Anhydrobiosis in Yeasts: Changes in Mitochondrial Membranes Improve the Resistance of Saccharomyces cerevisiae Cells to Dehydration–Rehydration


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals, Yeast Strains, and Growth Conditions
2.2. Yeast Dehydration–Rehydration
2.3. Yeast Viability Determination
2.4. Determination of Plasma Membranes Permeability
2.5. Statistical Methods
3. Results and Discussion
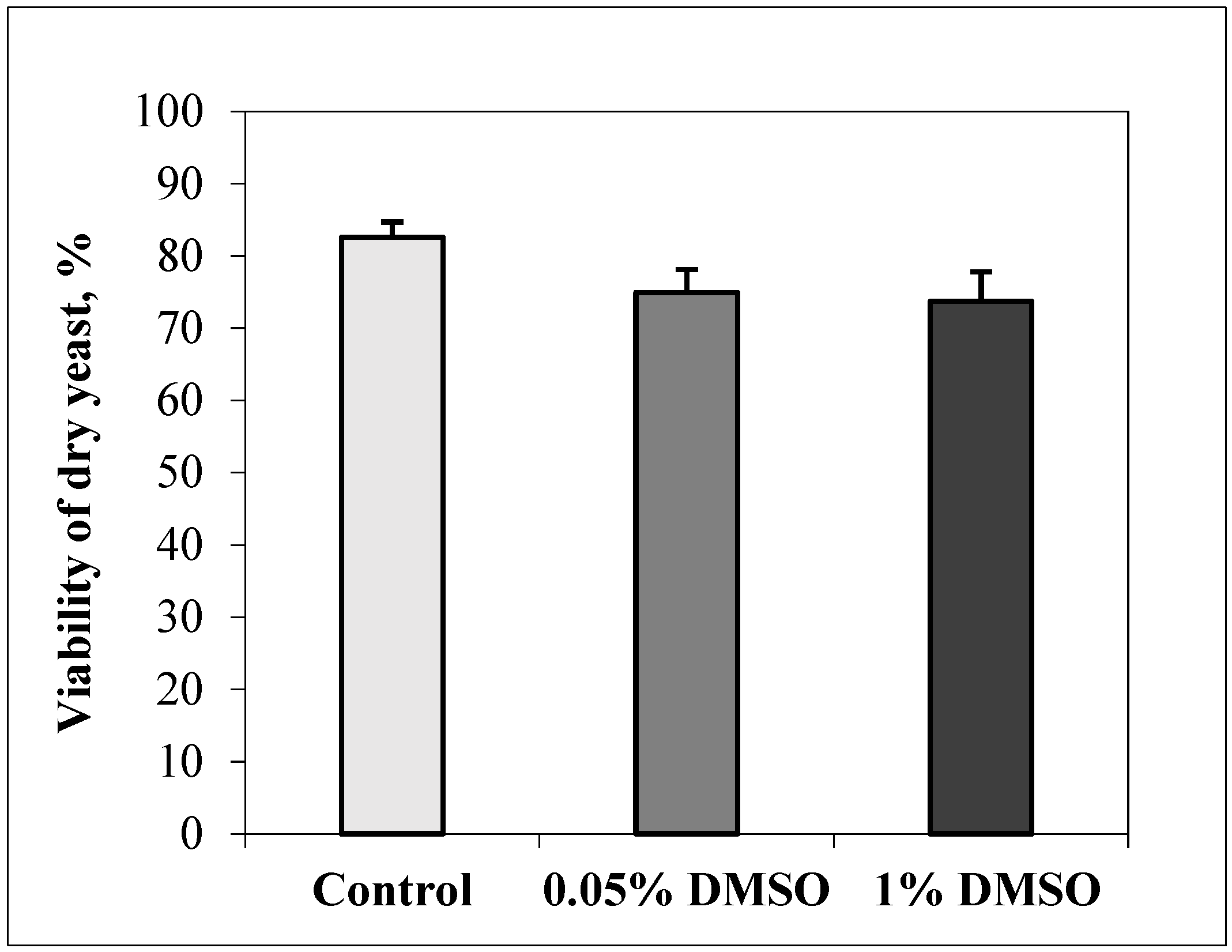
3.1. DMSO Influence on The Viability of Dry Yeast Cells
3.2. Determination of The Optimal Concentration of LCA
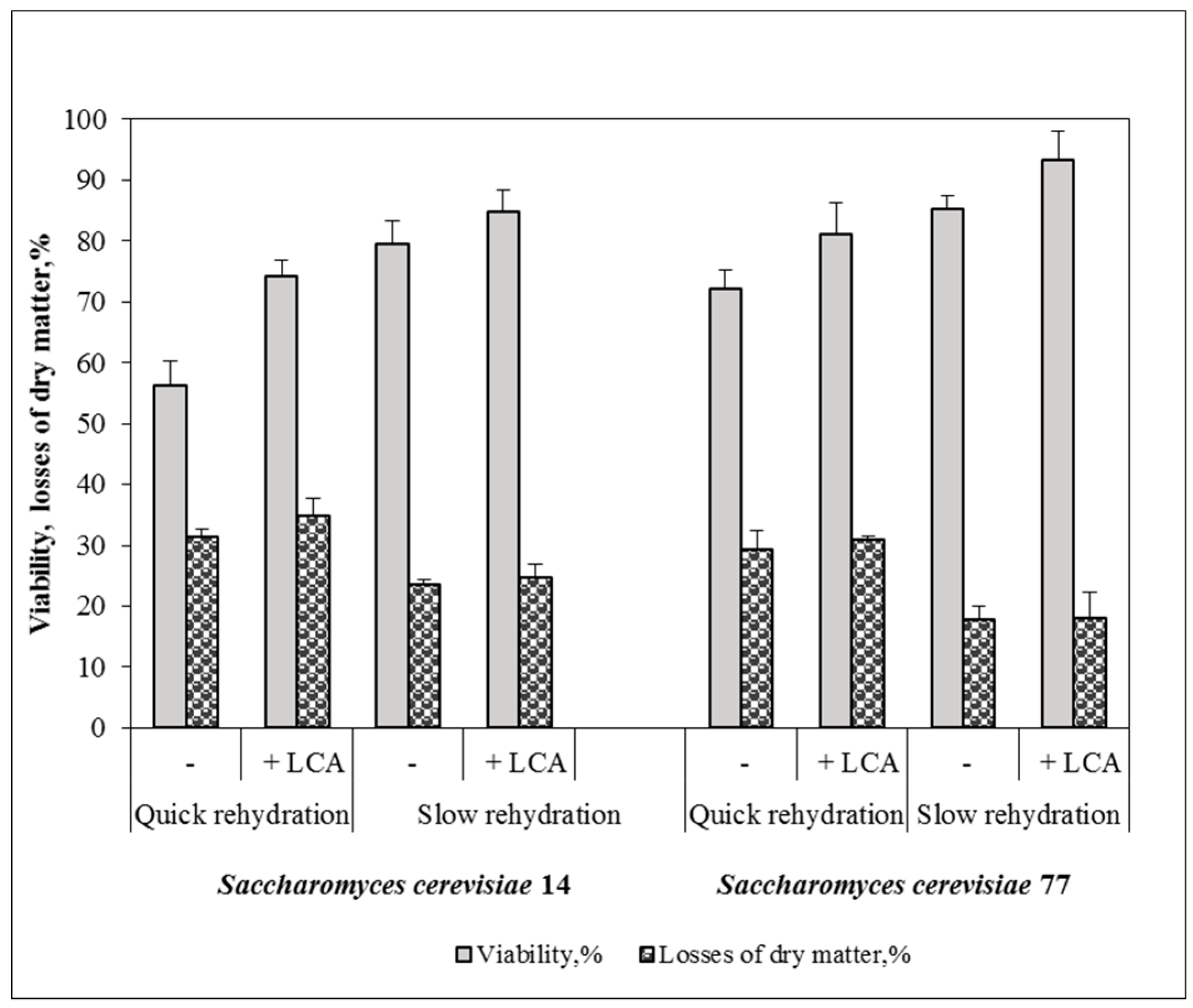
3.3. LCA Effects upon Resistance of Yeast to Dehydration–Rehydration Treatments
Author Contributions
Funding
Conflicts of Interest
References
- Crowe, J.H. Anhydrobiosis: An unsolved problem. Am. Nat. 1971, 105, 563–574. [Google Scholar] [CrossRef]
- Crowe, J.H.; Hoekstra, F.A.; Crowe, L.M. Anhydrobiosis. Annu. Rev. Physiol. 1992, 54, 579–599. [Google Scholar] [CrossRef] [PubMed]
- Crowe, J.H. Trehalose as a “chemical chaperone”: Fact and fantasy. Adv. Exp. Med. Biol. 2008, 594, 143–158. [Google Scholar]
- Crowe, J.H. Anhydrobiosis: An unsolved problem with applications in human welfare. Subcell. Biochem. 2015, 71, 263–280. [Google Scholar] [PubMed]
- Beker, M.J.; Rapoport, A.I. Conservation of yeasts by dehydration. Adv. Biochem. Eng. Biotechnol. 1987, 35, 127–171. [Google Scholar]
- Dupont, S.; Rapoport, A.; Gervais, P.; Beney, L. The survival kit of Saccharomyces cerevisiae for anhydrobiosis. Appl. Microbiol. Biotechnol. 2014, 98, 8821–8834. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, A. Anhydrobiosis and dehydration of yeasts. In Biotechnology of Yeasts and Filamentous Fungi; Sibirny, A., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 87–116. [Google Scholar]
- Rapoport, A.; Golovina, E.A.; Gervais, P.; Dupont, S.; Beney, L. Anhydrobiosis: Inside yeast cells. Biotechnol. Adv. 2019, 37, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Khroustalyova, G.M.; Adler, L.; Rapoport, A.I. Exponential growth phase cells of the osmotolerant yeast Debaryomyces hansenii are extremely resistant to dehydration stress. Proc. Biochem. 2001, 36, 1163–1166. [Google Scholar] [CrossRef]
- Rapoport, A.; Rusakova, A.; Khroustalyova, G.; Walker, G. Thermotolerance in Saccharomyces cerevisiae is linked to resistance to anhydrobiosis. Proc. Biochem. 2014, 49, 1889–1892. [Google Scholar] [CrossRef]
