
Effects of Aged Biochar on Remediation of Cd-Contaminated Soil and Greenhouse Gas Emission in Chinese Cabbage (Brassica chinensis L.) Growth

 ,
, 


Abstract
1. Introduction
2. Material and Methods
2.1. Preparation of Biochar
2.1.1. Preparation of Pristine Biochar
2.1.2. Preparation of Aged Biochar
2.2. Experimental Setup
2.3. Greenhouse Gas Emission Determination
2.4. Biochar Characterization
2.5. Soil Property Characterization
2.6. Illumina-Based 16S rRNA Sequencing for Identification of Microbial Species
2.7. Statistical Analyses
3. Results
3.1. Selected Properties of Biochar
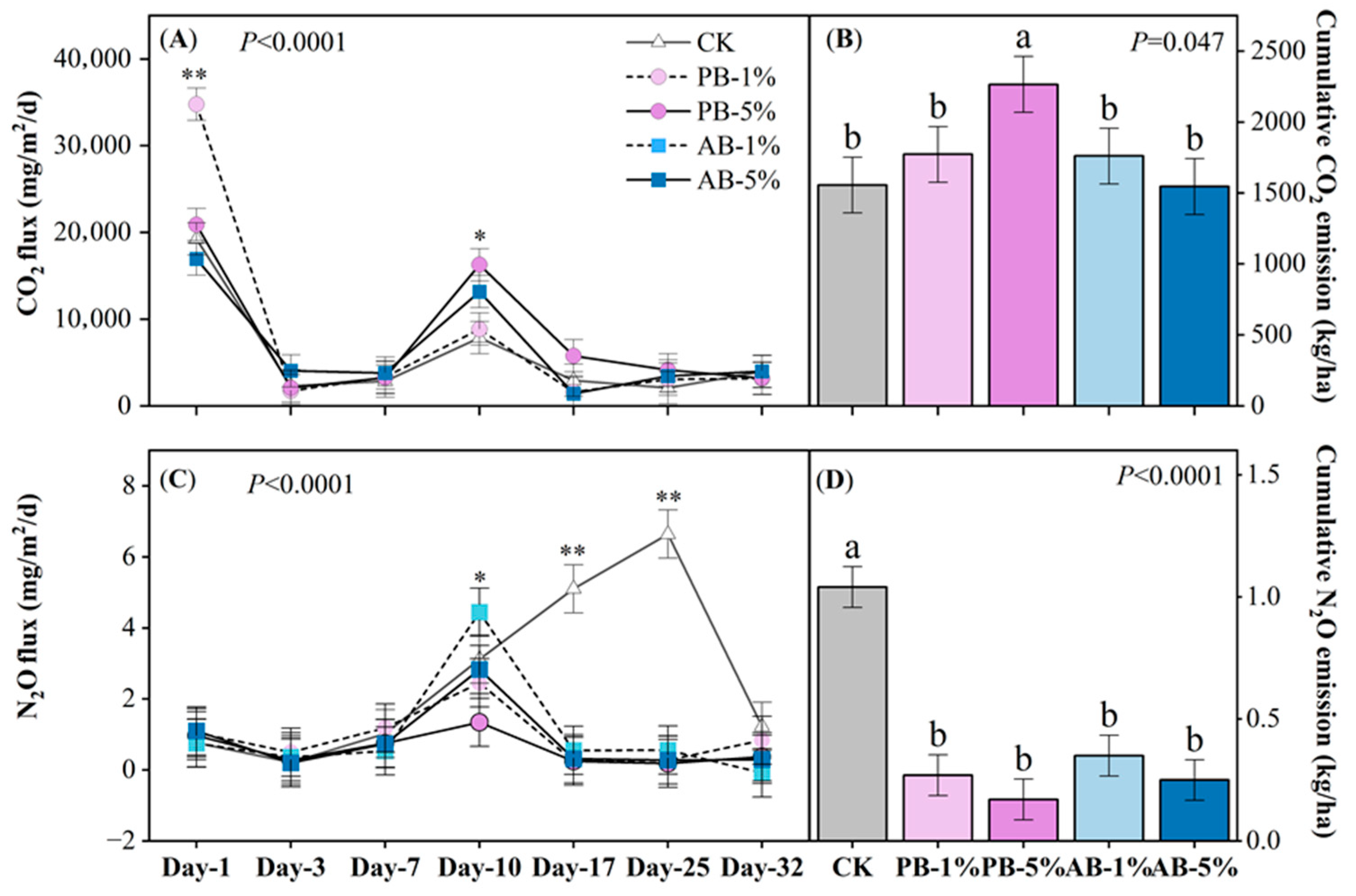
3.2. Soil CO2 and NO2 Emissions
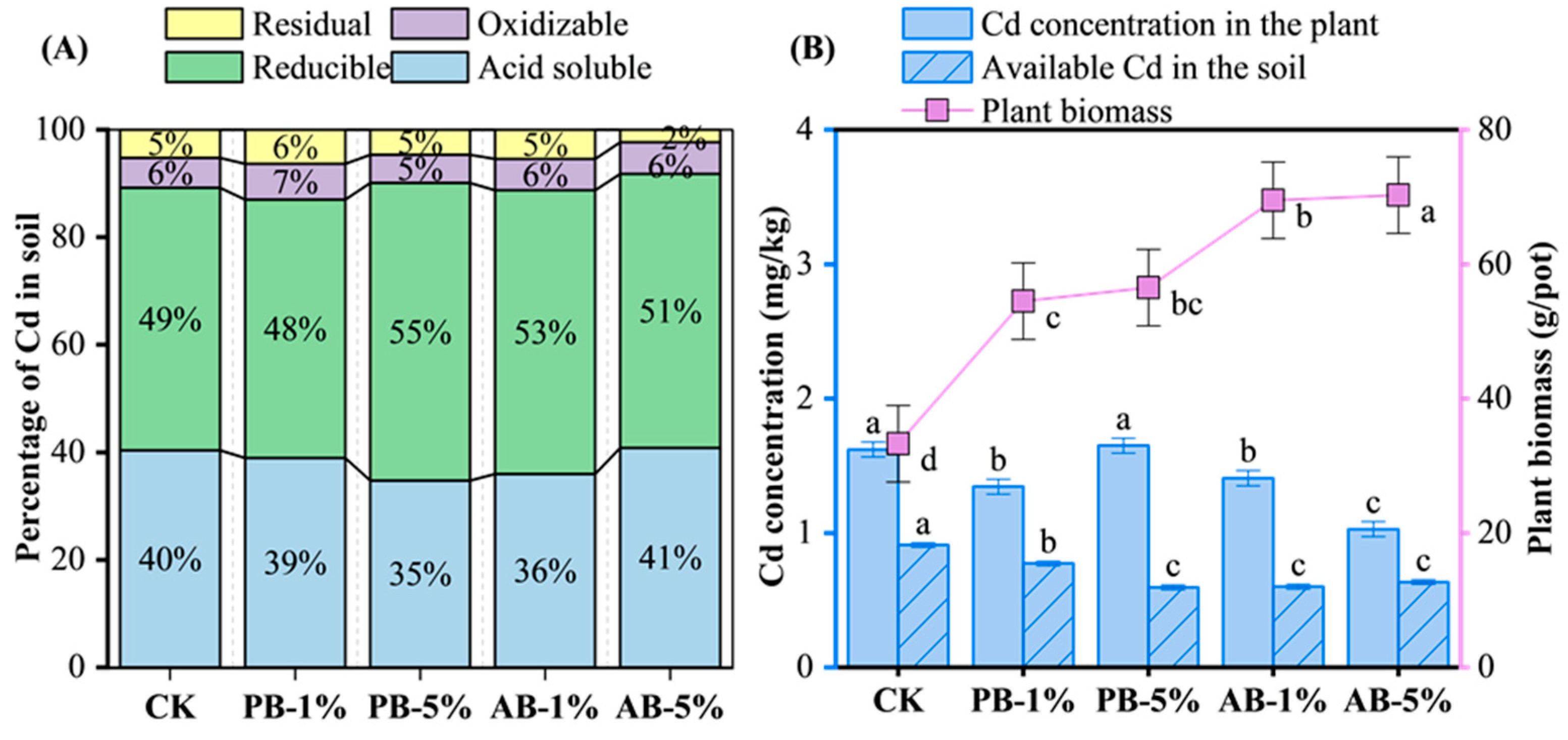
3.3. Soil Cd, Plant Cd Uptake, and Plant Biomass
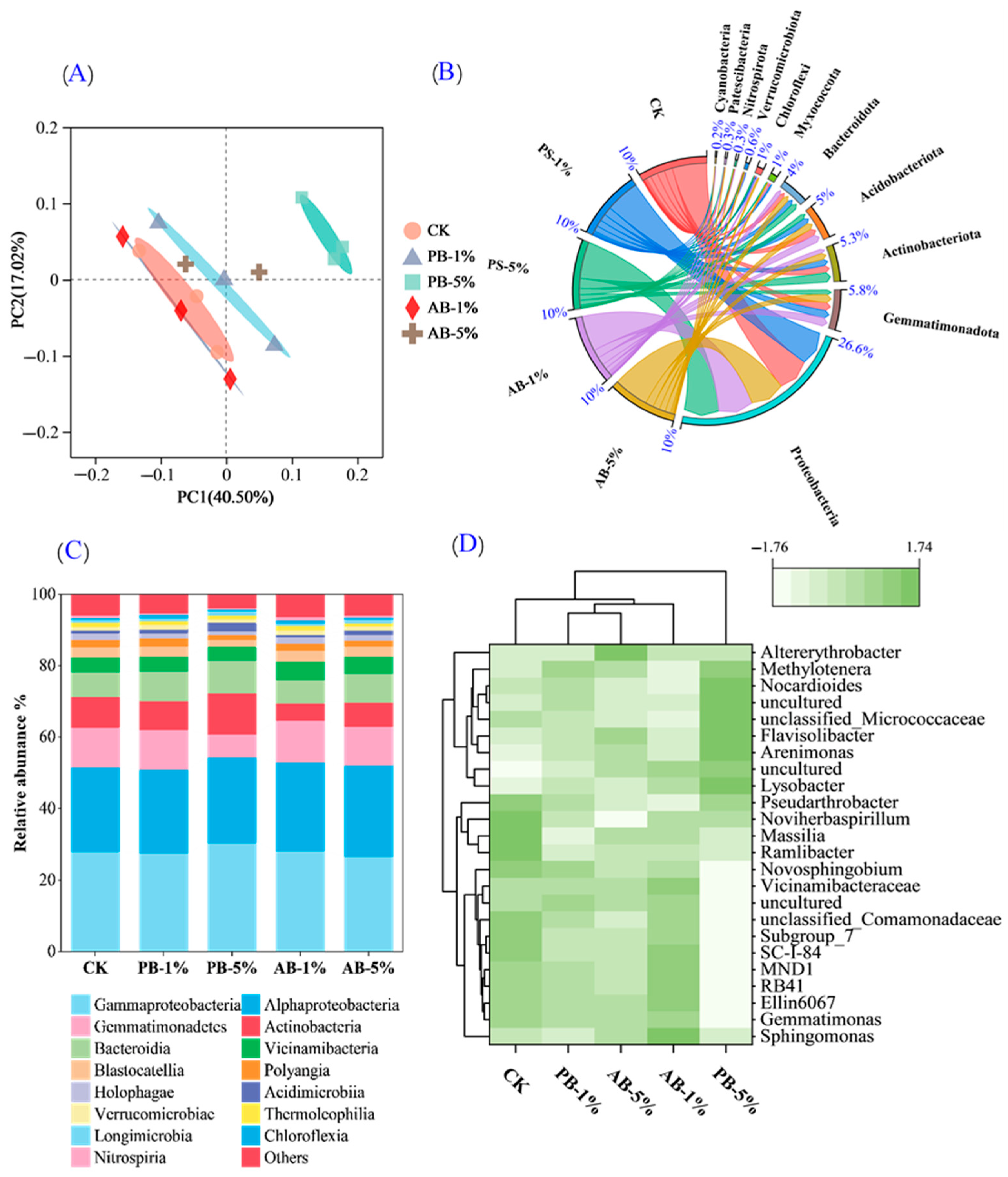
3.4. Changes in Soil Microbial Communities and Their Life Strategies
3.5. Relationship Between Soil Factors and Plant Biomass, Plant Cd, and Greenhouse Gas Emission
4. Discussion
4.1. Greenhouse Gas Emissions as Affected by Biochar Application
4.2. Soil Cd Content and Plant Cd Uptake as Affected by Biochar Application
4.3. Linking Microbial Communities with Greenhouse Gas Emission in Biochar-Treated Soils
4.4. Linking Microbial Communities with Cd Remediation in Biochar-Treated Soils
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lehmann, J.; Joseph, S. Biochar for Environmental Management: An Introduction. In Biochar for Environmental Management: Science, Technology and Implementation; Lehmann, J., Joseph, S., Eds.; Taylor and Francis: London, UK, 2015; pp. 1–13. [Google Scholar] [CrossRef]
- Duan, R.; Ma, S.; Ma, Y.; Xu, S.; Li, G.; Fu, H.; Wu, X.; Du, J.; Zhao, P. Efficient inactivation of antibiotic resistant bacteria by iron-modified biochar and persulfate system: Potential for controlling antimicrobial resistance spread and mechanism insights. J. Hazard. Mater. 2025, 492, 138182. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Jiang, M.; He, L.; Zhang, Z.; Gustave, W.; Vithanage, M.; Niazi, N.K.; Chen, B.; Zhang, X.; Wang, H.; et al. Challenges in safe environmental applications of biochar: Identifying risks and unintended consequence. Biochar 2025, 7, 12. [Google Scholar] [CrossRef]
