Abstract
Long non-coding RNAs (lncRNAs) belong to a class of non-protein-coding RNAs with their lengths longer than 200 nucleotides. Most of the mammalian genome is transcribed as RNA, yet only a small percent of the transcribed RNA corresponds to exons of protein-coding genes. Thus, the number of lncRNAs is predicted to be several times higher than that of protein-coding genes. Because of sheer number of lncRNAs, it is often difficult to elucidate the functions of all lncRNAs, especially those arising from their relationship to their binding partners, such as DNA, RNA, and proteins. Due to their binding to other macromolecules, it has become evident that the structures of lncRNAs influence their functions. In this regard, the recent development of epitranscriptomics (the field of study to investigate RNA modifications) has become important to further elucidate the structures and functions of lncRNAs. In this review, the current status of lncRNA structures and functions influenced by epitranscriptomic marks is discussed.
1. Introduction
By definition, long non-coding RNAs (lncRNAs) are any ncRNAs that are longer than 200 nucleotides (nt). With the advancement of high-throughput techniques [microarrays, next generation sequencing (NGS), especially RNA sequencing (RNA-seq)], many lncRNAs have been discovered [1]. To date, a number of functions of lncRNAs have been proposed and experimentally validated; ranging from decoy, epigenetic, transcriptional, post-transcriptional, and translational controls [2,3,4,5]. The general understanding in the field is that lncRNAs exert their actions by binding to other macromolecules, which are DNA, RNA, and proteins [6,7]. Thus, it is essential to identify the potential binding partners to elucidate the mechanism of action of lncRNAs. To this end, the most popular method is using an affinity tag on an in vitro purified RNA and using this RNA as a bait to pull-down proteins/nucleic acids from cellular extracts. There are other more elaborated methods currently available, including ChIRP (Chromatin isolation by RNA purification), CHART (Capture Hybridization Analysis of RNA Targets), CLIP (cross-linking and immunoprecipitation), and RAP (RNA antisense purification), which are comprehensively reviewed elsewhere [8,9,10].
Just as DNA and proteins, RNA can be modified by a variety of enzymes. The classic example is the RNA modifications of ribosomal RNAs (rRNAs) and transfer RNAs (tRNAs), which affect the efficiency of translation [11,12,13]. To date, there are more than 170 RNA modifications known across species [14], which has opened up a new field of study called, epitranscriptomics [15,16], whose name is based on the well-studied field of DNA modification, epigenetics. Much of the concepts of epigenetics are applied to dissect the ever-growing field of epitranscriptomics, including the epitranscriptomic enzymes being categorized as writers, readers, and erasers. Among epitranscriptomic marks, the most well studied one in recent years is N6-methyladenosine (m6A), which is a methylation of nitrogen-6 position of adenosine (A) found in messenger RNAs (mRNAs) and non-protein-coding RNAs (ncRNAs). Other epitranscriptomics marks in mammals include the A-to-I RNA editing, 2′-O-methylation (2′-O-Me), N1-methyladenosine (m1A), 3-methylcytidine (m3C), 5-methylcytosine (m5C), N7-methylguanosine (m7G), pseudouridylation (Ψ) to name a few [17,18]. These epitranscriptomic marks affect all realms of RNA lifecycle, including splicing, subcellular localization, microRNA (miRNA) biogenesis and bindings, RNA stability, and translation efficiency [19,20]. More importantly, dysregulation of epitranscriptomic marks affect many diseases, including cardiovascular [21], liver [22], and neurodegenerative diseases [18] as well as cancers [23]. These epitranscriptomic marks are also found in lncRNAs [24]. Given that epitranscriptomic marks affect the binding between lncRNA and other macromolecules is still a matter of ongoing investigation, which this review aims to summarize.
2. Epitranscriptomic Marks Affect RNA Structures as in the Case of Immune Responses
RNA exists in a single-stranded (ssRNA) or double-stranded RNA (dsRNA) state. The balance between these states may be influenced by cellular conditions, such as stress and viral infection [25,26,27,28]. Furthermore, more than half of the human genome consists of repetitive sequences, such as those derived from transposons and ALU elements [29]. These repetitive sequences form palindromic repeats, resulting in the formation of dsRNAs [30]. To detect dsRNAs, there are several high-throughput methods available, including PARS (Parallel Analysis of RNA Structure) by sequencing RNA digested with RNases S1 and V1 that specifically recognize single-stranded RNA (ssRNAs) and dsRNAs, respectively [31]. Other methods to analyze RNA structures are DMS-Seq to label RNA structures by dimethyl sulfate (DMS) [32], LIGR-seq (LIGation of interacting RNA followed by high-throughput sequencing) to globally map RNA–RNA duplexes crosslinked in vivo [33], PARIS to detect dsRNA [34], RIC-seq (RNA in situ conformation sequencing) to globally profile intra- and intermolecular RNA–RNA interactions [35,36], SHAPE-Seq (selective 2′-hydroxyl acylation analyzed by primer extension sequencing) [37], and SHAPE-MaP (selective 2′-hydroxyl acylation analyzed by primer extension and mutational profiling) to chemically probe RNA by adding RNA-specific small molecules in cell culture [38,39]. Recently, a comprehensive RNA structure probing database, RASP, was released, which contains 18 species (e.g., animals, plants, bacteria, fungi, and viruses) and 18 different experimental methods measuring RNA secondary structures in a transcriptome-wide manner [40]. Furthermore, there are databases for epitranscriptomic marks (comprehensively reviewed in [41]), including RMBase v2.0 [42] and RMVar [43] that contain several epitranscriptomic marks for different organisms. It will be of great interest to further analyze the collected data sets by merging them with high-throughput data that map known epitranscriptomic marks. This will enable the analysis of the preferential distribution of each epitranscriptomic mark to ssRNAs and dsRNAs in different species (thus, evolutional-conservation, if any).
Upon viral infection, the innate immune system is triggered, which recognizes pathogen-associated molecular patterns (PAMPs, which are unique molecular ligands on or within microbes, including viral DNA and RNA) leading to activation of intracellular signaling pathways to initiate antiviral response [44,45]. These PAMPs are detected by the host through pattern recognition receptors, such as Nod-like receptors (NLRs), RIG-I-like receptors (RLRs), and Toll-like receptors (TLRs) [46]. In the case of RLRs, RIG-I senses short dsRNAs, while the RLR, MDA5 (melanoma differentiation-associated protein 5), detects long dsRNAs. These recognitions of PAMPs by RLRs are followed by MAVS (mitochondrial antiviral-signaling protein)-mediated activation of signaling cascades, including type I interferon responses [47,48,49]. The epitranscriptomic mark, m6A, plays active roles in innate immunity by reducing type I interferon production [50,51]. Winkler et al. reported that m6A marks deposited by the m6A METTL3 and read by the m6A reader YTHDF2 negatively regulate interferon response by facilitating the fast turnover of interferon mRNAs leading to viral propagation [50] (Figure 1A). Interestingly, increasing evidence suggests that lncRNAs are shown to be involved in virus infections and antiviral immune responses [52]. Furthermore, many lncRNAs have m6A marks [53,54,55], influencing secondary structures of lncRNAs. For example, MALAT1 (metastasis associated lung adenocarcinoma transcript 1) is involved in inflammatory responses and innate immunity [56,57,58] along with its enzymatic processing product, MALAT1-associated small cytoplasmic RNA (mascRNA) [59,60,61,62,63]. These findings highlight that further investigation of epitranscriptomic marks on lncRNAs and their secondary structural changes may reveal the active involvement of lncRNAs in innate immunity. In this regard, it will be of high interest to understand the relationship between lncRNAs and another epitranscriptomic mark, pseudouridylation (Ψ) [64], as it is demonstrated recently in COVID-19 mRNA vaccines using N1-methylpseudouridine (m1Ψ) to increase their effectiveness [65].
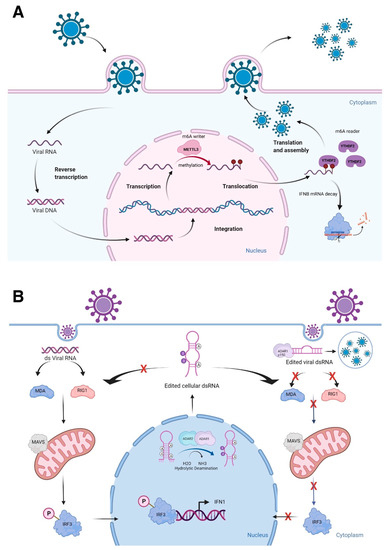
Figure 1.
