1. Introduction
A recent study shows that not only is the temperature a limiting factor for head formation in this type of lettuce, but mainly solar radiation, with a range of 17.6 to 19.5 MJ·m
−2·day
−1 of solar radiation global being ideal for iceberg lettuce production at average temperatures of 27.8 °C [
3]. High irradiance causes photoinhibition in this crop, as the radiation flow above the trophic limit can lead to increased transpiration through the upper leaves and possible stomatal closure [
4].
Iceberg lettuce production is directly influenced by temperature since it is a species from temperate regions. Head lettuce formation is impaired at temperatures above 20 °C. High temperatures stimulate its bolting, which is accelerated as this climatic factor increases [
1]. This is why the production of this vegetable in high-temperature regions, mainly semi-arid, is still not very expressive. In these regions, the weather conditions are characterized by high solar radiation (average 21.6 MJ·m
−2·day
−1), high temperatures (average 26 °C), low relative humidity, and reduced rainfall (average 700 mm per year) [
2].
The use of shading screens reduces the solar radiation direct incidence in plants, thus causing temperature reduction, which favors crops that need lower radiant energy flow. Lower incidence of solar energy can contribute to reducing the extreme effects of radiation, especially on photorespiration, and provide better environmental conditions for increasing the yield and quality of leaves for consumption [
5].
Thus, the shading screens alter the microclimatic conditions of the environments (temperature, relative humidity, and solar radiation). The light intensity is also affected, which directly affects several photoreceptors present in plants [
6], causing metabolic alterations and, consequently, influencing plant growth and development. Lettuce does not require high light intensity, about 500 μmol·m
−2·s
−1 in summer [
7]. Light intensity is an important factor for plant metabolism. The reduction of this factor causes photosynthesis not to work efficiently, leading to etiolation, but excessive light intensity can generate reactive oxygen species (ROS) and cause photoinhibition [
8].
ROS cause oxidative damage to the lipids and membrane proteins. To prevent this damage, plants activate various defense mechanisms. One of the forms of defense is the activation of the enzymatic antioxidant system that includes superoxide dismutase (SOD, EC 1.15.1.1), catalase (CAT, EC 1.11.1.6), and peroxidase (POD,1.11.1.7) [
9]. The activity of enzymes SOD, CAT, and POD can indicate whether the lettuce plants are under environmental stress. Studies involving the activity of antioxidant enzymes in lettuce plants under water deficit, salinity, and artificial lighting have already been reported and the activity of the enzymes SOD, CAT, and POD increase significantly under stress conditions [
10,
11].
Through plant breeding, cultivars that are more tolerant to high temperatures and solar radiation have been developed, and are thus adapted to tropical conditions without stimulating bolting and latex production. Allied to this, the technique of using shading screens to protect crops can increase the production of lettuce, allowing the growers to produce lettuce throughout the year, including during high temperatures or in the offseason [
12]. However, there is a lack of information on the relationship between these enzymes’ activity and the levels of sugars in the leaves during formation of the head of iceberg lettuce in tropical conditions of high temperature and solar radiation.
Based on the presented arguments, our research hypothesizes that the use of shading screens can be an efficient management for the cultivation of iceberg lettuce in regions of high temperature linked to the indication of cultivars adapted to these conditions. Therefore, this study aimed to evaluate the biochemical responses: sugar levels, the activity of antioxidant enzymes; and the commercial production of iceberg lettuce cultivars under environments with different shadings.
2. Materials and Methods
The experiment was carried out in the Experimental Vegetable Production Area of the Department of Technology and Social Sciences, Juazeiro Campus, of the State University of Bahia, in Juazeiro, Brazil, located at 9°25′14″ S and 40°29′08″ W with an average altitude of 360 m above sea level. According to the Köppen classification, the region has a semi-arid climate, BSh-type, with an average temperature and precipitation of 23.6 °C and 455 mm. From October to December 2019, the following events occurred: sowing, 10 October 2019; transplanting, 5 November 2019; harvest, 20 December 2019. In the spring season, the northeast region of Brazil experienced a period of high temperatures.
