Abstract
Aerobically cultivated cells of Corynebacterium glutamicum produce mixed organic acids, including succinic acid (SA), from glucose when the cells are transferred to oxygen-deprived conditions. Genetic modification, including inactivation of lactate dehydrogenase and overexpression of pyruvate carboxylase, allows this microbe to be an efficient SA producer under the conditions of oxygen deprivation. High productivity and high titers can be achieved in the production process by using the genetically engineered strain of C. glutamicum under the given conditions. However, glucose consumption for cell preparation decreases process yield (defined as the quantity of SA produced divided by the total quantity of glucose used in cell preparation and SA production). In this study, we investigated cell recycle fed-batch fermentation for SA production to improve the process yield by reducing the effect of glucose consumption for cell preparation on the process yield. A genetically stable and markerless strain, harboring nine genomic copies of the pyruvate carboxylase gene, was newly constructed and used for cell recycle fermentation. During 26 reaction cycles, only 0.7% decrease in specific productivity per reaction was observed. Overall, the process yield was improved by 79% compared to that in a single fed-batch reaction without cell recycling.
1. Introduction
Succinic acid (SA), a C4 dicarboxylic acid, is used as a building block for specialty chemicals such as surfactants and chelators and as an additive in the pharmaceutical and food industries [1]. SA has emerged as a feedstock to produce valuable molecules, especially in the polymer industry, such as 1,4-butanediol, tetrahydrofuran, and adipic acid [2]. Although SA has traditionally been produced from fossil resources, biomass-based SA has been produced on a commercial scale since 2014 [3].
Corynebacterium glutamicum is used for the industrial production of amino acids [4]. Recently, this Gram-positive microbe has also been utilized for the production of fuels and commodity chemicals including ethanol [5,6], isobutanol [7,8,9], d-lactic acid [10], cadaverine [11], putrescine [12], xylitol [13], 2,3-butanediol [14,15], and SA. SA-producing strains of C. glutamicum have been developed separately for aerobic [16,17,18,19] and anaerobic processes [20,21,22,23,24]. To date, anaerobic processes with C. glutamicum have been found to be superior to aerobic processes in terms of the productivity, titer, and yield of SA [20,21]. C. glutamicum naturally produces mixed acids including SA, lactic acid, and acetic acid from sugars under oxygen-deprived conditions, though cellular growth is basically suppressed during the conversion reaction [25,26]. Metabolic engineering has enabled the development of a dedicated C. glutamicum strain for SA production under conditions of oxygen deprivation [20,21]. In production processes using these engineered strains, the cells are cultured aerobically and subsequently subjected to oxygen deprivation to produce SA from sugar. A similar process has also been applied to SA-producing Escherichia coli to improve productivity [27]. A major shortcoming of this two-step process is that sugar consumption for cell preparation reduces the integrated process yield, which becomes a crucial issue in industrialization. In this study, we employed the cell recycle method to improve process yield in the two-step process for the production of SA by C. glutamicum. A cell recycle bioprocess exhibits very high volumetric productivity since cell density in the cell recycle process is capable of being maintained higher than that of a typical batch fermentation process, resulting in reduction of capital costs by shortening fermentation time [28]. As a result, high specific productivity was maintained over 26 rounds of cell recycle, and the amount of glucose used for cell preparation accounted for only 5.1% of the total glucose used in the process. Furthermore, we newly developed a plasmid-free SA-producing strain, as plasmids should be removed from industrial strains to maintain genetic stability and reduce the costs of antibiotics. This study demonstrates the potential utility of C. glutamicum for the industrial production of SA.
2. Materials and Methods
2.1. Bacterial Strain and Media
C. glutamicum strain R (JCM 18229) and its derivatives (shown in Table 1) were grown on a nutrient-rich medium (A-medium) containing (per liter) 7 g (NH4)2SO4, 2 g urea, 0.5 g KH2PO4, 0.5 g K2HPO4, 0.5 g MgSO4·7H2O, 6 mg FeSO4·7H2O, 4.2 mg MnSO4·H2O, 0.2 mg biotin (Wako Pure Chemical, Osaka, Japan), 0.2 mg thiamine (Wako Pure Chemical), 2 g yeast extract and 7 g Casamino acids, and supplemented with 40 g glucose [29]. Where appropriate, media were supplemented with kanamycin (50 μg/mL).