Epitranscriptomic marks and RNA structures in immune responses. (A) Viral RNA methylation deposited by the m6A writer METTL3 and read by the m6A reader YTHDF2 negatively regulate cellular defense response by facilitating the fast turnover of interferon mRNA leading to viral replication. (B) The role of ADARs in differentiating self-from non-self dsRNA by modulating canonical antiviral pathways induced by dsRNA. During an infection, the viral dsRNA enters into the cytoplasm. Non-edited dsRNA binds to MDA5 (melanoma differentiation-associated protein 5) and RIG-I (retinoic acid-inducible gene I like receptor). This complex activates MAVS (mitochondrial antiviral-signaling protein), leading to the phosphorylation of IRF3 (interferon regulatory transcription factor 3) and its translocation into the nucleus, thus inducing a type 1 interferon response. Endogenous cellular dsRNA that is generated during transcription is A-to-I edited by ADARs. The ADAR1 isoform p150 is cytoplasmic and is induced by interferon. It edits dsRNA either of viral or cellular origin. This dsRNA contains inosine and inhibits the activation of MDA5 and RIG-1, thus turning off the interferon response and apoptosis to prevent autoimmune reaction. However, this mechanism could favor virus replication, if it is not tightly regulated. Figure is created with BioRender.com, accessed on 15 March 2022.
A-to-I RNA editing is a type of epitranscriptomic mark that involves the RNA editing enzymes, ADARs [adenosine deaminases acting on RNA, consisting of three genes: ADAR1, ADARB1 (ADAR2), and catalytically inactive ADARB2 (ADAR3)], recognize dsRNAs to catalyze adenosine to inosine (A-to-I) conversion, mostly at ALU repeats and introns [21]. ALU repeats are ~300 bp that belong to the family of repetitive elements in primates. There are more than one million ALU repeats in primate genomes [66]. Two transcribed ALU repeats form a quasi-palindrome, which becomes double-stranded RNA to recruit ADARs to catalyze A-to-I RNA editing [67]. I is recognized as guanosine (G) by splicing and translational machineries as well as in reverse transcription reactions; allowing detection of A-to-G changes in RNA-seq reads when these reads are mapped to the reference genome [68]. Mutations in the human ADAR1 gene result in the autoimmune disease, Aicardi-Goutières syndrome, while the whole-body knockout mice of Adar1 results in embryonic death due to massive apoptosis and aberrant interferon induction, which can be rescued to live birth by ablating the RLRs, Mavs or Mda5 (melanoma differentiation-associated protein 5) [69,70,71]. Both ADAR1 and ADAR2 are important in differentiating self- from non-self dsRNAs [70,72,73] (Figure 1B). Furthermore, silencing of ADAR1 in the human hepatocellular carcinoma cell line, HepG2, resulted in shifting of dsRNAs to ssRNAs at the transcriptome-wide level [74]. As many lncRNAs have A-to-I RNA editing sites [75,76], further characterization of RNA editing sites will uncover the secondary structures of lncRNAs, especially the conversion of A to I at the nitrogen-6 position of adenosine, which can be methylated as m6A, if not edited [77]. Thus, both epitranscriptomic marks, A-to-I RNA editing and m6A, could competitively affect the secondary structures of lncRNAs, thereby, influencing the binding of other macromolecules.
3. Understanding the Actions of RNA-Binding Proteins Is Important for Functions of lncRNAs
RNA-binding proteins (RBPs) bind ssRNAs or dsRNAs to modulate their stability and translation [78]. RBPs also bind lncRNAs as in the case of the lncRNA NORAD sequestering the RBP, PUMILIO, to regulate genomic stability [79,80,81]. Classically, RBPs are defined to have specific motifs called, RNA-binding domains (RBDs). Through proteome-wide screenings (e.g., RNA interactome capture), over 2000 RBPs have been identified, of which many lack known RBDs [82,83,84]. In regards to lncRNAs, the catalytic subunit of polycomb repressor complex 2, EZH2, is a good example of a protein binding to RNA, including lncRNAs, without known RBDs, although promiscuously [85,86]. Indeed, there are databases available to examine the binding of RBPs to lncRNAs, including CLIPdb [87], POSTAR2 [88], and starBase [89]. Thus, increasing studies now investigate the possible binding of lncRNAs to RBPs thereby regulating RNA metabolism and functions (possibly as RBP sponges to sequester the available RBPs in the cell), instead of RBPs merely functioning in the biogenesis of lncRNAs [90].
RBPs can either stabilize or degrade the bound RNA [91] (Figure 2). Given that mRNAs have different epitranscriptomic marks, whether such marks signal the recruitment of RBPs to the mRNAs that ultimately affect the mRNA stability needs further investigation. A recent systematic analysis of RBP-bound regions and A-to-I RNA editing sites suggest that such RNA-edited sites are preferentially bound by the RNA-stabilizing RBP, HuR (encoded by ELAVL1 gene) [92]. This study further confirms the possible functional role of RNA-edited sites of individual genes in regards to mRNA stability via the action of HuR [93,94]. Furthermore, several lncRNAs are found to bind HuR, including the lncRNA, LINC02381, that stabilizes the 3′-untranslated region (UTR) of the HOXC10 mRNA via HuR [95]. Interestingly, although there is no A-to-I RNA editing (based on the DARNED database [96,97], https://darned.ucc.ie, accessed on 2 February 2022), nor m6A site (based on the m6A-Atlas database [98], http://180.208.58.19/m6A-Atlas/, accessed on 2 February 2022) in the 3′-UTR of HOXC10 gene, an 5-methylcytosine (m5C) site (based on the m5C-Atlas database [99], http://180.208.58.19/m5c-atlas/, accessed on 2 February 2022) is present in the 3′-UTR of HOXC10 gene, which calls for further investigation of this region to understand the causal relationship between lncRNA, RBP, and epitranscriptomic marks.
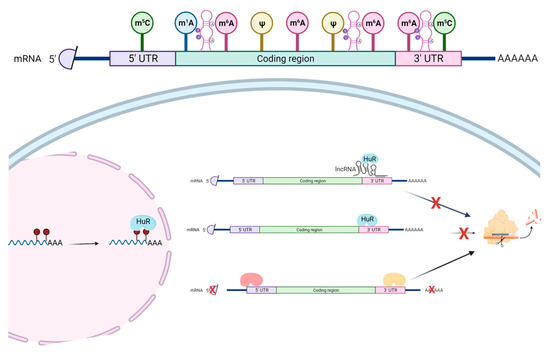
Figure 2.
The roles of RBPs in the epitranscriptomic context. The most common epitranscriptomic marks include 5-methylcytosine (m5C), N1-methyladenosine (m1A), N6-methyladenosine (m6A), pseudouridine (Ψ), as well as A-I RNA editing. These marks affect the structures of RNA and, thus, influence the binding of RBPs. RNA-edited sites are preferentially bound by HuR—a RNA-stabilizing RBP. Furthermore, several lncRNAs, including LINC02381, are found to be bound by HuR, which stabilizes the 3′-UTR of the mRNA. Other RBPs could promote RNA degradation. Figure is created with BioRender.com, accessed on 16 March 2022.
Given that many epitranscriptomic readers are RBPs (e.g., m6A readers: YTHDF1-3, YTHDC1, YTHDC2, HNRNPC, RBMX (HNRNPG), IGF2BP1-3 [100,101]), it is not surprising that the RBP binding sites overlap that of epitranscriptomic marks on mRNAs to regulate mRNA stability. In the case of RBP binding to the m6A sites, a convenient tool called, m6Adecom, was introduced recently [102]. However, it is of utmost importance to experimentally validate the relationships among epitranscriptomic marks, RBPs, and lncRNAs by performing gain/loss-of-function experiments for each of the components to understand the causal relation between them, rather than simply performing and analyzing high-throughput data.
