
Pseudophylloporus Gen. nov. and Rubroleccinum Gen. nov., Two New Genera Revealed by Morphological and Phylogenetic Evidences in the Family Boletaceae from Subtropical China

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. Molecular Procedures
2.3. Dataset Assembly
2.4. Phylogenetic Analyses
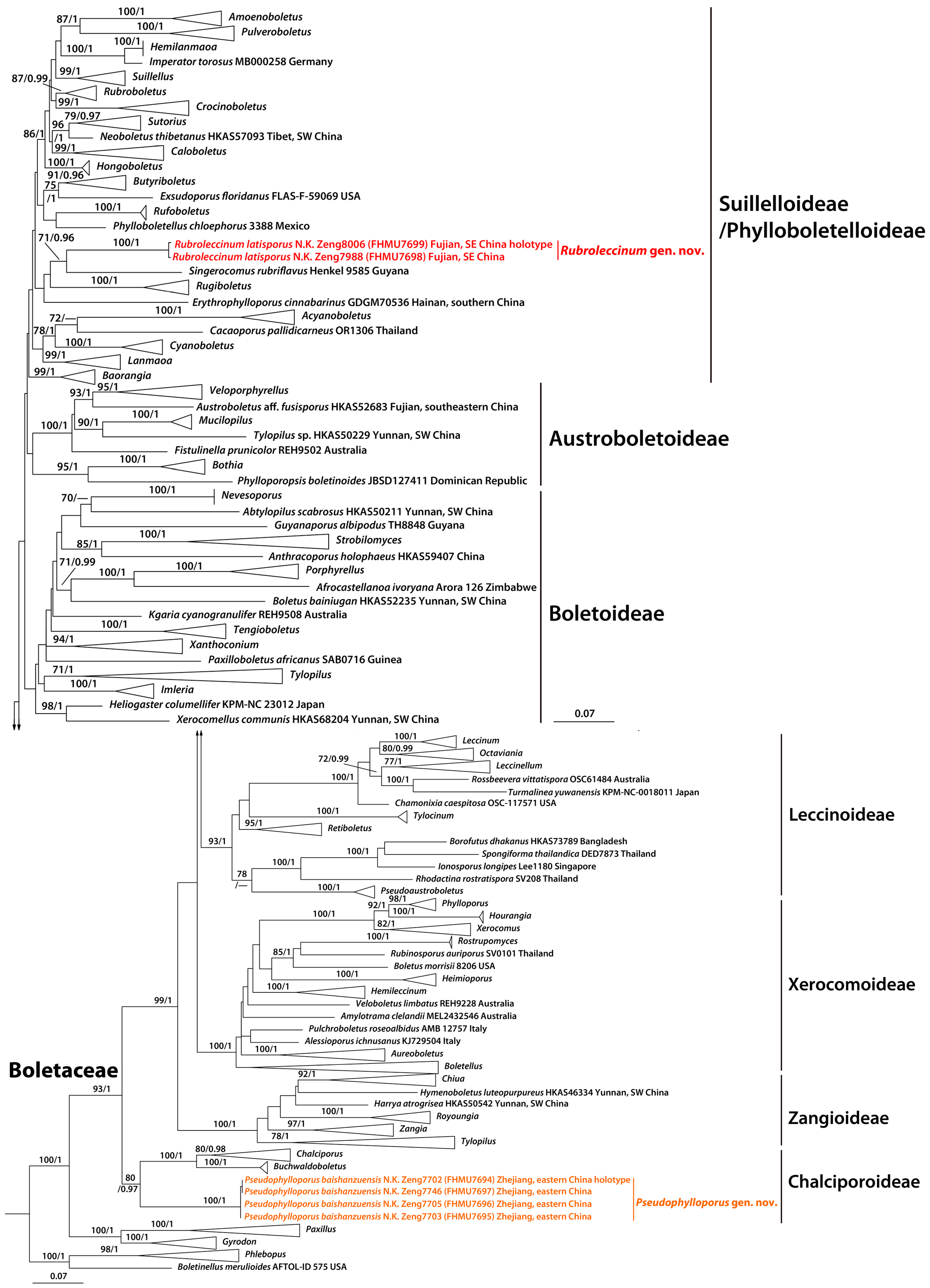
3. Results
3.1. Molecular Data
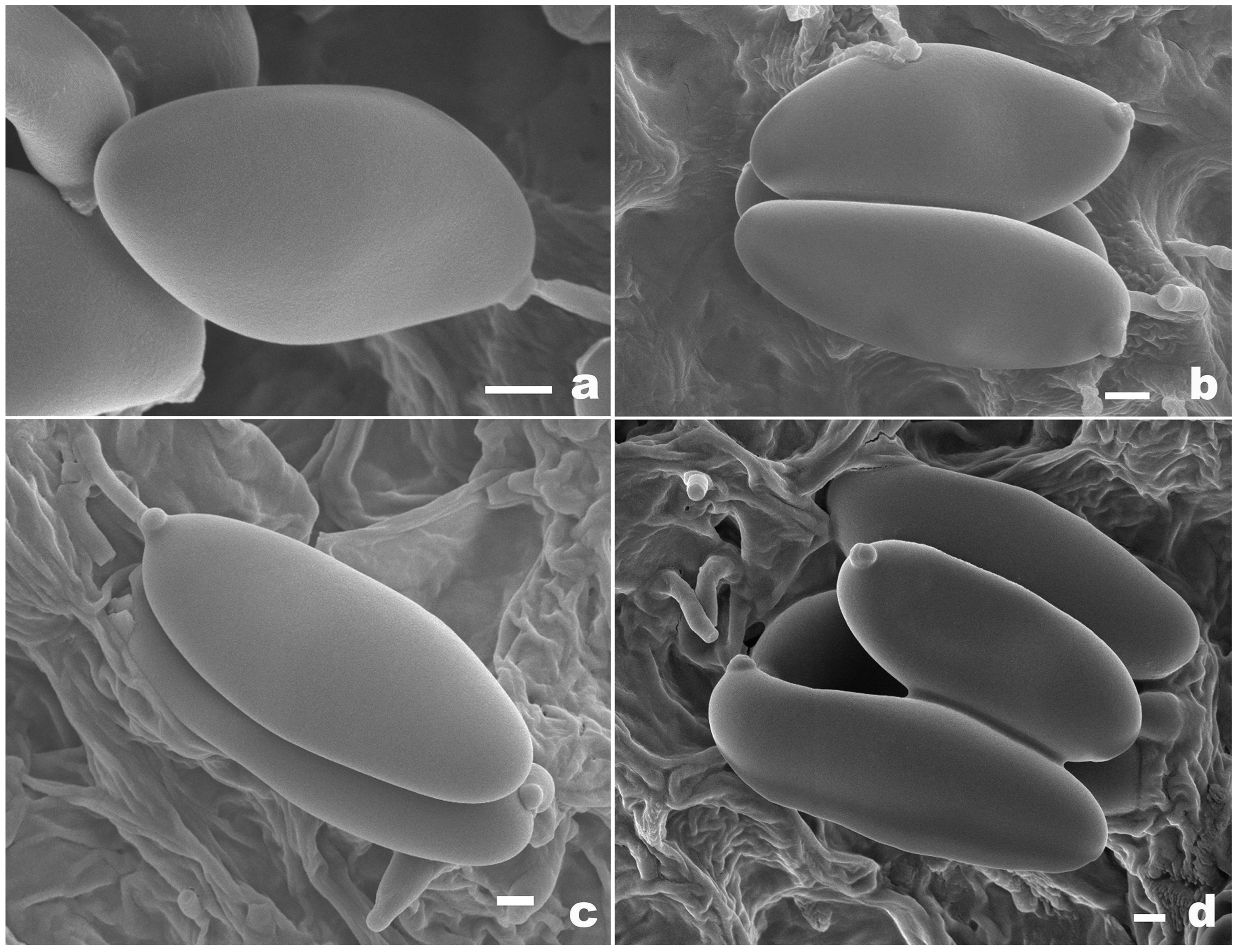
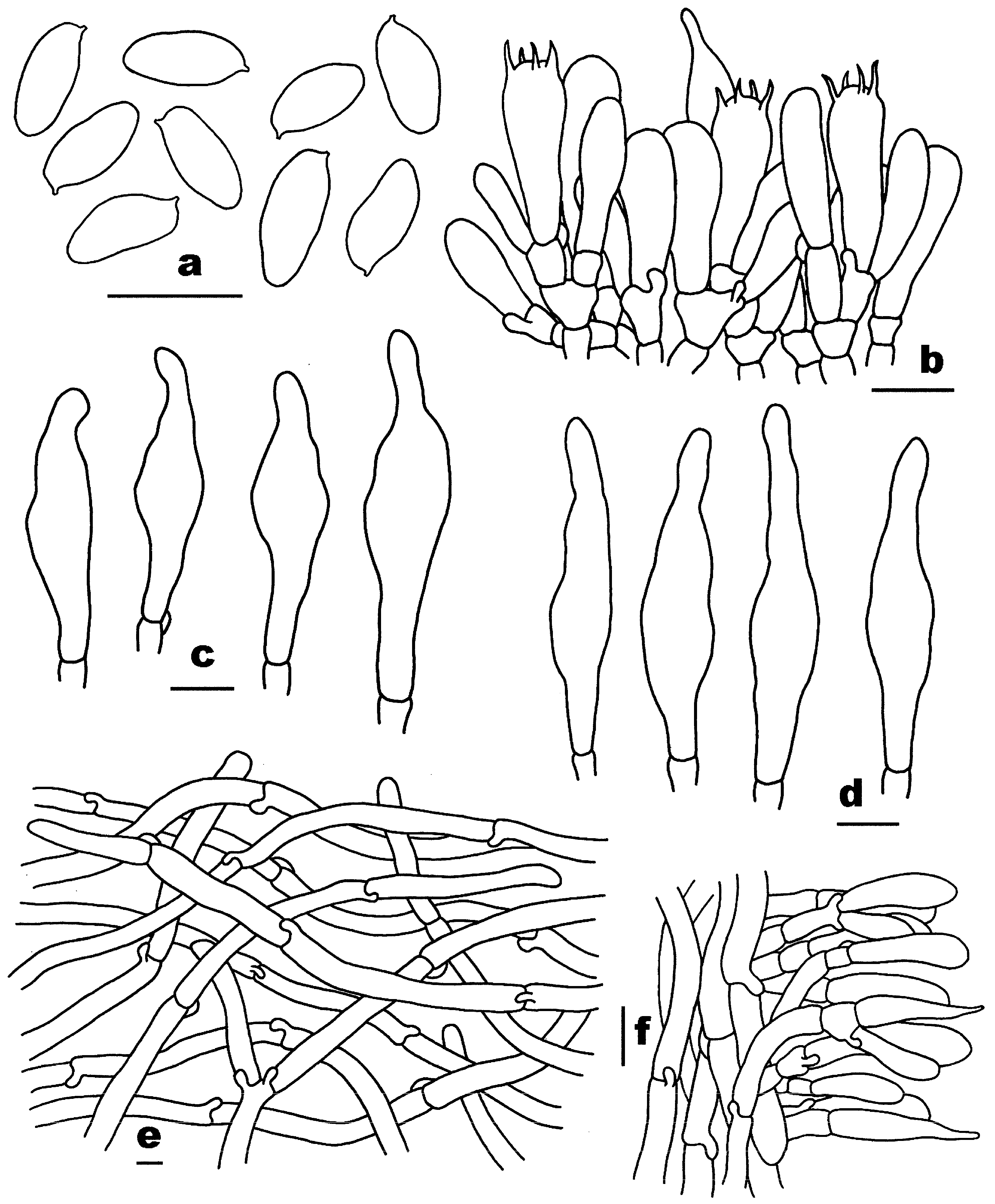
3.2. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Wang, L.Y.; Dai, D.; Qi, Z.X.; Zhang, Z.H.; Liu, Y.J.; Hu, J.J.; Zhang, P.; Li, Y.; Zhang, B. Boletaceae in China: Taxonomy and phylogeny reveal a new genus, two new species, and a new record. Front. Microbiol. 2023, 13, 1052948. [Google Scholar] [CrossRef] [PubMed]
- Gelardi, M.; Angelini, C.; Biketova, A.Y.; Ercole, E.; Svetasheva, T.Y.; Miller, K.O.; de la Fuente, J.I.; García-Jiménez, J.; Vizzini, A. Coccoloba-associated xerocomoid boletes (Boletaceae) from the Caribbean and Mexico: Tropicoboletus ruborculus gen. et comb. nov., revision of Xerocomus coccolobae, phylogenetic assessment of Singerocomus guadelupae comb. nov., and type studies of Xerocomus caeruleonigrescens, X. cuneipes, and X. pseudoboletinus var. pini-caribaeae. Mycol. Prog. 2023, 22, 29. [Google Scholar] [CrossRef]
- Wu, G.; Li, H.J.; Horak, E.; Wu, K.; Li, G.M.; Yang, Z.L. New taxa of Boletaceae from China. Mycosphere 2023, 14, 754–776. [Google Scholar] [CrossRef]
- Xue, R.; Zhang, X.; Xu, C.; Xie, H.J.; Wu, L.L.; Wang, Y.; Tang, L.P.; Hao, Y.J.; Zhao, K.; Jiang, S.; et al. The subfamily Xerocomoideae (Boletaceae, Boletales) in China. Stud. Mycol. 2023, 106, 95–197. [Google Scholar] [CrossRef]
