

Prokaryote Composition and Structure of Rumen Fluid before and after In Vitro Rumen Fermentation

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Background
2. Materials and Methods
2.1. In Vitro Fermentation
2.2. DNA Extraction
2.3. Bioinformatics
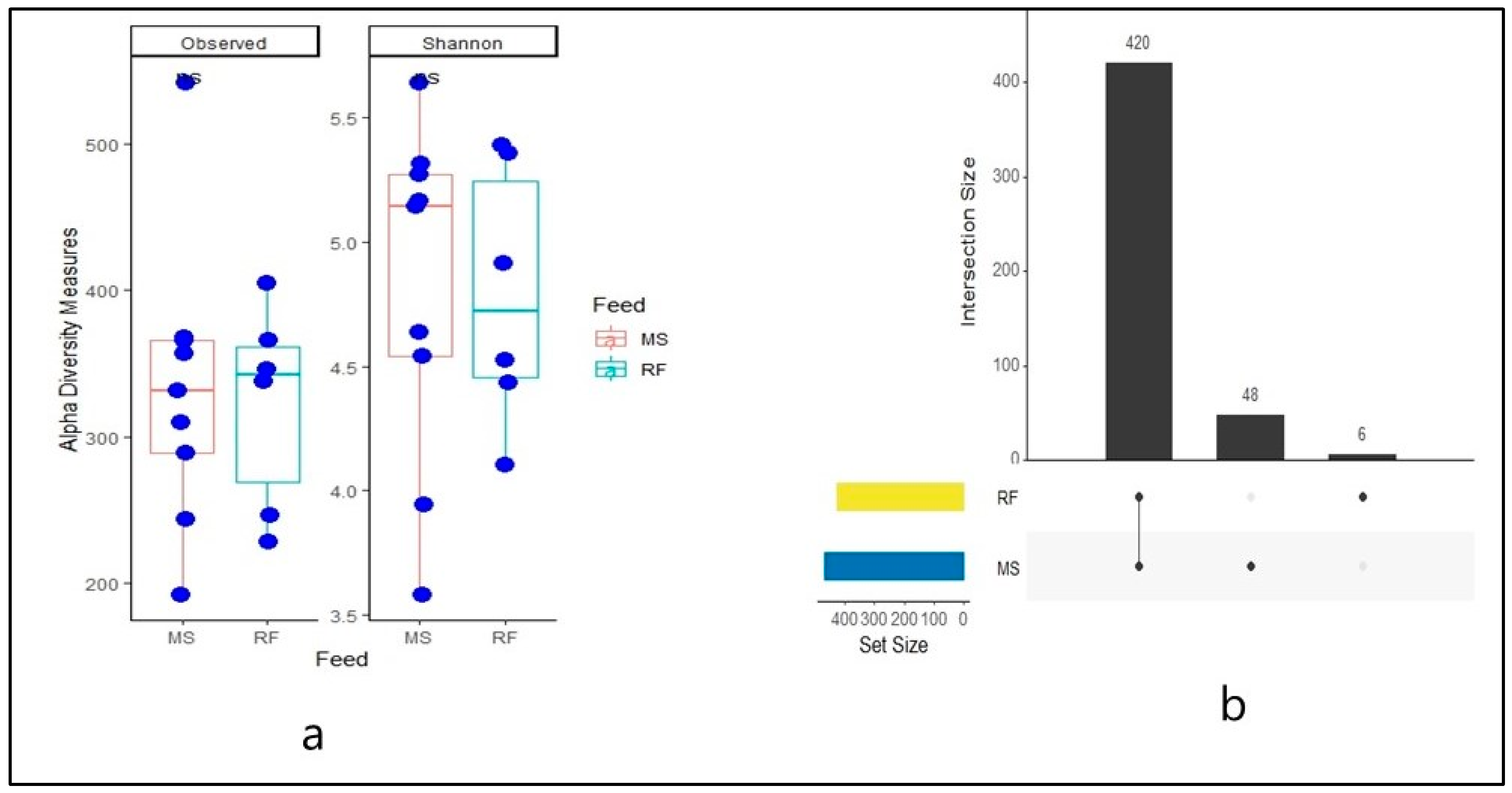
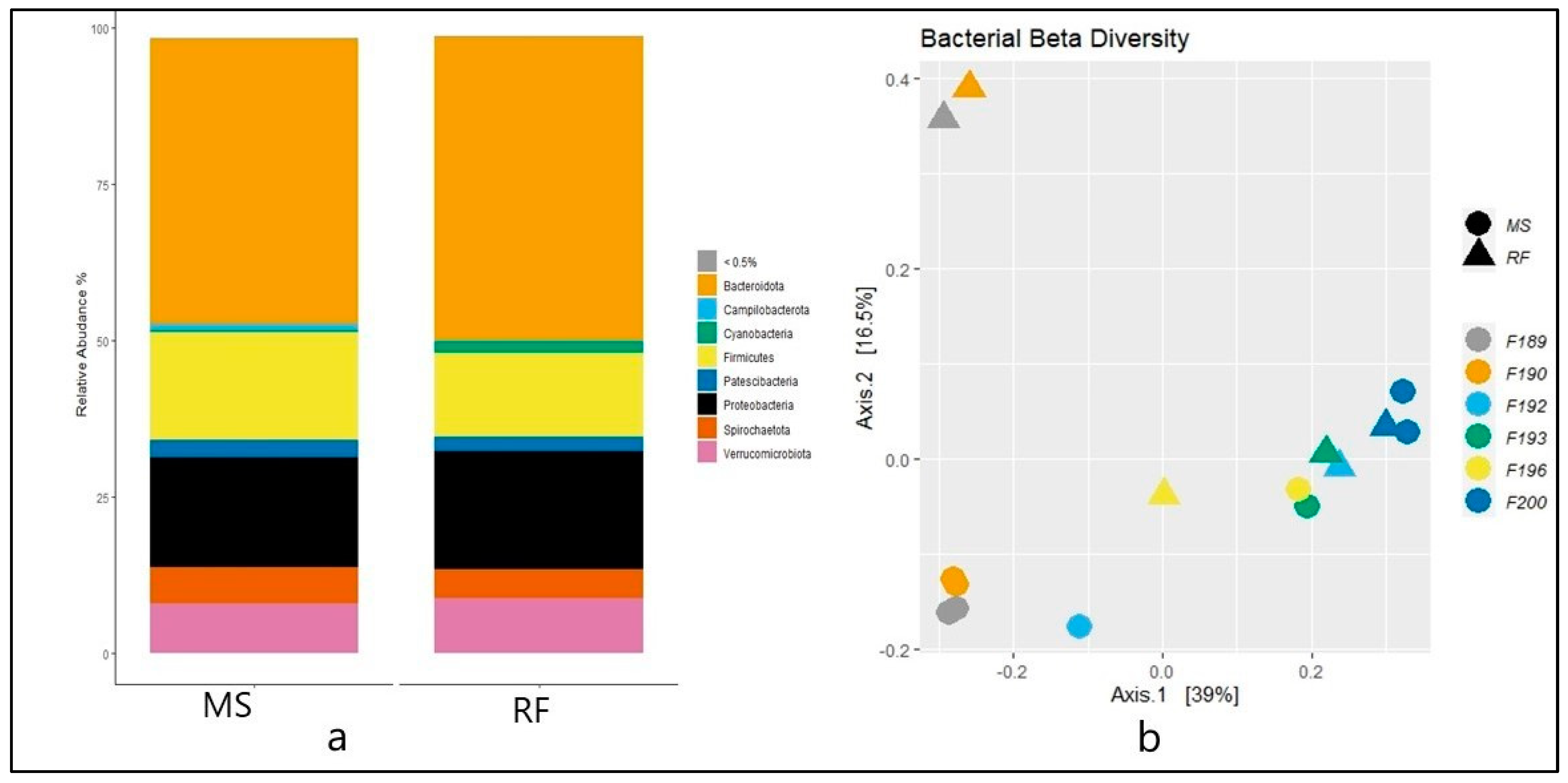
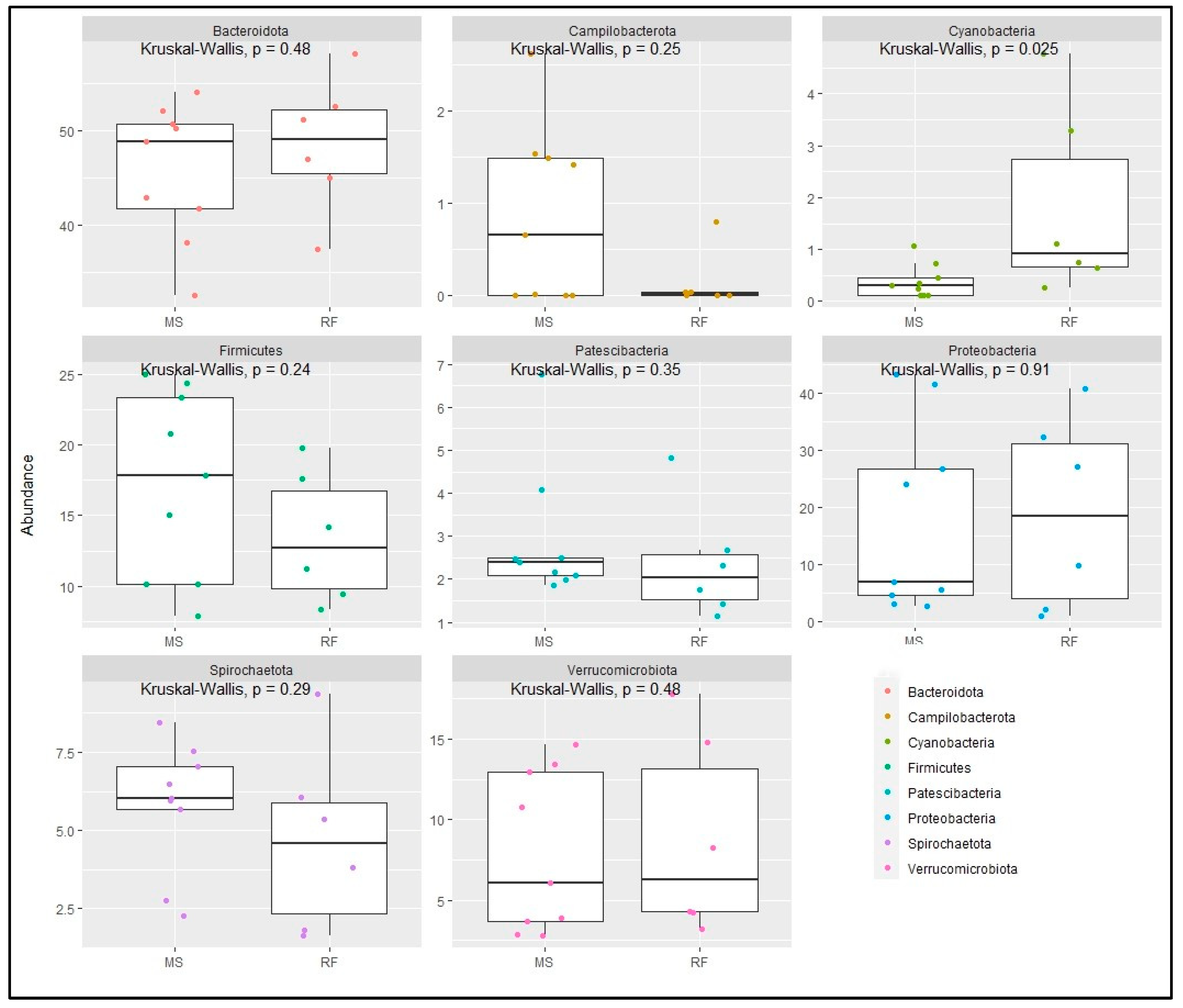
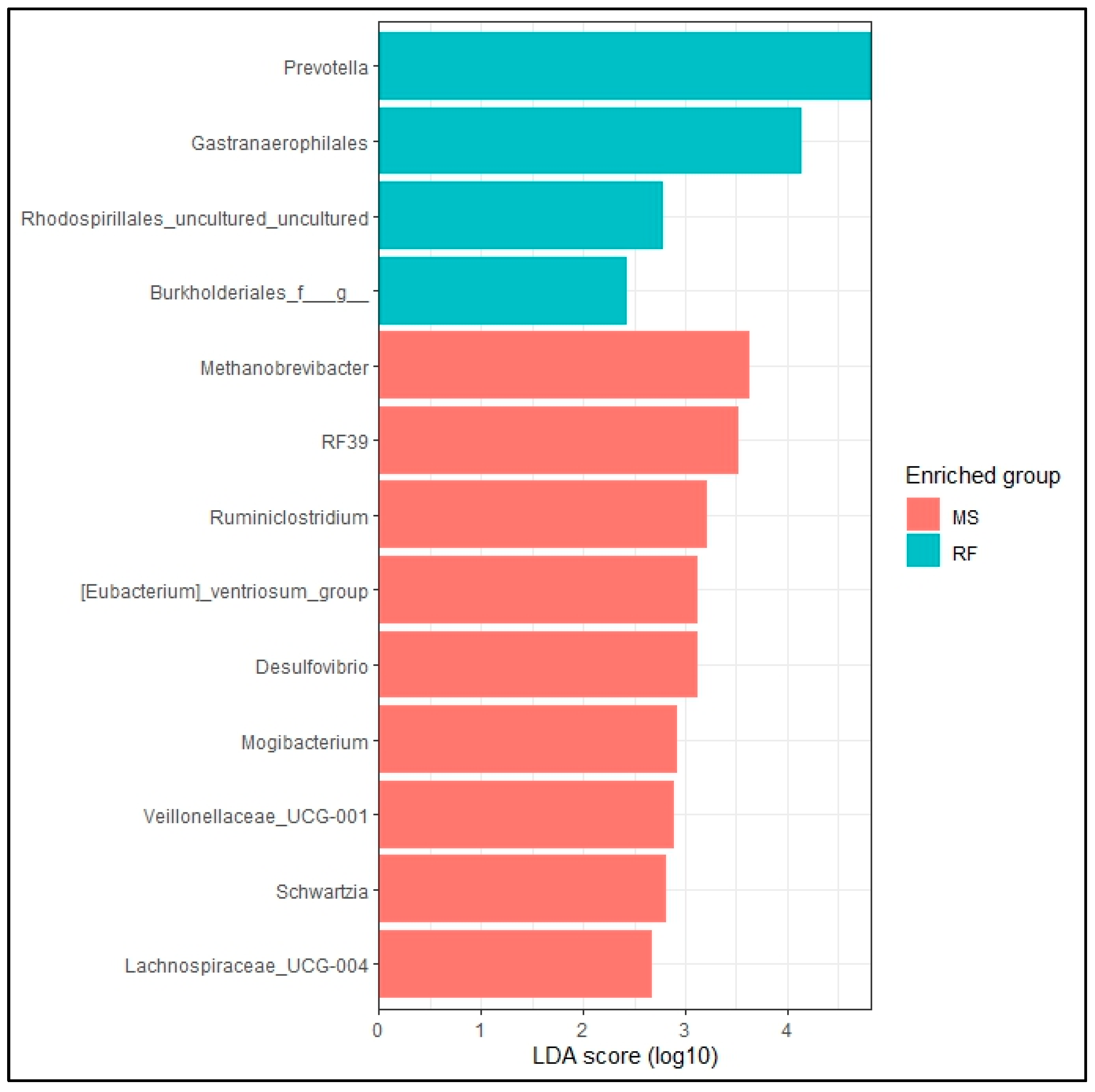
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASV | Amplicon sequence variant |
| DADA2 | Deficiency of adenosine deaminase 2 |
| DNA | Deoxyribonucleic acid |
| IVRF | In vitro rumen fermentation |
| MS | Maize silage |
| PCoA | Principle coordinate analysis |
| PCR | Polymerase chain reaction |
| QIIME2 | Quantitative insights into microbial ecology 2 |
| RF | Rumen fluid |
| rRNA | Ribosomal ribonucleic acid |
| VFAs | Volatile fatty acids |
References
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Applications of the In Vitro Gas Method in the evaluation of feed resources, and enhancement of nutritional value of tannin-rich tree/browse leaves and agro-industrial by-products. Anim. Prod. Heal. Sect. Int. At. Energy Agency Vienna 2009, 2, 23–41. [Google Scholar]
- Menke, K.H.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef]
- Blümmel, M.; Makkar, H.P.S.; Becker, K. In vitro gas production: A technique revisited. J. Anim. Physiol. Anim. Nutr. 1997, 77, 24–34. [Google Scholar] [CrossRef]