4. Factors Influencing lncRNA Mediated R-Loop Formation—Sequence, Structure and Chemical Marks
R-loops (RNA loops) are three-stranded nucleic acid structures formed from the hybridization of RNA and DNA, leading to a displaced single-stranded DNA (ssDNA). These structures typically result from nascent transcription at regions of high GC-skew, such as CpG islands in gene promoters [103]. When improperly regulated, R-loops can be pathological [104,105,106]. A well-described consequence of pathological R-loops is collision of the transcriptional machinery with the replisome (transcription-replication conflict) [107]. Not surprisingly, because of these deleterious consequences, most interest has focused on investigating factors involved in R-loop processing or dissolution, such as the RNase H1 and RNase H2 enzymes that specifically degrade the RNA in R-loops [108]. However, under basal conditions, R-loops are benign; and, in some cases, they play important physiological roles [109,110]. Recent studies also indicate that lncRNAs are involved in the formation of R loops, such as lncRNAs APOLO [111], HOTTIP [112], and TERRA [113,114,115,116,117,118,119], and enhancer RNAs (a subclass of lncRNAs arising from the enhancer regions of protein-coding genes) [120] (Figure 3).
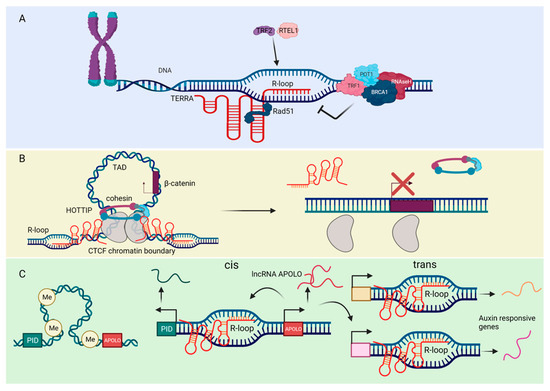
Figure 3.
The roles of ncRNAs in the formation of R-loops. (A) The lncRNA TERRA binds to telomeres through direct base-pairing with telomeric DNA, forming the R-loop structure. The RAD51 DNA recombinase binds TERRA and thus catalyzes TERRA R-loop formation. The helicase RTEL1 and the telomeric shelterin component TRF2 also promote TERRA association with chromosome ends and stimulate R-loop formation. TRF1 and POT1 play a role in preventing the accumulation of such structures. The DNA recombination factor BRCA1 modulate TERRA binding to telomeres, preventing R-loop-associated telomeric DNA damage. TERRA R-loops are regulated by endonucleolytic cleavage activity of RNase H, which removes the RNA-DNA hybrids. (B) The lncRNA HOTTIP is highly expressed in acute myeloid leukemia. HOTTIP directly interacts with and regulates CTCF/cohesin complex and form R-loops. HOTTIP-mediated R-loops facilitate formation of topologically associating domain (TAD) to drive gene transcription of β-catenin. Eliminating HOTTIP-guided R-loops by targeting RNase H or removing CTCF-binding sites affect TAD structure, leading to alleviating leukemia severity. (C) The cis and trans regulatory mechanisms of the lncRNA APOLO. Under normal physiological conditions, APOLO is not transcribed due to hypermethylation of its promoter and chromosomal looping. However, during the lateral root development, the plant hormone Auxin induces the transcription of APOLO. The transcribed APOLO forms R-loops and thus modulates the chromatin looping in cis and trans, thereby enhancing transcription of auxin responsive genes and lateral root development. The figure was created with BioRender.com, accessed on 17 March 2022.
A critical aspect of R-loop biology that may impact functionality and dynamics are characteristics of the RNA moiety in these structures. There is emerging evidence that m6A methylation [121,122,123] plays a key role in regulating R-loop dynamics. In a recent study, Yang et al. showed that the m6A writer, METTL3, is required for R-loop formation at m6A+ gene termination sites to prevent transcript read-through [123]. These results were extended in a subsequent study by Abakir et al., which found that m6A readers like YTHDF2 regulate R-loop levels to prevent genome instability in dividing cells [122]. Curiously, Kang et al. found in their recent work that a pleiotropic transcriptional regulator (either an activator or a suppressor depending on individual genes), tonicity-responsive enhancer binding protein (TonEBP), recognizes and binds R-loops, recruits METTL3, leading to RNase H1 recruitment to facilitate R-loop resolution [121]. In addition to m6A methylation, recent evidence indicates that RNA secondary structure [124,125,126] influences R-loop dynamics. de Almeida et al. demonstrated that the DDX1 RNA helicase resolves RNA G4 quadruplexes to promote R-loop formation and promote IgH class-switching recombination [126]. Moreover, Chakraborty et al. recently demonstrated that the DHX9 helicase unwinds RNA secondary structure, leading to R-loop formation [125]. Finally, similar to mutations in ADAR1, mutations in the R-loop-processing complex RNase H2 also results in Aicardi-Goutières syndrome [127], indicating a convergence of mechanism that may relate to both A-to-I editing and R-loop processing.
R-loops form readily both from protein-coding and non-coding RNA species, particularly lncRNAs [109,128]. However, unlike protein-coding RNA species, lncRNAs form R-loops both in cis (re-annealing to their DNA template) and in trans (annealing to a distal region). Antisense lncRNAs form cis R-loops that modulate the transcription of sense protein-coding genes [129,130,131,132,133]. For example, the lncRNA VIM-AS1 (VIM antisense RNA 1) forms an R-loop, leading to chromatin remodeling and expression of the sense vimentin (VIM) gene [130]. In a similar example, the lncRNA TARID (TCF21 antisense RNA inducing promoter demethylation, also known as EYA4-AS1) forms an R-loop which leads to GADD45A recruitment, demethylation, and expression of TCF21 mRNA [131]. Finally, the lncRNA GATA3-AS1 (GATA3 antisense RNA 1) forms an R-loop which promotes permissive chromatin marks and expression of GATA3 mRNA [129]. Conversely, lncRNA-mediated R-loops can also repress antisense transcription. The lncRNA ANRASSF1 (RASSF1 antisense RNA 1) forms an R-loop that recruits PRC2 to repress the RASSF1 gene [132]. Additionally, the lncRNA COOLAIR forms a cis R-loop to repress transcription at the FLC locus in Arabidopsis [133]. These examples illustrate how lncRNA forms cis R-loops to influence chromatin state and modulate transcription of protein-coding genes. In a different mechanism, Tan-Wong et al. demonstrated that sense transcription of protein-coding genes leads to cis R-loops that can promote antisense lncRNA transcription [134]. A proposed explanation for the action of cis R-loops in promoting antisense transcription is that the displaced ssDNA acts as a de novo promoter for general transcription factors [134]. However, the mechanism of trans-acting lncRNA R-loops is less clear.
lncRNA forms an R-loop in trans via the invasion of duplex DNA distal to the site of lncRNA transcription. Wahba et al. first described this phenomenon in a 2013 study, which found that lncRNAs form trans R-loops via the action of Rad51p in yeast [135]. Subsequent work in human cell lines showed that the lncRNA TERRA (telomeric-repeat-containing RNA) forms R-loops in trans at telomeres, also in a RAD51 (DNA repair protein)-dependent manner [136]. However, lncRNA R-loops may also act independently of the RAD51 trans mechanism. In budding yeast, the lncRNA GAL forms R-loops which promote rapid adaptation to changing nutrient availability [137]. Curiously, these lncRNA R-loops form in both cis and trans upon depletion of the RNA helicase Dbp2 [137], suggesting an alternative mechanism that does not depend on Rad51p. Moreover, in the Arabidopsis genome, the APOLO lncRNA forms an R-loop which regulates the PID gene in cis and multiple auxin-responsive target genes in trans to promote lateral root development [111].