A randomized block design arranged in a 4 × 4 factorial scheme with four replications was used. Four iceberg lettuce cultivars (Great Lakes, Winslow, Delícia, and Balsamo) and four environments with different shadings (without shading under the full sun (FS), shading with a red screen (RS), shading with ChromatiNet
® (CS), and shading with a black screen (BS)) were evaluated. According to the manufacturer, the ChromatiNet
® screen is a mesh that manipulates the spectrum of sunlight, breaking direct light by converting it to diffused light [
13]. Each plot consisted of twelve plants, and the four plants in the middle of plots were used for the evaluations.
The iceberg lettuce seedlings were produced in Styrofoam trays, filled with commercial substrate Plantmax® with 4.90 g·kg−1 of N, 1.47 g·kg−1 of P, 7.0 g·kg−1 K, 12.50 g·kg−1 of Ca, 120 g·kg−1 of Mg, 1.80 g·kg−1 of S, C/N of 34/1, 285 g·kg−1 of organic matter, and pH of 6.20. The trays were placed on benches with 0.5 m height, under a protected environment covered with 50% black shading screen, and after 25 days, they were transplanted to the seedbeds with a spacing of 0.25 × 0.25 m. The shading environments had a ceiling height of 2.00 m, and the screens were fixed on eucalyptus stakes with wire at 1.50 m height.
The soil had the following chemical properties: pH = 6.42, P = 56 mg dm−3, K = 0.22 cmolc dm−3, Ca = 5.0 cmolc dm−3, Mg = 1.70 cmolc dm−3, Al = 0.0 cmolc dm−3, CTC = 7.01 cmolc dm−3, and EC = 0.77 dS.m−1. Soil preparation was carried out with plowing, harrowing, and seedbed preparation at 30 cm height. Each seedbed was 16 m long by 1.5 m wide. Each experimental plot had 0.75 × 1.50 m with twelve plants. The four plants in the middle of plot were used for the evaluations. Planting fertilization was based on the recommendation of fertilization of the state of Bahia for lettuce cultivation by applying 6.3 kg ha−1 of N and 21.3 kg of P2O5 ha−1, using urea (44% of N) and simple superphosphate (18% of P2O5). For the topdressing fertilization, we also followed the recommendation of fertilization of the state of Bahia for lettuce cultivation by applying 38.0 kg ha−1 of N and 26.0 kg ha−1 of Ca, using calcium nitrate (15% of N and 19% Ca) as a source, performed 20 days after transplanting.
The air temperature (average, maximum, and minimum), relative humidity (average, maximum, and minimum), and solar radiation from the shading treatments and full sun were monitored using meteorological sensors installed at 50 cm height. The sensors were connected to the automatic data system (Datalogger), programmed to take readings every five seconds and hourly averages.
Plants were harvested 45 days after transplanting (DAT) and were taken to the laboratory where the plants were weighed to determine the fresh consumable part (i.e., heads) (FCP), removing the external. The results are expressed in g·plant−1. Immediately after harvesting, two replications with four central leaves from two plants per plot were collected and wrapped in aluminum foil, submerged in liquid nitrogen, and stored in an ultra-freezer at −80 °C for enzymatic analysis and sugar content evaluation.
The raw extract was obtained for all analyzes by macerating 1 g of the sample in 5 mL of 0.1 M potassium phosphate buffer (pH 7.5) containing 1.0 mM of ethylenediaminetetra acetic acid (EDTA), 3.0 mM dithiothreitol (DTT), 5% polyvinylpolypyrrolidone (PVPP), and 10.0 mM ascorbic acid and centrifuged at 10,000 rpm for 10 min at 4.0 °C. The supernatant was used to determine total protein content (TP), total soluble sugars (TSS), reducing sugars (RS), and activities of the enzymes superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD).
The total protein content (TP) was obtained using the method described by [
14], and was measured to determine specific enzyme activity. For the total soluble sugar (TSS) content, the anthrone method described in [
15,
16] was used. The content of reducing sugars (RS) was quantified by the method described by [
17], using 3,5-dinitrosalicylic (DNS).
The activity of the superoxide dismutase (SOD) enzyme was determined according to [
18], taking into account the enzyme’s ability to inhibit the photo-reduction of nitrotetrazolium blue chloride (NBT), expressed in UE·min
−1·mgTP
−1 (unit of enzyme per minute per milligram of protein). For the catalase enzyme (CAT) activity, the methodology described by [
19] was used. The results are expressed as μmolH
2O
2·min
−1·μgTP
−1. POD activity was determined in a spectrophotometer by measuring the formation of purpurogallin at 430 nm [
20], and the activity is expressed in UE·min
−1·g
−1.
