Deoxynivalenol (DON) Accumulation and Nutrient Recovery in Black Soldier Fly Larvae (Hermetia illucens) Fed Wheat Infected with Fusarium spp.

 , , and
, , and
Abstract
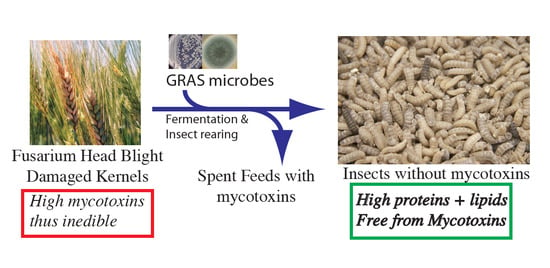
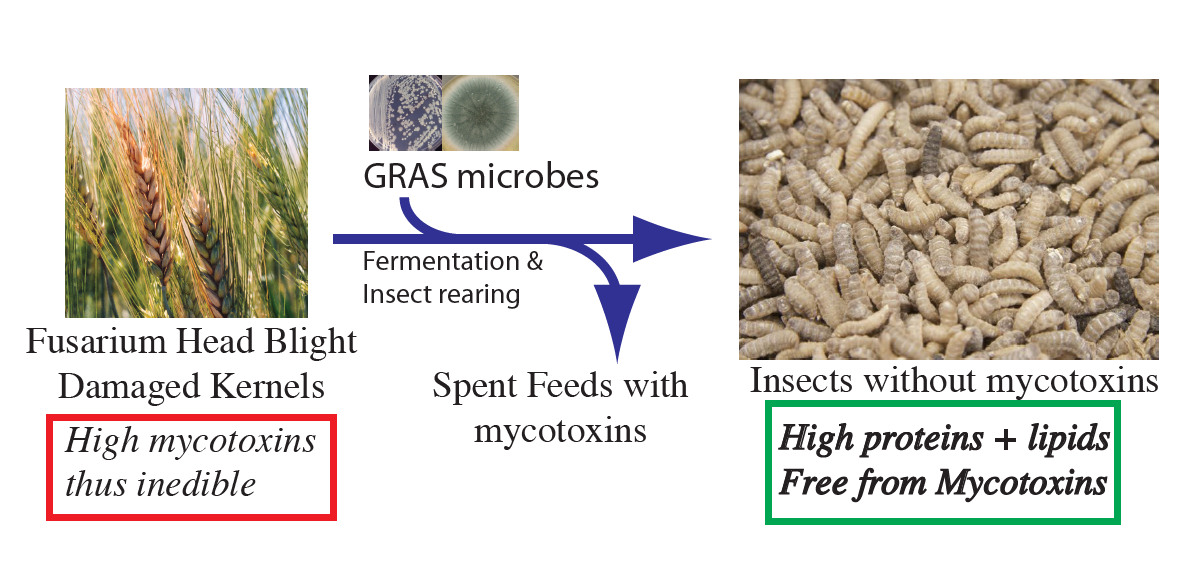
1. Introduction
2. Material and Methods
2.1. Materials
2.2. Solid-State Fermentation
2.3. BSFL Digestion
2.4. BSFL Separation after Digestion
2.5. Larval Weight Gain Determination and Survival Rate of Larvae
2.6. Proximate Analysis of BSFL and Spent Feed
2.7. Mycotoxin Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. Growth Rate of BSFL
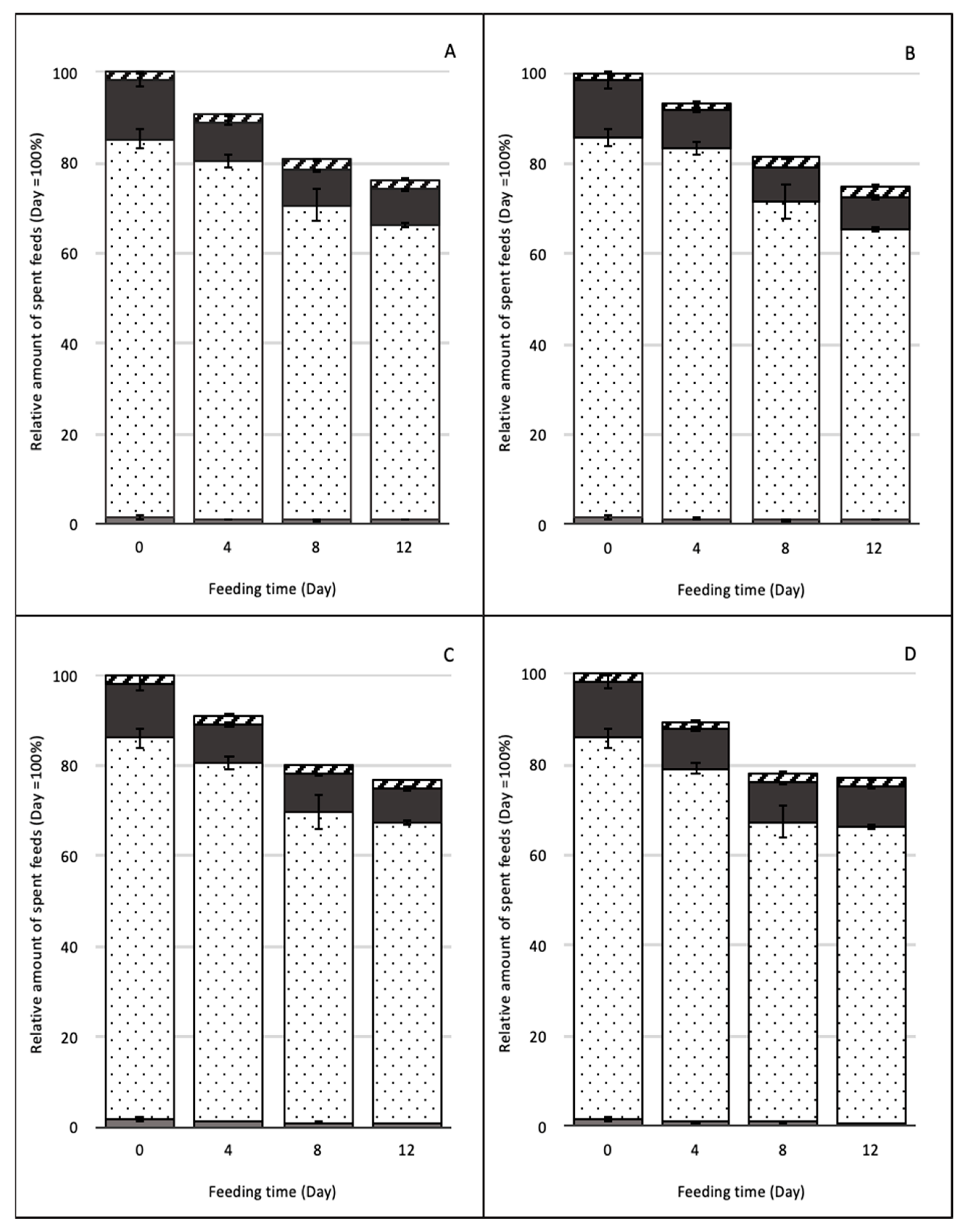
3.2. The Proximate Composition of BSFL and Spent Feed
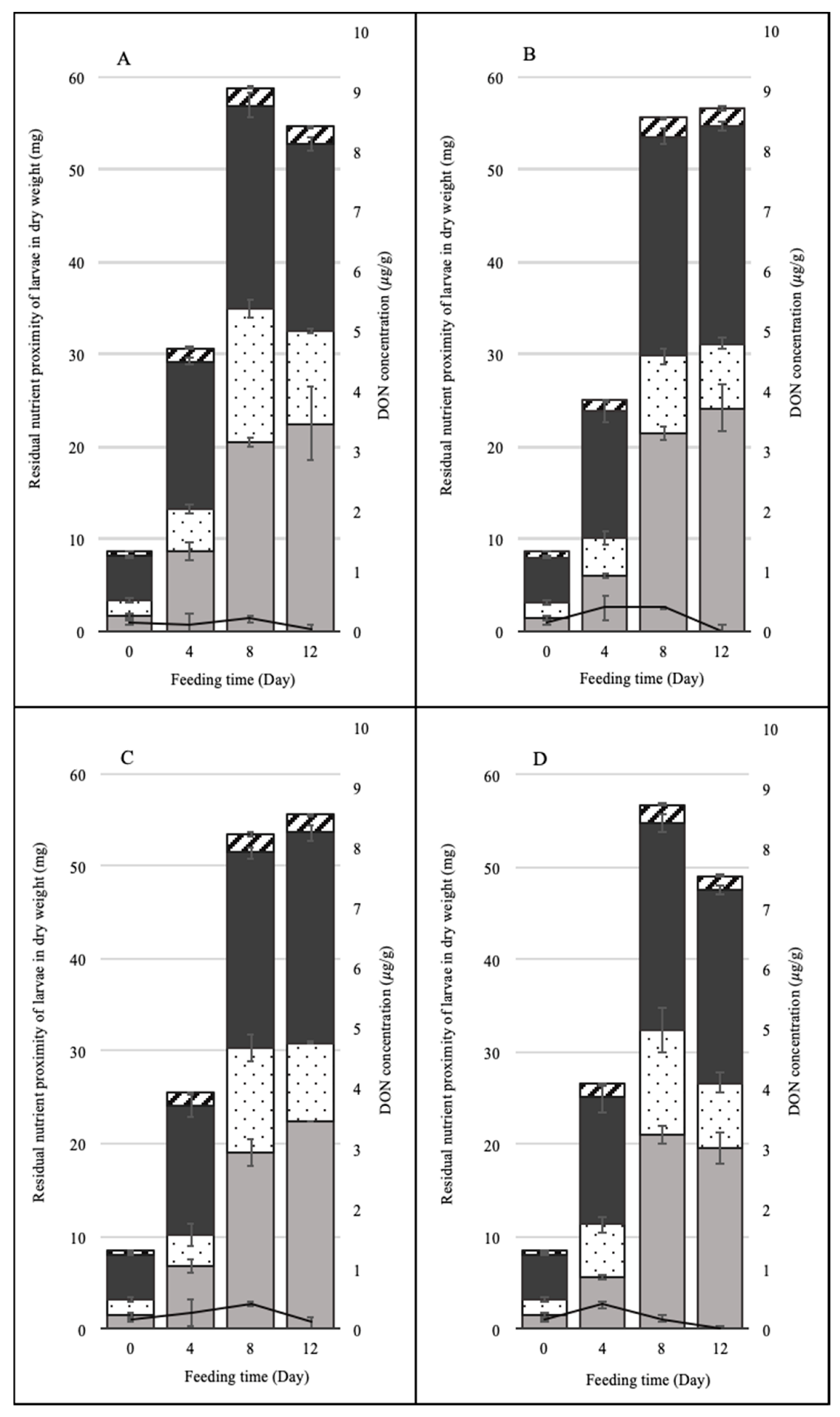
3.3. Amounts of DON in Spent Feed and BSFL Biomass
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yoshida, M.; Nakajima, T. Deoxynivalenol and nivalenol accumulation in wheat infected with Fusarium graminearum during grain development. Phytopathology 2010, 100, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Hsiao, F.S.H.; Proskura, W.S.; Dybus, A.; Siao, Y.H.; Cheng, Y.H. An impact of deoxynivalenol produced by Fusarium graminearum on broiler chickens. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Bosch, G.; Fels-Klerx, H.J.V.; Rijk, T.C.; Oonincx, D. Aflatoxin B1 tolerance and accumulation in black soldier fly larvae (Hermetia illucens) and yellow mealworms (Tenebrio molitor). Toxins 2017, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Kuttiyatveetil, J.R.A.; Mitra, P.; Goldin, D.; Nickerson, M.T.; Tanaka, T. Recovery of residual nutrients from agri-food byproducts using a combination of solid-state fermentation and insect rearing. Int. J. Food Sci. Technol. 2018, 54, 1130–1140. [Google Scholar] [CrossRef]
