may mean that its parents (interspecific or not) are not very closely related to the Bienmesabe tinto variety parentals.
4.5. Majorera Variety Relation with Respect to World Population
The availability of a complete database for Vitis vinifera ssp. vinifera in the TECNENOL research group allowed us to compare the MP-SSR of the Majorera variety with 308 other varieties from 22 countries around the world and, in this way, attempt to confirm the indications that point to this genetic profile as an interspecific cross.
The first strategy that was tested was purely genetic. Structure 2.3 prioritised the distribution K = 2, forming two populations that are markedly differentiated: POP1 (with mostly varieties from the rest of the world) and POP2 (with mostly Spanish individuals and from which the Canarian population was extracted (POP3)) (Figure 6b,c and Figure 7a,b). This behaviour is observed in all TECNENOL works; in the same way, the CI population singularity with respect to the Spanish population and also with the world population is also observed in all of them (Figure 7(a.2,a.3)) when working with total populations. However, in the representations with all individuals in the form of a circular dendrogram (Figure 6b), phylogenetic tree (Figure 6c) and two-dimensional PCoA (Figure 7(a.1)), it is no longer possible to demonstrate the uniqueness of IC with respect to POP2. If the behaviour of the varieties Bienmesabe tinto and Majorera is observed in these graphical representations, with the former being a DPH and the latter having the potential to be one, there are no significant differences with respect to the rest of the varieties either. Bienmesabe tinto and Majorera varieties’ locations overlap with the populations of POP2 and POP3 (IC) without giving relevant information. Only in the three-dimensional representation (Figure 7b) do these two varieties appear as significantly distant from the rest, especially the Bienmesabe tinto variety. It should be noted that an additional dimension adds reliability to the final result. In this case, the reliability went from 9.36% in the two-dimensional representation to 12.88% in the three-dimensional graph, which represents an increase of 27.33%. Despite this, the final goodness of fit is still significantly low and similar to that described by most authors for PCoA in two dimensions [59,60].
In addition, another strategy was tested. This included a geographic component. Thus, this world population was grouped by geographical areas as proposed by other authors [53,56], according to the origin of the variety registered in the VIVC.
In this particular case, the 309 varieties, 307 belonging to Vitis vinifera ssp. vinifera, the Bienmesabe tinto variety as DPH and the Majorera variety that could have originated from another interspecific cross, were grouped into six areas. The objective was to see what influence the geographical component had in supporting the possibility that the only MP-SSR on the island of Fuerteventura was not a pure vinifera.
When distributing the world population into six geographical areas, the global behaviour observed was similar to the previous one. The representation by population, once again, shows the uniqueness not only of the IC varieties but also the uniqueness of the PI varieties with respect to the rest of the varieties in the world (Figure 8a,b and Figure 9a.2,a.3,b). In addition, the singularity between IC and PI can also be observed, although they are closely related. In contrast, the individual representation in Figure 9(a.1) shows no differentiation between IC and PI. Figure 8a,b show how some PI varieties are located in branches belonging to the EAST-MED-CAU, BP and ITA areas, and in the one belonging to the FRA-CE area. This transposition may well be due to the influence of the MP-SSRs in question on these reference areas. Possibly, this influence is due not only to the conquest of new territories by the Crown of Aragon, which in the Middle Ages reached as far as the Balkan Peninsula [61], but also to the Camino de Santiago (FRA-CE) [62]. It is obvious to think that, in these situations, there was a bidirectional transfer of varieties. Regarding the varieties Bienmesabe tinto and Majorera, again it will be the three-dimensional PCoA representation (Figure 9b) that distinguishes these varieties significantly from the rest of the world varieties.
It must also be noted that, for both strategies, (a) coordinate 1 of Figure 7(a.1) and Figure 9(a.1) is the one that separates the specific populations for each case, while Coordinate 2 does not have a notable role or does not have any paper; (b) the major Spanish populations (peninsular and insular), as well as the Bienmesabe tinto and Majorera varieties, overlap without distinction (Figure 7(a.1) and Figure 9(a.1)); (c) it is always the third dimension collected in Figure 7b and Figure 9b that is able to differentiate between Bienmesabe tinto and Majorera varieties, and consequently show their uniqueness.
In conclusion, it can be hypothesised that there is a possibility that the Majorera variety from Fuerteventura Island is a DPH. This will have to be confirmed in further studies.
