

Exploring Salinity Tolerance in Three Halophytic Plants: Physiological and Biochemical Responses to Agronomic Management in a Half-Strength Seawater Aquaponics System

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Aquaculture
2.2. Plant Cultivation
2.3. Experimental Design and Treatments
2.4. Rooting Media Management
2.5. Plant Biometric Measurements
2.6. Cation and Anion Content in Plant Aqueous Extracts
2.7. Leaf Osmotic Potential
2.8. Chlorophyll and Betalain Content
2.9. Oxidative Stress Markers
2.9.1. Leaf MDA Content
2.9.2. Leaf Hydrogen Peroxide (H2O2) Content
2.10. Activities of Antioxidant Enzymes
2.10.1. Crude Enzyme Extraction
2.10.2. CAT, APX, GR, and SOD Activities
2.10.3. Protein Assay
2.11. Statistical Analysis
3. Results
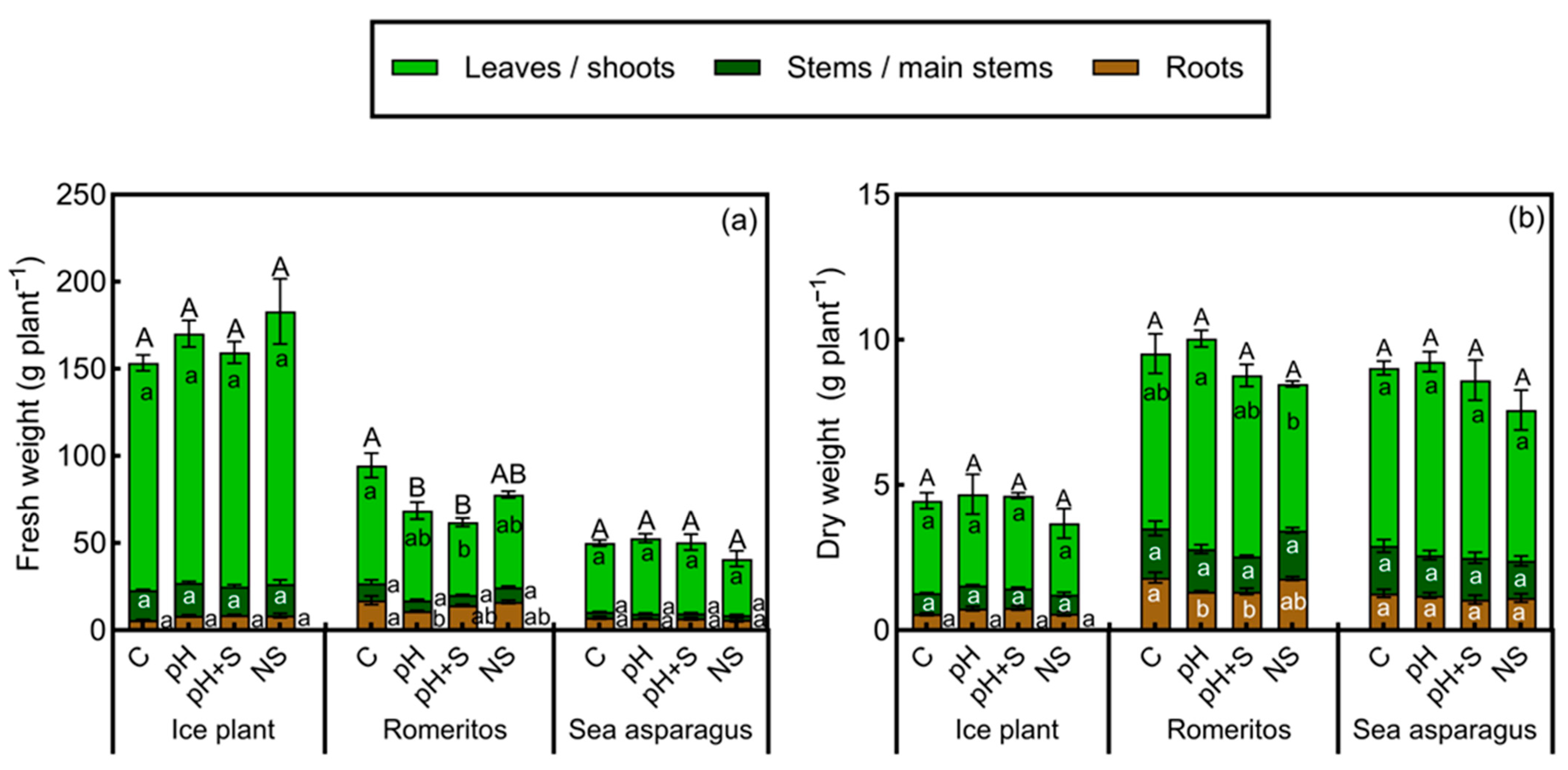
3.1. Growth Parameters
3.2. Cation and Anion Content in Leaves or Shoots
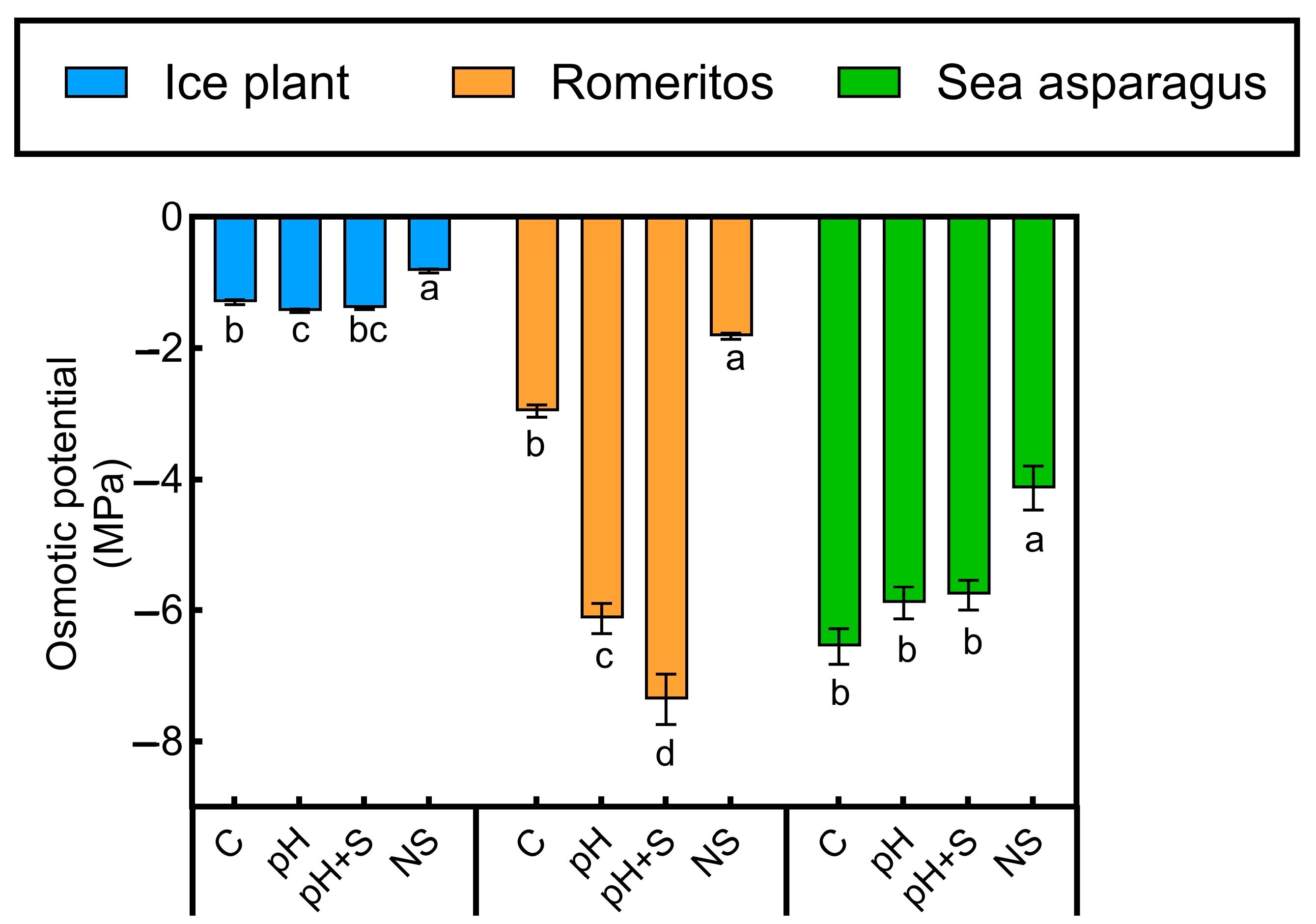
3.3. Osmotic Potential Response
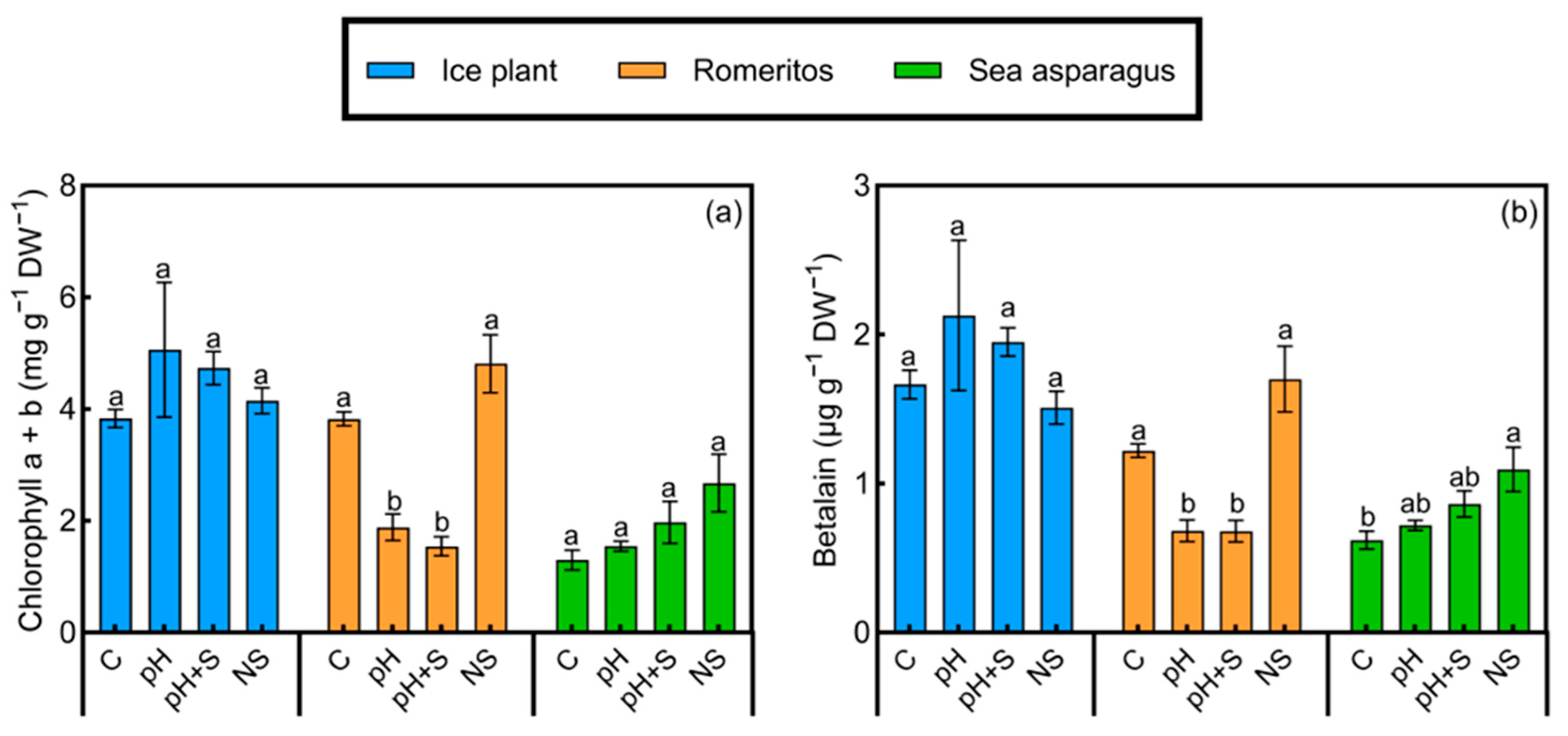
3.4. Pigment Response
3.5. Malondialdehyde and Hydrogen Peroxide Levels
3.6. Antioxidant Enzyme Responses
3.7. Overall Effects
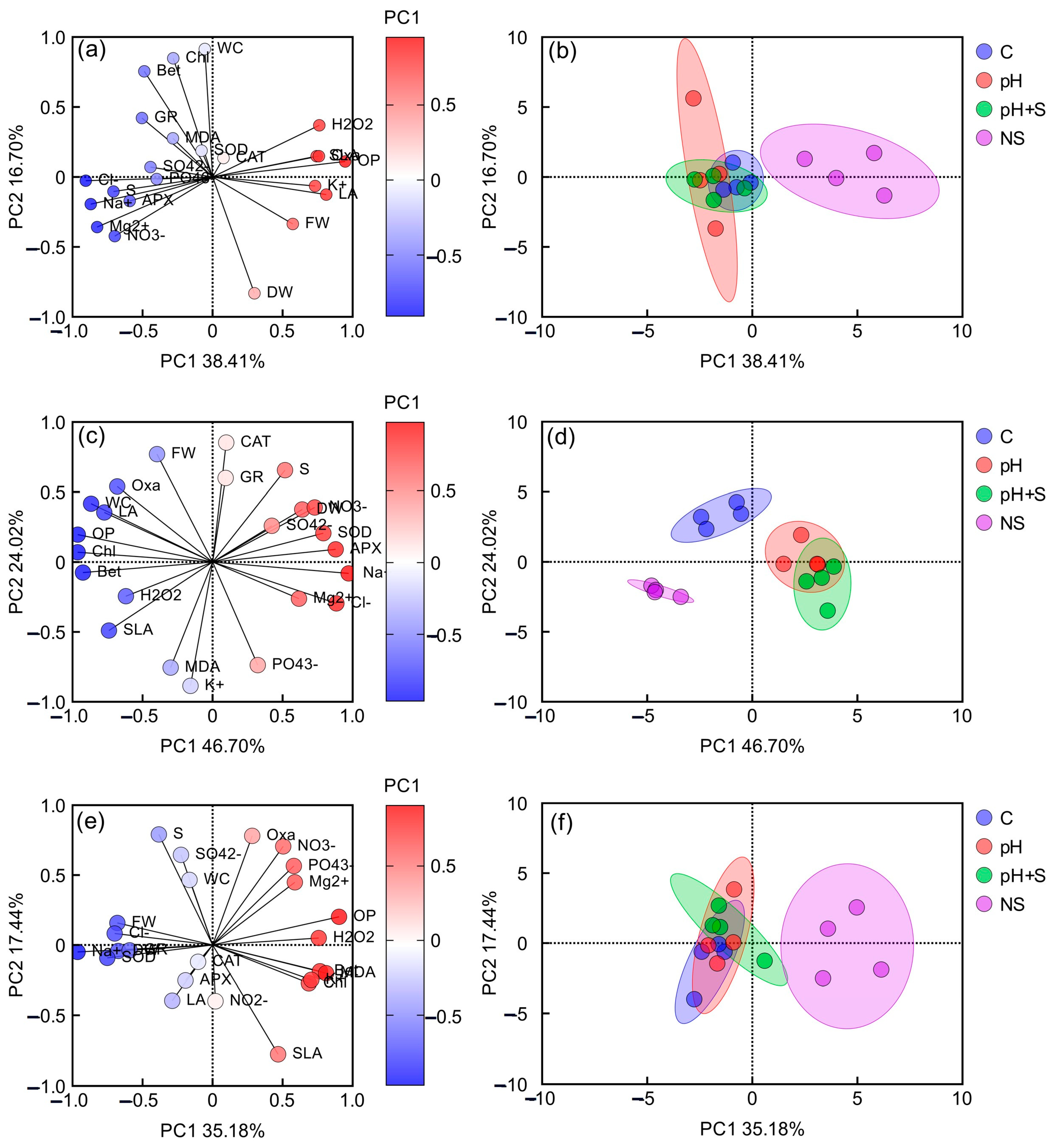
3.7.1. Principal Component Analysis
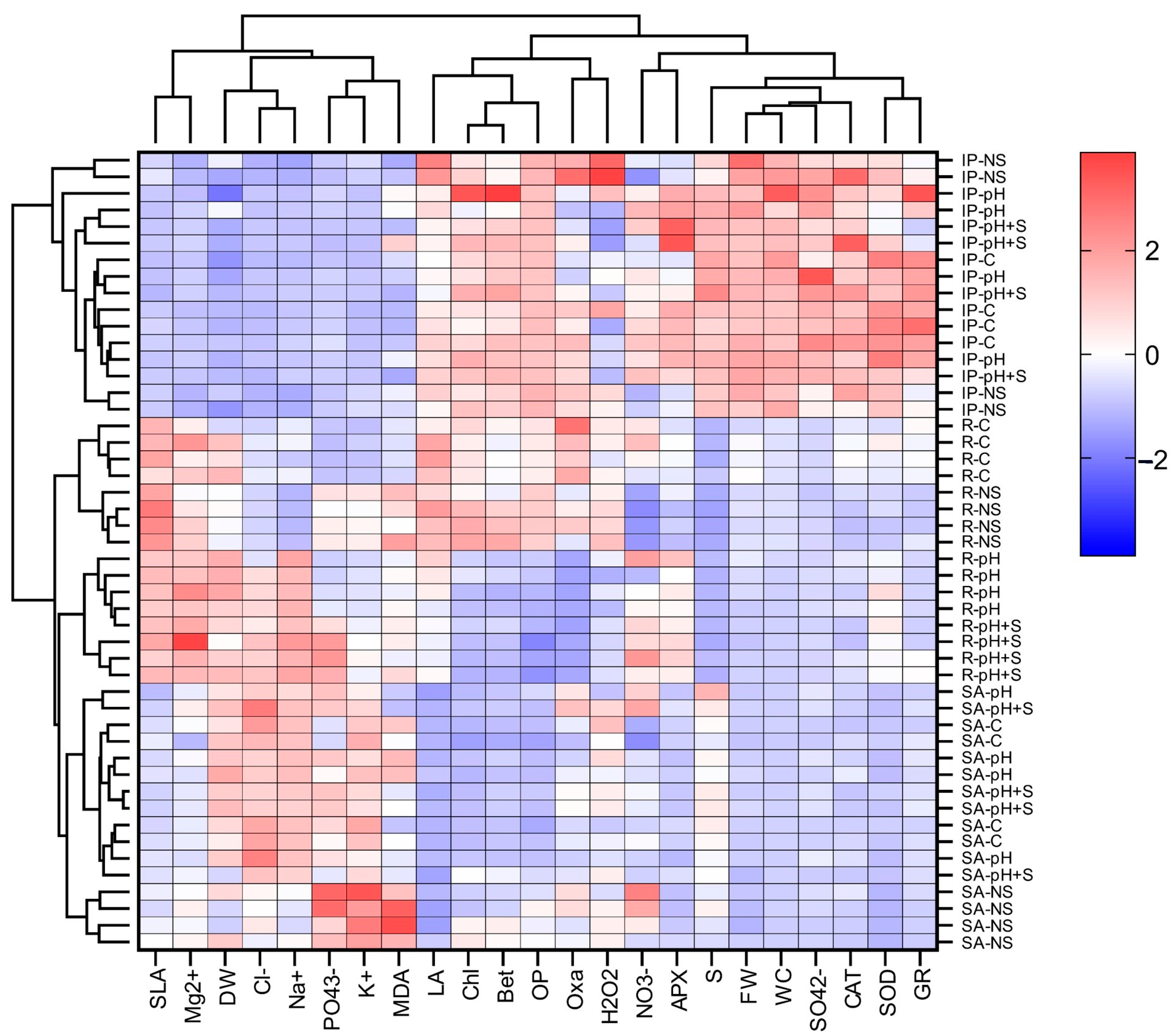
3.7.2. Hierarchical Clustering
4. Discussion
4.1. Growth Parameters and Water Relations
4.2. Ionic Pool and Osmotic Regulation
4.3. Pigments, Stress Markers, and Antioxidant Responses
4.4. General Outcomes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, K.; Noori, S.A.S.; Darbandi, A.I.; Akbari, A. Molecular mechanisms of plant salinity tolerance: A review. Aust. J. Crop Sci. 2015, 9, 321–336. [Google Scholar]
- Abobatta, W.F. Plant responses and tolerance to extreme salinity: Learning from halophyte tolerance to extreme salinity. In Salt and Drought Stress Tolerance in Plants: Signaling Networks and Adaptive Mechanisms; Hasanuzzaman, M., Tanveer, M., Eds.; Springer: Cham, Switzerland, 2020; pp. 177–210. [Google Scholar] [CrossRef]
- Hassanuzzaman, M.; Tanveer, M. Salt and Drougth Stress Tolerance in Plants: Signaling Networks and Adaptive Mechanisms; Springer: Cham, Switzerland, 2020; pp. i–x. [Google Scholar]
- Jacoby, B. Mechanisms involved in salt tolerance of plants. In Handbook of Plant and Crop Stress, 2nd ed.; Pessarakli, M., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1999; pp. 97–124. [Google Scholar]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Yeo, A.R. Breeding for salinity resistance in crop plants: Where next? Aust. J. Plant Physiol. 1995, 22, 875–884. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Munns, R.; Schachtman, D.P.; Condon, A.G. The significance of a two-phase growth response to salinity in wheat and barley. Aust. J. Plant Physiol. 1995, 22, 561–569. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Rozema, J.; Schat, H. Salt tolerance of halophytes, research questions reviewed in the perspective of saline agriculture. Environ. Exp. Bot. 2013, 82, 83–95. [Google Scholar] [CrossRef]
- Nikalje, G.C.; Srivastava, A.K.; Pandey, G.K.; Suprasanna, P. Halophytes in biosaline agriculture: Mechanism, utilization and value addition. Land Degrad. Dev. 2017, 29, 1081–1095. [Google Scholar] [CrossRef]
- Lokhande, V.H.; Suprasanna, P. Prospects of halophytes in understanding and managing abiotic stress tolerance. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 29–56. [Google Scholar]
- Mahmoud, A.H. Production of quinoa (Chenopodium quinoa) in the marginal environments of south mediterranean region: Nile Delta, Egypt. Egypt J. Soil Sci. 2017, 57, 329–337. [Google Scholar] [CrossRef]
- Attia-Ismail, S. Nutritional and feed value of halophytes and salt tolerant plants. In Halophytic and Salt-Tolerant Feedstuffs, Impacts on Nutrition, Physiology and Reproduction of Livestock; El shaer, H.M., Squires, V.R., Eds.; CRC Press: New York, NY, USA, 2015; Volume 106, pp. 106–126. [Google Scholar] [CrossRef]
- Ahmed, F.A.; Lofty, R.A. Phytochemical evaluation of some selected medicinal plants growing wildly in southeastern Egypt. Middle East J. Appl. Sci. 2015, 5, 1239–1246. [Google Scholar]
- Thevs, N.; Zerbe, S.; Kyosev, Y.; Rozi, A.; Tang, B.; Abdusalih, N.; Novitskiy, Z. Apocynum venetum L. and Apocynum pictum Schrenk (Apocynaceae) as multi-functional and multi-service plant species in Central Asia: A review on biology, ecology, and utilization. J. Appl. Bot. Food Qual. 2012, 85, 159–167. [Google Scholar]
- Sharma, R.; Wungrampha, S.; Singh, V.; Pareek, A.; Sharma, M.K. Halophytes as bioenergy crops. Front. Plant Sci. 2016, 7, 1372. [Google Scholar] [CrossRef]