Table 1.
Microbial strains and plasmids used in this study.
2.2. DNA Manipulation
To overexpress the pyc gene, DNA fragments containing the coding sequence and the promoter region of the pyc gene were excised from a pyc-expression plasmid pCRA717 [21] by restriction digestion with XbaI and SphI. The amplicon was integrated into C. glutamicum R strain-specific islands (SSIs) via a Cre/loxP-mediated system [30] or a multiple-gene transfer system using mutant lox sequence [31,32]. C. glutamicum R SSIs are discovered through comparative analysis of the genome sequences for three C. glutamicum strains [29]. Strain specific genome regions exist as “islands” in the common genome backbone, suggesting that SSIs are possibly dispensable regions for basic functions of the cells [30]. The latter technique showed higher recombination efficiency by 2 orders of magnitude compared to that of the homologous recombination technique. The resulting recombinant strain carries no antibiotic marker. In this study, a series of markerless mutants bearing up to 9 copies of the pyc gene were constructed. Detailed procedures for constructing SA-producing strains are described in supplementary material.
2.3. SA Production under Conditions of Oxygen Deprivation
For single batch reaction, C. glutamicum strains were aerobically cultivated in a 2 L flask containing 500 mL of A-medium with 4% glucose for 13 h. Cells were harvested by centrifugation (5000× g, 10 min, 4 °C) and were washed once with BT-solution (A-medium without yeast extract, casamino acids and urea). The cells were subsequently resuspended in BT-solution to a concentration corresponding to 40 g cell dry weight (CDW)/liter (L) incubated at 33 °C. Succinic acid production was started by addition of 400 mM glucose and 300 mM NaHCO3. Glucose and NaHCO3 were intermittently added to avoid depletion. Oxygen deprivation (dissolved oxygen concentration lower than 0.01 ppm) was achieved by no aeration and gentle agitation. The pH was maintained at 7.5 by addition of 2.5 M NaOH using a pH controller (DT-10023, Biott Co. Ltd, Tokyo, Japan). Cell mass was calculated by using a spectrophotometer and an absorbance of 1 at 610 nm (A610 = 1) corresponded to 0.39 g CDW per liter.
For cell recycle reaction, the C. glutamicum strain was aerobically cultivated in a 1 L jar fermenter (Biott) containing 500 mL of A-medium without urea, and supplemented with 4% glucose at 33 °C. Aeration rate and rotation rate of disc-turbine impellers were 0.5 L/min and 1000 rpm, respectively. The pH of the medium was maintained at 7.5 by addition of 2.5 M NaOH. After cultivation for 10 h, the culture broth was used to inoculate a 5 L jar fermenter (Biott) containing 3 L of BT-solution with 2% corn steep liquor. Glucose was initially supplemented to be 4% and followed by two times of 4% addition. The fermenter was operated at 33 °C, and the pH was maintained at 7.5 using 5 M NaOH. The rates of aeration and impeller blade rotation were 3 L/min and 700 rpm, respectively. Following cultivation for 7.5 h, the cells were collected and washed as described above. The cells were then resuspended in the minimal salts solution at a cell concentration of 20 g CDW/L and incubated at 33 °C. Reaction for SA production was initiated by the addition of glucose (400 mM) and sodium hydrogen carbonate (300 mM). Equal amounts of glucose and sodium hydrogen carbonate were added after 5 h. The cells were collected by centrifugation after 22 h and were used for the next reaction cycle.
2.4. Analytical Techniques
SA concentration was determined by HPLC (Model 8020, Tosoh, Yamaguchi, Japan) performed on a TSKgel OApak-A column (Tosoh) operating at 40 °C, with a 0.75 mM H2SO4 mobile phase at a flow rate of 1.0 mL/min. Glucose concentration was determined using an enzyme electrode glucose sensor (BF-4, Oji Scientific Instruments, Hyogo, Japan).