5. Conclusions
After carrying out this study, it can be concluded that, once again, the SSR kit used for this study worked correctly. Regarding the MP-SSR, this study has described: (a) 10 different variety profiles, where 9 correspond to known varieties and 1 corresponds to a new variety; (b) 4 new mutations of known varieties (Listan prieto from Antigua, Listan prieto from Vega, Hoja moral from El Rosario and Hoja moral from Betancuria); (c) identified mutations of known varieties that have been described in other works (Listan negro santanero, Mollar bonilla, Listan blanca chicharrera); and (d) 13 MP-SSR of accessions registered as unknown that have been identified. At a lexicographic level, five erratic names have been detected. Furthermore, this work proposes the inclusion in the global database (VIVC) of seven new names: one main name of a variety (Majorera); one new name synonymous with the Canarian variety Listan negro (Hoja moral); four names of mutations (Listan prieto from Antigua, Listan prieto from Vega, Hoja moral from El Rosario and Hoja moral from Betancuria); and the new name synonymous with the Moscatel variety Hamburg (Muscat). The uniqueness of the population of Canary Islands varieties as a potential centre for creating biodiversity for cultivated vines is also confirmed. Finally, the possibility is raised that the new variety from the island of Fuerteventura derives from an intraspecific crossing.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9121301/s1, Table S1: Information (original and conclusive) on 40 accessions from Fuerteventura Island. Table S2: Results of 7 international SSRs of 15 genetic profiles from Fuerteventura that correspond to 10 different varieties. The yellow box indicates a mutated allele. Table S3: Annealing temperature (Ta) of SSR. Table S4: Characterisation of the twenty microsatellite markers used in this study. Figure S1: The four steps of the graphical method of Evanno et al., (2005), allowing for the estimation of the true number of ancestral K groups for a population with 32 individuals from the Canary Islands collection (IC, including Fuerteventura Island). Figure S2: Plots of the best distributions of the 32 individuals of the Canary Islands (including Fuerteventura) from Structure 2.3. programme for 4K and 6K. Table S5: Genetic structure of the Canary Islands population. Distribution K = 6 (individuals belonging to each group or population). Breakdown of the ratio of pure and admixed individuals according to the q value (pure (q ≥ 85%) and admixed (q < 85%)). Figure S3: Genetic structure of the Canary Islands population (32 varieties). Distribution K = 4 (individuals belonging to each group or population). Breakdown of the ratio of pure and admixed individuals according to the q value (pure (q ≥ 85%) and admixed (q < 85%)). Status of each population. Figure S4: The four steps of the graphical method of Evanno et al., (2005), allowing for the estimation of the true number of ancestral K groups for a population of 309 varieties from the TECNENOL database. Figure S5: Genetic structure of the world population (309 individuals). Distribution K = 2 (individuals belonging to each group or population). Breakdown of the ratio of pure and admixed individuals according to the q value (pure (q ≥ 85%) and admixed (q < 85%)). Status of each population. Figure S6: Genetic structure of the world population (188 individuals). Distribution in 6 geographical areas. Breakdown of the well-assigned (pure) and wrong-assigned (admixed) individuals. Nations that compose each of the groups: EASTMED-CAU (Algeria, Cyprus, Georgia, Israel, Lebanon, Tunisia and Turkey), BP (Bosnia and Herzegovina, Bulgaria, Croatia, Greece, Serbia, Slovenia and Montenegro), ITA (Italy), FRA-CEU (Austria, France, Germany, Hungary and Switzerland), IP (Spain and Portugal) and IC (Canary archipelago).