- Schisler, D.A.; Khan, N.I.; Boehm, M.J. Biological control of Fusarium head blight of wheat and deoxynivalenol levels in grain via use of microbial antagonists. Adv. Exp. Med. Biol. 2002, 504, 53–69. [Google Scholar] [PubMed]
- Huis, A.V.; Itterbeeck, J.V.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; FAO Forestry Paper (No. 171); FAO, United Nations: Rome, Italy, 2013. [Google Scholar]
- Cickova, H.; Newton, G.L.; Lacy, R.C.; Kozanek, M. The use of fly larvae for organic waste treatment. J. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Shelomi, M. Review of black soldier fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y.; Minor, M.; Morel, P.C.H.; Najar-Rodriguez, A.J. Bioconversion of three organic wastes by black soldier fly (Diptera: Stratiomyidae) larvae. Environ. Entomol. 2018, 47, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Howdeshell, T.; Tanaka, T. Recovery of glucose from dried distiller’s grain with solubles, using combinations of solid-state fermentation and insect culture. Can. J. Microbiol. 2018, 64, 706–715. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 18th ed.; AOAC International: Rockville, MD, USA, 2005. [Google Scholar]
- AACC. International Approved Methods 1–2; Cereal and Grain Association: St. Paul, MN, USA, 2009; ISBN 978-1-891127-68-2. [Google Scholar]
- Manurung, R.; Suprianta, A.; Rizkita, R.E.; Ramadhani, E.P. Bioconversion of rice straw waste by black soldier fly larvae (Hermetia illucens L.): Optimal feed rate for biomass production. J. Entomol. Zool. Stud. 2016, 4, 1036–1041. [Google Scholar]