2.5. Detection of Biotinylated Protein
The cells cultivated in 10 mL of A-medium were collected by centrifugation (5,000 × g, 10 min) and were resuspended in 2.0 mL of buffer solution containing 100 mM Tris–HCl (pH 7.5), 20 mM KCl, 5 mM MnSO4, 20 mM MgCl2, 1 mM EDTA, and 2 mM dithiothreitol. The cells were subsequently disrupted by using a bead beater (Multi-Beads Shocker, Yasui Kikai, Osaka, Japan) at 2500 rpm for 10 cycles of 30 s each, at 4 °C. Soluble protein was separated by centrifugation (20,000 × g, 20 min) and then heat-denatured to resolve the components on12% SDS-PAGE. Separated proteins were blotted onto a polyvinylidene difluoride membrane (Clear Blot Membrane-P, ATTO, Tokyo, Japan) by using a Trans-Blot SD cell (Bio-Rad, Hercules, CA, USA). The membrane was incubated withblocking buffer (Blocking One, Nacalai Tesque, Kyoto, Japan) for 1 h. It was then transferred to a solution containing 0.2 μg/mLstreptavidin-alkaline phosphatase conjugate (Vector Laboratories, Burlingame, CA, USA) in TBS-T buffer (20 mM Tris, 150 mM NaCl and 0.1% Tween 20) for 1 h at 25 °C. After rinsing thrice with TBS-T buffer for 5 min, the membrane was incubated with detection buffer (0.2 mg/mL 4-Nitro blue tetrazolium chloride, 0.1 mg/mL 5-Bromo-4-chloro-3-indolyl-phosphate in TBS-T buffer). The gel image was semi-quantitatively analyzed with ImageJ [33].
3. Results
3.1. Construction of a Plasmid-Free Strain for SA Production
A 3.8-kb nucleotide fragment, containing the native promoter and the coding sequences for the pyc gene was integrated into non-essential SSIs of the C. glutamicum genome to construct a plasmid-free strain for SA production [30]. We first employed a Cre/loxP-mediated gene transfer system [30] to integrate the pyc gene into the genome and succeeded in constructing a strain harboring two exogenous copies of the pyc gene. The resulting strain, named CRZ19, showed lower volumetric productivity than that of the plasmid-bearing CRZ1-pCRA717 strain (Table 2). These results suggest that more copies of the pyc gene were required to exhibit SA volumetric productivity similar to that of the CRZ1-pCRA717 strain, as a copy number of the pBL1-based plasmid pCRA717 is estimated to be 10–30 [34]. To introduce more copies of pyc, a multiple-gene transfer system using a mutant lox sequence [31,32] was employed since this technique has a very high recombination efficiency. Using this technique, up to nine copies of the pyc gene were successfully integrated into the genome. Pyruvate carboxylase (PYC) of C. glutamicum is unstable and its enzyme activity was not detected in oxygen-deprived cells [25]. Therefore, we analyzed biotinylated proteins by using streptavidin-alkaline phosphatase conjugate since PYC is a biotin-containing enzyme. Figure 1 shows analysis of biotinylated proteins in mutant strains harboring exogenous 2 (CRZ19), 4 (CRZ20), and 9 copies (CRZ21) of the pyc gene, and the parental CRZ1 strain to compare the amounts of PYC. The 125-kDa band was designated as PYC protein based on our previous study that showed absence of this band in a pyc deletion mutant [25]. It was observed that the band became progressively denser with increasing copy numbers of pyc and was the densest in the lane corresponding to CRZ1-pCRA717. On the other hand, the bands close to 66 kDa, expected to correspond to the biotinylated α-subunit of acetyl-CoA carboxylase (ACC) [35,36], were virtually unchanged. These results suggest that the amount of PYC protein increased in proportion to the copy number of the introduced gene. The production of SA increased with increase in the copy number of the pyc gene, and the volumetric productivity of the CRZ21 strain was almost identical to that of the plasmid-bearing strain (Table 2). Thus, we chose this strain as a biocatalyst in cell recycle fermentation.