- Cárdenas-Pérez, S.; Rajabi, D.A.; Leszczynski, K.; Lubinska-Mielinska, S.; Ludwiczak, A.; Piernik, A. Salicornia europaea L. functional traits indicate its optimum growth. Plants 2022, 11, 1051. [Google Scholar] [CrossRef]
- Homayouni, H.; Razi, H.; Izadi, M.; Alemzadeh, A.; Kazemeini, S.A.; Niazi, A.; Vicente, O. Temporal changes in biochemical responses to salt stress in three salicornia species. Plamts 2024, 13, 979. [Google Scholar] [CrossRef]
- Atzori, G.; de Vos, A.C.; van Rijsselberghe, M.; Vignolini, P.; Rozema, J.; Mancuso, S.; van Bodegom, P.M. Effects of increased seawater salinity irrigation on growth and quality of the edible halophyte Mesembryanthemum crystallinum L. under field conditions. Agric. Water Manag. 2017, 187, 37–46. [Google Scholar] [CrossRef]
- Rosales-Nieblas, A.C.; Yamada, M.; Murillo-Amador, B.; Endo, M.; Yamada, S. Evaluation of the cultivation of three halophytic plants under half-strength seawater aquaponics. Agronomy 2025, 15, 277. [Google Scholar] [CrossRef]
- Pucinelli, M.; Marchioni, I.; Botrini, L.; Carmassi, G.; Pardossi, A.; Pistelli, L. Growing Salicornia europaea L. with saline hydroponic or aquaculture wastewater. Horticulturae 2024, 10, 196. [Google Scholar] [CrossRef]
- Spradlin, A.; Saha, S. Saline aquaponics: A review of challenges, opportunities, components, and system design. Aquaculture 2022, 555, 738173. [Google Scholar] [CrossRef]
- Endo, M. Chapter 9.2—Aquaponics in plant factory. In Plant Factory Using Artificial Light; Anpo, M., Fukuda, H., Wada, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 339–352. [Google Scholar] [CrossRef]
- Fimbres-Acedo, Y.E.; Traversari, S.; Cacini, S.; Costamagna, G.; Ginepro, M.; Massa, D. Testing the effect of high pH and low nutrient concentration in four vegetables in hydroponics. Agronomy 2023, 13, 41. [Google Scholar] [CrossRef]
- Chu, Y.T.; Brown, P.B. Evaluation of pacific whiteleg shrimp and three halophytic plants in marine aquaponic systems under three salinities. Sustainability 2021, 13, 269. [Google Scholar] [CrossRef]
- Maciel, E.; Domingues, P.; Domingues, M.R.M.; Calado, R.; Lillebo, A. Halophyte plants from sustainable marine aquaponics are a valuable source of omega-3 polar lipids. Food Chem. 2020, 320, 126560. [Google Scholar] [CrossRef]
- Kaburagi, E.; Yamada, M.; Baba, T.; Fujiyama, H.; Murillo-Amador, B.; Yamada, S. Aquaponics using saline groundwater: Effect of adding microelements to fish wastewater on the growth of Swiss chard (Beta vulgaris L. spp. Cicla). Agric. Water Manag. 2020, 227, 105851. [Google Scholar] [CrossRef]
- Doncato, K.B.; Costa, C.S.B. Micronutrient supplementation needs for halophytes in saline aquaponics with BFT system water. Aquaculture 2021, 531, 735815. [Google Scholar] [CrossRef]
- Doncato, K.B.; Costa, C.S.B. Effects of cutting on vegetative development and biomass quality of perennial halophytes grown in saline aquaponics. Hortic. Bras. 2022, 40, 432–440. [Google Scholar] [CrossRef]
- Doncato, K.B.; Costa, C.S.B. Evaluation of nitrogen and phosphorus nutritional needs of halophytes for saline aquaponics. Hortic. Environ. Biotechnol. 2023, 64, 355–370. [Google Scholar] [CrossRef]
- Tuteja, N. Mechanisms of high salinity tolerance in plants. Methods Enzymol. 2007, 428, 419–438. [Google Scholar] [CrossRef]
- Zhu, J.K.; Hasegawa, P.M.; Bressan, R.A.; Bohnert, H.J. Molecular aspects of osmotic stress in plants. Crit. Rev. Plant Sci. 1997, 16, 253–277. [Google Scholar] [CrossRef]
- Hamdy, A. Saline irrigation: Assessment and management techniques. In Halophytes and Biosaline Agriculture; Chouck-Allah, R., Malcolm, C.V., Hamdy, A., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1996; pp. 147–180. [Google Scholar]
- Carillo, P.; Annunziata, M.G.; Pontecorvo, G.; Fuggi, A.; Woodrow, P. Salinity stress and salt tolerance. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A., Venkateswarlu, B., Eds.; InTech: London, UK, 2011. [Google Scholar] [CrossRef]
- Kaburagi, E.; Morikawa, Y.; Yamada, M.; Fujiyama, H. Sodium enhances nitrate uptake in swiss chard (Beta vulgaris var. cicla). Soil Sci. Plant Nutr. 2014, 60, 651–658. [Google Scholar] [CrossRef]
- Wintermans, J.F.; Mots, A. Spectrophotometric characteristics of chlorophylls a and b and their pheophytins in ethanol. Biochim. Biophys. Acta 1965, 109, 448–453. [Google Scholar] [CrossRef]