- Yang, X.; Dai, Z.; Ge, C.; Yu, H.; Bolan, N.; Tsang, D.C.W.; Song, H.; Hou, D.; Shaheen, S.M.; Wang, H.; et al. Multiple-functionalized biochar affects rice yield and quality via regulating arsenic and lead redistribution and bacterial community structure in soils under different hydrological conditions. J. Hazard. Mater. 2023, 443, 130308. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, W.; Ding, C.; Zhang, J.; Zhang, N.; Li, Y.C.; Gao, B.; Wang, B.; Wang, X.Z. Biochar-supported zero-valent iron enhanced arsenic immobilization in a paddy soil: The role of soil organic matter. Biochar 2024, 6, 26. [Google Scholar] [CrossRef]
- Jeewani, P.H.; Brown, R.W.; Evans, C.D.; Cook, J.; Roberts, B.P.; Fraser, M.D.; Chadwick, D.R.; Jones, D.L. Rewetting alongside biochar and sulphate addition mitigates greenhouse gas emissions and retain carbon in degraded upland peatlands. Soil Biol. Biochem. 2025, 207, 109814. [Google Scholar] [CrossRef]
- Cong, M.; Hu, Y.; Sun, X.; Yan, H.; Yu, G.; Tang, G.; Chen, S.; Xu, W.; Jia, H. Long-term effects of biochar application on the growth and physiological characteristics of maize. Front. Plant Sci. 2023, 14, 1172425. [Google Scholar] [CrossRef]
- Liu, X.; He, Y.; Li, J.; Li, J.; Zhang, J.; Tang, X. Does biochar field aging reduce the kinetic retention for weakly hydrophobic antibiotics in purple soil? Biochar 2025, 7, 69. [Google Scholar] [CrossRef]
- Song, L.; Cao, W.; Ren, Y.; Zhang, R.; Zhang, Z.; Qian, J. Aging influence and long-term stable mechanism of biofuel ash immobilizing cadmium in alkaline soil combining artificial aging simulation. Environ. Res. 2025, 277, 121570. [Google Scholar] [CrossRef]
- Cheng, H.; Xing, D.; Twagirayezu, G.; Lin, S.; Gu, S.; Tu, C.; Hill, P.W.; Chadwick, D.R.; Jones, D.L. Effects of field-aging on the impact of biochar on herbicide fate and microbial community structure in the soil environment. Chemosphere 2024, 348, 140682. [Google Scholar] [CrossRef]