Table 2.
Volumetric productivity of succinic acid and exogenous copies of pyc in engineered strains a.
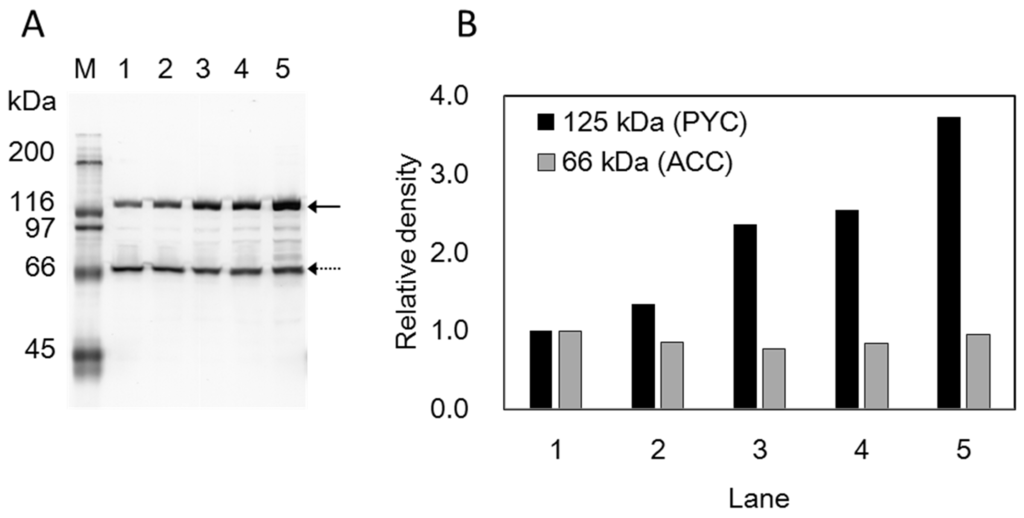
Figure 1.
Analysis of biotinylated proteins by using streptavidin-alkaline phosphatase conjugate. Cell-free extracts (2 μg-proteinper lane) from CRZ1 (lane 1), CRZ19 (lane 2), CRZ20 (lane 3), CRZ21 (lane 4), and CRZ1-pCRA717 (lane 5) strains were separated by SDS-PAGE on 12% gel (A). MW 125 kDa (solid arrow) and 66 kDa (dotted arrow) were estimated in reference to the protein marker (M). (B) shows relative densities of the 125 kDa (black) and 66 kDa bands (gray) in panel A. Density of each band (PYC or ACC) in lanes 2 to 5 was compared with the corresponding band in lane 1 by using the ImageJ software.
3.2. Cell Recycle Fed-Batch Reactions
Figure 2 shows the results of cell recycle fed-batch reactions for SA production using CRZ21. Each reaction was terminated at 22 h and the cells were subsequently collected by centrifugation and used for the next reaction. SA production in the first reaction was considerably lower than in the second reaction because the glucose feed failed, and consequently, the cells were starved for glucose for several hours. However, the cells collected from the first reaction showed pronounced recovery in productivities and yield of SA. In the course of the cell recycle reaction, SA volumetric productivity (amount of SA in g/(l h)) gradually decreased (Figure 2A), while specific productivity [amount of SA in g/(CDW in g h)](Figure 2B). The reaction yield was largely maintained until the 19th cycle and subsequently increased gradually from cycle 20th to 26th. This increase in the production yield might be accounted for decreased SA titer in those reaction cycles (Figure 2C,D). Since cells were collected by centrifugation, a small amount of cell loss occurred during each centrifugation step, and consequently cell concentration decreased in every reaction cycle. The slope of regression line generated from the data of the 2nd to 26th reaction cycles indicated 0.7% reduction in the specific productivity per cycle (Figure 2B). Table 3 shows mass balance analysis of the amount of consumed glucose to the amount of SA produced in the cell recycle reaction. Overall, 634 g of SA was produced in 26 reaction cycles. The total amount of glucose used for cell preparation and SA production was 939 g, and 48 g of glucose used for cell preparation accounted for only 5.1% of the total glucose used in the entire fermentation process. Out of the 26 reaction cycles, maximum volumetric productivity was observed in the 4th reaction cycle, where 38 g of SA was produced from 101 g of consumed glucose. Thus, if the cells are replaced at every reaction cycle, the process yield is calculated to be 0.38 g/g (Figure 2C), which is much lower than the process yield of 0.68 g/g obtained in the cell recycle reaction. This calculation indicates that cell recycle methodology helps in 79% improving the process yield for SA production by C. glutamicum under conditions of oxygen deprivation.
