
Trichocladium solani sp. nov.—A New Pathogen on Potato Tubers Causing Yellow Rot

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
3. Results
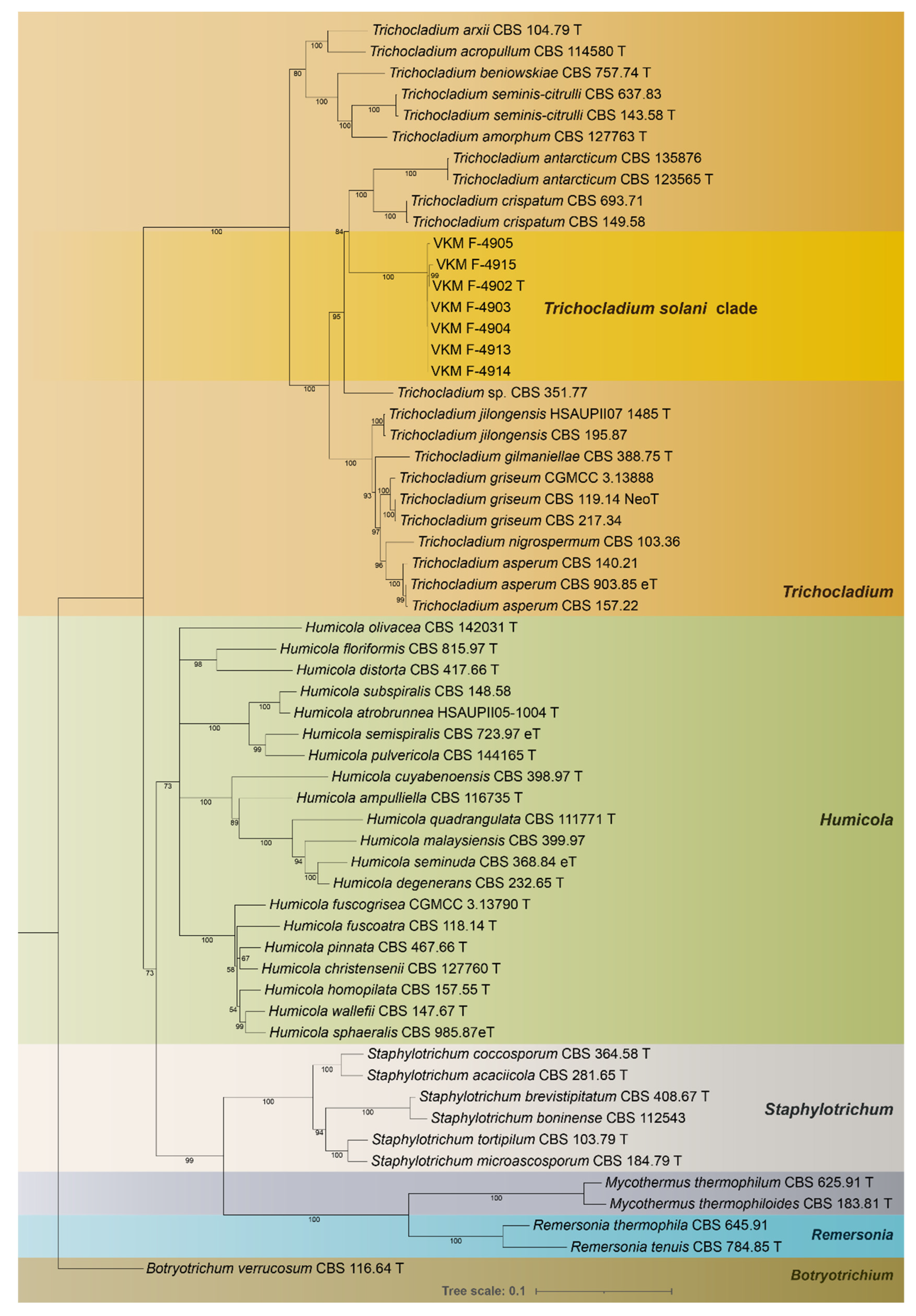
3.1. Phylogeny
3.2. Pathogenicity towards Potato Tubers
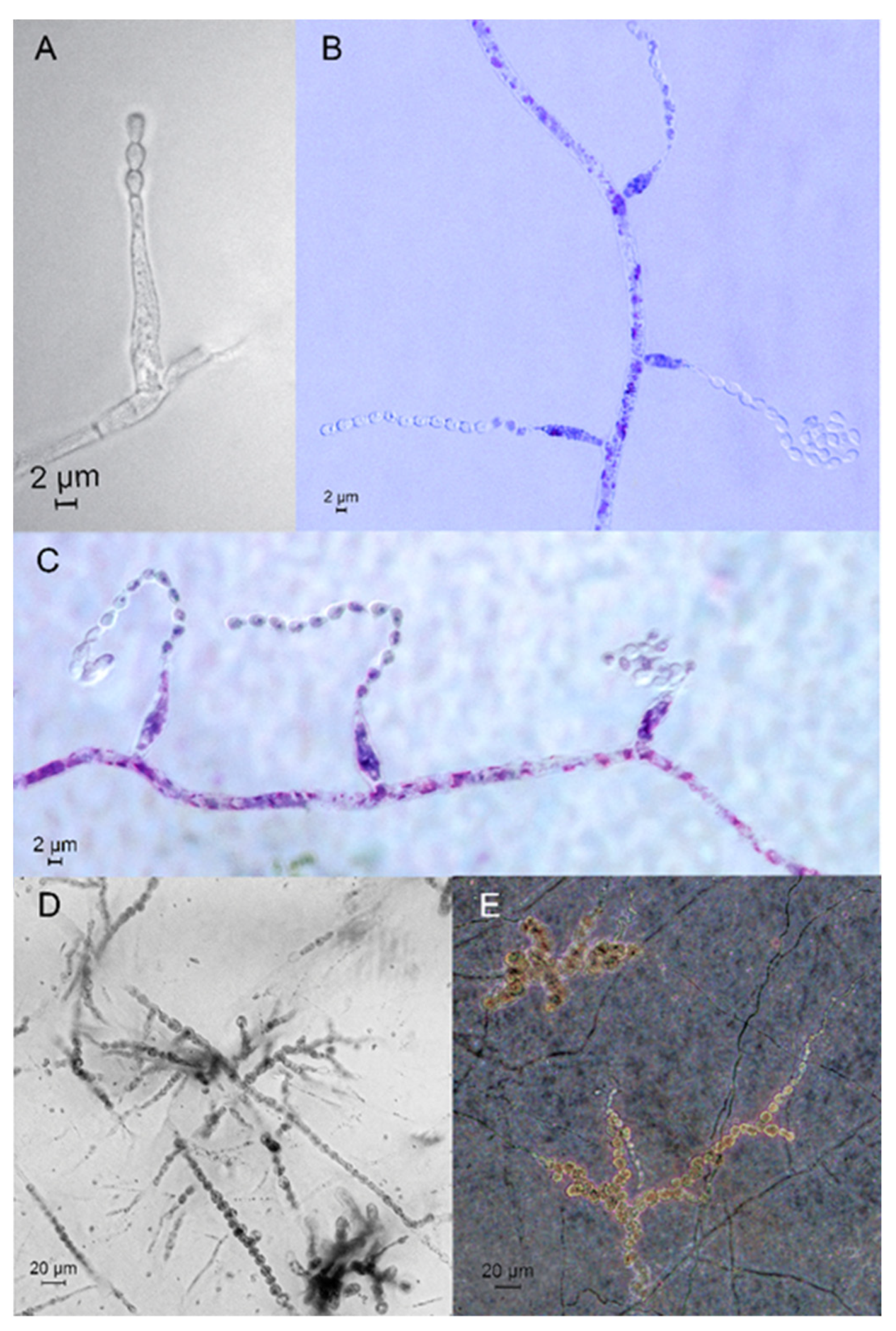
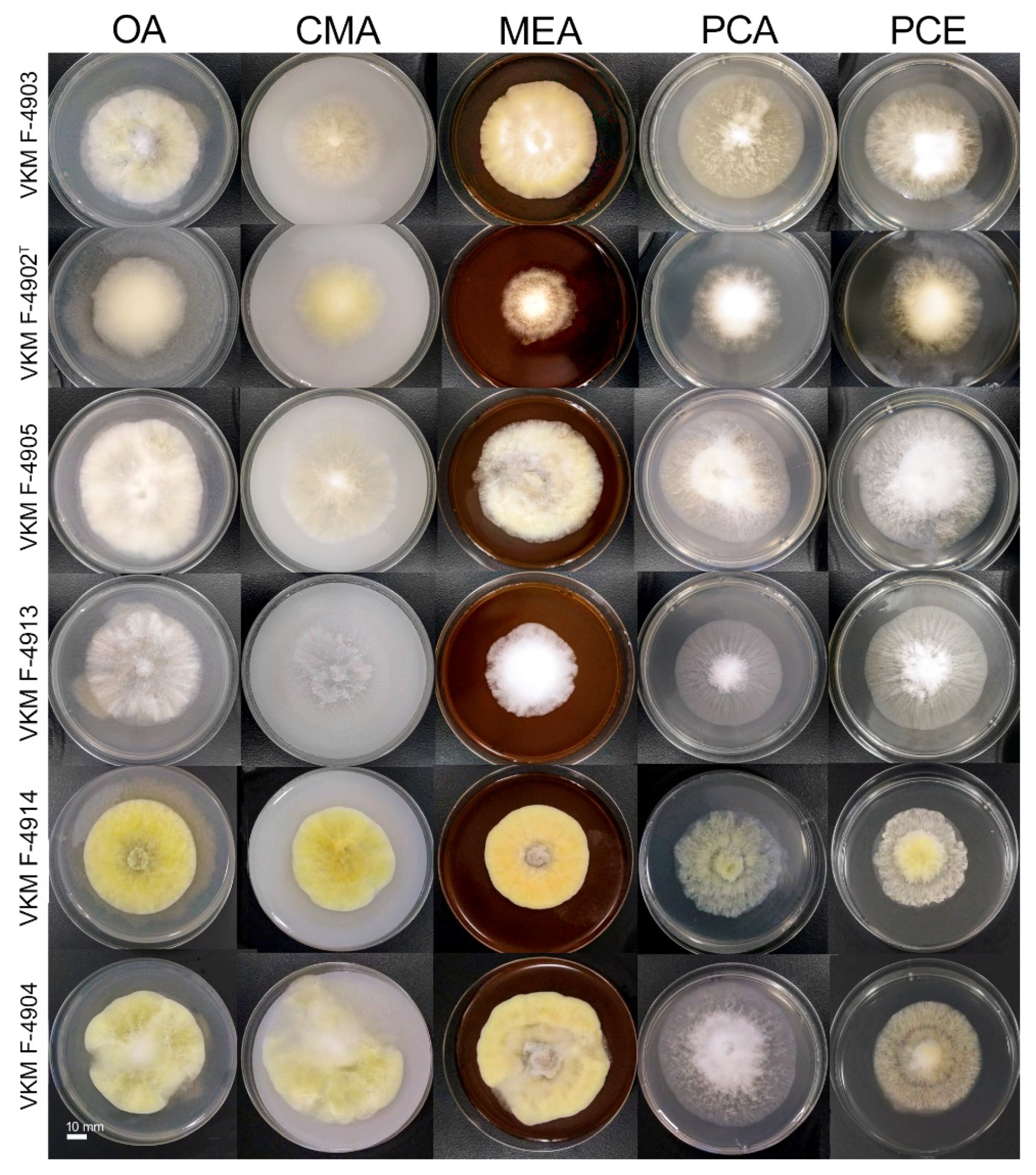
3.3. Taxonomy
4. Discussion
4.1. Relations and Features of the Species
4.2. Ecology
4.3. Monitoring of the Infection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant Pathogenic Fungi. Microbiol. Spectr. 2017, 5, 703–726. [Google Scholar] [CrossRef] [PubMed]
- Kapsa, J.S. Important Threats in Potato Production and Integrated Pathogen/Pest Management. Potato Res. 2008, 51, 385–401. [Google Scholar] [CrossRef]
- Fiers, M.; Edel-Hermann, V.; Chatot, C.; Le Hingrat, Y.; Alabouvette, C.; Steinberg, C. Potato Soil-Borne Diseases. A Review. Agron. Sustain. Dev. 2012, 32, 93–132. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal Rna Genes for Phylogenetics. In PCR Protocols; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Hambleton, S.; Nickerson, N.L.; Seifert, K.A. Leohumicola, a New Genus of Heat-Resistant Hyphomycetes. Stud. Mycol. 2005, 53, 29–52. [Google Scholar] [CrossRef]
- Schaad, N.W.; Frederick, R.D. Real-Time PCR and Its Application for Rapid Plant Disease Diagnostics. Can. J. Plant Pathol. 2002, 24, 250–258. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging Infectious Diseases of Plants: Pathogen Pollution, Climate Change and Agrotechnology Drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global Trends in Emerging Infectious Diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Von Arx, J.A.; Guarro, J.; Figueras, M.J. The Ascomycete Genus Chaetomium. Beih. Zur Nov. Hedwig. 1986, 43, 1–162. [Google Scholar]