- Chang, R.H.; Sohi, S.P.; Jing, F.Q.; Liu, Y.Y.; Chen, J.W. A comparative study on biochar properties and Cd adsorption behavior under effects of ageing processes of leaching, acidification and oxidation. Environ. Pollut. 2019, 254, 113123. [Google Scholar] [CrossRef]
- Li, H.; Ye, X.; Geng, Z.; Zhou, H.; Guo, X.; Zhang, Y.; Zhao, H.; Wang, G. The influence of biochar type on long-term stabilization for Cd and Cu in contaminated paddy soils. J. Hazard. Mater. 2016, 304, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Alves, B.S.Q.; Fernandes, L.A.; Southard, R.J. Biochar-cadmium retention and its effects after aging with hydrogen peroxide (H2O2). Heliyon 2021, 7, e08476. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhao, L.; Rui, X.; Hu, J.; Zhu, N. A review of pristine and modified biochar immobilizing typical heavy metals in soil: Applications and challenges. J. Hazard. Mater. 2022, 432, 128668. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gao, P.; Lu, Y.; Cui, X.; Peng, F. Hydrogen peroxide-aged biochar mitigating greenhouse gas emissions during co-composting of swine manure with rice bran. Environ. Pollut. 2025, 374, 126255. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Zhao, X.; Li, Y.; Deng, Y.; He, Q.; Dai, W. Aging alters the physicochemical properties of biochar, enhances its adsorption performance for tris-(1-chloro-2-propyl) phosphate, and changes the adsorption mechanism. Environ. Technol. Innov. 2025, 37, 104053. [Google Scholar] [CrossRef]
- Li, G.; Zhang, F.; Ma, Y.; Zhang, X.; Liu, Q.; Ye, R.; Ma, Y.; Liu, X. Enhanced stabilization of lead in soil using novel biochar composites under simulated accelerated aging conditions. Sustainability 2025, 17, 2563. [Google Scholar] [CrossRef]
- Mia, S.; Dijkstra, F.A.; Singh, B. Chapter One—Long-Term Aging of Biochar: A molecular understanding with agricultural and environmental implications. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 141, pp. 1–51. [Google Scholar] [CrossRef]