The data were submitted to the Anderson Darling’s homogeneity test through the Minitab software for statistical analysis. Once the normality of the data was verified, the analysis of variance (F-test) was performed, in which the means were grouped by the Scott Knott test at 5% probability, using the AGROESTAT® software. Correlation analysis was performed among all evaluated variables.
3. Results
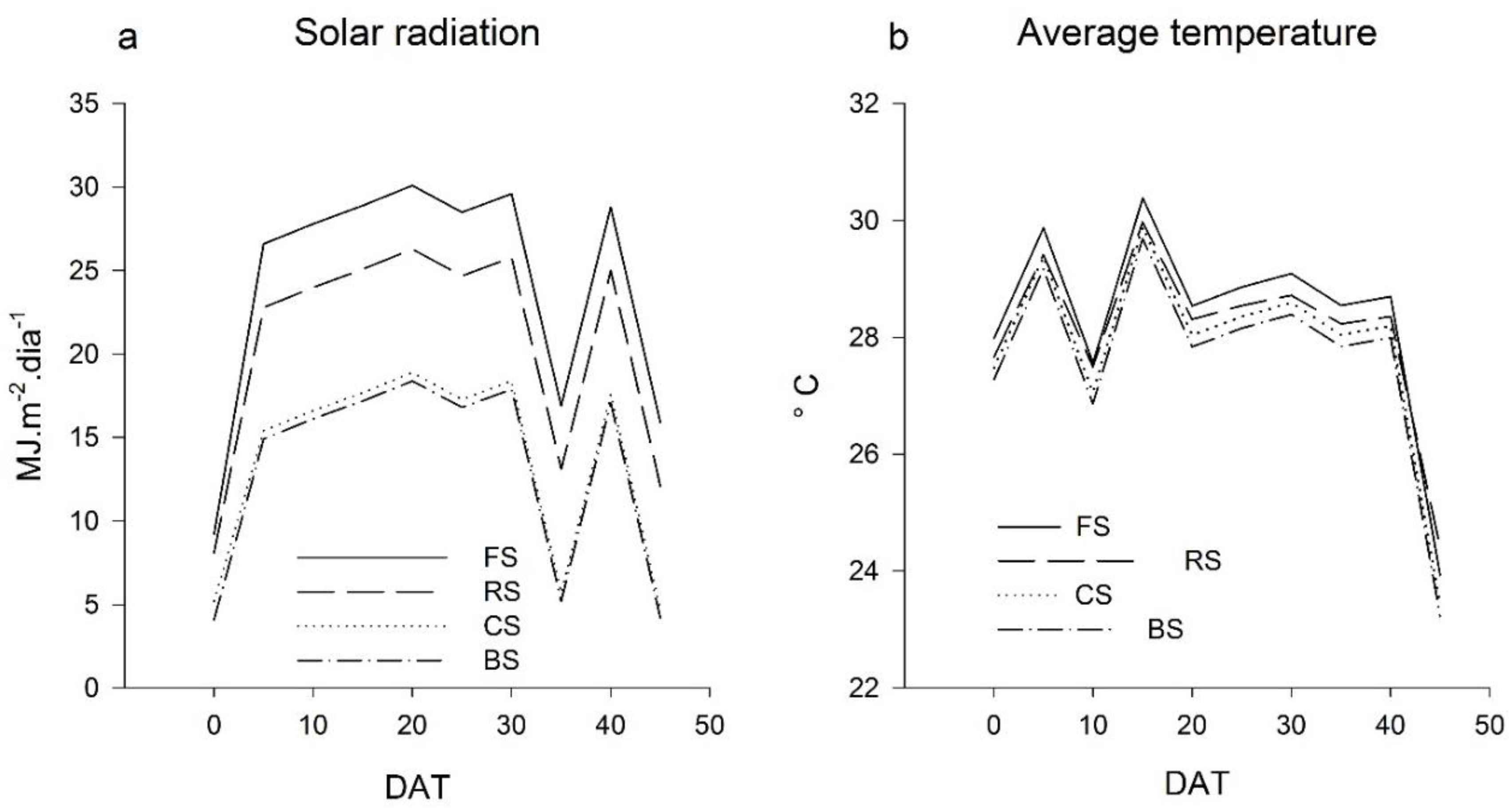
Among the microclimatic characteristics evaluated, global solar radiation was the most affected by shading screens (
Figure 1a), which, in turn, affected plant development. The average radiation observed in the period was about 16 MJ·m
−2·day
−1, 18 MJ·m
−2·day
−1, 25 MJ·m
−2·day
−1, and 30 MJ·m
−2·day
−1 in black screen (BS), ChromatiNet
® screen (CS), red screen (RS), and full sun (FS), respectively. There was a reduction of 0.5 °C in the shaded environments with BS and CS concerning the FS. The average temperature values during the period were 28.1 °C, 28.0 °C, 27.6 °C, and 27.6 °C for FS, VS, CS, and BS, respectively (
Figure 1b). For the transmittance, the red, ChromatiNet
®, and black shading screens presented 80%, 60%, and 55%, respectively.