Recent work suggests lncRNA formation of trans R-loops may also play a role in supporting long-range chromatin interactions and enhancer formation via the cohesin complex. The cohesin complex is a multiunit complex comprising the SMC1/3, RAD21, and STAG1/2 proteins. It forms a ring structure responsible for establishing and maintaining 3D chromatin architecture [138]. Previous work has shown that the cohesin complex subunits STAG1/2 bind to R-loops and co-localize with them genome wide [139], suggesting the possibility that R-loop/cohesin interactions may regulate 3D chromatin conformation at enhancers. In 2018, Tsai et al. found that an enhancer lncRNA (eRNA) forms a trans R-loop that recruits cohesin to regulate the activity of the Myogenin locus [140]. Moreover, genomic analysis suggests that eRNA R-loops may act in trans to facilitate enhancer-promoter interaction via Alu repeats [141]. Finally, a recent study showed that the lncRNA HOTTIP (HOXA distal transcript antisense RNA) forms trans R-loops and promotes cohesin binding and long-range enhancer interactions [112]. Taken together, these findings suggest a possible role for some lncRNA R-loops as trans-acting promoters of long-range chromatin looping via their interactions with the cohesin complex. However, future work is necessary to better understand this potential mechanism and reveal other potential roles for lncRNA R-loops.
5. Conclusions
While functional characterizations of lncRNAs have intensified in the past decade, the identification and characterization of various epitranscriptomic marks are still at their infancy as only m6A marks are highly studied. As it is evident that lncRNAs are modified by various epitranscriptomic enzymes, it is still difficult to understand what these epitranscriptomic marks mean for the functions of lncRNAs with largely unknown functions. Furthermore, a recent study about epitranscriptomic marks on tRNAs show that epitranscriptomic marks affect tertiary structures of tRNAs [142], suggesting that not only secondary structures but also tertiary structures of lncRNAs must be carefully analyzed for the presence of epitranscriptomic marks on lncRNAs. Given that sequence information alone cannot infer the functions of lncRNAs, careful inspections of secondary and tertiary structures of lncRNAs are necessary to uncover the functions of lncRNAs as their mechanisms of actions depend on their bindings to macromolecules. Further development of experimental techniques as well as re-analysis of published data from the perspective of epitranscriptomic marks and lncRNAs are necessary to uncover the causal relationships of marked lncRNAs to their binding partners, especially RBPs, and R-loop formation (Figure 4).
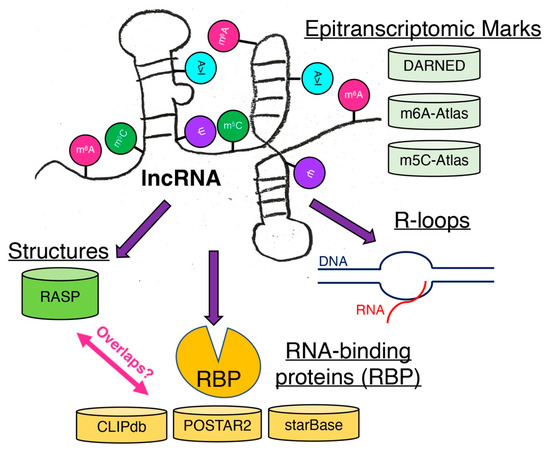
Figure 4.
Possible modifications of lncRNAs. These epitranscriptomic marks affect the structures of lncRNAs as well as their binding to RNA-binding proteins (RBPs) and the formation of R loops. Some known databases are listed to facilitate further bioinformatic analysis to connect the information.
Author Contributions
H.E.M., M.I., A.J.R.B. and S.U. wrote the manuscript, generated figures, and approved the final version of this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Novo Nordisk Foundation (NNF18OC0033438 to M.I. and S.U.); the National Institutes of Health (1R01CA241554 to A.J.R.B., F31AG072902 to H.E.M.); Stand Up to Cancer—Cancer Research UK (RT6187 to A.J.R.B); and the Greehey Family Foundation (Greehey graduate fellowship to H.E.M.).
Acknowledgments
We would like to thank the members of Bioinformatics Research Network for their continued support to further advance the development of bioinformatics and tools.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Seal, R.L.; Chen, L.L.; Griffiths-Jones, S.; Lowe, T.M.; Mathews, M.B.; O’Reilly, D.; Pierce, A.J.; Stadler, P.F.; Ulitsky, I.; Wolin, S.L.; et al. A guide to naming human non-coding RNA genes. EMBO J. 2020, 39, e103777. [Google Scholar] [CrossRef] [PubMed]
- Andergassen, D.; Rinn, J.L. From genotype to phenotype: Genetics of mammalian long non-coding RNAs in vivo. Nat. Rev. Genet. 2021, 23, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Long Noncoding RNAs: Molecular Modalities to Organismal Functions. Annu. Rev. Biochem. 2020, 89, 283–308. [Google Scholar] [CrossRef] [PubMed]
- Moore IV, J.B.; Uchida, S. Functional characterization of long noncoding RNAs. Curr. Opin. Cardiol. 2020, 35, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.W.; Wang, Y.; Chen, L.L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Garikipati, V.N.S.; Uchida, S. Elucidating the Functions of Non-Coding RNAs from the Perspective of RNA Modifications. Noncoding RNA 2021, 7, 31. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Cao, M.; Zhao, J.; Hu, G. Genome-wide methods for investigating long noncoding RNAs. Biomed. Pharm. 2019, 111, 395–401. [Google Scholar] [CrossRef]
- Hafner, M.; Katsantoni, M.; Köster, T.; Marks, J.; Mukherjee, J.; Staiger, D.; Ule, J.; Zavolan, M. CLIP and complementary methods. Nat. Rev. Methods Primers 2021, 1, 20. [Google Scholar] [CrossRef]
- Ramanathan, M.; Porter, D.F.; Khavari, P.A. Methods to study RNA-protein interactions. Nat. Methods 2019, 16, 225–234. [Google Scholar] [CrossRef]
- Ranjan, N.; Leidel, S.A. The epitranscriptome in translation regulation: mRNA and tRNA modifications as the two sides of the same coin? FEBS Lett. 2019, 593, 1483–1493. [Google Scholar] [CrossRef] [Green Version]
- Bastide, A.; David, A. Interaction of rRNA with mRNA and tRNA in Translating Mammalian Ribosome: Functional Implications in Health and Disease. Biomolecules 2018, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, S.M.; Fay, M.M.; Ivanov, P. The role of RNA modifications in the regulation of tRNA cleavage. FEBS Lett. 2018, 592, 2828–2844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piatkowski, P.; Baginski, B.; Wirecki, T.K.; de Crecy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018, 46, D303–D307. [Google Scholar] [CrossRef]
- Roundtree, I.A.; He, C. RNA epigenetics-chemical messages for posttranscriptional gene regulation. Curr. Opin. Chem. Biol. 2016, 30, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saletore, Y.; Meyer, K.; Korlach, J.; Vilfan, I.D.; Jaffrey, S.; Mason, C.E. The birth of the Epitranscriptome: Deciphering the function of RNA modifications. Genome Biol. 2012, 13, 175. [Google Scholar] [CrossRef] [Green Version]
- Wiener, D.; Schwartz, S. The epitranscriptome beyond m(6)A. Nat. Rev. Genet. 2021, 22, 119–131. [Google Scholar] [CrossRef]
- Angelova, M.T.; Dimitrova, D.G.; Dinges, N.; Lence, T.; Worpenberg, L.; Carre, C.; Roignant, J.Y. The Emerging Field of Epitranscriptomics in Neurodevelopmental and Neuronal Disorders. Front. Bioeng. Biotechnol. 2018, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Mohapatra, T. Deciphering Epitranscriptome: Modification of mRNA Bases Provides a New Perspective for Post-transcriptional Regulation of Gene Expression. Front. Cell Dev. Biol. 2021, 9, 628415. [Google Scholar] [CrossRef]