- Feng, Y.; Feng, Y.; Liu, Q.; Chen, S.; Hou, P.; Poinern, G.; Jiang, Z.; Fawcett, D.; Xue, L.; Lam, S.S.; et al. How does biochar aging affect NH3 volatilization and GHGs emissions from agricultural soils? Environ. Pollut. 2022, 294, 118598. [Google Scholar] [CrossRef]
- Zhang, K.; Gu, R.; Yang, Y.; Yan, J.; Ma, Y.; Shen, Y. Recent distribution changes of invasive Asteraceae species in China: A five-year analysis (2016–2020). J. Environ. Manag. 2025, 376, 124445. [Google Scholar] [CrossRef]
- Cai, J.-F.; Zhang, L.; Zhang, Y.; Zhang, M.-X.; Li, H.-L.; Xia, H.-J.; Kong, W.-J.; Yu, F.-H. Remediation of cadmium-contaminated coastal saline-alkaline soil by Spartina alterniflora derived biochar. Ecotoxicol. Environ. Saf. 2020, 205, 111172. [Google Scholar] [CrossRef]
- Wu, J.; Song, Q.; Zhou, J.; Wu, Y.; Liu, X.; Liu, J.; Zhou, L.; Wu, Z.; Wu, W. Cadmium threshold for acidic and multi-metal contaminated soil according to Oryza sativa L. Cadmium accumulation: Influential factors and prediction model. Ecotoxicol. Environ. Saf. 2021, 208, 111420. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.W.; Zia-ur-Rehman, M.; Zahir, Z.A.; Rinklebe, J.; Tack, F.M.G.; Ok, Y.S. A critical review on effects, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Ramzani, P.M.A.; Rasool, B.; Khan, M.A.; Rahman, M.; Akhtar, I.; Turan, V.; Tauqeer, H.M.; Farhad, M.; Khan, S.A.; et al. Efficacy of chitosan-coated textile waste biochar applied to Cd-polluted soil for reducing Cd mobility in soil and its distribution in moringa (Moringa oleifera L.). J. Environ. Manag. 2021, 284, 112047. [Google Scholar] [CrossRef] [PubMed]
- Aborisade, M.A.; Geng, H.; Oba, B.T.; Kumar, A.; Ndudi, E.A.; Battamo, A.Y.; Liu, J.; Chen, D.; Okimiji, O.P.; Ojekunle, O.Z.; et al. Remediation of soil polluted with Pb and Cd and alleviation of oxidative stress in Brassica rapa plant using nanoscale zerovalent iron supported with coconut-husk biochar. J. Plant Physiol. 2023, 287, 154023. [Google Scholar] [CrossRef] [PubMed]
- Ghani, M.I.; Ahanger, M.A.; Sial, T.A.; Haider, S.; Siddique, J.A.; Fan, R.; Liu, Y.; Ali, E.F.; Kumar, M.; Yang, X.; et al. Almond shell-derived biochar decreased toxic metals bioavailability and uptake by tomato and enhanced the antioxidant system and microbial community. Sci. Total Environ. 2024, 929, 172632. [Google Scholar] [CrossRef]