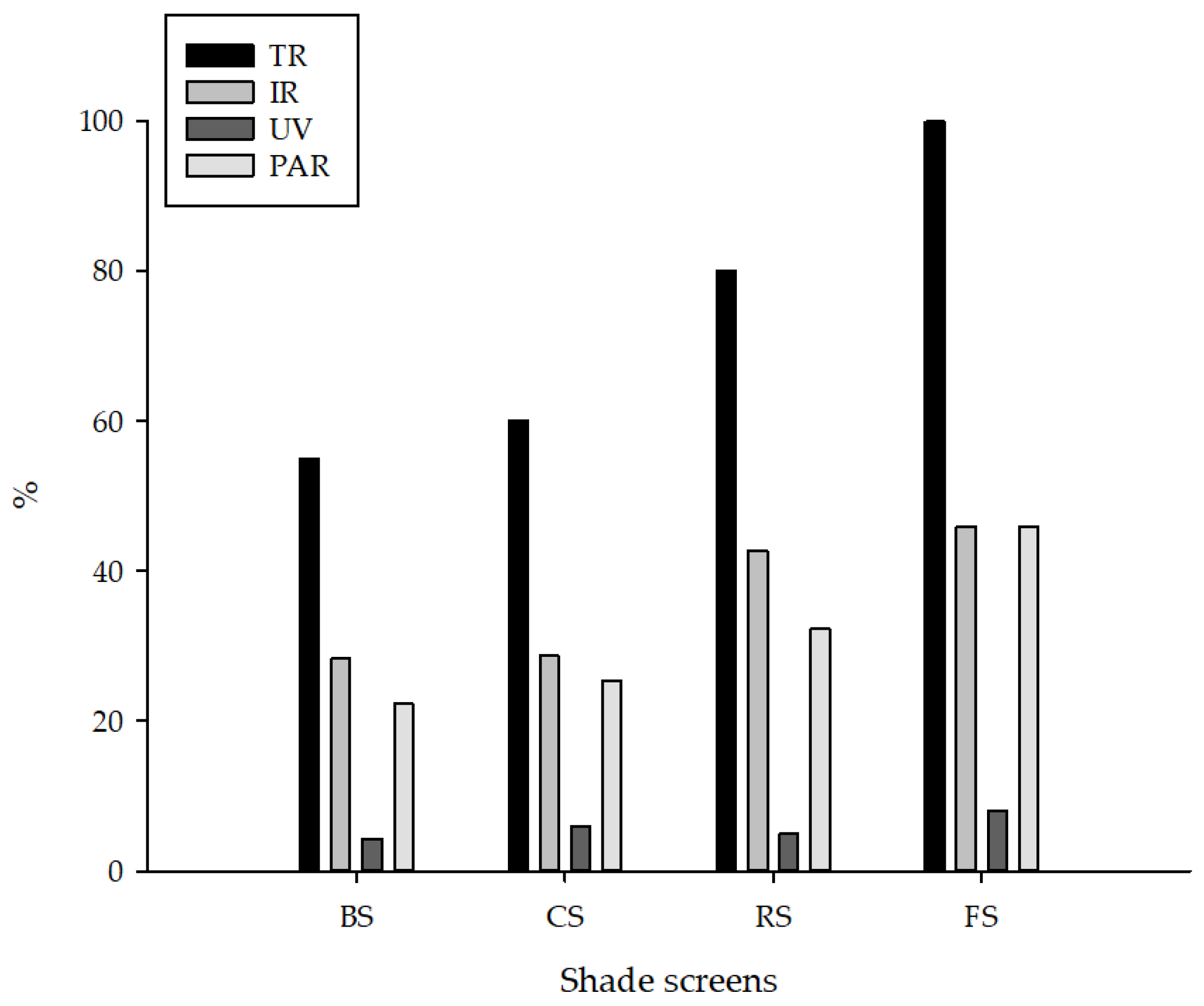
Figure 2 shows the distribution of spectral radiation under the three studied covers. Among the shades, the highest infrared radiation (IR) transmittance occurred in the environment covered with a red screen (42.72%), followed by ChromatiNet
® screen (28.69%), and the lowest IR in the environment with a black screen (28.39%). While the highest transmittance of ultraviolet radiation occurred on ChromatiNet
® (5.91%) and the lowest on red screen (4.98%). For photosynthetically active radiation transmittance, the highest value was observed in the environment with red screen (42.72%), followed by ChromatiNet
® screen (28.69%) and black screen (28.39%).