- Wang, X.W.; Houbraken, J.; Groenewald, J.Z.; Meijer, M.; Andersen, B.; Nielsen, K.F.; Crous, P.W.; Samson, R.A. Diversity and Taxonomy of Chaetomium and Chaetomium-like Fungi from Indoor Environments. Stud. Mycol. 2016, 84, 145–224. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, W.; Cai, L. Polyphasic Characterisation of Chaetomium Species from Soil and Compost Revealed High Number of Undescribed Species. Fungal Biol. 2017, 121, 21–43. [Google Scholar] [CrossRef]
- Van den Brink, J.; Facun, K.; de Vries, M.; Stielow, J.B. Thermophilic Growth and Enzymatic Thermostability Are Polyphyletic Traits within Chaetomiaceae. Fungal Biol. 2015, 119, 1255–1266. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Yang, F.Y.; Meijer, M.; Kraak, B.; Sun, B.D.; Jiang, Y.L.; Wu, Y.M.; Bai, F.Y.; Seifert, K.A.; Crous, P.W.; et al. Redefining Humicola Sensu Stricto and Related Genera in the Chaetomiaceae. Stud. Mycol. 2019, 93, 65–153. [Google Scholar] [CrossRef] [PubMed]
- Ames, L.M. A Monograph of the Chaetomiaceae; U.S. Army, Research and Development; The University of Virginia: Charlottesville, VA, USA, 1963; Volume 2. [Google Scholar]
- Ramarajan, R.; Manohar, C.S. Biological Pretreatment and Bioconversion of Agricultural Wastes, Using Ligninolytic and Cellulolytic Fungal Consortia. Bioremediat. J. 2017, 21, 89–99. [Google Scholar] [CrossRef]
- Ashwini, C. A Review on Chaetomium globosum Is Versatile Weapons for Various Plant Pathogens. J. Pharmacogn. 2019, 8, 946–949. [Google Scholar]
- Goh, T.K.; Hyde, K.D. A Synopsis of Trichocladium Species, Based on the Literature. Fungal Divers. 1999, 2, 101–118. [Google Scholar]
- FAOstat. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 8 January 2022).
- Muhammad, A.F.; Naz, F.; Irshad, G. Important Fungal Diseases of Potato and Their Management—A Brief Review. Mycopath 2013, 11, 45–50. [Google Scholar]
- Oyesola, O.L.; Aworunse, O.S.; Oniha, M.I.; Obiazikwor, O.H.; Bello, O.; Atolagbe, O.M.; Sobowale, A.A.; Popoola, J.O.; Obembe, O.O. Impact and Management of Diseases of Solanum tuberosum. In Solanum Tuberosum—A Promising Crop for Starvation Problem; IntechOpen: London, UK, 2021; ISBN 978-1-83969-167-6. [Google Scholar]