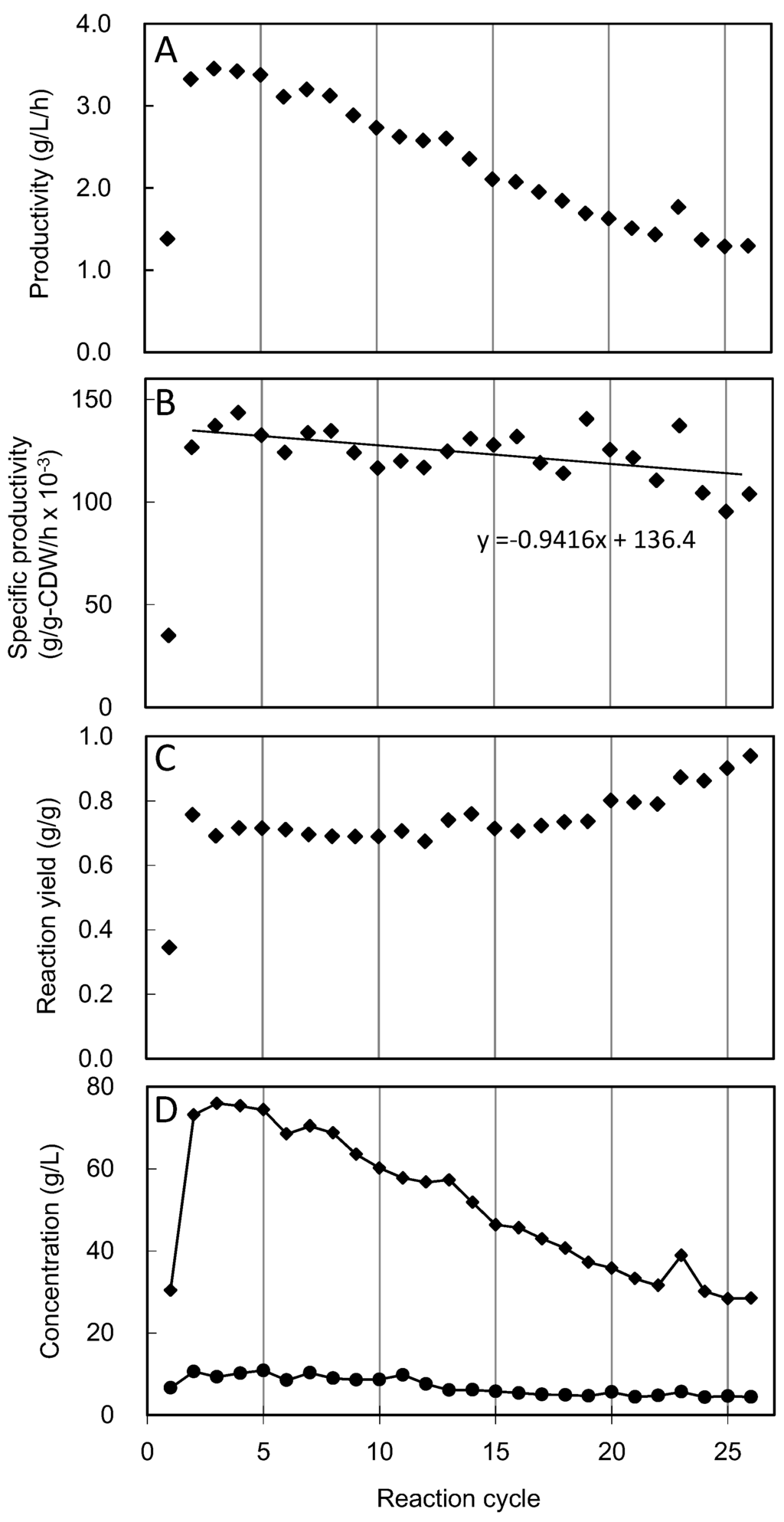
Figure 2.