- Haider, F.U.; Ain, N.; Khan, I.; Farooq, M.; Cai, H.L.; Li, Y. Co-application of biochar and plant growth regulators improves maize growth and decreases Cd accumulation in cadmium-contaminated soil. J. Clean. Prod. 2024, 440, 140515. [Google Scholar] [CrossRef]
- Qu, J.; Shi, J.; Wang, Y.; Tong, H.; Zhu, Y.; Xu, L.; Wang, Y.; Zhang, B.; Tao, Y.; Dai, X.; et al. Applications of functionalized magnetic biochar in environmental remediation: A review. J. Hazard. Mater. 2022, 434, 128841. [Google Scholar] [CrossRef]
- Qiu, M.; Liu, L.; Ling, Q.; Cai, Y.; Yu, S.; Wang, S.; Fu, D.; Hu, B.; Wang, X. Biochar for the removal of contaminants from soil and water: A review. Biochar 2022, 4, 19. [Google Scholar] [CrossRef]
- Sachdeva, S.; Kumar, R.; Sahoo, P.K.; Nadda, A.K. Recent advances in biochar amendments for immobilization of heavy metals in an agricultural ecosystem: A systematic review. Environ. Pollut. 2023, 319, 120937. [Google Scholar] [CrossRef]
- Xu, W.; Xie, X.; Li, Q.; Yang, X.; Ren, J.; Shi, Y.; Liu, D.; Shaheen, S.M.; Rinklebe, J. Biochar co-pyrolyzed from peanut shells and maize straw improved soil biochemical properties, rice yield, and reduced cadmium mobilization and accumulation by rice: Biogeochemical investigations. J. Hazard. Mater. 2024, 466, 133486. [Google Scholar] [CrossRef]
- Cross, A.; Sohi, S.P. A method for screening the relative long-term stability of biochar. Glob. Change Biol. Bioenergy 2013, 5, 215–220. [Google Scholar] [CrossRef]
- Chen, X.; Gao, X.; Yu, P.; Spanu, L.; Hinojosa, J.; Zhang, S.; Long, M.; Alvarez, P.J.J.; Masiello, C.A. Rapid simulation of decade-scale charcoal aging in soil: Changes in physicochemical properties and their environmental implications. Environ. Sci. Technol. 2022, 57, 128–138. [Google Scholar] [CrossRef]
- GB 15618-2018; Soil Environmental Quality: Risk Control Standard for Soil Contamination of Agricultural Land. Ministry of Ecology and Environment: Beijing, China, 2018.