The significant interaction between cultivars and studied environments was observed for all variables analyzed (
Table 1). It is noticeable that there were many cultivar × environment interactions, suggesting that cultivars responded differently to environment changes for different traits.
The accumulation of carbohydrates observed by the content of total soluble sugars (TSS) and reducing sugars (RS) was higher in the Great Lakes cultivar in the full sun (FS) environment with about 78 µmol·g
−1 of commercial fresh matter (CMF) and shaded with the red screen (RS), with 6.40 µmol·g
−1, respectively (
Table 2).
As an indication of stress in lettuce plants, the activity of antioxidant enzymes was evaluated. In this context, the lettuce cultivars showed different responses to the enzymes SOD, CAT, and POD in the different environments studied (
Table 3 and
Table 4). For the enzyme SOD activity, the cultivars Delícia and Balsamo stood out in the BS and RS environments, as did the Great Lakes cultivar in RS (
Table 3). The enzyme SOD had the highest activity under BS, with 16 MJ·m
−2·day
−1 of average solar radiation, followed by CS, which presented 18 MJ·m
−2·day
−1 (
Figure 1a). Thus, the environments with solar radiation values higher than 25 MJ·m
−2·day
−1, RS and FS, inhibited the action of the SOD enzyme in the iceberg lettuce cultivars tested (
Table 3).
For CAT enzyme activity, higher values, 12 µmol H2O2·min−1.µgTP−1, were observed in the plants of the cultivars Great Lakes and Balsamo under the red screen. The highest values for POD enzyme activity, about 20 UE·min−1·gTP−1, were observed in the Great Lakes cultivar in full sun and Winslow under RS. Analyzing the activity of CAT and POD enzymes, higher values of CAT enzyme activity were observed in plants grown under BS and CS, while POD activity was higher in FS. Thus, plants grown under BS and CS modulate antioxidant response via CAT, and those grown under FS modulate via POD.
All cultivars grown in full sun were less productive than the other environments (
Table 4). The highest values were reached by cultivars Delícia and Balsamo in the CS environment, about 210 g·plant
−1, followed by the Balsamo cultivar grown in BS. The Great Lakes cultivar did not produce lettuce head in any environments studied, and, as a result is not found in
Table 4. However, even without head formation (FCP), this cultivar developed leaves, and these were used for biochemical analysis.
Analyzing the correlation between the variables (
Table 5), mainly to know which of these influences commercial fresh matter, it was possible to verify a negative correlation of this variable with the RS content and POD enzyme activity and positive between FCP and SOD activity.
4. Discussion
The distribution of spectral radiation (
Figure 2) under the three shades studied was not the main reason for the biochemical and productive alterations of the studied iceberg lettuce cultivars, since the difference between IR, UV, and PAR between the screens was small. However, the high incidence of solar radiation recorded during the period of the experiment caused changes in the biochemical aspects of the iceberg lettuce, reducing the photosynthetic efficiency of the plants and negatively interfering with all the characteristics evaluated in full sun (
Figure 1). However, it was observed that the use of shading screens reduced the intensity of radiant energy, providing better quality and distribution of solar radiation within the protected environments, resulting in better crop performance.