- Li, S.; Mason, C.E. The pivotal regulatory landscape of RNA modifications. Annu. Rev. Genom. Hum. Genet. 2014, 15, 127–150. [Google Scholar] [CrossRef]
- Uchida, S.; Jones, S.P. RNA Editing: Unexplored Opportunities in the Cardiovascular System. Circ. Res. 2018, 122, 399–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Meng, J.; Su, R.; Zhang, J.; Chen, J.; Ma, X.; Xia, Q. Epitranscriptomics in liver disease: Basic concepts and therapeutic potential. J. Hepatol. 2020, 73, 664–679. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.; Wang, Q.H.; Zhu, C.B.; Ma, J.; Jin, W.L. Deciphering the Epitranscriptome in Cancer. Trends Cancer 2018, 4, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Kazimierczyk, M.; Wrzesinski, J. Long Non-Coding RNA Epigenetics. Int. J. Mol. Sci. 2021, 22, 6166. [Google Scholar] [CrossRef]
- Reineke, L.C.; Kedersha, N.; Langereis, M.A.; van Kuppeveld, F.J.; Lloyd, R.E. Stress granules regulate double-stranded RNA-dependent protein kinase activation through a complex containing G3BP1 and Caprin1. mBio 2015, 6, e02486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabo, S.; Meurs, E.F. dsRNA-dependent protein kinase PKR and its role in stress, signaling and HCV infection. Viruses 2012, 4, 2598–2635. [Google Scholar] [CrossRef] [Green Version]
- Mertens, P. The dsRNA viruses. Virus Res. 2004, 101, 3–13. [Google Scholar] [CrossRef]
- Sadeq, S.; Al-Hashimi, S.; Cusack, C.M.; Werner, A. Endogenous Double-Stranded RNA. Noncoding RNA 2021, 7, 15. [Google Scholar] [CrossRef]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [Green Version]
- Trigiante, G.; Blanes Ruiz, N.; Cerase, A. Emerging Roles of Repetitive and Repeat-Containing RNA in Nuclear and Chromatin Organization and Gene Expression. Front. Cell Dev. Biol. 2021, 9, 735527. [Google Scholar] [CrossRef]
- Kertesz, M.; Wan, Y.; Mazor, E.; Rinn, J.L.; Nutter, R.C.; Chang, H.Y.; Segal, E. Genome-wide measurement of RNA secondary structure in yeast. Nature 2010, 467, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Rouskin, S.; Zubradt, M.; Washietl, S.; Kellis, M.; Weissman, J.S. Genome-wide probing of RNA structure reveals active unfolding of mRNA structures in vivo. Nature 2014, 505, 701–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, E.; Sterne-Weiler, T.; O’Hanlon, D.; Blencowe, B.J. Global Mapping of Human RNA-RNA Interactions. Mol. Cell 2016, 62, 618–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Gong, J.; Zhang, Q.C. PARIS: Psoralen Analysis of RNA Interactions and Structures with High Throughput and Resolution. Methods Mol. Biol. 2018, 1649, 59–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, C.; Cai, Z.; Ye, R.; Su, R.; Hu, N.; Zhao, H.; Xue, Y. Global in situ profiling of RNA-RNA spatial interactions with RIC-seq. Nat. Protoc. 2021, 16, 2916–2946. [Google Scholar] [CrossRef]
- Cai, Z.; Cao, C.; Ji, L.; Ye, R.; Wang, D.; Xia, C.; Wang, S.; Du, Z.; Hu, N.; Yu, X.; et al. RIC-seq for global in situ profiling of RNA-RNA spatial interactions. Nature 2020, 582, 432–437. [Google Scholar] [CrossRef]
- Lucks, J.B.; Mortimer, S.A.; Trapnell, C.; Luo, S.; Aviran, S.; Schroth, G.P.; Pachter, L.; Doudna, J.A.; Arkin, A.P. Multiplexed RNA structure characterization with selective 2′-hydroxyl acylation analyzed by primer extension sequencing (SHAPE-Seq). Proc. Natl. Acad. Sci. USA 2011, 108, 11063–11068. [Google Scholar] [CrossRef] [Green Version]
- Smola, M.J.; Weeks, K.M. In-cell RNA structure probing with SHAPE-MaP. Nat. Protoc. 2018, 13, 1181–1195. [Google Scholar] [CrossRef]
- Smola, M.J.; Rice, G.M.; Busan, S.; Siegfried, N.A.; Weeks, K.M. Selective 2′-hydroxyl acylation analyzed by primer extension and mutational profiling (SHAPE-MaP) for direct, versatile and accurate RNA structure analysis. Nat. Protoc. 2015, 10, 1643–1669. [Google Scholar] [CrossRef]
- Li, P.; Zhou, X.; Xu, K.; Zhang, Q.C. RASP: An atlas of transcriptome-wide RNA secondary structure probing data. Nucleic Acids Res. 2021, 49, D183–D191. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, L.; Chen, S.; Liu, H. A brief review of RNA modification related database resources. Methods 2021, 21, S1046-2023. [Google Scholar] [CrossRef] [PubMed]
- Xuan, J.J.; Sun, W.J.; Lin, P.H.; Zhou, K.R.; Liu, S.; Zheng, L.L.; Qu, L.H.; Yang, J.H. RMBase v2.0: Deciphering the map of RNA modifications from epitranscriptome sequencing data. Nucleic Acids Res. 2018, 46, D327–D334. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Li, H.; Liang, J.; Zhao, Q.; Xie, Y.; Ren, J.; Zuo, Z. RMVar: An updated database of functional variants involved in RNA modifications. Nucleic Acids Res. 2021, 49, D1405–D1412. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Ding, T.; Zuo, Z.; Xu, Z.; Deng, J.; Wei, Z. Regulation of MAVS Expression and Signaling Function in the Antiviral Innate Immune Response. Front. Immunol. 2020, 11, 1030. [Google Scholar] [CrossRef] [PubMed]
- Chintakuntlawar, A.V.; Zhou, X.; Rajaiya, J.; Chodosh, J. Viral capsid is a pathogen-associated molecular pattern in adenovirus keratitis. PLoS Pathog. 2010, 6, e1000841. [Google Scholar] [CrossRef]
- Thompson, M.R.; Kaminski, J.J.; Kurt-Jones, E.A.; Fitzgerald, K.A. Pattern recognition receptors and the innate immune response to viral infection. Viruses 2011, 3, 920–940. [Google Scholar] [CrossRef] [Green Version]
- Kimura, S.; Matsumiya, T.; Shiba, Y.; Nakanishi, M.; Hayakari, R.; Kawaguchi, S.; Yoshida, H.; Imaizumi, T. The Essential Role of Double-Stranded RNA-Dependent Antiviral Signaling in the Degradation of Nonself Single-Stranded RNA in Nonimmune Cells. J. Immunol. 2018, 201, 1044–1052. [Google Scholar] [CrossRef]
- Okamoto, M.; Tsukamoto, H.; Kouwaki, T.; Seya, T.; Oshiumi, H. Recognition of Viral RNA by Pattern Recognition Receptors in the Induction of Innate Immunity and Excessive Inflammation During Respiratory Viral Infections. Viral Immunol. 2017, 30, 408–420. [Google Scholar] [CrossRef]
- Saito, T.; Gale, M., Jr. Differential recognition of double-stranded RNA by RIG-I-like receptors in antiviral immunity. J. Exp. Med. 2008, 205, 1523–1527. [Google Scholar] [CrossRef] [Green Version]
- Winkler, R.; Gillis, E.; Lasman, L.; Safra, M.; Geula, S.; Soyris, C.; Nachshon, A.; Tai-Schmiedel, J.; Friedman, N.; Le-Trilling, V.T.K.; et al. m(6)A modification controls the innate immune response to infection by targeting type I interferons. Nat. Immunol. 2019, 20, 173–182. [Google Scholar] [CrossRef]