Author Contributions
Conceptualisation, F.F. and Q.L.-Y.; methodology, F.F., Q.L.-Y. and C.V.; software, P.S.-G., J.M.C. and F.Z.; validation, F.F. and Q.L.-Y.; formal analysis, F.F., Q.L.-Y. and C.V.; investigation, F.F., Q.L.-Y. and C.V.; resources, F.F.; data curation, F.F. and Q.L.-Y.; writing—original draft preparation, F.F. and P.S.-G.; writing—review and editing, F.F. and P.S.-G.; visualisation, J.M.C. and F.Z.; supervision, F.F., Q.L.-Y., C.V., P.S.-G., J.M.C. and F.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This project has been funded by Cabildo Insular de la Isla de Fuerteventura.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors are grateful to Javier Ibañez and the viticulture specialists Alberto Gerardo González and Juan Manuel Brito for their valuable support. We would also like to thank Luis Ricardo Suárez de Abreu, Rosa Pastor, Isabel Araque, Braulio Esteve-Zarzoso, Laia Fañanás and Santiago Moreno for their support in the laboratory.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wolkovich, E.M.; García de Cortázar-Atauri, I.; Morales-Castilla, I.; Nicholas, K.A.; Lacombe, T. From Pinot to Xinomavro in the world’s future wine-growing regions. Nat. Clim. Chang. 2018, 8, 29–37. [Google Scholar] [CrossRef]
- Togores, J.H.; Fernández-Cano, L.H. Tratado de Viticultura, 2nd ed.; Ediciones Mundi-Prensa: Madrid, Spain, 2019; Volume 1. [Google Scholar]
- Jones, G.V. Climate Change: Observations, Projections, and General Implications for Viticulture and Wine Production. Economics Department, Oregon University. 2007, Paper No. 7, 15. Available online: https://www.researchgate.net/publication/253713937_Climate_Change_Observations_Projections_and_General_Implications_for_Viticulture_and_Wine_Production (accessed on 20 August 2023).
- Roca, P. Dos Bazas Para el Sector Vitivinícola en un Contexto Incierto, el Discurso de Pau Roca en el Congreso Mundial de la Viña y el Vino; Tecnovino: Bilbao, Spain, 2023; Available online: https://www.tecnovino.com/dos-bazas-para-el-sector-vitivinicola-en-un-contexto-incierto-el-discurso-de-pau-roca-en-el-congreso-mundial-de-la-vina-y-el-vino/ (accessed on 20 August 2023).
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [PubMed]
- Coordinadora de Organizaciones de Agricultores y Ganaderos (COAG). Cambio Climático y Viñedo en España. Available online: https://adaptecca.es/sites/default/files/documentos/cambio_climatico_y_vinedo_en_espana_2016.pdf (accessed on 20 August 2023).
- Bota, J.; Tomas, M.; Flexas, J.; Medrano, H.; Escalona, J.M. Differences among grapevine cultivars in their stomatal behavior and water use efficiency under progressive water stress. Agric. Water Manag. 2016, 164, 91–99. [Google Scholar] [CrossRef]
- Wikipedia. Available online: https://es.wikipedia.org/wiki/Fuerteventura (accessed on 7 July 2023).
- Wikimedia Commons. 2014. Available online: https://commons.wikimedia.org/w/index.php?curid=33566121#file (accessed on 7 July 2023).
- NASA. 2011. Available online: https://www.flickr.com/photos/gsfc/6630087415/in/photostream/ (accessed on 7 July 2023).
- Instituto Canario de Estadística (ISTAC). 2005. Available online: http://www.gobiernodecanarias.org:80/istac/publicaciones/pdfs/2006_005.pdf (accessed on 7 July 2023).
- Climate Data. 2021. Available online: https://es.climate-data.org/europe/espana/fuerteventura-10238/ (accessed on 7 July 2023).
- Lana, D. Diagnóstico de la Viticultura en Canarias. Consejería de Agricultura; Ganadería y Pesca: Sevilla, Spain, 2021; Available online: https://www.gobiernodecanarias.org/agricultura/detalle/Diagnostico-de-la-viticultura-en-Canarias/ (accessed on 7 July 2023).
- Otamendi, J.J. Historia del Vino de Lanzarote. Notas Para Su Estudio; Bodegas El Grifo: Lanzarote, Spain, 2013; Available online: https://docplayer.es/9690813-Historia-del-vino-de-lanzarote-notas-para-su-estudio.html (accessed on 7 July 2023).
- Las Gavias, un Tesoro en la Tierra de Fuerteventura y Lanzarote. In Ciencia Canaria; Agencia Canaria de Investigación, Innovación y Sociedad de la Información: Santa Cruz de Tenerife, Spain, 2017; Available online: https://www.cienciacanaria.es/secciones/a-fondo/865-las-gavias-un-tesoro-en-la-tierra-de-fuerteventura-y-lanzarote (accessed on 7 July 2023).