- Guchi, E. Disease Management Practice on Potato (Solanum tuberosum L.) in Ethiopia. World J. Agric. Res. 2015, 3, 34–42. [Google Scholar] [CrossRef]
- Du, M.; Ren, X.; Sun, Q.; Wang, Y.; Zhang, R. Characterization of Fusarium spp. Causing Potato Dry Rot in China and Susceptibility Evaluation of Chinese Potato Germplasm to the Pathogen. Potato Res. 2012, 55, 175–184. [Google Scholar] [CrossRef]
- Kinealy, C.; Moran, G.; King, J.; Kinealy, C. (Eds.) The History of the Irish Famine; Routledge: London, UK, 2020; Volume 1, ISBN 9781315513898. [Google Scholar]
- Merlington, A.; Hanson, L.E.; Bayma, R.; Hildebrandt, K.; Steere, L.; Kirk, W.W. First Report of Fusarium proliferatum Causing Dry Rot in Michigan Commercial Potato (Solanum tuberosum) Production. Plant Dis. 2014, 98, 843. [Google Scholar] [CrossRef]
- Chudinova, E.; Platonov, V.; Elansky, S.; Alexandrova, A.; Kokaeva, L.; Krutyakov, Y. First Report of Ilyonectria crassa on Potato. J. Plant Pathol. 2019, 101, 1293–1294. [Google Scholar] [CrossRef]
- Derviş, S.; Özer, G.; Türkölmez, Ş. First Report of Neoscytalidium dimidiatum Causing Tuber Rot of Potato in Turkey. J. Plant Pathol. 2020, 102, 1295–1296. [Google Scholar] [CrossRef]
- Song, S.; Lu, Z.; Wang, J.; Zhu, J.; Gu, M.; Tang, Q.; Zhang, Z.; Wang, W.; Zhang, L.; Wang, B. First Report of a New Potato Disease Caused by Galactomyces candidum F12 in China. J. Integr. Agric. 2020, 19, 2470–2476. [Google Scholar] [CrossRef]
- Chudinova, E.M.; Elansky, S.N. First Report of Septotinia populiperda on Potato Tubers in Russia. J. Plant Pathol. 2021, 103, 665. [Google Scholar] [CrossRef]
- Riaz, M.; Akhtar, N.; Khan, S.N.; Shakeel, M.; Tahir, A. Neocosmospora rubicola: An Unrecorded Pathogen from Pakistan Causing Potato Stem Rot. Sarhad J. Agric. 2020, 36, 906–912. [Google Scholar] [CrossRef]