- Khroustalyova, G.; Giovannitti, G.; Severini, D.; Scherbaka, R.; Turchetti, B.; Buzzini, P.; Rapoport, A. Anhydrobiosis in yeasts: Psychrotolerant yeasts are highly resistant to dehydration. Yeast 2019, 36, 375–379. [Google Scholar] [CrossRef]
- Rapoport, A.I.; Beker, M.E. Effect of sucrose and lactose on resistance of the yeast Saccharomyces cerevisiae to dehydration. Microbiology 1983, 52, 556–559. [Google Scholar]
- Rapoport, A.I.; Khroustalyova, G.M.; Crowe, L.M.; Crowe, J.H. Anhydrobiosis in yeast: Stabilization by exogenous lactose. Microbiology 2009, 78, 624–629. [Google Scholar] [CrossRef]
- Rozenfelde, L.; Rapoport, A. Anhydrobiosis in yeast: Is it possible to reach anhydrobiosis for yeast grown in conditions with severe oxygen limitation? Antonie van Leeuwenhoek J. Microbiol. 2014, 106, 211–217. [Google Scholar]
- Grube, M.; Gavare, M.; Rozenfelde, L.; Rapoport, A. Anhydrobiosis in yeast: FT-IR spectroscopic studies of yeast grown under conditions of severe oxygen limitation. Biotechnol. Appl. Biochem. 2014, 61, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Kulikova-Borovikova, D.; Lisi, S.; Dauss, E.; Alamae, T.; Buzzini, P.; Hallsworth, J.E.; Rapoport, A. Activity of the a-glucoside transporter Agt1 in Saccharomyces cerevisiae cells during dehydration-rehydration events. Fungal Biol. 2018, 122, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, A.; Turchetti, B.; Buzzini, P. Application of anhydrobiosis and dehydration of yeasts for non-conventional biotechnological goals. World J. Microbiol. Biotechnol. 2016, 32, 104. [Google Scholar] [CrossRef] [PubMed]
- Panek, A.D. Trehalose metabolism and its role in Saccharomyces cerevisiae. J. Biotechnol. 1985, 3, 121–130. [Google Scholar] [CrossRef]
- Panek, A.D. Trehalose metabolism—New horizons in technological applications. Braz. J. Med. Biol. Res. 1995, 28, 169–181. [Google Scholar]
- Eleutherio, E.C.A.; de Araujo, P.S.; Panek, A.D. Role of the trehalose carrier in dehydration resistance of Saccharomyces cerevisiae. Biochim. Biophys. Acta 1993, 1156, 263–266. [Google Scholar] [CrossRef]
- Eleutherio, E.; Panek, A.; de Mesquita, J.F.; Trevisol, D.; Magalhães, R. Revisiting yeast trehalose metabolism. Curr. Genet. 2015, 61, 263–274. [Google Scholar] [CrossRef]
- Leslie, S.B.; Teter, S.A.; Crowe, L.M.; Crowe, J.H. Trehalose lowers membrane phase transitions in dry yeast cells. Biochim. Biophys. Acta 1994, 1192, 7–13. [Google Scholar] [CrossRef]