- Castellanos-Santiago, E.; Yahia, E.M. Identification and quantification of betalains from the fruits of 10 mexican prickly pear cultivars by high-performance liquid chromatography and electrospray ionization mass spectrometry. J. Agric. Food Chem. 2008, 56, 5758–5864. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, M.E.; Albacete, A.; Martínez-Andújar, C.; Acosta, M.; Romero-Aranda, R.; Dodd, I.C.; Lutts, S.; Pérez-Alfocea, F. Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). J. Exp. Bot. 2008, 59, 3039–3050. [Google Scholar] [CrossRef]
- Tanaka, H.; Yamada, S.; Masunaga, T.; Yamamoto, S.; Tsuji, W.; Murillo-Amador, B. Comparison of nutrient uptake and antioxidative response among four Labiatae herb species under salt stress condition. Soil Sci. Plant Nutr. 2018, 64, 589–597. [Google Scholar] [CrossRef]
- Tanaka, K.; Otsubo, T.; Kondo, N. Participation of hydrogen peroxide in the inactivation of calvin-cycle SH enzymes in SO2-fumigated spinach leaves. Plant Cell Physiol. 1982, 36, 1089–1095. [Google Scholar] [CrossRef]
- Tanaka, K.; Sugahara, K. Role of superoxide dismutase in defense against SO2 toxicity and an increase in superoxide dismutase activity with SO2 fumigation. Plant Cell Physiol. 1980, 21, 601–611. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Asao, T. Hydroponics: A Standard Methodology for Plant Biological Researches; InTech: Rijeka, Croatia, 2012; pp. 1–10. [Google Scholar] [CrossRef]
- Urrestarazú, M. Tratado de Cultivo Sin Suelo, 3rd ed.; Mundi-Prensa: Madrid, Spain, 2004; p. 17. [Google Scholar]
- Frick, J.; Mitchell, C.A. Stabilization of pH in solid matrix hydroponic systems. HortScience 2004, 28, 981–984. [Google Scholar] [CrossRef]
- Bugbee, B. Nutrient management in recirculating hydroponic culture. Acta Hortic. 2004, 648, 99–112. [Google Scholar] [CrossRef]
- Blatt, M.R. A charged existence: A century of transmembrane ion transport in plants. Plant Physiol. 2024, 195, 79–110. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Dunn, B.; Payton, M. Hydroponic pH modifiers affect plant growth and nutrient content in leafy greens. J. Hortic. Res. 2019, 27, 31–36. [Google Scholar] [CrossRef]
- Ali Al Meselmani, M. Nutrient Solution for Hydroponics. In Recent Research and Advances in Soilless Culture; Turan, M., Ed.; InTech: London, UK, 2023. [Google Scholar] [CrossRef]
- Hewitt, E.J. Sand and water culture methods used in the study of plant nutrition. Soil Sci. Soc. Am. J. 1953, 17, 301. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Petropoulos, S.A.; Prvulovic, D.; Tzortzakis, N. Performance of hydroponically cultivated geranium and common verbena under salinity and high electrical conductivity levels. Agronomy 2021, 11, 1237. [Google Scholar] [CrossRef]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef]
- Sanadhya, P.; Agarwal, P.; Agarwal, P.K. Ion homeostasis in a salt-secreting halophytic grass. Aob Plants 2015, 7, plv055. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Han, J.; Mao, P.; Tian, X.; Zheng, M.; Mur, L.A.J. SOS1 is a key systemic regulator of salt secretion and K+/Na+ homeostasis in the recretohalophyte Karelinia caspia. Environ. Exp. Bot. 2020, 177, 104098. [Google Scholar] [CrossRef]
- Carmassi, G.; Incrocci, L.; Malorgio, M.; Tognoni, F.; Pardossi, A. A simple model for salt accumulation in closed-loop hydroponics. Acta Hortic. 2003, 614, 149–154. [Google Scholar] [CrossRef]
- Szabados, L.; Kovács, H.; Zilberstein, A.; Bouchereau, A. Plants in extreme environments: Importance of protective compounds in stress tolerance. In Plant Responses to Drought and Salinity Stress Developments in a Post-Genomic Era, Advances in Botanical Research; Turkan, I., Ed.; Academic Press: Orlando, FL, USA, 2011; Volume 57, pp. 105–150. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotox. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Yamada, M.; Kasami, K.; Urushigaki, R.; Murillo-Amador, B.; Yamada, S. Mechanisms of halophilic and salt tolerance in Suaeda edulis Flores Olv. & Noguez. Soil Sci. Plant Nutr. 2025, 1–10. [Google Scholar] [CrossRef]
- Belkheiri, O.; Mulas, M. The effects of salt stress on growth, water relations and ion accumulation in two halophyte Atriplex species. Environ. Exp. Bot. 2013, 86, 17–28. [Google Scholar] [CrossRef]
- Ungar, I.A. Ecophysiology of Vascular Halophytes; CRC Press: Boca Raton, FL, USA, 1991; p. 209. [Google Scholar] [CrossRef]
- Pitman, M.G. Transport across the root and shoot/root interactions. In Salinity Tolerance in Plants: Strategies for Crop Improvement; Staples, R.C., Toenniessen, G.H., Eds.; John Wiley & Sons: New York, NY, USA, 1984; p. 93. [Google Scholar]