- Chudinova, E.M.; Vedmedenko, D.V.; Platonov, V.A.; Elansky, A.S.; Belosokhov, A.F.; Elansky, S.N. First Report of Potato Tuber Disease Caused by Acrostalagmus luteoalbus. J. Plant Pathol. 2022, 104. in print. [Google Scholar] [CrossRef]
- Crous, P.W.; Verkley, G.J.; Groenewald, J.Z.; Samson, R.A. Fungal Biodiversity. CBS Laboratory Manual Series 1. Centraalbureau Voor Schimmelcultures; Westerdijk Fungal Biodiversity Institute: Utrecht, The Netherlands, 2009; ISBN 9789070351779. [Google Scholar]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic RDNA ITS2 Types within a Monophyletic Lineage of the Fungus Fusarium Are Nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Groenewald, J.Z.; Nakashima, C.; Nishikawa, J.; Shin, H.D.; Park, J.H.; Jama, A.N.; Groenewald, M.; Braun, U.; Crous, P.W. Species Concepts in Cercospora: Spotting the Weeds among the Roses. Stud. Mycol. 2013, 75, 115–170. [Google Scholar] [CrossRef]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A Multi-Gene Phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of Localized Incongruence Using a Combinational Bootstrap Approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Miller, A.N.; Huhndorf, S.M. Multi-Gene Phylogenies Indicate Ascomal Wall Morphology Is a Better Predictor of Phylogenetic Relationships than Ascospore Morphology in the Sordariales (Ascomycota, Fungi). Mol. Phylogenet. Evol. 2005, 35, 60–75. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest V2. Progr. Distrib. by Author. 2004.
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree v. 1.3.1. Computer Program and Documentation Distributed by the Author.
- Dahlberg, K.R.; van Etten, J.L. Physiology and Biochemistry of Fungal Sporulation. Annu. Rev. Phytopathol. 1982, 20, 281–301. [Google Scholar] [CrossRef]
- Su, Y.Y.; Qi, Y.L.; Cai, L. Induction of Sporulation in Plant Pathogenic Fungi. Mycology 2012, 3, 195–200. [Google Scholar] [CrossRef]
- Tiwari, S.; Thakur, R.; Shankar, J. Role of Heat-Shock Proteins in Cellular Function and in the Biology of Fungi. Biotechnol. Res. Int. 2015, 2015, 132635. [Google Scholar] [CrossRef]