- Neves, M.A.; Binder, M.; Halling, R.; Hibbett, D.; Soytong, K. The phylogeny of selected Phylloporus species, inferred from NUC-LSU and ITS sequences, and descriptions of new species from the old world. Fungal Divers. 2012, 55, 109–123. [Google Scholar] [CrossRef]
- Arora, D.; Frank, J.L. Clarifying the butter Boletes: A new genus, Butyriboletus, is established to accommodate Boletus sect. Appendiculati, and six new species are described. Mycologia 2014, 106, 464–480. [Google Scholar] [CrossRef]
- Henkel, T.W.; Obase, K.; Husbands, D.; Uehling, J.K.; Bonito, G.; Aime, M.C.; Smith, M.E. New Boletaceae taxa from Guyana: Bnderoboletus segoi gen. and sp. nov., Guyanaporus albipodus gen. and sp. nov., Singerocomus rubriflavus gen. and sp. nov., and a new combination for Xerocomus inundabilis. Mycologia 2016, 108, 157–173. [Google Scholar] [CrossRef]
- Chai, H.; Liang, Z.Q.; Xue, R.; Jiang, S.; Luo, S.H.; Wang, Y.; Wu, L.L.; Tang, L.P.; Chen, Y.; Hong, D.; et al. New and noteworthy boletes from subtropical and tropical China. MycoKeys 2019, 46, 55. [Google Scholar] [CrossRef]
- Deng, H.; Wang, Y.; Lei, J.R.; Chen, Z.Z.; Liang, Z.Q.; Zeng, N.K. Four new species of Strobilomyces (Boletaceae, Boletales) from Hainan island, tropical China. J. Fungi 2023, 9, 1128. [Google Scholar] [CrossRef]
- Vadthanarat, S.; Lumyong, S.; Raspé, O. Cacaoporus, a new Boletaceae genus, with two new species from Thailand. MycoKeys 2019, 54, 1–29. [Google Scholar] [CrossRef]
- Magnago, A.C.; Alves-Silva, G.; Henkel, T.W.; da Silveira, R.M.B. New genera, species, and combinations of Boletaceae from Brazil and Guyana. Mycologia 2022, 114, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Farid, A.; Gelardi, M.; Angelini, C.; Franck, A.R.; Costanzo, F.; Kaminsky, L.; Ercole, E.; Baroni, T.J.; White, A.L.; Garey, J.R.; et