Cell recycle fed-batch reaction using CRZ21 strain of C. glutamicum under conditions of oxygen deprivation. Diamonds indicate volumetric productivity (A), specific productivity (B), reaction yield (C) and concentrations of succinic acid and acetic acid (D) in each reaction. The reaction yield is defined as the amount of SA produced (g)/amount of glucose consumed (g), during a single reaction. Regression line in panel B was fitted to the data from the 2nd to the 26th reaction cycles.
Table 3.
Mass balance analysis of cell recycle fed-batch reactions for succinic acid production by C. glutamicum.
4. Discussion
In this study, we successfully applied cell recycle fed-batch reaction for SA production by C. glutamicum under oxygen deprivation. It has been observed that in the process using oxygen-deprived C. glutamicum cells, glucose consumption for cell preparation depresses the process yield [20]. To address this issue, a metabolically engineered strain (BOL-3/pAN6-gap), which can use formate as an electron donor, was developed [20]. This strain showed high process yield (0.89 g/g), though 36 g of formate was needed to produce 134 g of SA, thus adding to the cost of raw materials. The results obtained in this study clearly demonstrate the effectiveness of cell recycle fermentation to improve the process yield for SA production by C. glutamicum.
Although the specific productivity was largely maintained throughout the 26 reaction cycles (Figure 2B), SA volumetric productivity was depressed at every batch reaction due to the loss of cells owing to centrifugation (Figure 2A). A separation membrane has been successfully used in a continuous process for the production of SA by Actinobacillus succinogenes [37]; therefore, it can be expected that the integration of a membrane separation system with the cell recycle reaction developed in this study will result in improved volumetric productivity. Downstream processing to purify SA is a remaining challenge for industrialization of the process developed in this study.
5. Conclusions
We developed a plasmid-free strain of C. glutamicum for SA production by cell recycle fermentation and demonstrated significant improvement in the process yield during 26 reaction cycles. Furthermore, C. glutamicum cells maintained metabolic activity for 572 h (22 h × 26), which is the longest reaction time for fermentation using oxygen-deprived C. glutamicum cells.
Supplementary Materials
Supplementary materials can be found at http://www.mdpi.com/2311-5637/2/1/5/s1.
Acknowledgments
This work was partially supported by New Energy and Industrial Technology Development Organization (NEDO) of Japan and the Ministry of Economy, Trade and Industry (Japan).
Author Contributions
Maysayuki Inui, Hideaki Yukawa and Shohei Okino conceived and designed the experiments; Ryoji Noburyu and Masako Suda performed the experiments; Toru Jojima analyzed the data and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McKinlay, J.B.; Vieille, C.; Zeikus, J.G. Prospects for a bio-based succinate industry. Appl. Microbiol. Biotechnol. 2007, 76, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Thakker, C.; Martinez, I.; San, K.Y.; Bennett, G.N. Succinate production in Escherichia coli. Biotechnol. J. 2012, 7, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Succinity. Succinity Produces First Commercial Quantities of Biobased Succinic Acid. Available online: http://www.succinity.com/9-news/9-first-commercial-succinic-acid (accessed on 1 December 2015).