- Hutchinson, H.B.; Clayton, J. On the Decomposition of Cellulose by an Aerobic Organism (Spirochaeta cytophaga, N. SP.). J. Agric. Sci. 1919, 9, 143–172. [Google Scholar] [CrossRef]
- Bensch, K.; Groenewald, J.Z.; Dijksterhuis, J.; Starink-Willemse, M.; Andersen, B.; Summerell, B.A.; Shin, H.D.; Dugan, F.M.; Schroers, H.J.; Braun, U.; et al. Species and Ecological Diversity within the Cladosporium cladosporioides Complex (Davidiellaceae, Capnodiales). Stud. Mycol. 2010, 67, 1–94. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart. A Mycol. Colour Chart. 1970, 64, 230–233. [Google Scholar]
- Gherbawy, Y.A.; Gashgari, R.M. Mycobiota Associated with Superficial Blemishes of Potato Tubers. Food Biotechnol. 2013, 27, 137–151. [Google Scholar] [CrossRef]
- Hughes, S.J. An Undescribed Species of Chaetomium, with Four-Spored Asci. Trans. Br. Mycol. Soc. 1946, 29, 70–73. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant Cell Wall—Degrading Enzymes and Their Secretion in Plant-Pathogenic Fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Lombard, L.; Groenewald, J.Z.; Li, J.; Videira, S.I.R.; Samson, R.A.; Liu, X.Z.; Crous, P.W. Phylogenetic Reassessment of the Chaetomium globosum Species Complex. Pers. Mol. Phylogeny Evol. Fungi 2016, 36, 83–133. [Google Scholar] [CrossRef] [PubMed]
- Eltawil, M.A.; Samuel, D.V.K.; Singhal, O.P. Potato Storage Technology and Store Design Aspects. Agric. Eng. Int. CIGR Ejournal 2006, VIII, 1–18. [Google Scholar]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Lal, M.K.; Chourasia, K.N.; Kumar, D.; Kumar, M. Potato Dry Rot Disease: Current Status, Pathogenomics and Management. 3 Biotech 2020, 10, 503. [Google Scholar] [CrossRef]
- Wharton, P.; Willie, K. Fusarium Dry Rot Posing Problems in Potatoes—MSU Extension. Available online: https://www.canr.msu.edu/news/fusarium_dry_rot_posing_problems_in_potatoes (accessed on 8 January 2022).