- Cornou, C.; Storm, I.M.L.D.; Hindrichsen, I.K.; Worgan, H.; Bakewell, E.; Ruiz, D.R.Y.; Abecia, L.; Tagliapietra, F.; Cattani, M.; Ritz, C.; et al. A ring test of a wireless in vitro gas production system. Anim. Prod. Sci. 2013, 53, 585–592. [Google Scholar] [CrossRef]
- Dijkstra, J.; Kebreab, E.; Bannink, A.; France, J.; López, S. Application of the gas production technique to feed evaluation systems for ruminants. Anim. Feed Sci. Technol. 2005, 123–124 Pt 1, 561–578. [Google Scholar] [CrossRef]
- Pandey, D.; Hansen, H.H.; Dhakal, R.; Aryal, N.; Rai, S.P.; Sapkota, R.; Nielsen, M.O.; Novoa-Garrido, M.; Khanal, P. Interspecies and seasonal variations in macroalgae from the Nordic region: Chemical composition and impacts on rumen fermentation and microbiome assembly. J. Clean. Prod. 2022, 363, 132456. [Google Scholar] [CrossRef]
- Sembach, L.E.; Hansen, H.H.; Dhakal, R.; Eriksson, T.; Nielsen, N.I.; Nielsen, M.O. Estimates for rumen dry matter degradation of concentrates are higher, but not consistently, when evaluated based on in sacco as compared to in vitro methods. In Proceedings of the 10th Nordic Feed Science Conference, Uppsala, Sweden, 11–12 June 2019; pp. 123–128. [Google Scholar]
- Getachew, G.; Robinson, P.H.; DePeters, E.J.; Taylor, S.J. Relationships between chemical composition, dry matter degradation and in vitro gas production of several ruminant feeds. Anim. Feed Sci. Technol. 2004, 111, 57–71. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Pedersen, N.C.; Khushvakov, J.; Ye, Y.; Dhakal, R.; Hansen, H.H.; Ahrné, L.; Khakimov, B. Effect of Supplementing Dairy Goat Diets with Rapeseed Oil or Sunflower Oil on Performance, Milk Composition, Milk Fatty Acid Profile, and in vitro Fermentation Kinetics. Front. Vet. Sci. 2022, 9, 614. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, R.; Ronquillo, M.G.; Vargas-Bello-Pérez, E.; Hansen, H.H. Effect of Autochthonous Nepalese Fruits on Nutrient Degradation, Fermentation Kinetics, Total Gas Production, and Methane Production in In-Vitro Rumen Fermentation. Animals 2022, 12, 2199. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- R Core Team. R A Lang. Environ. Stat. Comput. R Found. Stat. Comput. 2022. Available online: https://www.r-project.org/ (accessed on 29 December 2023).
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; Hara, R.B.O.; Simpson, G.L.; Solymos, P.; et al. Package ‘vegan’. 2020. Available online: https://github.com/vegandevs/vegan (accessed on 3 February 2023).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Cao, Y.; Dong, Q.; Wang, D.; Zhang, P.; Liu, Y.; Niu, C. microbiomeMarker: An R/Bioconductor package for microbiome marker identification and visualization. Bioinformatics 2022, 38, 4027–4029. [Google Scholar] [CrossRef]
- Wei, X.; Ouyang, K.; Long, T.; Liu, Z.; Li, Y.; Qiu, Q. Dynamic Variations in Rumen Fermentation Characteristics and Bacterial Community Composition during In Vitro Fermentation. Fermentation 2022, 8, 5–7. [Google Scholar] [CrossRef]
- Terry, S.A.; Badhan, A.; Wang, Y.; Chaves, A.V.; McAllister, T.A. Fibre digestion by rumen microbiota—A review of recent metagenomic and metatranscriptomic studies. Can. J. Anim. Sci. 2019, 99, 678–692. [Google Scholar] [CrossRef]