- Ikeda, M.; Takeno, S. Amino acid production by Corynebacterium glutamicum. In Corynebacterium Glutamicum; Yukawa, H., Inui, M., Eds.; Springer: Heidelberg, Germany, 2013; Volume 23, pp. 107–147. [Google Scholar]
- Inui, M.; Kawaguchi, H.; Murakami, S.; Vertès, A.A.; Yukawa, H. Metabolic engineering of Corynebacterium glutamicum for fuel ethanol production under oxygen-deprivation conditions. J. Mol. Microbiol. Biotechnol. 2004, 8, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Jojima, T.; Noburyu, R.; Sasaki, M.; Tajima, T.; Suda, M.; Yukawa, H.; Inui, M. Metabolic engineering for improved production of ethanol by Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2015, 99, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Suda, M.; Niimi, S.; Inui, M.; Yukawa, H. Strain optimization for efficient isobutanol production using Corynebacterium glutamicum under oxygen deprivation. Biotechnol. Bioeng. 2013, 110, 2938–2948. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.M.; Cho, K.M.; Liao, J.C. Engineering Corynebacterium glutamicum for isobutanol production. Appl. Microbiol. Biotechnol. 2010, 87, 1045–1055. [Google Scholar] [CrossRef] [PubMed]
- Blombach, B.; Riester, T.; Wieschalka, S.; Ziert, C.; Youn, J.W.; Wendisch, V.F.; Eikmanns, B.J. Corynebacterium glutamicum tailored for efficient isobutanol production. Appl. Environ. Microbiol. 2011, 77, 3300–3310. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, Y.; Yamamoto, S.; Suda, M.; Inui, M.; Yukawa, H. Reactions upstream of glycerate-1,3-bisphosphate drive Corynebacterium glutamicum d-lactate productivity under oxygen deprivation. Appl. Microbiol. Biotechnol. 2013, 97, 6693–6703. [Google Scholar] [CrossRef] [PubMed]
- Mimitsuka, T.; Sawai, H.; Hatsu, M.; Yamada, K. Metabolic engineering of Corynebacterium glutamicum for cadaverine fermentation. Biosci. Biotechnol. Biochem. 2007, 71, 2130–2135. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Wendisch, V.F. Putrescine production by engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2010, 88, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Jojima, T.; Inui, M.; Yukawa, H. Xylitol production by recombinant Corynebacterium glutamicum under oxygen deprivation. Appl. Microbiol. Biotechnol. 2010, 86, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kim, B.; Kim, H.; Kweon, Y.; Lee, S.; Lee, J. Industrial Production of 2,3-Butanediol from the Engineered Corynebacterium glutamicum. Appl. Biochem. Biotechnol. 2015, 176, 2303–2313. [Google Scholar] [CrossRef] [PubMed]
- Radoš, D.; Turner, D.L.; Fonseca, L.L.; Carvalho, A.L.; Blombach, B.; Eikmanns, B.J.; Neves, A.R.; Santos, H. Carbon flux analysis by 13C nuclear magnetic resonance to determine the effect of CO2 on anaerobic succinate production by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2014, 80, 3015–3024. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhu, N.; Xia, H. Aerobic production of succinate from arabinose by metabolically engineered Corynebacterium glutamicum. Bioresour. Technol. 2014, 151, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Litsanov, B.; Brocker, M.; Bott, M. Glycerol as a substrate for aerobic succinate production in minimal medium with Corynebacterium glutamicum. Microb. Biotechnol. 2013, 6, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Litsanov, B.; Kabus, A.; Brocker, M.; Bott, M. Efficient aerobic succinate production from glucose in minimal medium with Corynebacterium glutamicum. Microb. Biotechnol. 2012, 5, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Xia, H.; Wang, Z.; Zhao, X.; Chen, T. Engineering of acetate recycling and citrate synthase to improve aerobic succinate production in Corynebacterium glutamicum. PLoS ONE 2013, 8, e60659. [Google Scholar] [CrossRef] [PubMed]
- Litsanov, B.; Brocker, M.; Bott, M. Toward homosuccinate fermentation: Metabolic engineering of Corynebacterium glutamicum for anaerobic production of succinate from glucose and formate. Appl. Environ. Microbiol. 2012, 78, 3325–3337. [Google Scholar] [CrossRef] [PubMed]