- Rapoport, A.I.; Puzyrevskaya, O.M.; Saubenova, M.G. Polyols and resistance of yeasts to dehydration. Microbiology 1988, 57, 269–271. [Google Scholar]
- Krallish, I.; Jeppsson, H.; Rapoport, A.; Hahn-Hagerdal, B. Effect of xylitol and trehalose on dry resistance of yeasts. Appl. Microbiol. Biotechnol. 1997, 47, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Sakai, K.; Morida, K.; Nakamori, S. Proline accumulation by mutation or disruption of the proline oxidase gene improves resistance to freezing and desiccation stresses in Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2000, 184, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H. Proline as a stress protectant in yeast: Physiological functions, metabolic regulations, and biotechnological applications. Appl. Microbiol. Biotechnol. 2008, 81, 211–223. [Google Scholar] [CrossRef]
- Espindola, A.D.S.; Gomes, D.S.; Panek, A.D.; Eleutherio, E.C.A. The role of glutathione in yeast dehydration tolerance. Cryobiology 2003, 47, 236–241. [Google Scholar] [CrossRef]
- Gamero-Sandemetrio, E.; Paya-Tormo, L.; Gomez-Pastor, R.; Aranda, A.; Matallana, E. Non-canonical regulation of glutathione and trehalose biosynthesis characterizes non-Saccharomyces wine yeasts with poor performance in active dry yeast production. Microb. Cell 2018, 5, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Câmara Júnior, A.D.A.; Nguyen, T.D.; Jossier, A.; Endrizzi, A.; Saurel, R.; Simonin, H.; Husson, F. Improving total glutathione and trehalose contents in Saccharomyces cerevisiae cells to enhance their resistance to fluidized bed drying. Proc. Biochem. 2018, 69, 45–51. [Google Scholar] [CrossRef]
- Câmara, A.D.A.; Maréchal, P.-A.; Tourdot-Maréchal, R.; Husson, F. Dehydration stress responses of yeasts Torulaspora delbrueckii, Metschnikowia pulcherrima and Lachancea thermotolerans: Effects of glutathione and trehalose biosynthesis. Food Microbiol. 2019, 79, 137–146. [Google Scholar] [CrossRef]
- Kulikova-Borovikova, D.; Khroustalyova, G.; Chang, C.-R.; Daugelavicius, R.; Yurkiv, M.; Ruchala, J.; Sibirny, A.; Rapoport, A. Anhydrobiosis in yeast: Glutathione overproduction improves resistance to dehydration of a recombinant Ogataea (Hansenula) polymorpha strain. Proc. Biochem. 2018, 71, 41–44. [Google Scholar] [CrossRef]
- Guzhova, I.; Krallish, I.; Khroustalyova, G.; Margulis, B.; Rapoport, A. Dehydration of yeast: Changes in the intracellular content of Hsp70 family proteins. Proc. Biochem. 2008, 43, 1138–1141. [Google Scholar] [CrossRef]