al. Phylloporus and Phylloboletellus are no longer alone: Phylloporopsis gen. nov. (Boletaceae), a new smooth-spored lamellate genus to accommodate the American species Phylloporus boletinoides. Fungal Syst. Evol. 2018, 2, 341–359. [Google Scholar] [CrossRef] [PubMed]
- Badou, S.A.; Furneaux, B.; De Kesel, A.; Khan, F.K.; Houdanon, R.D.; Ryberg, M.; Yorou, N.S. Paxilloboletus gen. nov., a new lamellate bolete genus from tropical Africa. Mycol. Prog. 2022, 21, 243–256. [Google Scholar] [CrossRef]
- Ayala-Vásquez, O.; Pérez-Moreno, J.; Pinzón, J.P.; Garibay-Orijel, R.; García-Jiménez, J.; de la Fuente, J.I.; Venegas-Barrera, C.S.; Martínez-Reyes, M.; Montoya, L.; Bandala, V.; et al. Broadening the knowledge of Mexican boletes: Addition of a new genus, seven new species, and three new combinations. J. Fungi 2023, 9, 1126. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Feng, B.; Xu, J.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Tremble, K.; Henkel, T.; Bradshaw, A.; Domnauer, C.; Brown, L.M.; Thám, L.X.; Furci, G.; Aime, M.C.; Moncalvo, J.M.; Dentinger, B. A revised phylogeny of Boletaceae using whole genome sequences. Mycologia 2024, 116, 392–408. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, CT, USA, 2008. [Google Scholar]
- Wu, G.; Li, M.X.; Horak, E.; Yang, Z.L. Phylogenetic analysis reveals the new genus Amoenoboletus from Asia and New Zealand. Mycologia 2022, 114, 144–156. [Google Scholar] [CrossRef]
- Becerra, A.G.; Zak, M.R. The ectomycorrhizal symbiosis in South America: Morphology, colonization, and diversity. In Diversity and Biotechnology of Ectomycorrhizae; Springer: Berlin/Heidelberg, Germany, 2011; pp. 19–41. [Google Scholar] [CrossRef]