- Lu, Y.; Silveira, M.L.; Vendramini, J.M.B.; Erickson, J.E.; Li, Y. Biosolids and biochar application effects on bahiagrass herbage accumulation and nutritive value. J. Agron. 2020, 112, 1330–1345. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Abbas, T.; Adrees, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farooq Qayyum, M.; Nawaz, R. Residual effects of biochar on growth, photosynthesis and cadmium uptake in rice (Oryza sativa L.) under Cd stress with different water conditions. J. Environ. Manag. 2018, 206, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Alves, B.J.R.; Smith, K.A.; Flores, R.A.; Cardoso, A.S.; Oliveira, W.R.D.; Jantali, C.P.; Boddey, R.M. Selection of the most suitable sampling time for static chambers for the estimation of daily mean N2O flux from soils. Soil Biol. Biochem. 2012, 46, 129–135. [Google Scholar] [CrossRef]
- Walkley, A.J.; Black, I.A. Estimation of soil organic carbon by the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Mulvaney, R.L. Nitrogen-Inorganic Forms. In Methods of Soil Analysis, Part 3, Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpoor, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; SSSA Book Series No. 5; SSSA: Madison, WI, USA, 1996; pp. 1123–1184. [Google Scholar]
- Alsaleh, K.A.M.; Meuser, H.; Usman, A.R.A.; Al-Wabel, M.I.; Al-Farraj, A.S. A comparison of two digestion methods for assessing heavy metals level in urban soils influenced by mining and industrial activities. J. Environ. Manag. 2018, 206, 731–739. [Google Scholar] [CrossRef]
- Zhao, M.; Xu, L.; Wang, X.X.; Li, C.; Zhao, Y.; Cao, B.; Zhang, C.; Zhang, J.; Wang, J.; Chen, Y.; et al. Microplastics promoted cadmium accumulation in maize plants by improving active cadmium and amino acid synthesis. J. Hazard. Mater. 2023, 447, 130788. [Google Scholar] [CrossRef] [PubMed]
- Ure, A.M.; Quevauviller, P.; Muntau, H.; Griepink, B. Speciation of heavy metals in soils and sediments. An account of the improvement and harmonization of extraction techniques undertaken under the auspices of the BCR of the commission of the European communities. Int. J. Environ. Anal. Chem. 1993, 51, 135–151. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93, fix006. [Google Scholar] [CrossRef] [PubMed]
- Parasar, B.J.; Agarwala, N. Unravelling the role of biochar-microbe-soil tripartite interaction in regulating soil carbon and nitrogen budget: A panacea to soil sustainability. Biochar 2025, 7, 37. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef]
- Morrisey, E.M.; Mau, R.L.; Schwartz, E.; Caporaso, J.G.; Dijkstra, P.; van Gestel, N.; Koch, B.J.; Liu, C.M.; Hayer, M.; McHugh, T.A.; et al. Phylogenetic organization of bacterial activity. ISME J. 2016, 10, 2336–2340. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Change Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Pepe-Ranney, C.; Campbell, A.N.; Koechll, C.N.; Berthrong, S.; Buckley, D.H. Unearthing the ecology of soil microorganisms using a high resolution DNASIP approach to explore cellulose and xylose metabolism in soil. Front. Microbiol. 2016, 7, e703. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. SAS/STAT 9.2 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Lu, Y.; Silveira, M.L.; Cavigelli, M.; O’Connor, G.A.; Vendramini, J.M.B.; Erickson, J.E.; Li, Y.C. Biochar impacts on nutrient dynamics in a subtropical grassland soil: 2. Greenhouse gas emissions. J. Environ. Qual. 2020, 49, 1421–1434. [Google Scholar] [CrossRef]
- Zhang, Y.; Hou, W.; Chi, M.; Sun, Y.; An, J.; Yu, N.; Zou, H. Simulating the effects of soil temperature and soil moisture on CO2 and CH4 emissions in rice straw-enriched paddy soil. Catena 2020, 194, 104677. [Google Scholar] [CrossRef]
- Lu, Y.; Silveira, M.L.; Vendramini, J.M.B.; Li, Y.C. Biochar impacts on soil nitrogen and carbon dynamics in a Spodosol amended with biosolids and inorganic fertilizer. J. Environ. Qual. 2023, 52, 999–1010. [Google Scholar] [CrossRef]
- Lu, Y.; Silveira, M.L.; O’Connor, G.A.; Vendramini, J.M.B.; Erickson, J.E.; Li, Y.C. Assessing the impacts of biochar and fertilizer management strategies on N and P balances in subtropical pastures. Geoderma 2021, 394, 115038. [Google Scholar] [CrossRef]
- Hamamoto, T.; Nhamo, N.; Chikoye, D.; Mukumbuta, I.; Uchida, Y. Responses of CO2 emissions and soil microbial community structures to organic amendment in two contrasting soils in Zambia. Sci. Rep. 2022, 12, 6368. [Google Scholar] [CrossRef]