- Marsal, G.; Baiges, I.; Canals, J.M.; Zamora, F.; Fort, F. A fast, efficient method for extracting DNA from leaves, stems, and seeds of Vitis vinifera L. Am. J. Enol. Vitic. 2011, 62, 376–381. [Google Scholar] [CrossRef]
- Marsal, G.; Boronat, N.; Canals, J.M.; Zamora, F.; Fort, F. Comparison of the efficiency of some of the most usual DNA extraction methods for woody plants in different tissues of Vitis vinifera L. J. Int. Sci. Vigne Vin 2013, 47, 227–237. [Google Scholar] [CrossRef]
- Fort, F.; Hayoun, L.; Valls, J.; Canals, J.M.; Arola, L.; Zamora, F. A new and simple method for rapid extraction and isolation of high-quality RNA from grape (Vitis vinifera) berries. J. Sci. Food Agric. 2008, 88, 179–184. [Google Scholar] [CrossRef]
- This, P.; Jung, A.; Boccacci, P.; Borrego, J.; Botta, R.; Costantini, L.; Crespan, M.; Dangl, G.S.; Eisenheld, C.; Ferreira-Monteiro, F.; et al. Development of a standard set of microsatellite reference alleles for the identification of grape cultivars. Theor. Appl. Genet. 2004, 109, 1448–1458. [Google Scholar] [CrossRef]
- Thomas, M.R.; Scott, N.S. Microsatellite repeats in grapevine reveal DNA polymorphisms when analyzed as sequence-tagged sites (STSs). Theor. Appl. Genet. 1993, 86, 985–990. [Google Scholar] [CrossRef]
- Bowers, J.E.; Dangl, G.S.; Vignani, R.; Meredith, C.P. Isolation and characterization of new polymorphic simple sequence repeat loci in grape (Vitis vinifera L.). Genome 1996, 39, 628–633. [Google Scholar] [CrossRef]
- Bowers, J.E.; Dangl, G.S.; Meredith, C.P. Development and characterization of additional microsatellite DNA markers for grape. Am. J. Enol. Vitic. 1999, 50, 243–246. [Google Scholar] [CrossRef]
- Sefc, K.M.; Regner, F.; Turetschek, E.; Glössl, J.; Steinkellner, H. Identification of microsatellite sequences in Vitis riparia and their applicability for genotyping of different Vitis species. Genome 1999, 42, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.D.; Eggler, P.; Seaton, G.; Rosseto, M.; Abblet, E.M.; Lee, L.S.; Henry, R.J. Analysis of SSRs derived from grape ESTs. Theor. Appl. Genet. 2000, 100, 723–726. [Google Scholar] [CrossRef]
- Lefort, F.; Kyvelos, C.; Zervou, M.; Edwards, K.; Roubelakis-Angelakis, K. Characterization of new microsatellite loci from Vitis vinifera and their conservation in some Vitis species and hybrids. Mol. Ecol. Resour. 2002, 2, 20–21. [Google Scholar] [CrossRef]
- Cipriani, G.; Spadotto, A.; Jurman, I.; Di Gaspero, G.; Crespan, M.; Meneghetti, S.; Frare, E.; Vignani, R.; Cresti, M.; Morgante, M.; et al. The SSR-based molecular profile of 1005 grapevine (Vitis vinifera L.) accessions uncovers new synonymy and parentages and reveals a large admixture amongst varieties of different geographic origins. Theor. Appl. Genet. 2010, 121, 1569–1585. [Google Scholar] [CrossRef] [PubMed]
- Dalbó, M.A.; Ye, G.N.; Weeden, N.F.; Steinkellner, H.; Sefc, K.M.; Reisch, B.I. A gene-controlling sex in grapevines is placed on a molecular marker-based genetic map. Genome 2000, 43, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Paetkau, D.; Calvert, W.; Stirling, I.; Strobeck, C. Microsatellite analysis of population structure in Canadian polar bears. Mol. Ecol. 1995, 4, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Paetkau, D.; Slade, R.; Burden, M.; Estoup, A. Genetic assignment methods for the direct, real-time estimation of migration rate: A simulation-based exploration of accuracy and power. Mol. Ecol. 2004, 13, 55–65. [Google Scholar] [CrossRef]