- Capece, A.; Votta, S.; Guaragnella, N.; Zambuto, M.; Romaniello, R.; Romano, P. Comparative study of Saccharomyces cerevisiae wine strains to identify potential marker genes correlated to desiccation stress tolerance. FEMS Yeast Res. 2016, 16, fow015. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, A.I.; Kostrikina, N.A. Cytological investigation of an anabiosis state of yeast organisms. Proc. Acad. Sci. USSR Biol. 1973, 5, 770–773. [Google Scholar]
- Shima, J.; Ando, A.; Takagi, H. Possible roles of vacuolar H+-ATPase and mitochondrial function in tolerance to air-drying stress revealed by genome-wide screening of Saccharomyces cerevisiae deletion strains. Yeast 2008, 25, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Calahan, D.; Dunham, M.; Desevo, C.; Koshland, D.E. Genetic analysis of desiccation tolerance in Saccharomyces cerevisiae. Genetics 2011, 189, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Picazo, C.; Gamero-Sandemetrio, E.; Orozco, H.; Albertin, W.; Marullo, P.; Matallana, E.; Aranda, A. Mitochondria inheritance is a key factor for tolerance to dehydration in wine yeast production. Lett. Appl. Microbiol. 2015, 60, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Koupaki, O.; Boukh-Viner, T.; Gregg, C.; Juneau, M.; English, A.M.; Thomas, D.Y.; Titorenko, V.I. Chemical genetic screen identifies lithocholic acid as an anti-aging compound that extends yeast chronological life span in a TOR-independent manner, by modulating housekeeping longevity assurance processes. Aging 2010, 2, 393–414. [Google Scholar]
- Beach, A.; Richard, V.R.; Leonov, A.; Burstein, M.T.; Bourque, S.D.; Koupaki, O.; Juneau, M.; Feldman, R.; Iouk, T.; Titorenko, V.I. Mitochondrial membrane lipidome defines yeast longevity. Aging 2013, 5, 551–574. [Google Scholar] [CrossRef] [PubMed]
- Beach, A.; Richard, V.R.; Bourque, S.; Boukh-Viner, T.; Kyryakov, P.; Gomez-Perez, A.; Arlia Ciommo, A.; Feldman, R.; Leonov, A.; Piano, A.; et al. Lithocholic bile acid accumulated in yeast mitochondria orchestrates a development of an anti-aging cellular pattern by causing age-related changes in cellular proteome. Cell Cycle 2015, 14, 1643–1656. [Google Scholar] [CrossRef]
- Medkour, Y.; Dakik, P.; McAuley, M.; Mohammad, K.; Mitrofanova, D.; Titorenko, V.I. Mechanisms underlying the essential role of mitochondrial membrane lipids in yeast chronological aging. Oxid. Med. Cell. Long. 2017, 2017, 2916985. [Google Scholar] [CrossRef]