- Belanche, A.; Patra, A.K.; Morgavi, D.P.; Suen, G.; Newbold, C.J.; Yáñez-Ruiz, D.R. Editorial: Gut Microbiome Modulation in Ruminants: Enhancing Advantages and Minimizing Drawbacks. Front. Microbiol. 2021, 11, 622002. [Google Scholar] [CrossRef]
- Mizrahi, I.; Wallace, R.J.; Moraïs, S. The rumen microbiome: Balancing food security and environmental impacts. Nat. Rev. Microbiol. 2021, 19, 553–566. [Google Scholar] [CrossRef]
- Macome, F.M.; Pellikaan, W.F.; Hendriks, W.H.; Warner, D.; Schonewille, J.T.; Cone, J.W. In vitro gas and methane production in rumen fluid from dairy cows fed grass silages differing in plant maturity, compared to in vivo data. J. Anim. Physiol. Anim. Nutr. 2018, 102, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Fortina, R.; Glorio Patrucco, S.; Barbera, S.; Tassone, S. Rumen Fluid from Slaughtered Animals: A Standardized Procedure for Sampling, Storage and Use in Digestibility Trials. Methods Protoc. 2022, 5, 59. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.Y.; Zhou, J.W.; Yi, S.Y.; Wang, M.; Tan, Z.L. In vitro Inoculation of Fresh or Frozen Rumen Fluid Distinguishes Contrasting Microbial Communities and Fermentation Induced by Increasing Forage to Concentrate Ratio. Front. Nutr. 2022, 8, 772645. [Google Scholar] [CrossRef] [PubMed]