- Three-Dimensional Plotting in Matplotlib (Python Data Science Handbook). Available online: https://jakevdp.github.io/PythonDataScienceHandbook/04.12-three-dimensional-plotting.html (accessed on 20 August 2023).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic threes. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Marsal, G.; Mateo, J.M.; Canals, J.M.; Zamora, F.; Fort, F. SSR analysis of 338 accessions planted in Penedes (Spain) reveals 28 unreported molecular profiles of Vitis vinifera L. Am. J. Enol. Vitic. 2016, 67, 466–470. [Google Scholar] [CrossRef]
- Marsal, G.; Bota, J.; Martorell, A.; Canals, J.M.; Zamora, F.; Fort, F. Local cultivars of Vitis vinifera L. in Spanish Islands: Balearic Archipelago. Sci. Hortic. 2017, 226, 122–132. [Google Scholar] [CrossRef]
- Marsal, G.; Mendez, J.J.; Mateo-Sanz, J.M.; Ferrer, S.; Canals, J.M.; Zamora, F.; Fort, F. Molecular characterization of Vitis vinifera L. local cultivars from volcanic areas (the Canary Islands and Madeira) using SSR markers. OENO One 2019, 4, 667–680. [Google Scholar] [CrossRef]
- Fort, F.; Marsal, G.; Mateo-Sanz, J.M.; Pena, V.; Canals, J.M.; Zamora, F. Molecular characterisation of the current cultivars of Vitis vinifera L. in Lanzarote (Canary Islands, Spain) reveals nine individuals which correspond to eight new varieties and two new sports. OENO One 2022, 56, 281–295. [Google Scholar] [CrossRef]
- Fort, F.; Lin, Q.; Suárez-Abreu, L.R.; Sancho-Galán, P.; Canals, J.M.; Zamora, F. Study of Molecular Biodiversity and Population Structure of Vitis vinifera L. ssp. vinifera on the volcanic island of El Hierro (Canary Islands (Spain)) by using Microsatellite Markers. Horticulturae, 2024; submitted. [Google Scholar]
- Zerolo, J.; Cabello, F.; Espino, A.; Borrego, J.; Ibañez, J.; Rodriguez-Torres, I.; Muñoz-Organero, G.; Rubio, C.; Hernández, M. Variedades de Vid de Cultivo Tradicional en Canarias, 1st ed.; Instituto Canario de Calidad Agroalimentaria, Gobierno de Canarias: Santa Cruz de Tenerife, Spain, 2006. [Google Scholar]
- Rodriguez-Torres, I. Variedades de vid cultivadas en Canarias. Descriptores Morfológicos. In Caracterización Morfológica, Molecular, Agronómica y Enológica, 1st ed.; Instituto Canario de Investigaciones Agrarias, Gobierno de Canarias: Santa Cruz de Tenerife, Spain, 2018. [Google Scholar]
- Maul, E.; Röckel, F. Vitis International Variety Catalogue. 2015. Available online: http://www.vivc.de (accessed on 7 July 2023).
- Ibañez, J.; De Andrés, M.T.; Molino, A.; Borrego, J. Genetic study of key Spanish grapevine varieties using microsatellite analysis. Am. J. Enol. Vitic. 2003, 54, 22–30. [Google Scholar] [CrossRef]
- Lopes, M.S.; Rodrigues dos Santos, M.; Eiras Dias, J.E.; Mendonça, D.; da Câmara Machadoa, A. Discrimination of Portuguese grapevines based on microsatellite markers. J. Biotechnol. 2006, 127, 34–44. [Google Scholar] [CrossRef]
- Aliquo, G.; Torres, R.; Lacombe, T.; Boursiquot, J.M.; Laucou, V.; Gualpa, J.; Fanzone, M.; Sari, S.; Pérez-Peña, J.; Prieto, J.A. Identity and parentage of some South American grapevine cultivars present in Argentina. Aust. J. Grape Wine Res. 2017, 23, 452–460. [Google Scholar] [CrossRef]
- Moita, A.; Santos, R.; Catarina, A. Unraveling the origin of Vitis vinifera L. Verdelho. Aust. J. Grape Wine Res. 2018, 24, 450–460. [Google Scholar] [CrossRef]
- Žulj Mihaljević, M.; Maletić, E.; Preiner, D.; Zdunić, G.; Bubola, M.; Zyprian, E.; Pejić, I. Genetic Diversity, Population Structure, and Parentage Analysis of Croatian Grapevine Germplasm. Genes 2020, 11, 737. [Google Scholar] [CrossRef]