- Rubio, R.M.; Depledge, D.P.; Bianco, C.; Thompson, L.; Mohr, I. RNA m(6) A modification enzymes shape innate responses to DNA by regulating interferon beta. Genes Dev. 2018, 32, 1472–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; Wang, T.; Tang, Q.; Li, G.; Wu, P.; Chen, K. Long Non-coding RNAs: Regulators of Viral Infection and the Interferon Antiviral Response. Front. Microbiol. 2018, 9, 1621. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Parisien, M.; Dai, Q.; Zheng, G.; He, C.; Pan, T. Probing N6-methyladenosine RNA modification status at single nucleotide resolution in mRNA and long noncoding RNA. RNA 2013, 19, 1848–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [Green Version]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Wang, M.C.; McCown, P.J.; Schiefelbein, G.E.; Brown, J.A. Secondary Structural Model of MALAT1 Becomes Unstructured in Chronic Myeloid Leukemia and Undergoes Structural Rearrangement in Cervical Cancer. Noncoding RNA 2021, 7, 6. [Google Scholar] [CrossRef]
- McCown, P.J.; Wang, M.C.; Jaeger, L.; Brown, J.A. Secondary Structural Model of Human MALAT1 Reveals Multiple Structure-Function Relationships. Int. J. Mol. Sci. 2019, 20, 5610. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.I.; Parisien, M.; Dai, Q.; Liu, N.; Diatchenko, L.; Sachleben, J.R.; Pan, T. N(6)-Methyladenosine Modification in a Long Noncoding RNA Hairpin Predisposes Its Conformation to Protein Binding. J. Mol. Biol. 2016, 428, 822–833. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Wei, C.; Wang, D.; Wang, X.; Wang, J.; Hu, Y.; Mao, X. The small RNA mascRNA differentially regulates TLR-induced proinflammatory and antiviral responses. JCI Insight 2021, 6, e150833. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Z.; Liu, L.; Yang, Z.; Liu, S.; Ma, Z.; Liu, Y.; Ma, Y.; Zhang, L.; Zhang, X.; et al. LncRNA Malat1 inhibition of TDP43 cleavage suppresses IRF3-initiated antiviral innate immunity. Proc. Natl. Acad. Sci. USA 2020, 117, 23695–23706. [Google Scholar] [CrossRef]
- Feng, L.L.; Xin, W.N.; Tian, X.L. MALAT1 modulates miR-146′s protection of microvascular endothelial cells against LPS-induced NF-kappaB activation and inflammatory injury. Innate Immun. 2019, 25, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gast, M.; Rauch, B.H.; Nakagawa, S.; Haghikia, A.; Jasina, A.; Haas, J.; Nath, N.; Jensen, L.; Stroux, A.; Bohm, A.; et al. Immune system-mediated atherosclerosis caused by deficiency of long non-coding RNA MALAT1 in ApoE-/-mice. Cardiovasc. Res. 2019, 115, 302–314. [Google Scholar] [CrossRef]
- Gast, M.; Schroen, B.; Voigt, A.; Haas, J.; Kuehl, U.; Lassner, D.; Skurk, C.; Escher, F.; Wang, X.; Kratzer, A.; et al. Long noncoding RNA MALAT1-derived mascRNA is involved in cardiovascular innate immunity. J. Mol. Cell Biol. 2016, 8, 178–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Eckwahl, M.J.; Zhou, K.I.; Pan, T. Sensitive and quantitative probing of pseudouridine modification in mRNA and long noncoding RNA. RNA 2019, 25, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- Nance, K.D.; Meier, J.L. Modifications in an Emergency: The Role of N1-Methylpseudouridine in COVID-19 Vaccines. ACS Cent. Sci. 2021, 7, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Batzer, M.A.; Deininger, P.L. Alu repeats and human genomic diversity. Nat. Rev. Genet. 2002, 3, 370–379. [Google Scholar] [CrossRef]
- Fumagalli, D.; Gacquer, D.; Rothe, F.; Lefort, A.; Libert, F.; Brown, D.; Kheddoumi, N.; Shlien, A.; Konopka, T.; Salgado, R.; et al. Principles Governing A-to-I RNA Editing in the Breast Cancer Transcriptome. Cell Rep. 2015, 13, 277–289. [Google Scholar] [CrossRef] [Green Version]
- John, D.; Weirick, T.; Dimmeler, S.; Uchida, S. RNAEditor: Easy detection of RNA editing events and the introduction of editing islands. Brief. Bioinform. 2017, 18, 993–1001. [Google Scholar] [CrossRef]
- Pestal, K.; Funk, C.C.; Snyder, J.M.; Price, N.D.; Treuting, P.M.; Stetson, D.B. Isoforms of RNA-Editing Enzyme ADAR1 Independently Control Nucleic Acid Sensor MDA5-Driven Autoimmunity and Multi-organ Development. Immunity 2015, 43, 933–944. [Google Scholar] [CrossRef] [Green Version]
- Liddicoat, B.J.; Piskol, R.; Chalk, A.M.; Ramaswami, G.; Higuchi, M.; Hartner, J.C.; Li, J.B.; Seeburg, P.H.; Walkley, C.R. RNA editing by ADAR1 prevents MDA5 sensing of endogenous dsRNA as nonself. Science 2015, 349, 1115–1120. [Google Scholar] [CrossRef] [Green Version]
- Mannion, N.M.; Greenwood, S.M.; Young, R.; Cox, S.; Brindle, J.; Read, D.; Nellaker, C.; Vesely, C.; Ponting, C.P.; McLaughlin, P.J.; et al. The RNA-editing enzyme ADAR1 controls innate immune responses to RNA. Cell Rep. 2014, 9, 1482–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanai, M.; Kojima, S.; Sakai, M.; Komorizono, R.; Tomonaga, K.; Makino, A. ADAR2 Is Involved in Self and Nonself Recognition of Borna Disease Virus Genomic RNA in the Nucleus. J. Virol. 2020, 94, e01513–e01519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pujantell, M.; Riveira-Munoz, E.; Badia, R.; Castellvi, M.; Garcia-Vidal, E.; Sirera, G.; Puig, T.; Ramirez, C.; Clotet, B.; Este, J.A.; et al. RNA editing by ADAR1 regulates innate and antiviral immune functions in primary macrophages. Sci. Rep. 2017, 7, 13339. [Google Scholar] [CrossRef] [PubMed]
- Solomon, O.; Di Segni, A.; Cesarkas, K.; Porath, H.T.; Marcu-Malina, V.; Mizrahi, O.; Stern-Ginossar, N.; Kol, N.; Farage-Barhom, S.; Glick-Saar, E.; et al. RNA editing by ADAR1 leads to context-dependent transcriptome-wide changes in RNA secondary structure. Nat. Commun. 2017, 8, 1440. [Google Scholar] [CrossRef]
- Porath, H.T.; Knisbacher, B.A.; Eisenberg, E.; Levanon, E.Y. Massive A-to-I RNA editing is common across the Metazoa and correlates with dsRNA abundance. Genome Biol. 2017, 18, 185. [Google Scholar] [CrossRef]
- Galipon, J.; Ishii, R.; Suzuki, Y.; Tomita, M.; Ui-Tei, K. Differential Binding of Three Major Human ADAR Isoforms to Coding and Long Non-Coding Transcripts. Genes 2017, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Xiang, J.F.; Yang, Q.; Liu, C.X.; Wu, M.; Chen, L.L.; Yang, L. N(6)-Methyladenosines Modulate A-to-I RNA Editing. Mol. Cell 2018, 69, 126–135.e6. [Google Scholar] [CrossRef]
- Corley, M.; Burns, M.C.; Yeo, G.W. How RNA-Binding Proteins Interact with RNA: Molecules and Mechanisms. Mol. Cell 2020, 78, 9–29. [Google Scholar] [CrossRef]
- Elguindy, M.M.; Mendell, J.T. NORAD-induced Pumilio phase separation is required for genome stability. Nature 2021, 595, 303–308. [Google Scholar] [CrossRef]
- Elguindy, M.M.; Kopp, F.; Goodarzi, M.; Rehfeld, F.; Thomas, A.; Chang, T.C.; Mendell, J.T. PUMILIO, but not RBMX, binding is required for regulation of genomic stability by noncoding RNA NORAD. Elife 2019, 8, e48625. [Google Scholar] [CrossRef]
- Lee, S.; Kopp, F.; Chang, T.C.; Sataluri, A.; Chen, B.; Sivakumar, S.; Yu, H.; Xie, Y.; Mendell, J.T. Noncoding RNA NORAD Regulates Genomic Stability by Sequestering PUMILIO Proteins. Cell 2016, 164, 69–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castello, A.; Fischer, B.; Frese, C.K.; Horos, R.; Alleaume, A.M.; Foehr, S.; Curk, T.; Krijgsveld, J.; Hentze, M.W. Comprehensive Identification of RNA-Binding Domains in Human Cells. Mol. Cell 2016, 63, 696–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckmann, B.M.; Horos, R.; Fischer, B.; Castello, A.; Eichelbaum, K.; Alleaume, A.M.; Schwarzl, T.; Curk, T.; Foehr, S.; Huber, W.; et al. The RNA-binding proteomes from yeast to man harbour conserved enigmRBPs. Nat. Commun. 2015, 6, 10127. [Google Scholar] [CrossRef] [PubMed]