- Zeng, N.K.; Tang, L.P.; Li, Y.C.; Tolgor, B.; Zhu, X.T.; Zhao, Q.; Yang, Z.L. The genus Phylloporus (Boletaceae, Boletales) from China: Morphological and multilocus DNA sequence analyses. Fungal Divers. 2013, 58, 73–101. [Google Scholar] [CrossRef]
- Pérez-Moreno, J.; Guerin-Laguette, A.; Flores, A.R.; Qiang, F. Mushrooms, Humans and Nature in a Changing World; Springer Nature: Basel, Switzerland, 2020; p. 473. [Google Scholar] [CrossRef]
- Pérez-Moreno, J.; Guerin-Laguette, A.; Rinaldi, A.C.; Yu, F.; Verbeken, A.; Hernández-Santiago, F.; Martínez Reyes, M. Edible mycorrhizal fungi of the world: What is their role in forest sustainability, food security, biocultural conservation and climate change? Plants People Planet 2020, 3, 471–490. [Google Scholar] [CrossRef]
- Husbands, D.R.; Henkel, T.W.; Bonito, G.; Vilgalys, R.; Smith, M.E. New species of Xerocomus (Boletales) from the Guiana Shield, with notes on their mycorrhizal status and fruiting occurrence. Mycologia 2013, 105, 422–435. [Google Scholar] [CrossRef]
- Yang, H.; Yang, Z.; Wang, Q.C.; Wang, Y.L.; Hu, H.W.; He, J.Z.; Zheng, Y.; Yang, Y. Compartment and plant identity shape tree mycobiome in a subtropical forest. Microbiol. Spectr. 2022, 10, e0134722. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, J.; Deng, T.; Boufford, D.E. Origins and evolution of plant diversity in the Hengduan Mountains, China. Plant Divers. 2017, 39, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben, 3rd ed.; Muster-Schmidt Verlag: Göttingen, Germany, 1981; p. 242. [Google Scholar]
- Zeng, N.K.; Wu, G.; Li, Y.C.; Liang, Z.Q.; Yang, Z.L. Crocinoboletus, a new genus of Boletaceae (Boletales) with unusual boletocrocin polyene pigments. Phytotaxa 2014, 175, 133–140. [Google Scholar] [CrossRef]