- de Oliveira, G.L.; Niederauer, G.F.; de Oliveira, F.A.; Rodrigues, C.S.; Hernandes, J.L.; de Souza, A.P.; Moura, M.F. Genetic diversity, population structure and parentage analysis of Brazilian grapevine hybrids after half a century of genetic breeding. Sci. Hortic. 2023, 311, 111825. [Google Scholar] [CrossRef]
- Arslan, N.; Yılmaz Baydu, F.; Hazrati, N.; Yüksel Özmen, C.; Ergönül, O.; Uysal, T.; Ya¸sasın, A.S.; Özer, C.; Boz, Y.; Kuleyin, Y.S.; et al. Genetic Diversity and Population Structure Analysis of Anatolian Kara Grapevine (Vitis vinifera L.) Germplasm Using Simple Sequence Repeats. Horticulturae 2023, 9, 743. [Google Scholar] [CrossRef]
- Jiménez-Cantizano, A.; Puig-Pujol, A.; Arroyo-García, R. Identification of Vitis vinifera L. Local Cultivars Recovered in Andalusia (Spain) by Using Microsatellite Markers. Horticulturae 2023, 9, 316. [Google Scholar] [CrossRef]
- Arroyo-García, R.; Ruiz-García, L.; Bolling, L.; Ocete, R.; López, M.A.; Arnold, C.; Ergul, A.; Söylemezoğlu, G.; Uzun, H.I.; Cabello, F.; et al. Multiple origins of cultivated grapevine (Vitis vinifera L. ssp. sativa) based on chloroplast DNA polymorphisms. Mol. Ecol. 2006, 15, 3707–3714. [Google Scholar] [CrossRef]
- Galet, P. Dictionnaire Encylcopédique des Cépages, 1st ed.; Hachette: Paris, France, 2000. [Google Scholar]
- Lacombe, T.; Boursiquot, J.M.; Laucou, V.; Dechesne, F.; Varès, D.; This, P. Relationships and genetic diversity within the accessions related to Malvasia held in the Domaine de Vassal grape germplasm repository. Am. J. Enol. Vitic. 2007, 58, 124–131. [Google Scholar] [CrossRef]
- Bacilieri, R.; Lacombe, T.; Cunff, L.L.; Di Vecchi-Staraz, M.; Laucou, V.; Genna, B.; Perós, J.P.; This, P.; Boursiquot, J.M. Genetic structure in cultivated grapevines is linked to geography and human selection. BMC Plant Biol. 2013, 13, 25. [Google Scholar] [CrossRef]
- Moreno-Sanz, P.; Loureiro, M.D.; Suárez, B. Microsatellite characterization of grapevine (Vitis vinifera L.) genetic diversity in Asturias (Northern spain). Sci. Hortic. 2011, 129, 433–440. [Google Scholar] [CrossRef]
- Schneider, A.; Mainardi, G.; Raimondi, S. Ampelografia Universale Storica Illustrata. I Vitigni del Mondo, 1st ed.; L’Artistica: Savigliano, Italy, 2012. [Google Scholar]
- Dong, Y.; Duan, S.; Xia, Q.; Liang, Z.; Dong, X.; Margaryan, K.; Musayev, M.; Goryslavets, S.; Zdunić, G.; Bert, P.-F.; et al. Dual domestications and origin of traits in grapevine evolution. Science 2023, 379, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martínez-Zapater, J.M.; Zyprian, E.; Moreira, F.M.; et al. Genetic diversity and population structure assessed by SSR and SNP markers in large germplasm collection of grape. BMC Plant Biol. 2013, 13, 39. [Google Scholar] [CrossRef] [PubMed]
- García-Muñoz, S.; Lacombe, T.; De Andrés, M.T.; Gaforio, L.; Muñoz-Organero, G.; Laucou, V.; This, P.; Cabello, F. Grape varieties (Vitis vinifera L.) from the Balearic Islands: Genetic characterization and relationship with Iberian Peninsula and Mediterranean Basin. Genet. Resour. Crop Evol. 2012, 59, 589–605. [Google Scholar] [CrossRef]
- Casanova, J.; Mozas, P.; Ortiz, J.M. Ampelography and microsatellite DNA analysis of autochthonous and endangered grapevine cultivars in the province of Huesca (Spain). Span. J. Agric. Res. 2011, 9, 790–800. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).