High photosynthetic rates promote the synthesis of sugars that reflect greater growth and storage of carbohydrates in the physiological sinks [
21]. Lettuce grown in a hydroponic system showed high contents of total sugars in the basal leaves because these leaves are functioning as the main source of photoassimilates [
22]. Therefore, the cultivar Great Lakes may have shown better photosynthetic efficiency in environments with FS and RS when compared to other screens. However, these sugars were used for bolting—or inhibiting the lettuce head formation, the commercial product of lettuce—in this cultivar.
In temperate climate conditions and winter, greater solar radiation has a beneficial effect on the greater accumulation of dry matter and total soluble sugars in lettuce plants [
23]. For tropical and semi-arid conditions, as in the present research, a more appropriate range of solar radiation is observed that allows greater liquid photosynthesis, increase in TSS, and head formation of iceberg lettuce. This behavior was evidenced in the Balsamo cultivar in environments with BS and CS, where greater commercial biomass production accompanied the highest TSS content (
Table 2).
In a study with lettuce, maximum temperature above 25 °C caused less accumulation of carbohydrates and less growth and mass at the end of the cycle [
24]. Thus, for lettuce types that do not produce head, the increase in these variables can indicate ideal cultivation conditions. On the other hand, the decline in the values of sugar and starch in lettuce leaves may be indicative of stress, caused by the restricted absorption of water and nutrients and limited photosynthesis [
24], which can reduce stomatal conductance, and, consequently, photosynthesis, thus reducing productivity.
In this research, it was not possible to observe the relationship between the reducing sugars content and indicative stress due to the restriction of photosynthesis since the plants in environments with CS and BS presented lower reducing sugars and greater production (
Table 2 and
Table 4) when compared to the FS and RS. According to [
25], global solar radiation around 20.7 MJ·m
−2·day
−1 favors the photosynthetic rate and greater production in lettuce experiments. Approximate values between 16 and 18 MJ·m
−2·day
−1 were observed for BS and CS, suggesting that this range is efficient for raising RS contents in the Balsamo cultivar and results from a better photosynthetic performance than the other cultivars studied.
A well-known consequence of high temperature in plants is a reduction in photosynthesis rate, while respiration and photorespiration rates increase considerably under high temperatures [
26]. However, one must also consider the energy balance of the environment. The increase in temperature and solar radiation leads to a rise in the environment’s vapor pressure deficit, reducing photosynthesis and increasing photorespiration, forming reactive oxygen species (ROS).
When plants are subjected to some biotic or abiotic stress, the first response observed in the plant is the drastic increase in ROS production that causes oxidative damage in proteins, lipids, and nucleic acids, characterizing oxidative stress [
27]. The first ROS formed is superoxide (•O
−2), which gives rise to the other reactive oxygen species. The •O
−2, through the enzyme SOD, is dismutated to H
2O
2 [
28]. Radiation causes the expression of defense genes, initiating ROS generation by the enzyme nicotinamide adenine dinucleotide phosphate (NADPH) oxidase on the plasma membrane cytoplasm; these species are formed in mitochondria and chloroplasts [
29]. Regulation of H
2O
2 levels occurs through a variety of enzymes, such as ascorbate peroxidase (APX), catalase (CAT), peroxidase (POD), glutathione S-transferase (GST), glutathione reductase (GR), and glutathione peroxidase (GPX) [
30].
Considering the environments in this study, solar radiation was responsible for modulating the activity response of the antioxidant enzymes SOD, CAT, and POD in lettuce plants (
Table 3 and
Table 4) since the temperature and relative air humidity did not experience significant changes due to the use of shading screens (
Figure 1b). The cultivars studied showed different responses, showing tolerance to high solar radiation environments for expressing genes for activating antioxidative metabolism.
Regarding the reduction in the harmful effects of H
2O
2, the action of the CAT enzyme is similar to that of some other enzymes, such as peroxidases. However, CAT is more effective in relatively high concentrations of H
2O
2 (mM) in a condition of severe stresses since CAT does not need a reducing agent, being efficient for the removal of H
2O
2 [
31].