- Rapoport, A.I.; Khrustaleva, G.M.; Chamanis, G.J.; Beker, M.E. Yeast anhydrobiosis: Permeability of the plasma membrane. Mikrobiology 1995, 64, 275–278. [Google Scholar]
- Rapoport, A.I.; Meissel, M.N. Survival rates of yeast organisms after dehydration as determined by fluorescence microscopy. Microbiology 1985, 54, 53–55. [Google Scholar]
- Brayton, C.F. Dimethyl sulfoxide (DMSO): A review. Cornell Vet. 1986, 76, 61–90. [Google Scholar] [PubMed]
- Beker, M.J.; Blumbergs, J.E.; Ventina, E.J.; Rapoport, A.I. Characteristics of cellular membranes at rehydration of dehydrated yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 1984, 19, 347–352. [Google Scholar] [CrossRef]
- Borovikova, D.; Teparic, R.; Mrsa, V.; Rapoport, A. Anhydrobiosis in yeast: Cell wall mannoproteins are important for yeast Saccharomyces cerevisiae resistance to dehydration. Yeast 2016, 33, 347–353. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample | Viability, % |
|---|---|
| S. cerevisiae 77 | 61.0 ± 3.8 |
| S. cerevisiae 77 + LCA 50 µM (0.05% DMSO) | 54.0 ± 4.7 |
| S. cerevisiae 77 + LCA 50 µM (1% DMSO) | 68.6 ± 1.6 |
| S. cerevisiae 77 + LCA 100 µM (0.05% DMSO) | 50.9 ± 3.1 |
| S. cerevisiae 77 + LCA 100 µM (1% DMSO) | 69.5 ± 2.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khroustalyova, G.; Rapoport, A. Anhydrobiosis in Yeasts: Changes in Mitochondrial Membranes Improve the Resistance of Saccharomyces cerevisiae Cells to Dehydration–Rehydration. Fermentation 2019, 5, 82. https://doi.org/10.3390/fermentation5030082
Khroustalyova G, Rapoport A. Anhydrobiosis in Yeasts: Changes in Mitochondrial Membranes Improve the Resistance of Saccharomyces cerevisiae Cells to Dehydration–Rehydration. Fermentation. 2019; 5(3):82. https://doi.org/10.3390/fermentation5030082
Chicago/Turabian StyleKhroustalyova, Galina, and Alexander Rapoport. 2019. "Anhydrobiosis in Yeasts: Changes in Mitochondrial Membranes Improve the Resistance of Saccharomyces cerevisiae Cells to Dehydration–Rehydration" Fermentation 5, no. 3: 82. https://doi.org/10.3390/fermentation5030082
APA StyleKhroustalyova, G., & Rapoport, A. (2019). Anhydrobiosis in Yeasts: Changes in Mitochondrial Membranes Improve the Resistance of Saccharomyces cerevisiae Cells to Dehydration–Rehydration. Fermentation, 5(3), 82. https://doi.org/10.3390/fermentation5030082