- Kaya, M.; Sofi, K.; Sargin, I.; Mujtaba, M. Changes in physicochemical properties of chitin at developmental stages (larvae, pupa and adult) of Vespa crabro (wasp). Carbohydr. Polym. 2016, 145, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Schaarschmidt, S.; Fauhl-Hassek, C. The fate of mycotoxins during the processing of wheat for human consumption. Compr. Rev. Food Sci. Food Saf. 2018, 17, 556–593. [Google Scholar] [CrossRef]
- Goral, T.; Wisniewska, H.; Ochodzki, P.; Nielsen, L.K.; Walentyn-Goral, D.; Stepien, L. Relationship between Fusarium head blight, kernel damage, concentration of Fusarium biomass, and Fusarium toxins in grain of winter wheat inoculated with Fusarium culmorum. Toxins 2019, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; He, X.; Matsuo, Y.; Singh, P.K.; Kushiro, M. Analysis of the masked metabolite of deoxynivalenol and Fusarium resistance in CIMMYT wheat germplasm. Toxins 2017, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.K.R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Camenzuli, L.; Dam, R.V.; Rijk, T.; Andriessen, R.; Schelt, J.V.; Fles-Klerx, H.J.V. Tolerance and excretion of the mycotoxins aflatoxin B1, zearalenone, deoxynivalenol, and ochratoxin A by Alphitobius diaperinus and Hermetia illucens from contaminated substrates. Toxins 2018, 10, 91. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| DON Concentration (µg/g Spent Feed) * | ||||
|---|---|---|---|---|
| Day 0 | Day 4 | Day 8 | Day 12 | |