Solar radiation may have modulated the antioxidant system’s response, in that under higher values of this microclimatic factor, the activation of POD occurred, while with lower values, CAT activation occurred. The average temperature of the shaded environments and the full sun was higher than that recommended to cultivate iceberg lettuce (
Figure 1b). This thermal stress led to an increase in ROS production, which, due to the action of the SOD enzyme, may have been converted into H
2O
2. In environments with higher radiation, POD acted for the conversion of H
2O
2 (
Table 4), while in environments with less radiation, CAT was the main reason for this conversion (
Table 3). CAT is more efficient for reducing the effects of H
2O
2; thus, cultivars under CS and BS, with less solar radiation, may have presented less damage to the cell membrane and, consequently, better responses to the formation of commercial matter (
Table 4).
Weiguo et al. [
11] observed that the POD enzyme action was more evident under a condition of greater light intensity when compared to CAT in lettuce plants. These same authors also observed that the excess of energy of light intensity, 600 µmol·m
−2·s
−1 led to the production of ROS and a relatively substantial increase in lipid peroxidation. In a second assessment, the activities of SOD, CAT, and POD increased; a certain proportion of ROS was eliminated; and lipid peroxidation decreased.
Thus, when the solar radiation is between 16 and 18 MJ·m
−2·day
−1, as observed in environments with BS, the enzyme SOD activity increases. There may also be a reduction in damage to the cell membrane. There was also no photoinhibition in the cultivars Balsamo, Winslow, and Delícia, leading to greater accumulation of biomass and commercial production in these environments (
Table 4).
The microclimate generated by shading screens in the environments, with lower average temperature and solar radiation, provided a 241% increase in the commercial fresh matter for Delicia cultivar and 254% for the Balsamo cultivar compared to CS and FS. A higher fresh matter of iceberg lettuce was observed in a shaded environment with a 37% reduction in solar radiation when compared to the full sun. The shaded environments provide excellent light conditions in the summer production of this vegetable [
32]. However, the Great Lakes cultivar did not produce lettuce head at all under any environment conditions, indicating that lettuce is not productive in tropical regions, regardless of what kind of shading method is used.
The positive relationship between the activity of the SOD enzyme and FCP can be related to the role of this enzyme in protecting plant cells from the harmful consequences of the increase in respiratory metabolism caused by the high temperature observed during this study. This enzyme is active in the first line of antioxidative defense by dismuting the superoxide radical to hydrogen peroxide and molecular oxygen [
33]. Although not correlated with FCP, the CAT enzyme is considered a crucial enzyme to prevent the effects caused by stress due to its high catalytic activity, acting in the removal of hydrogen peroxide (H
2O
2) and, consequently, reducing lipid peroxidation and damage to cell membranes [
34].
As for the negative relationship between POD activity and FCP, this effect may be related to this enzyme’s ability to reduce H
2O
2, which may not have been efficient enough to reduce abiotic stress and increase commercial fresh matter values of lettuce. It is known that the activity of the POD enzyme is not as efficient as CAT; for example, the authors of [
35] point out that CAT is the main eliminating enzyme of this reactive oxygen species.
Thus, through the correlation analysis, it is possible to affirm that the high content of reducing sugars is used as an energy source for the bolting of iceberg lettuce, which leads to no head formation, mainly in the Great Lakes cultivar. For antioxidant enzymes, SOD has a positive relationship with FCP due to the detoxification function of ROS, which reduces stress and allows a greater accumulation of commercial fresh matter. The POD, having a lower capacity to detoxify H2O2, did not present a positive correlation with the FCP.
As for the cultivars, there was a different answer for each of them in each different environment, with this variation resulting from the adaptive capacity of the genetic material used; even though all are indicated for tropical conditions, full sun results in only 80 g.plant−1 of FCP. The cultivars Balsamo and Delícia can be indicated, as they present the highest yields when cultivated in environments covered with ChromatiNet®.
The average solar radiation in South America between the parallels 15° N and 15° S is 250 Wm
−2 or 21.6 MJ·m
−2·day
−1 [
36], above those found in this study (
Figure 1a) for growing lettuce in high-temperature conditions. Solar radiation is an important meteorological component for iceberg lettuce production, and values above 20.7 MJ·m
−2·day
−1 reduced commercial production and the activity of antioxidant enzymes, and increased iceberg lettuce bolting in this study. The use of shading screens is a technology that must be used to make the cultivation of this vegetable feasible in certain regions.


 ,
, 



{kind=link}
{kind=link}