- Wharton, P.; Hammerschmidt, R.; Kirk, W. Fusarium Dry Rot; Mich. State Univ. Ext. Bull. E-2995; Michigan State University: East Lansing, MI, USA, 2007. [Google Scholar]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a Node or a Foot-Shaped Basal Cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Jager, G.; Ten Hoopen, A.; Velvis, H. Hyperparasites of Rhizoctonia solani in Dutch Potato Fields. Neth. J. Plant Pathol. 1979, 85, 253–268. [Google Scholar] [CrossRef]
- Abou-Heilah, A.N. Postharvest Fungal Diseases of Some Vegetables in the Two Main Markets of Riyadh (Saudi Arabia). J. Univ. Kuwait 1985, 12, 103–112. [Google Scholar]
- Mazurkiewicz-Zapałowicz, K.; Wróbel, J.; Janowicz, K. Interaction between Golden Nematode Globodera rostochiensis (Woll.), Behrens and Soil Saprophytic Fungi—Its Influence on the Processes of Gaseous Exchange and Crop Yield of Potato (Solanum tuberosum L.): Interaktionen Zwischen Kartoffelnematoden Globod. Int. J. Phytoremediation 2002, 21, 279–286. [Google Scholar] [CrossRef]
- Kinga, M.Z.; Jacek, W.; Krystyna, J. Influence of Selected Soil Saprotrophes on Gas Exchange, Growth and Yield of Solanum Tuberosum. Acta Physiol. Plant. 2004, 26, 157–164. [Google Scholar] [CrossRef]
- Fiers, M.; Chatot, C.; Edel-Hermann, V.; Le Hingrat, Y.; Konate, A.Y.; Gautheron, N.; Guillery, E.; Alabouvette, C.; Steinberg, C. Diversity of Microorganisms Associated with Atypical Superficial Blemishes of Potato Tubers and Pathogenicity Assessment. Eur. J. Plant Pathol. 2010, 128, 353–371. [Google Scholar] [CrossRef]
- Treikale, O.; Vigule, Z.; Javoiša, B.; Grantiņa-Ieviņa, L.; Skrabule, I.; Plant, L.; Grantina-ievina, L.; Health, A.; Skrabule, I. Pathogenic Mycoflora of Potatoes during Storage. In Proceedings of the Zinātniski praktiskā konference ‘Līdzsvarota Lauksaimniecība’, Latvijas Lauksaimniecības universitātes Lauksaimniecības fakultātes Latvijas Agronomu biedrības un Latvijas Lauksaimniecības un meža zinātņu akadēmijas organizētās zinātniski praktiskās konferences, Jelgavā, Latvia, 19–20 February 2015; pp. 126–131. [Google Scholar]
- Golinski, P.; Wnuk, S.; Chełkowski, J.; Schollenberger, M. Formation of Avenacein Y by Fusarium avenaceum Fries Sacc. Isolates from Poland and Biological Properties of the Compound. Mycotoxin Res. 1987, 3, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Yeboah, S.; Xu, X.; Liu, Y.; Yu, B. Analysis on Fungal Diversity in Rhizosphere Soil of Continuous Cropping Potato Subjected to Different Furrow-Ridge Mulching Managements. Front. Microbiol. 2017, 8, 845. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Culture Accession Number | Origin | GeneBank Accession Numbers | ||
|---|---|---|---|---|---|
| ITS | rpb2 | tub2 | |||
| Trichocladium solani | VKM F-4903 | Air from potato storage facility, Moscow region, Russia. July 2017 | OL691126 | OL314249 | OL314243 |
| VKM F-4902T | Potato tubers, cultivar ‘Djura’ with symptoms of yellow rot. Moscow region, Russia. March 2018 | OL691125 | OM974683 | OL314244 | |
| VKM F-4913 | Potato tubers, cultivar ‘Gala’ with symptoms of yellow rot. Moscow region, Russia. March 2018 | OL691123 | OL314251 | OL314246 | |
| VKM F-4904 | Potato tubers, cultivar ‘Romano’ with symptoms of yellow rot. Kaluga region, Russia. May 2018 | OL691122 | OM974684 | OL314247 | |
| VKM F-4905 | Potato tubers, cultivar ‘Gala’ with symptoms of yellow rot. Moscow region, Russia. September 2018 | OL691124 | OL314250 | OL314245 | |