| Control | 0.63 ± 0.20 c,C | 0.15 ± 0.05 c,A | 4.17 ± 0.83 b,B | 6.72 ± 0.70 a,C |
| Lactobacillus plantarum | 2.60 ± 0.07 b,B | 0.20 ± 0.07 c,A | 13.66 ± 0.37 a,A | 14.17 ± 1.55 a,A |
| Aspergillus oryzae | 2.76 ± 0.27 b,B | 0.12 ± 0.02 c,A | 5.80 ± 0.88 a,B | 6.82 ± 1.82 a,B,C |
| L. plantarum + A. oryzae | 3.58 ± 0.18 b,A | 0.12 ± 0.05 b,A | 10.69 ± 1.77 a,A | 11.41 ± 2.25 a,A,B |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gulsunoglu, Z.; Aravind, S.; Bai, Y.; Wang, L.; Kutcher, H.R.; Tanaka, T. Deoxynivalenol (DON) Accumulation and Nutrient Recovery in Black Soldier Fly Larvae (Hermetia illucens) Fed Wheat Infected with Fusarium spp. Fermentation 2019, 5, 83. https://doi.org/10.3390/fermentation5030083
Gulsunoglu Z, Aravind S, Bai Y, Wang L, Kutcher HR, Tanaka T. Deoxynivalenol (DON) Accumulation and Nutrient Recovery in Black Soldier Fly Larvae (Hermetia illucens) Fed Wheat Infected with Fusarium spp. Fermentation. 2019; 5(3):83. https://doi.org/10.3390/fermentation5030083
Chicago/Turabian StyleGulsunoglu, Zehra, Smitha Aravind, Yuchen Bai, Lipu Wang, H. Randy Kutcher, and Takuji Tanaka. 2019. "Deoxynivalenol (DON) Accumulation and Nutrient Recovery in Black Soldier Fly Larvae (Hermetia illucens) Fed Wheat Infected with Fusarium spp." Fermentation 5, no. 3: 83. https://doi.org/10.3390/fermentation5030083
APA StyleGulsunoglu, Z., Aravind, S., Bai, Y., Wang, L., Kutcher, H. R., & Tanaka, T. (2019). Deoxynivalenol (DON) Accumulation and Nutrient Recovery in Black Soldier Fly Larvae (Hermetia illucens) Fed Wheat Infected with Fusarium spp. Fermentation, 5(3), 83. https://doi.org/10.3390/fermentation5030083