- Spanghero, M.; Chiaravalli, M.; Colombini, S.; Fabro, C.; Froldi, F.; Mason, F.; Moschini, M.; Sarnataro, C.; Schiavon, S.; Tagliapietra, F. Rumen inoculum collected from cows at slaughter or from a continuous fermenter and preserved in warm, refrigerated, chilled or freeze-dried environments for in vitro tests. Animals 2019, 9, 815. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H.; Abecia, L.; Angarita, E.; Aravena, P.; Arenas, G.N.; et al. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [PubMed]
- Ilina, L.A.; Filippova, V.A.; Brazhnik, E.A.; Dubrovin, A.V.; Yildirim, E.A.; Dunyashev, T.P.; Laptev, G.Y.; Novikova, N.I.; Sobolev, D.V.; Yuzhakov, A.A.; et al. The comparative analysis of the ruminal bacterial population in reindeer (Rangifer tarandus L.) from the Russian arctic zone: Regional and seasonal effects. Animals 2021, 11, 911. [Google Scholar] [CrossRef]
- Nandi, R.; Sengupta, S. Microbial Production of Hydrogen: An Overview. Crit. Rev. Microbiol. 1998, 24, 61–84. [Google Scholar] [CrossRef]
- Neves, A.L.A.; Li, F.; Ghoshal, B.; McAllister, T.; Guan, L.L. Enhancing the resolution of rumen microbial classification from metatranscriptomic data using Kraken and Mothur. Front. Microbiol. 2017, 8, 2445. [Google Scholar] [CrossRef]
- Amin, N.; Seifert, J. Dynamic progression of the calf’s microbiome and its influence on host health. Comput. Struct. Biotechnol. J. 2021, 19, 989–1001. [Google Scholar] [CrossRef]
- Tröscher-Mußotter, J.; Saenz, J.S.; Grindler, S.; Meyer, J.; Kononov, S.U.; Mezger, B.; Borda-Molina, D.; Frahm, J.; Dänicke, S.; Camarinha-Silva, A.; et al. Microbiome Clusters Disclose Physiologic Variances in Dairy Cows Challenged by Calving and Lipopolysaccharides. mSystems 2021, 6, e00856-21. [Google Scholar] [CrossRef]