- LiangPeng, Y.; Jian, M.A.; Yan, L.I. Impact of salt stress on the features and activities of root system for three desert halophyte species in their seedling stage. Sci. China Ser. D 2007, 50, 97–106. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Z.; Ge, S.; Peng, B.; Zhang, K.; Hu, M.; Mai, W.; Tian, C. Root morphology and rhizosphere characteristics are related to sat tolerance of Suaeda salsa and Beta vulgaris L. Front. Plant Sci. 2021, 12, 677767. [Google Scholar] [CrossRef]
- Lüttge, U.; Fischer, E.; Steudle, E. Membrane potentials and salt distribution in epidermal bladders and photosynthetic tissue of Mesembryanthemum crystallinum L. Plant Cell Environ. 1978, 1, 121–129. [Google Scholar] [CrossRef]
- Agarie, S.; Shimoda, T.; Shimizu, Y.; Baumann, K.; Sunagawa, H.; Kondo, A.; Ueno, O.; Nakahara, T.; Nose, A.; Cushman, J.C. Salt tolerance, salt accumulation, and ionic homeostasis in an epidermal bladder-cell-less mutant of the common ice plant Mesembryanthemum crystallinum. J. Exp. Bot. 2007, 58, 1957–1967. [Google Scholar] [CrossRef]
- Abd El-Gawad, A.M.; Shehata, H.S. Ecology and development of Mesembryanthemum crystallinum L. in the deltaic mediterranean coast of Egypt. Egypt. J. Basic Appl. Sci. 2014, 1, 29–37. [Google Scholar] [CrossRef]
- Adams, P.; Nelson, D.E.; Yamada, S.; Chmara, W.; Jensen, R.G.; Bohnert, H.J.; Griffiths, H. Growth and development of Mesembryanthemum crystallinum (Aizoaceae). New Phytol. 1998, 138, 171–190. [Google Scholar] [CrossRef]
- Lv, S.; Jiang, P.; Chen, X.; Fan, P.; Wang, X.; Li, Y. Multiple compartmentalization of sodium conferred salt tolerance in Salicornia europaea. Plant Physiol. Biochem. 2012, 51, 47–52. [Google Scholar] [CrossRef]
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Aziz, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of salinity stress on chloroplast structure and function. Cells 2021, 10, 2023. [Google Scholar] [CrossRef]
- Rozema, J. An ecophysiological study on the response to salt of four halophytic and glycophytic Juncus species. Flora 1976, 165, 197–209. [Google Scholar] [CrossRef]
- Su, H.; Golldack, D.; Katsuhara, M.; Zhao, C.; Bohnert, H.J. Expression and stress-dependent induction or potassium channel transcripts in the common ice plant. Plant Physiol. 2001, 125, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Golldack, D.; Zhao, C.; Bohnert, H.J. The expression of HAK-type K+ transporters is regulated in response to salinity stress in common ice plant. Plant Physiol. 2002, 129, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- Rus, A.; Yokoi, S.; Sharkuu, A.; Reddy, M.; Lee, B.H.; Matsumoto, T.K.; Koiwa, H.; Zhi, J.K.; Bressan, R.A.; Hasegawa, P.M. AtHKT1 is a salt tolerance determinant that controls Na+ entry into plant roots. Proc. Natl. Acad. Sci. USA 2001, 98, 14150–14155. [Google Scholar] [CrossRef] [PubMed]
- Barkla, B.J.; Zingarelli, L.; Blumwald, E.; Smith, J.A.C. Tonoplast Na+/H+ antiport activity and its energization by the vacuolar H+-ATPase in the halophytic plant Mesembryanthemum crystallinum L. Plant Physiol. 1995, 109, 549–556. [Google Scholar] [CrossRef]
- Ratajczak, R.; Richter, J.; Lüttge, U. Adaptation of the tonoplast V-type H+-ATPase of Mesembryanthemum crystallinum to salt stress, C3-CAM transition and plant age. Plant Cell Environ. 1994, 17, 1101–1112. [Google Scholar] [CrossRef]
- Vernon, D.M.; Bohnert, H.J. A novel methyl transferase induced by osmotic stress in the facultative halophyte Mesembryanthemum crystallinum. EMBO J. 1992, 11, 2077–2085. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Schroeder, J.I. Structure and transport mechanism of a high-affinity potassium uptake transporter from higher plants. Nature 1994, 370, 655–658. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Y.; Duan, H.R.; Yin, X.X.; Cui, Y.N.; Chai, W.W.; Song, X.; Flowers, T.J.; Wang, S. SsHKT1;1 is coordinated with SsSOS1 and SsNHX1 to regulate Na+ homeostasis in Suaeda salsa under saline conditions. Plant Soil 2020, 449, 117–131. [Google Scholar] [CrossRef]
- Lebaudy, A.; Véry, A.A.; Sentenac, H. K+ channel activity in plants: Genes, regulations and functions. FEBS Lett. 2007, 581, 2357–2366. [Google Scholar] [CrossRef]
- Qiu, N.; Chen, M.; Guo, J.; Bao, H.; Ma, X.; Wang, B. Coordinate up-regulation of V-H+-ATPase and vacuolar Na+/H+ antiporter as a response to NaCl treatment in a C3 halophyte Suaeda salsa. Plant Sci. 2007, 172, 1218–1225. [Google Scholar] [CrossRef]