- Wang, L.; O’Connor, D.; Rinklebe, J.; Ok, Y.S.; Tsang, D.C.W.; Shen, Z.; Hou, D. Biochar aging: Mechanisms, physicochemical changes, assessment, and implications for field applications. Environ. Sci. Technol. 2020, 54, 14797–14814. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, X.; Dong, Y.; Xu, X.; Xiong, Z. Microbial explanations for field-aged biochar mitigating greenhouse gas emissions during a rice-growing season. Environ. Sci. Pollut. Res. 2018, 25, 31307–31317. [Google Scholar] [CrossRef] [PubMed]
- Le Mer, J.; Roge, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Wang, M.; Fu, Y.; Wang, Y.; Li, Y.; Shen, J.; Liu, X.; Wu, J. Pathways and mechanisms by which biochar application reduces nitrogen and phosphorus runoff losses from a rice agroecosystem. Sci. Total Environ. 2021, 797, 149193. [Google Scholar] [CrossRef]
- Fan, J.; Duan, T.; Wu, X.; Liao, M.; Sun, J. Can the aging process necessarily weaken the effect of biochar on cadmium-contaminated soil remediation: Considering biochar at different pyrolysis temperatures and aging treatment. Environ. Geochem. Health 2025, 47, 66. [Google Scholar] [CrossRef]
- Saikia, S.; Prit, H.; Maturi, K.C.; Kalamdhad, A.S. Amendment of nutrient-deficient alluvial soil by municipal solid waste char and compost for the remediation of contaminants and enhancement of plant growth. Water Air Soil Pollut. 2025, 236, 238. [Google Scholar] [CrossRef]
- Zhang, Z.; Rengel, Z.; Chang, H.; Meney, K.; Pantelic, L.; Tomanovic, R. Bioremediation potential of Juncus subsecundus in soils contaminated with cadmium and polynuclear aromatic hydrocarbons (PAHs). Geoderma 2012, 175–176, 1–8. [Google Scholar] [CrossRef]
- Hao, X.; Bai, L.; Liu, X.; Zhu, P.; Liu, H.; Xiao, Y.; Geng, J.; Liu, Q.; Huang, L.; Jiang, H. Cadmium speciation distribution responses to soil properties and soil microbes of plow layer and plow pan soils in cadmium-contaminated paddy fields. Front. Microbiol. 2021, 12, 774301. [Google Scholar] [CrossRef]
- Xing, D.; Cheng, H.; Ning, Z.; Liu, Y.; Lin, S.; Li, Y.; Wang, X.; Hill, P.; Chadwick, D.; Jones, D.L. Field aging declines the regulatory effects of biochar on cadmium uptake by pepper in the soil. J. Environ. Manag. 2022, 321, 115832. [Google Scholar] [CrossRef]
- Zheng, N.; Yu, Y.; Li, Y.; Ge, C.; Chapman, S.J.; Yao, H. Can aged biochar offset soil greenhouse gas emissions from crop residue amendments in saline and non-saline soils under laboratory conditions? Sci. Total Environ. 2022, 806, 151256. [Google Scholar] [CrossRef]
- Liao, X.; Chen, Y.; Hi, J.; Zhang, C.; Mao, S.; Ruan, H.; Malghani, S. Effects of fresh and aged biochar on N2O emission from a poplar plantation soil. Pedosphere 2025, 35, 435–447. [Google Scholar] [CrossRef]
- Yan, R.; Wu, H.; Yang, X.; Yang, C.; Lyu, H.; Zhang, H.; Li, S.; Liu, T.; Li, R.; Yao, Y. Soil decreases N2O emission and increases TN content during combined composting of wheat straw and cow manure by inhibiting denitrification. Chem. Eng. J. 2023, 477, 147306. [Google Scholar] [CrossRef]
- Tian, W.; Liang, F.; Tu, L.; Xu, Z.; Li, R.; Ma, R.; Huang, Y.; Wu, S.; Li, S.; Wang, J.; et al. Mineral fertilizer substitution and application of Bacillus velezensis SQR9 reduced nitrogen-oxide emissions in tropical vegetable fields. Agricu. Ecosyst. Environ. 2025, 384, 109554. [Google Scholar] [CrossRef]
- Tang, B.; Wu, L.; Wang, J.; Sun, W.; Zhao, Y.; Liu, F. Separation of heat-stable antifungal factor from Lysobacter enzymogenes fermentation broth via photodegradation and macroporous resin adsorption. Front. Microbiol. 2021, 13, 663065. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, D.; Song, M.; Guan, G.; Sun, Y.; Li, J.; Cheng, X.; Luo, C.; Zhang, G. The positive role of root decomposition on the bioremediation of organic pollutants contaminated soil: A case study using PCB-9 as a model compound. Soil Biol. Biochem. 2022, 171, 108726. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, W.; Fan, X.; Gao, H.; Xu, X.; Liu, C.; Chai, Y.; Zhang, M.; Drosos, M.; Shan, S. The molecular composition of soil organic matter is regulated by bacterial community under biochar application. Geoderma 2025, 457, 117308. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, G.; Robinson, D.; Yang, Z.; Guo, J.; Xie, J.; Fu, S.; Zhou, L.; Yang, Y. Large amounts of easily decomposable carbon stored in subtropical forest subsoil are associated with r-strategy-dominated soil microbes. Soil Biol. Biochem. 2016, 95, 233–242. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, K.; Wang, Y.; Zhang, B.; Mao, K.; Zhang, H. Health risk assessments and microbial community analyses of groundwater from a heavy metal-contaminated site in Hezhou city, Southwest China. Int. J. Environ. Res. Public Health 2023, 20, 604. [Google Scholar] [CrossRef]