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Beckmann, B.M.; Strein, C.; Davey, N.E.; Humphreys, D.T.; Preiss, T.; Steinmetz, L.M.; et al. Insights into RNA biology from an atlas of mammalian mRNA-binding proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Ohsumi, T.K.; Kung, J.T.; Ogawa, Y.; Grau, D.J.; Sarma, K.; Song, J.J.; Kingston, R.E.; Borowsky, M.; Lee, J.T. Genome-wide identification of polycomb-associated RNAs by RIP-seq. Mol. Cell 2010, 40, 939–953. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Goodrich, K.J.; Gooding, A.R.; Naeem, H.; Archer, S.; Paucek, R.D.; Youmans, D.T.; Cech, T.R.; Davidovich, C. Targeting of Polycomb Repressive Complex 2 to RNA by Short Repeats of Consecutive Guanines. Mol. Cell 2017, 65, 1056–1067.e5. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.C.; Di, C.; Hu, B.; Zhou, M.; Liu, Y.; Song, N.; Li, Y.; Umetsu, J.; Lu, Z.J. CLIPdb: A CLIP-seq database for protein-RNA interactions. BMC Genom. 2015, 16, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Xu, G.; Yang, Y.T.; Xu, Z.; Chen, X.; Shi, B.; Xie, D.; Lu, Z.J.; Wang, P. POSTAR2: Deciphering the post-transcriptional regulatory logics. Nucleic Acids Res. 2019, 47, D203–D211. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [Green Version]
- Hentze, M.W.; Castello, A.; Schwarzl, T.; Preiss, T. A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell Biol. 2018, 19, 327–341. [Google Scholar] [CrossRef]
- Dodson, R.E.; Shapiro, D.J. Regulation of pathways of mRNA destabilization and stabilization. Prog. Nucleic Acid Res. Mol. Biol. 2002, 72, 129–164. [Google Scholar] [CrossRef] [PubMed]
- Weirick, T.; Militello, G.; Hosen, M.R.; John, D.; Moore Iv, J.B.; Uchida, S. Investigation of RNA Editing Sites within Bound Regions of RNA-Binding Proteins. High Throughput 2019, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stellos, K.; Gatsiou, A.; Stamatelopoulos, K.; Perisic Matic, L.; John, D.; Lunella, F.F.; Jae, N.; Rossbach, O.; Amrhein, C.; Sigala, F.; et al. Adenosine-to-inosine RNA editing controls cathepsin S expression in atherosclerosis by enabling HuR-mediated post-transcriptional regulation. Nat. Med. 2016, 22, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.X.; So, E.; Devlin, J.L.; Zhao, Y.; Wu, M.; Cheung, V.G. ADAR regulates RNA editing, transcript stability, and gene expression. Cell Rep. 2013, 5, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Priyanka, P.; Sharma, M.; Das, S.; Saxena, S. The lncRNA HMS recruits RNA-binding protein HuR to stabilize the 3′-UTR of HOXC10 mRNA. J. Biol. Chem. 2021, 297, 100997. [Google Scholar] [CrossRef]
- Kiran, A.M.; O’Mahony, J.J.; Sanjeev, K.; Baranov, P.V. Darned in 2013: Inclusion of model organisms and linking with Wikipedia. Nucleic Acids Res. 2013, 41, D258–D261. [Google Scholar] [CrossRef]
- Kiran, A.; Baranov, P.V. DARNED: A DAtabase of RNa EDiting in humans. Bioinformatics 2010, 26, 1772–1776. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Chen, K.; Song, B.; Ma, J.; Wu, X.; Xu, Q.; Wei, Z.; Su, J.; Liu, G.; Rong, R.; et al. m6A-Atlas: A comprehensive knowledgebase for unraveling the N6-methyladenosine (m6A) epitranscriptome. Nucleic Acids Res. 2021, 49, D134–D143. [Google Scholar] [CrossRef]
- Ma, J.; Song, B.; Wei, Z.; Huang, D.; Zhang, Y.; Su, J.; de Magalhaes, J.P.; Rigden, D.J.; Meng, J.; Chen, K. m5C-Atlas: A comprehensive database for decoding and annotating the 5-methylcytosine (m5C) epitranscriptome. Nucleic Acids Res. 2022, 50, D196–D203. [Google Scholar] [CrossRef]
- Von Hagen, F.; Gundert, L.; Strick, A.; Klumper, N.; Schmidt, D.; Kristiansen, G.; Tolkach, Y.; Toma, M.; Ritter, M.; Ellinger, J. N(6) -Methyladenosine (m(6) A) readers are dysregulated in renal cell carcinoma. Mol. Carcinog. 2021, 60, 354–362. [Google Scholar] [CrossRef]
- Patil, D.P.; Pickering, B.F.; Jaffrey, S.R. Reading m(6)A in the Transcriptome: M(6)A-Binding Proteins. Trends Cell Biol. 2018, 28, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, L.; Zhou, Y. m6Adecom: Analysis of m(6)A profile matrix based on graph regularized non-negative matrix factorization. Methods 2022, 22, S1046-2023. [Google Scholar] [CrossRef] [PubMed]
- Ginno, P.A.; Lim, Y.W.; Lott, P.L.; Korf, I.; Chedin, F. GC skew at the 5′ and 3′ ends of human genes links R-loop formation to epigenetic regulation and transcription termination. Genome Res. 2013, 23, 1590–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, R.P.; Xu, Q.; Weinberger, P.M. R-Loop Physiology and Pathology: A Brief Review. DNA Cell Biol. 2020, 39, 1914–1925. [Google Scholar] [CrossRef] [PubMed]
- Crossley, M.P.; Bocek, M.; Cimprich, K.A. R-Loops as Cellular Regulators and Genomic Threats. Mol. Cell 2019, 73, 398–411. [Google Scholar] [CrossRef] [Green Version]
- Richard, P.; Manley, J.L. R Loops and Links to Human Disease. J. Mol. Biol. 2017, 429, 3168–3180. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.L.; Pasero, P. Transcription-Replication Conflicts: Orientation Matters. Cell 2017, 170, 603–604. [Google Scholar] [CrossRef]
- Lockhart, A.; Pires, V.B.; Bento, F.; Kellner, V.; Luke-Glaser, S.; Yakoub, G.; Ulrich, H.D.; Luke, B. RNase H1 and H2 Are Differentially Regulated to Process RNA-DNA Hybrids. Cell Rep. 2019, 29, 2890–2900. [Google Scholar] [CrossRef]
- Niehrs, C.; Luke, B. Regulatory R-loops as facilitators of gene expression and genome stability. Nat. Rev. Mol. Cell Biol. 2020, 21, 167–178. [Google Scholar] [CrossRef]
- Garcia-Muse, T.; Aguilera, A. R Loops: From Physiological to Pathological Roles. Cell 2019, 179, 604–618. [Google Scholar] [CrossRef]
- Ariel, F.; Lucero, L.; Christ, A.; Mammarella, M.F.; Jegu, T.; Veluchamy, A.; Mariappan, K.; Latrasse, D.; Blein, T.; Liu, C.; et al. R-Loop Mediated trans Action of the APOLO Long Noncoding RNA. Mol. Cell 2020, 77, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zhu, G.; Eshelman, M.A.; Fung, T.K.; Lai, Q.; Wang, F.; Zeisig, B.B.; Lesperance, J.; Ma, X.; Chen, S.; et al. HOTTIP-dependent R-loop formation regulates CTCF boundary activity and TAD integrity in leukemia. Mol. Cell 2022, 82, 833–851. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.V.; Feretzaki, M.; Lingner, J. The makings of TERRA R-loops at chromosome ends. Cell Cycle 2021, 20, 1745–1759. [Google Scholar] [CrossRef] [PubMed]
- Vohhodina, J.; Goehring, L.J.; Liu, B.; Kong, Q.; Botchkarev, V.V., Jr.; Huynh, M.; Liu, Z.; Abderazzaq, F.O.; Clark, A.P.; Ficarro, S.B.; et al. BRCA1 binds TERRA RNA and suppresses R-Loop-based telomeric DNA damage. Nat. Commun. 2021, 12, 3542. [Google Scholar] [CrossRef]