- Zhang, X.; Tian, R.; Tang, L.P.; Liang, Z.Q.; Zhang, W.H.; Jiang, S.; Wang, C.K.; Zeng, N.K. Morphological and phylogenetic evidence reveal three new species of Aureoboletus (Boletaceae, Boletales) from China. Phytotaxa 2022, 567, 127–148. [Google Scholar] [CrossRef]
- Zeng, N.K.; Liang, Z.Q.; Tang, L.P.; Li, Y.C.; Yang, Z.L. The genus Pulveroboletus (Boletaceae, Boletales) in China. Mycologia 2017, 109, 422–442. [Google Scholar] [CrossRef]
- Xu, C.; Liang, Z.Q.; Su, M.S.; Jiang, S.; Chen, Y.; Fan, Y.G.; Zeng, N.K. Austroboletus brunneisquamus (Boletaceae, Boletales), a new ectomycorrhizal fungus from a tropical rainforest, China. Forests 2021, 12, 1438. [Google Scholar] [CrossRef]
- Xie, H.J.; Tang, L.P.; Mu, M.; Fan, Y.G.; Jiang, S.; Su, M.S.; Liang, Z.Q.; Zeng, N.K. A contribution to knowledge of Gyroporus (Gyroporaceae, Boletales) in China: Three new taxa, two previous species, and one ambiguous taxon. Mycol. Prog. 2022, 21, 71–92. [Google Scholar] [CrossRef]
- Bas, C. Morphology and subdivision of Amanita and a monograph of its section Lepidella. Persoonia 1969, 5, 285–573. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- James, T.Y.; Kauff, F.; Schoch, C.; Matheny, P.B.; Hofstetter, V.; Cox, C.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of the fungi using a six gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol. Phylogenetics Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analyses program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.A.; Dunn, C.W. Phyutility: A phyloinformatics tool for trees, alignments andmolecular data. Bioinformatics 2008, 24, 715–716. [Google Scholar] [CrossRef]