- Want, B.; Lüttge, V.; Ratajczak, R. Effects of salt treatment and osmotic stress on V-ATPase and V-PPase in leaves of the halophyte Suaeda salsa. J. Exp. Bot. 2001, 52, 2355–2365. [Google Scholar] [CrossRef]
- Nedelyaeva, O.I.; Popova, L.G.; Volkov, V.S.; Balnokin, Y.V. Molecular cloning and characterization of SaCLCd, SaCLCf, and SaCLCg, novel proteins of the chloride channel family (CLC) from the halophyte Suaeda altissima (L.) Pall. Plants 2022, 11, 409. [Google Scholar] [CrossRef] [PubMed]
- Nedelyaeva, O.I.; Shuvalov, A.V.; Mayorova, O.V.; Yurchenko, A.A.; Popova, L.G.; Balnokin, Y.V.; Karpichev, I.V. Cloning and functional analysis of SaCLCc1, a gene belonging to the chloride channel family (CLC), from the halophyte Suaeda altissima (L.) Pall. Dokl. Biochem. Biophys. 2018, 481, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, J.; Deng, C.; Liu, Z.; Yin, K.; Zhang, Y.; Zhao, Z.; Zhao, R.; Zhao, N.; Zhou, X.; et al. Effect of NaCl on ammonium and nitrate uptake and transport in salt-tolerant and salt-sensitive poplars. Three Physiol. 2024, 44, tpae020. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S. Salinity effects on nitrogen metabolism in plants—Focusing on the activities of nitrogen metabolizing enzymes: A review. J. Plant Nutr. 2018, 41, 1065–1081. [Google Scholar] [CrossRef]
- Libert, B.; Franceschi, V.R. Oxalate in crop plants. J. Agric. Food Chem. 1987, 35, 926–938. [Google Scholar] [CrossRef]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef]
- Barker, D.H.; Marszalek, J.; Zimpfer, J.F.; Adams, W.W., III. Changes in photosynthetic pigment composition and absorbed energy allocation during salt stress and CAM induction in Mesembryanthemum crystallinum. Funct. Plant Biol. 2004, 31, 781–787. [Google Scholar] [CrossRef]
- Koyro, H.W.; Huchzermeyer, B. Breeding for abiotic stress tolerance in maize. In Abiotic Stress: Plant Resistance Through Breeding and Molecular Approaches; Ashraf, M., Harris, P.J.C., Eds.; The Haworth Press: New York, NY, USA, 2005; pp. 545–576. [Google Scholar]
- Sudhir, P.; Murthy, S.D.S. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Koyro, H.W.; Zörb, C.; Debez, A.; Huchzermeyer, B. The effect of hyper-osmotic salinity on protein pattern and enzyme activities of halophytes. Funct. Plant Biol. 2013, 40, 787–804. [Google Scholar] [CrossRef]
- Akcin, A.; Yalçin, E. Effect of salinity stress on chlorophyll, carotenoid content, and proline in Salicornia prostrata Pall. And Suaeda prostrata Pall. subsp. prostrata (Amaranthaceae). Braz. J. Bot. 2016, 39, 101–106. [Google Scholar] [CrossRef]
- Cerillo-Rojas, G.V.; Tiscareño-Andrade, M.; Ochoa-Alfaro, A.E.; Pérez-Molphe, B.E.; Soria-Guerra, R.E.; Morales-Domínguez, J.F. In vitro propagation, isolation and expression studies of Suaeda edulis genes involved in the osmoprotectans biosynthesis. Phyton-Int. J. Exp. Bot. 2020, 89, 715–726. [Google Scholar] [CrossRef]
- Parida, A.K.; Kumari, A.; Panda, A.; Rangani, J.; Agarwal, P.K. Photosynthetic pigments, betalains, proteins, sugars, and minerals during Salicornia brachiata senescence. Biol. Plant. 2018, 62, 343–352. [Google Scholar] [CrossRef]
- Ungar, I.A.; Benner, D.K.; McGraw, D.C. The distribution and growth of Salicornia europaea on an inland salt pan. Ecology 1979, 60, 329–336. [Google Scholar] [CrossRef]
- Li, J.T.; Qiu, Z.B.; Zhang, X.W.; Wang, L.S. Exogenous hydrogen peroxide can enhance tolerance of wheat seedlings to salt stress. Acta physiol. Plant 2011, 33, 835–842. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015; p. 270. [Google Scholar]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Jha, B. Salt tolerance mechanisms in mangroves: A review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B.; Mohanty, P. Defense potentials to NaCl in a mangrove, Bruguiera parviflora: Differential changes of isoforms of some antioxidative enzymes. J. Plant Physiol. 2004, 161, 531–542. [Google Scholar] [CrossRef]
- Voothuluru, P.; Mäkelä, P.; Zhu, J.; Yamaguchi, M.; Cho, I.J.; Oliver, M.J.; Simmonds, J.; Sharp, R.E. Apoplastic hydrogen peroxide in the growth zone of the maize primary root. Increased levels differentially modulate root elongation under well-watered and water-stressed conditions. Front. Plant Sci. 2020, 11, 392. [Google Scholar] [CrossRef]
- Kawano, T. Roles of the reactive oxygen species-generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep. 2003, 21, 829–837. [Google Scholar] [CrossRef]
- Gupta, R.; Verma, N.; Tewari, R.J. Micronutrient deficiency-induced oxidative stress in plants. Plant Cell Rep. 2024, 43, 213. [Google Scholar] [CrossRef]