| VKM F-4914 | Potato tubers, cultivar ‘La Strada’ with symptoms of yellow rot. Moscow region, Russia. November 2019 | OL691121 | OL314252 | OL314248 | |
| VKM F-4915 | Potato tubers of cultivar ‘Gala’ with symptoms of yellow rot. Moscow Region. March 2020 | OL638311 | OL442086 | OL674250 | |
| Botryotrichum verrucosum | CBS 116.64 T | Salt marsh soil, mature dunes, UK. January 1962 | LT993567 | LT993486 | LT993648 |
| Humicola ampulliella | CBS 116735 T | Discarded sock, China. August 2013 | LT993568 | LT993487 | LT993649 |
| Humicola atrobrunnea | HSAUPII05-1004 T | Soil, China, Guizhou | LT993570 | LT993489 | LT993651 |
| Humicola christensenii | CBS 127760 T | Soil, USA, Minnesota. 1988 | LT993571 | LT993490 | LT993652 |
| Humicolacuyabenoensis | CBS 398.97 T | Rain forest, Ecuador. March 1993 | LT993573 | LT993492 | LT993654 |
| Humicola degenerans | CBS 232.65 T | Soil under mixed forest, Canada, Ontario. August 1964 | LT993574 | LT993493 | LT993655 |
| Humicola distorta | CBS 417.66 T | Populus tremuloides dead leaf, USA, Iowa | LT993577 | LT993496 | LT993658 |
| Humicola floriformis | CBS 815.97 T | Fallen leaves, Thailand, Sukhothai | LT993578 | LT993497 | LT993659 |
| Humicola fuscoatra | CBS 118.14 T | Soil, Norway. 1914 | LT993579 | LT993498 | LT993660 |
| Humicola fuscogrisea | CGMCC 3.13790 T | Soil, China, Shennongjia | LT993581 | LT993500 | LT993662 |
| Humicola homopilata | CBS 157.55 T | Filter paper in soil, Norway | LT993582 | LT993501 | LT993663 |
| Humicola leptodermospora | CBS 120095 T | Forestal soil, Brazil | LT993584 | LT993503 | LT993665 |
| Humicola malaysiensis | CBS 399.97 | Elaeis guineensi, Mylaysia, Selangor. March 1974 | LT993586 | LT993505 | LT993667 |
| Humicola mutabilis | CBS 779.71 T | Soil, Israel | LT993588 | LT993507 | LT993669 |
| Humicola olivacea | CBS 142031 T | Dust, USA | LT993589 | LT993508 | LT993670 |
| Humicola pinnata | CBS 467.66 T | Dead wood, USA, Coronado National Forest | LT993590 | LT993509 | LT993671 |
| Humicola pulvericola | CBS 144165 T | Dust, Mexico | LT993591 | LT993510 | LT993672 |
| Humicola quadrangulata | CBS 111771 T | Soil, Brazil | LT993593 | LT993512 | LT993674 |
| Humicola seminuda | CBS 368.84 eT | Soil, Canada, Ontario. April 1981 | LT993594 | LT993513 | LT993675 |
| Humicola semispiralis | CBS 723.97 eT | Paper, Canada, Toronto | LT993597 | LT993516 | LT993678 |
| Humicola sphaeralis | CBS 985.87 eT | Soil, France | LT993598 | LT993517 | LT993679 |
| Humicola subspiralis | CBS 148.58 | Leaf fragments in soil, China | LT993599 | LT993518 | LT993680 |
| Humicola wallefii | CBS 147.67 T | Soil, Zaire | LT993602 | LT993521 | LT993683 |
| Mycothermus thermophiloides | CBS 183.81 T | Soil, USA, Indiana | LT993603 | LT993522 | LT993684 |
| Mycothermus thermophilum | CBS 625.91 T | Chicken nest straw, USA, Nevada | LT993604 | LT993523 | LT993685 |
| Remersonia tenuis | CBS 784.85 T | Dung of horse, India | LT993609 | LT993528 | LT993690 |
| Remersonia thermophila | CBS 645.91 | Compost, Netherlands | LT993611 | LT993530 | LT993692 |
| Staphylotrichum acaciicola | CBS 281.65 T | Acacia karroo leaf litter, South Africa | LT993613 | LT993532 | LT993694 |
| Staphylotrichum boninense | CBS 112543 | Leaf litter, Brazil | LT993617 | LT993536 | LT993698 |