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Han, L.H.; Cui, Y.Y.; Li, F.; Xu, J.P.; Yang, Z.L. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Orihara, T.; Smith, M.E. Unique phylogenetic position of the African truffle-like fungus, Octaviania ivoryana (Boletaceae, Boletales), and the proposal of a new genus, Afrocastellanoa. Mycologia 2017, 109, 323–332. [Google Scholar] [CrossRef]
- Gelardi, M.; Simonini, G.; Ercole, E.; Vizzini, A. Alessioporus and Pulchroboletus (Boletaceae, Boletineae), two novel genera for Xerocomus ichnusanus and X. roseoalbidus from the European Mediterranean basin: Molecular and morphological evidence. Mycologia 2014, 106, 1168–1187. [Google Scholar] [CrossRef]
- Lebel, T.; Davoodian, N.; Bloomfield, M.; Syme, K.; May, T.W.; Hosaka, K.; Castellano, M.A. A mixed bag of sequestrate fungi from five different families: Boletaceae, Russulaceae, Psathyrellaceae, Strophariaceae, and Hysterangiaceae. Swainsona 2022, 36, 33–65. [Google Scholar]
- Nuhn, M.E.; Binder, M.; Taylor, A.F.; Halling, R.E.; Hibbett, D.S. Phylogenetic overview of the Boletineae. Fungal Biol. 2013, 117, 479–511. [Google Scholar] [CrossRef]
- Wu, G.; Zhao, K.; Li, Y.C.; Zeng, N.K.; Feng, B.; Halling, R.E.; Yang, Z.L. Four new genera of the fungal family Boletaceae. Fungal Divers. 2015, 81, 1–24. [Google Scholar] [CrossRef]
- Nuhn, M.E. Molecular ecology of Boletinellus merulioides and systematics of the Boletineae; Clark University ProQuest Dissertations Publishing: Worcester, MA, USA, 2016. [Google Scholar]
- Hosen, M.; Feng, B.; Wu, G.; Zhu, X.T.; Li, Y.C.; Yang, Z.L. Borofutus, a new genus of Boletaceae from tropical Asia: Phylogeny, morphology and taxonomy. Fungal Divers. 2013, 58, 215–226. [Google Scholar] [CrossRef]
- Halling, R.E.; Baroni, T.J.; Binder, M. A new genus of Boletaceae from eastern North America. Mycologia 2007, 99, 310–316. [Google Scholar] [CrossRef]