- Hsieh, L.C.; Lee, C.C.; Zhang, K.F.; Chang, H.H.; Li, C.H.; Huang, H.J. Transcriptomic and enzymatic analysis of peroxidase families at the early growth stage of halophyte ice plant (Mesembryanthemum crystallinum L.) under salt stress. Bot. Stud. 2025, 66, 5. [Google Scholar] [CrossRef]
- Shevyakova, N.I.; Rakitin, V.Y.; Stetsenko, L.A.; Aronova, E.E.; Kuznetsov, V.V. Oxidative stress and fluctuations of free and conjugated polyamines in the halophyte Mesembryanthemum crystallinum L. under NaCl salinity. Plant Growth Regul. 2006, 50, 69–78. [Google Scholar] [CrossRef]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic stress: Interplay between ROS, hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Shakirova, F.M.; Allagulova, C.R.; Maslennikova, D.R.; Klyuchnikova, E.O.; Avalbaev, A.M.; Bezrukova, M.V. Salicylic acid-induced protection against cadmium toxicity in wheat plants. Environ. Exp. Bot. 2016, 122, 19–28. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, A.; Yadav, S.; Singh, I.K. Reactive oxygen species-mediated signaling during abiotic stress. Plant Gene 2019, 18, 100–173. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant response to salt stress and role of exogenous protectants to mitigate salt-induced damages. In Ecophysiology and Responses of Plants under Salt Stress; Ahmed, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 25–87. [Google Scholar]
- Zandalinas, S.I.; Fichman, Y.; Devireddy, A.R.; Sengupta, S.; Azad, R.K.; Mittler, R. System signaling during abiotic stress combination in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 13810–13820. [Google Scholar] [CrossRef]
- Sui, N. Photoinhibition of Suaeda salsa to chilling stress is related to energy dissipation and water-water cycle. Photosynthetica 2015, 53, 207–212. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van Montagu, M.; Inzé, D.; Van Camp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef]
- Aghaleh, M.; Niknam, V.; Ebrahimzadeh, H.; Razavi, K. Salt stress effects on growth, pigments, proteins and lipid peroxidation in Salicornia persica and S. europaea. Biol. Plant 2009, 53, 243–248. [Google Scholar] [CrossRef]
- Aghaleh, M.; Niknam, V.; Ebrahimzadeh, H.; Razavi, K. Effect of salt stress on physiological and antioxidative responses in two species of Salicornia (S. persica and S. europaea). Acta Physiol. Plant 2011, 33, 1261–1270. [Google Scholar] [CrossRef]
- Moghaddam, A.; Larijani, H.R.; Oveysi, M.; Moghaddam, H.R.T.; Nasri, M. Alleviating the adverse effects of salinity stress on Salicornia persica using sodium nitroprusside and potassium nitrate. BMC Plant Biol. 2023, 23, 166. [Google Scholar] [CrossRef] [PubMed]
- Slesak, I.; Miszalski, Z.; Karpinska, B.; Niewiadomska, E.; Ratajczak, R.; Karpinski, S. Redox control of oxidative stress responses in the C3-CAM intermediate plant Mesembryanthemum crystallinum. Plant Physiol. Biochem. 2002, 40, 669–677. [Google Scholar] [CrossRef]
- Agarie, S.; Kawaguchi, A.; Kodera, A.; Sunagawa, H.; Kojima, H.; Nose, A.; Nakahara, T. Potential of the common ice plant, Mesembryanthemum crystallinum as a new high-functional food as evaluated by polyol accumulation. Plant Prod. Sci. 2009, 12, 37–46. [Google Scholar] [CrossRef]
- Ibdah, M.; Krins, A.; Seidlitz, H.K.; Heller, W.; Strack, D.; Vogt, T. Spectral dependence of flavonol and betacyanin accumulation in Mesembryanthemum crystallinum under enhanced ultraviolet radiation. Plant Cell Environ. 2002, 25, 1145–1154. [Google Scholar] [CrossRef]
- Guo, J.; Wang, H.; Wang, B. NaCl enhanced the vegetative growth of halophyte Suaeda salsa by improving the ability of antioxidant and osmotic adjustment. IOP Conf. Ser. Earth Environ. Sci. 2019, 237, 052024. [Google Scholar] [CrossRef]
- Zang, W.; Miao, R.; Zhang, Y.; Yuan, Y.; Pan, Q.; Zhou, Z. Metabolic and molecular basis for the salt and alkali responses of Suaeda corniculate. Environ. Exp. Bot. 2021, 192, 104643. [Google Scholar] [CrossRef]
- Behr, J.H.; Bouchereau, A.; Berardocco, S.; Seal, C.E.; Flowers, T.J.; Zörb, C. Metabolic and physiological adjustment of Suaeda maritima to combined salinity and hypoxia. Ann. Bot. 2017, 119, 965–976. [Google Scholar] [CrossRef]
- Hussain, S.; Khalid, M.F.; Hussain, M.; Ali, M.A.; Nawaz, A.; Zakir, I.; Fatima, Z.; Ahmad, S. Role of micronutrients in salt stress tolerance to plants. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; p. 363. [Google Scholar] [CrossRef]
- Raddatz, N.; Morales de los Ríos, L.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated transport of nitrate, potassium, and sodium. Front. Plant Sci. 2020, 11, 247. [Google Scholar] [CrossRef]