- Bandara, T.; Krohn, C.; Jin, J.; Chathurika, J.B.A.J.; Franks, A.; Xu, J.; Potter, I.D.; Tang, C. The effects of biochar aging on rhizosphere microbial communities in cadmium-contaminated acid soil. Chemosphere 2022, 303, 135153. [Google Scholar] [CrossRef]
- Niu, H.; Leng, Y.; Li, X.; Yu, Q.; Wu, H.; Gong, J.; Li, H.; Chen, K. Behaviors of cadmium in rhizosphere soils and its interaction with microbiome communities in phytoremediation. Chemosphere 2021, 269, 128765. [Google Scholar] [CrossRef] [PubMed]
- Bankar, A.; Nagaraja, G. Chapter 18—recent trends in biosorption of heavy metals by Actinobacteria. In New and Future Developments in Microbial Biotechnology and Bioengineering; Singh, B.P., Gupta, V.K., Passari, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 257–275. [Google Scholar] [CrossRef]
- Yan, H.; Cong, M.; Hu, Y.; Qiu, C.; Yang, Z.; Tang, G.; Xu, W.; Zhu, X.; Sun, X.; Jia, H. Biochar-mediated changes in the microbial communities of rhizosphere soil alter the architecture of maize roots. Front. Microbiol. 2022, 4, 1023444. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, Z.; Li, Y.; Liu, J.; Liu, C.; Chai, Y.; Ai, C.; Hussain, Q.; Drosos, M.; Shan, S. Effects of rice straw biochar application rates on soil aggregate biogeochemistry and linkages to microbial community structure and enzyme activities. Soil Tillage Res. 2025, 252, 106589. [Google Scholar] [CrossRef]
- He, T.T.; Hao, X.; Jing, R.; Li, Z.; Chen, Y.; Yang, M.; Gu, C.; Wang, Y.; Lin, R.; Zheng, X. Pig bedding biochar enhances the Cd extraction capacity of Perilla frutescens by mitigating the toxicity of Cd-contaminated soil. J. Environ. Sci. 2025, 156, 684–698. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Huang, D.; Zhou, W.; Xiao, R.; Chen, H.; Huang, H.; Xu, W.; Wang, G.; Li, R. Higher remediation efficiency of Cd and lower CO2 emissions in phytoremediation systems with biochar application. Environ. Pollut. 2025, 376, 126345. [Google Scholar] [CrossRef]
- Lu, X.; Sun, J.; Pan, G.; Qi, W.; Zhang, Z.; Xing, J.; Gao, Y. Ball-milling-modified biochar with additives enhances soil Cd passivation, increases plant growth and restrains Cd uptake by Chinese cabbage. Horticulturae 2025, 11, 168. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Total C | Total N | NH4-N | NO3-N | Total Cd | Organic Matter |
|---|---|---|---|---|---|---|
| - | ------------–------- | --------------------mg/kg------------- | -----g/kg----- | |||
| 6.5 | 0.5 | 0.07 | 4.8 | 35.9 | 0.0 | 8.9 |
| Property | Unit | Pristine Biochar | Aged Biochar |
|---|---|---|---|
| pH | - | 8.8 | 8.1 |
| EC | μS/cm | 4.7 | 37.9 |
| Specific surface area | m2/g | 24.8 | 12.5 |
| Total pore volume | cm3/g | 0.01 | 0.01 |
| Pore size | Å | 20.3 | 32.2 |
| C | % | 76.6 | 52.0 |
| H | % | 2.5 | 3.9 |
| N | % | 0.6 | 0.4 |
| S | % | 0.1 | 0.2 |
| C/N | - | 137.1 | 124.3 |
| C/H | - | 30.9 | 13.3 |
| Total Cd | mg/kg | 0.0 | 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Zhao, X.; Li, Y.; Li, G.; Wu, G.; Wang, Q.; Li, J.; Du, D. Effects of Aged Biochar on Remediation of Cd-Contaminated Soil and Greenhouse Gas Emission in Chinese Cabbage (Brassica chinensis L.) Growth. Horticulturae 2025, 11, 800. https://doi.org/10.3390/horticulturae11070800
Lu Y, Zhao X, Li Y, Li G, Wu G, Wang Q, Li J, Du D. Effects of Aged Biochar on Remediation of Cd-Contaminated Soil and Greenhouse Gas Emission in Chinese Cabbage (Brassica chinensis L.) Growth. Horticulturae. 2025; 11(7):800. https://doi.org/10.3390/horticulturae11070800
Chicago/Turabian StyleLu, Yanyan, Xiaoyi Zhao, Yuxuan Li, Guanlin Li, Guizhu Wu, Qianwu Wang, Jian Li, and Daolin Du. 2025. "Effects of Aged Biochar on Remediation of Cd-Contaminated Soil and Greenhouse Gas Emission in Chinese Cabbage (Brassica chinensis L.) Growth" Horticulturae 11, no. 7: 800. https://doi.org/10.3390/horticulturae11070800
APA StyleLu, Y., Zhao, X., Li, Y., Li, G., Wu, G., Wang, Q., Li, J., & Du, D. (2025). Effects of Aged Biochar on Remediation of Cd-Contaminated Soil and Greenhouse Gas Emission in Chinese Cabbage (Brassica chinensis L.) Growth. Horticulturae, 11(7), 800. https://doi.org/10.3390/horticulturae11070800