- Okino, S.; Noburyu, R.; Suda, M.; Jojima, T.; Inui, M.; Yukawa, H. An efficient succinic acid production process in a metabolically engineered Corynebacterium glutamicum strain. Appl. Microbiol. Biotechnol. 2008, 81, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, H.; Cai, H.; Zhou, Z.; Chen, Y.; Ouyang, P. Succinic acid production from corn cob hydrolysates by genetically engineered Corynebacterium glutamicum. Appl. Biochem. Biotechnol. 2014, 172, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Hirasawa, T.; Nishii, M.; Furusawa, C.; Shimizu, H. Enhanced acetic acid and succinic acid production under microaerobic conditions by Corynebacterium glutamicum harboring Escherichia coli transhydrogenase gene pntAB. J. Gen. Appl. Microbiol. 2014, 60, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Xia, H.; Yang, J.; Zhao, X.; Chen, T. Improved succinate production in Corynebacterium glutamicum by engineering glyoxylate pathway and succinate export system. Biotechnol. Lett. 2014, 36, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Murakami, S.; Okino, S.; Kawaguchi, H.; Vertès, A.A.; Yukawa, H. Metabolic analysis of Corynebacterium glutamicum during lactate and succinate productions under oxygen deprivation conditions. J. Mol. Microbiol. Biotechnol. 2004, 7, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Okino, S.; Inui, M.; Yukawa, H. Production of organic acids by Corynebacterium glutamicum under oxygen deprivation. Appl. Microbiol. Biotechnol. 2005, 68, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Effects of growth mode and pyruvate carboxylase on succinic acid production by metabolically engineered strains of Escherichia coli. Appl. Environ. Microbiol. 2002, 68, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Sarks, C.; Jin, M.; Sato, T.K.; Balan, V.; Dale, B.E. Studying the rapid bioconversion of lignocellulosic sugars into ethanol using high cell density fermentations with cell recycle. Biotechnol. Biofuels 2014, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Yukawa, H.; Omumasaba, C.A.; Nonaka, H.; Kos, P.; Okai, N.; Suzuki, N.; Suda, M.; Tsuge, Y.; Watanabe, J.; Ikeda, Y.; et al. Comparative analysis of the Corynebacterium glutamicum group and complete genome sequence of strain R. Microbiology 2007, 153, 1042–1058. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Okayama, S.; Nonaka, H.; Tsuge, Y.; Inui, M.; Yukawa, H. Large-scale engineering of the Corynebacterium glutamicum genome. Appl. Environ. Microbiol. 2005, 71, 3369–3372. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Nonaka, H.; Tsuge, Y.; Inui, M.; Yukawa, H. New multiple-deletion method for the Corynebacterium glutamicum genome, using a mutant lox sequence. Appl. Environ. Microbiol. 2005, 71, 8472–8480. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Inui, M.; Yukawa, H. Site-directed integration system using a combination of mutant lox sites for Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2007, 77, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Pátek, M.; Nešvera, J. Promoters and Plasmid Vectors of Corynebacterium glutamicum. In Corynebacterium glutamicum; Yukawa, H., Inui, M., Eds.; Springer: Heidelberg, Germany, 2013; Volume 23, pp. 51–88. [Google Scholar]
- Gande, R.; Dover, L.G.; Krumbach, K.; Besra, G.S.; Sahm, H.; Oikawa, T.; Eggeling, L. The two carboxylases of Corynebacterium glutamicum essential for fatty acid and mycolic acid synthesis. J. Bacteriol. 2007, 189, 5257–5264. [Google Scholar] [CrossRef] [PubMed]
- Peters-Wendisch, P.G.; Wendisch, V.F.; Paul, S.; Eikmanns, B.J.; Sahm, H. Pyruvate carboxylase as an anaplerotic enzyme in Corynebacterium glutamicum. Microbiology 1997, 143, 1095–1103. [Google Scholar] [CrossRef]
- Kim, M.I.; Kim, N.J.; Shang, L.; Chang, Y.K.; Lee, S.Y.; Chang, H.N. Continuous production of succinic acid using an external membrane cell recycle system. J. Microbiol. Biotechnol. 2009, 19, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).