- Ghisays, F.; Garzia, A.; Wang, H.; Canasto-Chibuque, C.; Hohl, M.; Savage, S.A.; Tuschl, T.; Petrini, J.H.J. RTEL1 influences the abundance and localization of TERRA RNA. Nat. Commun. 2021, 12, 3016. [Google Scholar] [CrossRef]
- Lalonde, M.; Chartrand, P. TERRA, a Multifaceted Regulator of Telomerase Activity at Telomeres. J. Mol. Biol. 2020, 432, 4232–4243. [Google Scholar] [CrossRef]
- Saha, A.; Nanavaty, V.P.; Li, B. Telomere and Subtelomere R-loops and Antigenic Variation in Trypanosomes. J. Mol. Biol. 2020, 432, 4167–4185. [Google Scholar] [CrossRef]
- Cusanelli, E.; Chartrand, P. Telomeric repeat-containing RNA TERRA: A noncoding RNA connecting telomere biology to genome integrity. Front. Genet. 2015, 6, 143. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, V.; Crittin, J.; Grolimund, L.; Lingner, J. The THO complex component Thp2 counteracts telomeric R-loops and telomere shortening. EMBO J. 2013, 32, 2861–2871. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Xu, S.; Zhang, M.; Wu, Q. Systematic functional characterization of antisense eRNA of protocadherin alpha composite enhancer. Genes Dev. 2021, 35, 1383–1394. [Google Scholar] [CrossRef]
- Kang, H.J.; Cheon, N.Y.; Park, H.; Jeong, G.W.; Ye, B.J.; Yoo, E.J.; Lee, J.H.; Hur, J.H.; Lee, E.A.; Kim, H.; et al. TonEBP recognizes R-loops and initiates m6A RNA methylation for R-loop resolution. Nucleic Acids Res. 2021, 49, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Abakir, A.; Giles, T.C.; Cristini, A.; Foster, J.M.; Dai, N.; Starczak, M.; Rubio-Roldan, A.; Li, M.; Eleftheriou, M.; Crutchley, J.; et al. N(6)-methyladenosine regulates the stability of RNA:DNA hybrids in human cells. Nat. Genet. 2020, 52, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, Q.L.; Xu, W.; Zhang, Y.C.; Yang, Y.; Ju, L.F.; Chen, J.; Chen, Y.S.; Li, K.; Ren, J.; et al. m(6)A promotes R-loop formation to facilitate transcription termination. Cell Res. 2019, 29, 1035–1038. [Google Scholar] [CrossRef] [PubMed]
- Miglietta, G.; Russo, M.; Capranico, G. G-quadruplex-R-loop interactions and the mechanism of anticancer G-quadruplex binders. Nucleic Acids Res. 2020, 48, 11942–11957. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Huang, J.T.J.; Hiom, K. DHX9 helicase promotes R-loop formation in cells with impaired RNA splicing. Nat. Commun. 2018, 9, 4346. [Google Scholar] [CrossRef]
- Ribeiro de Almeida, C.; Dhir, S.; Dhir, A.; Moghaddam, A.E.; Sattentau, Q.; Meinhart, A.; Proudfoot, N.J. RNA Helicase DDX1 Converts RNA G-Quadruplex Structures into R-Loops to Promote IgH Class Switch Recombination. Mol. Cell 2018, 70, 650–662. [Google Scholar] [CrossRef] [Green Version]
- Crow, Y.J.; Leitch, A.; Hayward, B.E.; Garner, A.; Parmar, R.; Griffith, E.; Ali, M.; Semple, C.; Aicardi, J.; Babul-Hirji, R.; et al. Mutations in genes encoding ribonuclease H2 subunits cause Aicardi-Goutieres syndrome and mimic congenital viral brain infection. Nat. Genet. 2006, 38, 910–916. [Google Scholar] [CrossRef]
- Kuznetsov, V.A.; Bondarenko, V.; Wongsurawat, T.; Yenamandra, S.P.; Jenjaroenpun, P. Toward predictive R-loop computational biology: Genome-scale prediction of R-loops reveals their association with complex promoter structures, G-quadruplexes and transcriptionally active enhancers. Nucleic Acids Res. 2018, 46, 7566–7585. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, H.R.; Shaginurova, G.; Kim, L.C.; Chapman, N.; Spurlock, C.F., 3rd; Aune, T.M. Divergent lncRNA GATA3-AS1 Regulates GATA3 Transcription in T-Helper 2 Cells. Front. Immunol. 2018, 9, 2512. [Google Scholar] [CrossRef]
- Boque-Sastre, R.; Soler, M.; Oliveira-Mateos, C.; Portela, A.; Moutinho, C.; Sayols, S.; Villanueva, A.; Esteller, M.; Guil, S. Head-to-head antisense transcription and R-loop formation promotes transcriptional activation. Proc. Natl. Acad. Sci. USA 2015, 112, 5785–5790. [Google Scholar] [CrossRef] [Green Version]
- Arab, K.; Park, Y.J.; Lindroth, A.M.; Schafer, A.; Oakes, C.; Weichenhan, D.; Lukanova, A.; Lundin, E.; Risch, A.; Meister, M.; et al. Long noncoding RNA TARID directs demethylation and activation of the tumor suppressor TCF21 via GADD45A. Mol. Cell 2014, 55, 604–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckedorff, F.C.; Ayupe, A.C.; Crocci-Souza, R.; Amaral, M.S.; Nakaya, H.I.; Soltys, D.T.; Menck, C.F.; Reis, E.M.; Verjovski-Almeida, S. The intronic long noncoding RNA ANRASSF1 recruits PRC2 to the RASSF1A promoter, reducing the expression of RASSF1A and increasing cell proliferation. PLoS Genet. 2013, 9, e1003705. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Csorba, T.; Skourti-Stathaki, K.; Proudfoot, N.J.; Dean, C. R-loop stabilization represses antisense transcription at the Arabidopsis FLC locus. Science 2013, 340, 619–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan-Wong, S.M.; Dhir, S.; Proudfoot, N.J. R-Loops Promote Antisense Transcription across the Mammalian Genome. Mol. Cell 2019, 76, 600–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahba, L.; Gore, S.K.; Koshland, D. The homologous recombination machinery modulates the formation of RNA-DNA hybrids and associated chromosome instability. Elife 2013, 2, e00505. [Google Scholar] [CrossRef]
- Feretzaki, M.; Pospisilova, M.; Valador Fernandes, R.; Lunardi, T.; Krejci, L.; Lingner, J. RAD51-dependent recruitment of TERRA lncRNA to telomeres through R-loops. Nature 2020, 587, 303–308. [Google Scholar] [CrossRef]
- Cloutier, S.C.; Wang, S.; Ma, W.K.; Al Husini, N.; Dhoondia, Z.; Ansari, A.; Pascuzzi, P.E.; Tran, E.J. Regulated Formation of lncRNA-DNA Hybrids Enables Faster Transcriptional Induction and Environmental Adaptation. Mol. Cell 2016, 61, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Kuru-Schors, M.; Haemmerle, M.; Gutschner, T. The Cohesin Complex and Its Interplay with Non-Coding RNAs. Noncoding RNA 2021, 7, 67. [Google Scholar] [CrossRef]
- Pan, H.; Jin, M.; Ghadiyaram, A.; Kaur, P.; Miller, H.E.; Ta, H.M.; Liu, M.; Fan, Y.; Mahn, C.; Gorthi, A.; et al. Cohesin SA1 and SA2 are RNA binding proteins that localize to RNA containing regions on DNA. Nucleic Acids Res. 2020, 48, 5639–5655. [Google Scholar] [CrossRef]
- Tsai, P.F.; Dell’Orso, S.; Rodriguez, J.; Vivanco, K.O.; Ko, K.D.; Jiang, K.; Juan, A.H.; Sarshad, A.A.; Vian, L.; Tran, M.; et al. A Muscle-Specific Enhancer RNA Mediates Cohesin Recruitment and Regulates Transcription In trans. Mol. Cell 2018, 71, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Li, F.; Zhang, Z. A hypothetical model of trans-acting R-loops-mediated promoter-enhancer interactions by Alu elements. J. Genet. Genom. 2021, 48, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Biedenbander, T.; de Jesus, V.; Schmidt-Dengler, M.; Helm, M.; Corzilius, B.; Furtig, B. RNA modifications stabilize the tertiary structure of tRNAfMet by locally increasing conformational dynamics. Nucleic Acids Res. 2022, 50, 2334–2349. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).