- Ushakova, S.A.; Kovaleva, N.P.; Gribovskaya, I.V.; Dolgushev, V.A.; Tikhomirova, N.A. Effect of NaCl concentration on productivity and mineral composition of Salicornia europaea as a potential crop for utilization NaCl in LSS. Adv. Space Res. 2005, 36, 1349–1353. [Google Scholar] [CrossRef]
- Flowers, T.J.; Hajibagher, M.A.; Yeo, A.R. Ion accumulation in the cell walls of rice plants growing under saline conditions: Evidence for the Oertli hypothesis. Plant Cell Environ. 1991, 14, 319–325. [Google Scholar] [CrossRef]
- Moghaieb, R.E.A.; Saneoka, H.; Fujita, K. Effect of salinity on osmotic adjustment, glycine betaine accumulation and the betaine aldehyde dehydrogenase gene expression in two halophytic plants, Salicornia europaea and Suaeda maritima. Plant Sci. 2004, 166, 1345–1349. [Google Scholar] [CrossRef]
- Fussy, A.; Papenbrock, J. Molecular analysis of the reactions in Salicornia europaea to varying NaCl concentrations at various stages of development to better exploit its potential as a new crop plant. Front. Plant Sci. 2024, 15, 1454541. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ice Plant | Romeritos | Sea Asparagus | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | pH | pH+S | NS * | C | pH | pH+S | NS * | C | pH | pH+S | NS * | ||
| pH | 7.9 | 5.5 | 5.5 | 5.5 | 7.8 | 5.5 | 5.5 | 5.5 | 8.0 | 5.5 | 5.5 | 5.5 | |
| T-N | mM | 5.9 | 5.9 | 5.9 | 4.0 | 6.6 | 6.6 | 6.6 | 4.0 | 4.6 | 4.6 | 4.6 | 4.0 |
| -N | 4.9 | 4.9 | 4.9 | 4.0 | 4.2 | 4.2 | 4.2 | 4.0 | 3.7 | 3.7 | 3.7 | 4.0 | |
| -N | - | - | - | - | 0.1 | 0.1 | 0.1 | - | 0.1 | 0.1 | 0.1 | - | |
| -N | 0.07 | 0.07 | 0.07 | - | 0.05 | 0.05 | 0.05 | - | 0.2 | 0.2 | 0.2 | - | |
| P | 0.2 | 0.2 | 0.4 | 0.4 | 0.1 | 0.1 | 0.4 | 0.4 | 0.1 | 0.1 | 0.4 | 0.4 | |
| K | 9.05 | 9.05 | 9.05 | 2.0 | 9.1 | 9.1 | 9.1 | 2.0 | 8.9 | 8.9 | 8.9 | 2.0 | |
| Ca | 1.2 | 1.2 | 1.2 | 1.0 | 1.2 | 1.2 | 1.2 | 1.0 | 1.06 | 1.06 | 1.06 | 1.0 | |
| Mg | 22.0 | 22.0 | 22.0 | 2.0 | 25.4 | 25.4 | 25.4 | 2.0 | 23.2 | 23.2 | 23.2 | 2.0 | |
| Na | 262.4 | 262.4 | 262.4 | 5.0 | 263.6 | 263.6 | 263.6 | 5.0 | 250.6 | 250.6 | 250.6 | 5.0 | |
| Cl | 311.6 | 311.6 | 311.6 | 5.0 | 307.5 | 307.5 | 307.5 | 5.0 | 297.2 | 297.2 | 297.2 | 5.0 | |
| B | µM | 216.4 | 216.4 | 216.4 | 18.0 | 213.8 | 213.8 | 213.8 | 18.0 | 209.8 | 209.8 | 209.8 | 18.0 |
| Fe | - | - | 35.0 | 35.0 | - | - | 35.0 | 35.0 | - | - | 35.0 | 35.0 | |
| Mn | 0.6 | 0.6 | 9.0 | 9.0 | 0.5 | 0.5 | 9.0 | 9.0 | 0.6 | 0.6 | 9.0 | 9.0 | |
| Zn | - | - | 1.0 | 1.0 | - | - | 1.0 | 1.0 | - | - | 1.0 | 1.0 | |
| Cu | - | - | 0.1 | 0.1 | - | - | 0.1 | 0.1 | - | - | 0.1 | 0.1 | |
| Leaf Area (cm2 plant−1) | Specific Leaf Area (cm2 g−1 FW) | Leaf or Shoot Water Content (g g−1 DW) | Leaf or Shoot Succulence (g water cm2) | Shoot/Root Ratio | ||
|---|---|---|---|---|---|---|
| Ice | C | 785.01 ± 47.22 | 6.01 ± 0.30 | 40.97 ± 3.71 | 0.16 ± 0.009 | 7.30 ± 0.92 |
| plant | pH | 783.90 ± 35.90 | 5.49 ± 0.12 | 50.46 ± 9.77 | 0.17 ± 0.003 | 5.38 ± 0.89 |
| pH+S | 737.76 ± 59.10 | 5.49 ± 0.33 | 41.17 ± 0.87 | 0.18 ± 0.012 | 5.12 ± 0.40 | |
| NS | 1058.17 ± 154.74 | 6.71 ± 0.44 | 44.98 ± 3.24 | 0.14 ± 0.009 | 7.92 ± 1.90 | |
| ns | ns | ns | ns | ns | ||
| Romeritos | C | 1020.72 ± 101.68 a | 15.23 ± 1.06 b | 10.28 ± 0.20 a | 0.06 ± 0.005 a | 4.31 ± 0.25 bc |
| pH | 742.77 ± 73.14 ab | 14.42 ± 0.34 b | 6.07 ± 0.50 b | 0.05 ± 0.002 ab | 6.60 ± 0.39 a | |
| pH+S | 623.68 ± 23.90 b | 15.07 ± 0.75 b | 5.68 ± 0.12 b | 0.05 ± 0.003 ab | 5.68 ± 0.55 ab | |
| NS | 1025.43 ± 74.19 a | 19.25 ± 0.85 a | 9.54 ± 0.42 a | 0.04 ± 0.002 b | 3.77 ± 0.12 c | |
| ** | ** | *** | * | *** | ||
| Sea | C | 290.47 ± 1.60 | 7.39 ± 0.36 | 5.50 ± 0.45 | 0.11 ± 0.006 | 6.33 ± 0.67 |
| asparagus | pH | 297.18 ± 40.82 | 6.79 ± 0.66 | 5.49 ± 0.05 | 0.12 ± 0.015 | 6.76 ± 0.27 |
| pH+S | 270.29 ± 23.77 | 6.69 ± 0.24 | 5.68 ± 0.17 | 0.12 ± 0.004 | 7.28 ± 0.67 | |
| NS | 278.72 ± 51.85 | 8.48 ± 0.59 | 5.25 ± 0.29 | 0.10 ± 0.008 | 5.80 ± 0.05 | |
| ns | ns | ns | ns | ns |
| mM | mM | mM | mM | ||
|---|---|---|---|---|---|
| Ice | C | 217.48 ± 23.38 a | N.D. | 18.58 ± 1.33 b | 8.54 ± 0.29 a |
| plant | pH | 215.44 ± 7.27 a | N.D. | 25.01 ± 1.45 b | 9.23 ± 0.61 a |
| pH+S | 229.72 ± 11.67 a | N.D. | 23.46 ± 1.44 b | 9.66 ± 0.41 a | |
| NS | 81.94 ± 14.54 b | 0.72 ± 0.05 | 38.36 ± 2.47 a | 6.00 ± 0.28 b | |
| *** | *** | *** | |||
| Romeritos | C | 377.50 ± 51.15 b | N.D. | 24.35 ± 2.57 c | 23.86 ± 3.35 |
| pH | 907.11 ± 30.68 a | 2.36 ± 1.43 | 43.91 ± 2.16 b | 27.84 ± 2.48 | |