| Staphylotrichum brevistipitatum | CBS 408.67 T | Eucalyptus leaf litter, South Africa | LT993619 | LT993538 | LT993700 |
| Staphylotrichum coccosporum | CBS 364.58 T | Soil, Zaire | LT993620 | LT993539 | LT993701 |
| Staphylotrichum microascosporum | CBS 184.79 T | Soil from Mangifera orchard, Sudan | LT993624 | LT993543 | LT993705 |
| Staphylotrichum tortipilum | CBS 103.79 T | Dung of pine vole, USA, North Carolina | LT993625 | LT993544 | LT993706 |
| Trichocladium acropullum | CBS 114580 T | Soil, China | LT993626 | LT993545 | LT993707 |
| CBS 127763 T | Greenhouse Soil, USA | LT993628 | LT993547 | LT993709 | |
| Trichocladium antarcticum | CBS 135876 | Usnea cf. aurantio-atra, Antarctica | LT993630 | LT993549 | LT993711 |
| CBS 123565 T | Usnea cf. aurantio-atra, Antarctica | LT993629 | LT993548 | LT993710 | |
| Trichocladium arxii | CBS 104.79 T | Dung of kangaroo rat, USA, California | LT993631 | LT993550 | LT993712 |
| Trichocladium asperum | CBS 903.85 eT | Acidic soil, Germany | LT993632 | LT993551 | LT993713 |
| CBS 140.21 | Unknown substrate, Netherlands | LT993633 | LT993552 | LT993714 | |
| CBS 157.22 | Unknown substrate, unknown country | LT993634 | LT993553 | LT993715 | |
| Trichocladium beniowskiae | CBS 757.74 T | Grass, India | LT993635 | LT993554 | LT993716 |
| Trichocladium crispatum | CBS 149.58 | Estuarine sediment, Germany | LT993636 | LT993555 | LT993717 |
| CBS 693.71 | Agricultural soil, Netherlands | LT993637 | LT993556 | LT993718 | |
| Trichocladium gilmaniellae | CBS 388.75 T | Salt marsh soil, Kuwait | LT993638 | LT993557 | LT993719 |
| Trichocladium griseum | CBS 119.14 NeoT | Soil, Norway | LT993639 | LT993558 | LT993720 |
| CBS 217.34 | Unknown substrate, Germany | LT993640 | LT993559 | LT993721 | |
| CGMCC 3.13888 | Soil, China, Jilin | LT993641 | LT993560 | LT993722 | |
| Trichocladium jilongensis | HSAUPII07 1485 T | Mountain soil, China, Tibet | LT993642 | LT993561 | LT993723 |
| CBS 195.87 | Field soil, Germany | LT993643 | LT993562 | LT993724 | |
| Trichocladium nigrospermum | CBS 103.36 | Meal, Netherlands | LT993644 | LT993563 | LT993725 |
| Trichocladium seminis-citrulli | CBS 143.58 T | Dung of fox, Turkmenistan | LT993645 | LT993564 | LT993726 |
| CBS 637.83 | Dung of goat, Israel | LT993646 | LT993565 | LT993727 | |
| Trichocladium sp. | CBS 351.77 | Wheat field soil, Germany | LT993647 | LT993566 | LT993728 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belosokhov, A.; Yarmeeva, M.; Kokaeva, L.; Chudinova, E.; Mislavskiy, S.; Elansky, S. Trichocladium solani sp. nov.—A New Pathogen on Potato Tubers Causing Yellow Rot. J. Fungi 2022, 8, 1160. https://doi.org/10.3390/jof8111160
Belosokhov A, Yarmeeva M, Kokaeva L, Chudinova E, Mislavskiy S, Elansky S. Trichocladium solani sp. nov.—A New Pathogen on Potato Tubers Causing Yellow Rot. Journal of Fungi. 2022; 8(11):1160. https://doi.org/10.3390/jof8111160
Chicago/Turabian StyleBelosokhov, Arseniy, Maria Yarmeeva, Lyudmila Kokaeva, Elena Chudinova, Svyatoslav Mislavskiy, and Sergey Elansky. 2022. "Trichocladium solani sp. nov.—A New Pathogen on Potato Tubers Causing Yellow Rot" Journal of Fungi 8, no. 11: 1160. https://doi.org/10.3390/jof8111160
APA StyleBelosokhov, A., Yarmeeva, M., Kokaeva, L., Chudinova, E., Mislavskiy, S., & Elansky, S. (2022). Trichocladium solani sp. nov.—A New Pathogen on Potato Tubers Causing Yellow Rot. Journal of Fungi, 8(11), 1160. https://doi.org/10.3390/jof8111160