- Kim, S.H.; Mamuad, L.L.; Kim, E.J.; Sung, H.G.; Bae, G.S.; Cho, K.K.; Lee, C.; Lee, S.S. Effect of different concentrate diet levels on rumen fluid inoculum used for determination of in vitro rumen fermentation, methane concentration, and methanogen abundance and diversity. Ital. J. Anim. Sci. 2018, 17, 359–367. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhakal, R.; Neves, A.L.A.; Sapkota, R.; Khanal, P.; Hansen, H.H. Prokaryote Composition and Structure of Rumen Fluid before and after In Vitro Rumen Fermentation. Fermentation 2024, 10, 108. https://doi.org/10.3390/fermentation10020108
Dhakal R, Neves ALA, Sapkota R, Khanal P, Hansen HH. Prokaryote Composition and Structure of Rumen Fluid before and after In Vitro Rumen Fermentation. Fermentation. 2024; 10(2):108. https://doi.org/10.3390/fermentation10020108
Chicago/Turabian StyleDhakal, Rajan, André Luis Alves Neves, Rumakanta Sapkota, Prabhat Khanal, and Hanne Helene Hansen. 2024. "Prokaryote Composition and Structure of Rumen Fluid before and after In Vitro Rumen Fermentation" Fermentation 10, no. 2: 108. https://doi.org/10.3390/fermentation10020108
APA StyleDhakal, R., Neves, A. L. A., Sapkota, R., Khanal, P., & Hansen, H. H. (2024). Prokaryote Composition and Structure of Rumen Fluid before and after In Vitro Rumen Fermentation. Fermentation, 10(2), 108. https://doi.org/10.3390/fermentation10020108