| pH+S | 950.49 ± 50.14 a | 4.06 ± 1.51 | 66.57 ± 4.40 a | 34.11 ± 5.59 | |
| NS | 154.59 ± 8.12 c | 0.24 ± 0.15 | 80.92 ± 5.35 a | 20.96 ± 1.80 | |
| *** | ns | *** | ns | ||
| Sea | C | 827.45 ± 2.85 a | N.D. | 131.62 ± 7.34 b | 12.60 ± 2.00 |
| asparagus | pH | 796.83 ± 32.27 a | 2.74 ± 1.63 b | 96.80 ± 10.18 b | 13.33 ± 0.89 |
| pH+S | 770.98 ± 22.34 a | 2.69 ± 0.97 b | 101.72 ± 1.80 b | 15.04 ± 1.39 | |
| NS | 423.96 ± 50.48 b | 29.24 ± 2.56 a | 187.81 ± 19.31 a | 17.05 ± 0.69 | |
| *** | *** | *** | ns |
| mM | mM | mM | mM | mM | mM | ||
|---|---|---|---|---|---|---|---|
| Ice | C | 155.42 ± 17.04 a | 0.79 ± 0.06 b | 48.44 ± 4.11 a | 4.59 ± 0.55 | 5.42 ± 0.89 | 34.98 ± 5.30 ab |
| plant | pH | 217.47 ± 6.73 a | 1.08 ± 0.2 a | 51.51 ± 2.97 a | 4.59 ± 0.18 | 7.30 ± 1.09 | 23.10 ± 5.35 b |
| pH+S | 200.01 ± 27.68 a | 0.98 ± 0.6 ab | 48.43 ± 5.15 a | 3.57 ± 0.37 | 5.27 ± 0.61 | 30.14 ± 3.48 ab | |
| NS | 58.98 ± 3.40 b | N.D. | 28.68 ± 4.20 b | 3.51 ± 0.25 | 4.13 ± 0.75 | 49.11 ± 7.08 a | |
| *** | * | ** | ns | ns | * | ||
| Romeritos | C | 442.78 ± 47.12 b | 2.20 ± 0.11 | 47.30 ± 4.54 a | 3.01 ± 0.21c | 1.07 ± 0.03 | 50.59 ± 5.90 a |
| pH | 871.04 ± 139.58 a | 1.43 ± 0.78 | 45.59 ± 8.21 a | 5.51 ± 0.57 c | 1.20 ± 0.06 | 6.78 ± 0.26 c | |
| pH+S | 1209.98 ± 81.90 a | 0.62 ± 0.07 | 55.19 ± 4.82 a | 18.67 ± 1.98 a | 1.05 ± 0.03 | 6.90 ± 0.31 c | |
| NS | 346.29 ± 8.19 b | N.D. | 19.48 ± 1.04 b | 11.12 ± 0.74 b | 0.90 ± 0.13 | 29.49 ± 4.63 b | |
| *** | ns | ** | *** | ns | *** | ||
| Sea | C | 1491.79 ± 62.03 a | 1.20 ± 0.56 | 29.01 ± 5.31 | 8.83 ± 1.90 b | 0.81 ± 0.05 b | 19.29 ± 2.46 |
| asparagus | pH | 1310.94 ± 191.92 ab | 1.49 ± 0.53 | 39.69 ± 5.00 | 13.62 ± 1.39 ab | 1.56 ± 0.19 a | 23.12 ± 4.02 |
| pH+S | 1343.63 ± 207.20 ab | 0.87 ± 0.33 | 44.10 ± 7.29 | 13.43 ± 2.19 ab | 1.28 ± 0.18 ab | 30.68 ± 4.66 | |
| NS | 729.63 ± 70.67 b | 1.35 ± 0.63 | 55.49 ± 9.19 | 21.88 ± 3.80 a | 1.07 ± 0.12 ab | 29.84 ± 4.06 | |
| * | ns | ns | * | * | ns |
| CAT *1 (mmol H2O mg−1 Protein min−1) | APX *2 (mmol ASC mg−1 Protein min−1) | SOD *3 (unit mg−1 Protein min−1) | GR *4 (mmol NADPH mg−1 Protein min−1) | ||
|---|---|---|---|---|---|
| Ice | C | 203.15 ± 17.62 | 0.34 ± 0.07 ab | 62.73 ± 2.04 a | 0.39 ± 0.03 a |
| plant | pH | 165.22 ± 7.47 | 0.37 ± 0.06 ab | 42.54 ± 9.96 ab | 0.37 ± 0.06 a |
| pH+S | 241.44 ± 42.68 | 0.50 ± 0.13 a | 35.46 ± 4.87 b | 0.17 ± 0.07 ab | |
| NS | 207.41 ± 49.39 | 0.12 ± 0.01 b | 40.63 ± 2.57 ab | 0.13 ± 0.01 b | |
| ns | * | * | *** | ||
| Romeritos | C | 85.90 ± 7.35 a | 0.15 ± 0.01 ab | 18.70 ± 3.32 ab | 0.12 ± 0.007 a |
| pH | 57.07 ± 7.45 ab | 0.26 ± 0.04 a | 24.07 ± 3.53 a | 0.04 ± 0.002 b | |
| pH+S | 43.27 ± 11.57 b | 0.27 ± 0.02 a | 23.55 ± 1.91 a | 0.08 ± 0.029 ab | |
| NS | 31.41 ± 8.02 b | 0.07 ± 0.03 b | 9.06 ± 1.50 b | 0.03 ± 0.015 b | |
| ** | ** | ** | * | ||
| Sea | C | 40.34 ± 5.86 | 0.08 ± 0.004 | 7.25 ± 0.65 a | 0.05 ± 0.011 |
| asparagus | pH | 54.77 ± 7.60 | 0.04 ± 0.011 | 3.63 ± 0.43 bc | 0.07 ± 0.006 |
| pH+S | 30.80 ± 2.72 | 0.08 ± 0.019 | 5.88 ± 1.06 ab | 0.03 ± 0.006 | |
| NS | 43.24 ± 8.69 | 0.05 ± 0.014 | 1.06 ± 0.10 c | 0.05 ± 0.008 | |
| ns | ns | *** | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosales-Nieblas, A.C.; Yamada, M.; Murillo-Amador, B.; Yamada, S. Exploring Salinity Tolerance in Three Halophytic Plants: Physiological and Biochemical Responses to Agronomic Management in a Half-Strength Seawater Aquaponics System. Horticulturae 2025, 11, 623. https://doi.org/10.3390/horticulturae11060623
Rosales-Nieblas AC, Yamada M, Murillo-Amador B, Yamada S. Exploring Salinity Tolerance in Three Halophytic Plants: Physiological and Biochemical Responses to Agronomic Management in a Half-Strength Seawater Aquaponics System. Horticulturae. 2025; 11(6):623. https://doi.org/10.3390/horticulturae11060623
Chicago/Turabian StyleRosales-Nieblas, Ayenia Carolina, Mina Yamada, Bernardo Murillo-Amador, and Satoshi Yamada. 2025. "Exploring Salinity Tolerance in Three Halophytic Plants: Physiological and Biochemical Responses to Agronomic Management in a Half-Strength Seawater Aquaponics System" Horticulturae 11, no. 6: 623. https://doi.org/10.3390/horticulturae11060623
APA StyleRosales-Nieblas, A. C., Yamada, M., Murillo-Amador, B., & Yamada, S. (2025). Exploring Salinity Tolerance in Three Halophytic Plants: Physiological and Biochemical Responses to Agronomic Management in a Half-Strength Seawater Aquaponics System. Horticulturae, 11(6), 623. https://doi.org/10.3390/horticulturae11060623