- Zeng, N.K.; Su, M.S.; Liang, Z.Q.; Yang, Z.L. A geographical extension of the North American genus Bothia (Boletaceae, Boletales) to East Asia with a new species B. fujianensis from China. Mycol. Prog. 2015, 14, 1015. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, G.; Feng, B.; Yang, Z.L. Molecular phylogeny of Caloboletus (Boletaceae) and a new species in East Asia. Mycol. Prog. 2014, 13, 1127–1136. [Google Scholar] [CrossRef]
- Kuo, M.; Ortiz-Santana, B. Revision of leccinoid fungi, with emphasis on North American taxa, based on molecular and morphological data. Mycologia 2020, 112, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, T.H. Erythrophylloporus (Boletaceae, Boletales), a new genus inferred from morphological and molecular data from subtropical and tropical China. Mycosystema 2018, 37, 1111–1126. [Google Scholar] [CrossRef]
- Biketova, A.Y.; Gelardi, M.; Smith, M.E.; Simonini, G.; Healy, R.A.; Taneyama, Y.; Vasquez, G.; Kovács, Á.; Nagy, L.G.; Wasser, S.P.; et al. Reappraisal of the genus Exsudoporus (Boletaceae) worldwide based on multi-gene phylogeny, morphology and biogeography, and insights on Amoenoboletus. J. Fungi 2022, 8, 101. [Google Scholar] [CrossRef]
- Halling, R.E.; Nuhn, M.; Osmundson, T.; Fechner, N.; Trappe, J.M.; Soytong, K.; Arora, D.; Hibbett, D.S.; Binder, M. Affinities of the Boletus chromapes group to Royoungia and the description of two new genera, Harrya and Australopilus. Aust. Syst. Bot. 2012, 25, 418–431. [Google Scholar] [CrossRef]
- Vadthanarat, S.; Raspé, O.; Lumyong, S. Phylogenetic affinities of the sequestrate genus Rhodactina (Boletaceae), with a new species, R. rostratispora from Thailand. MycoKeys 2018, 29, 63. [Google Scholar] [CrossRef]
- Dentinger, B.T.; Ammirati, J.F.; Both, E.E.; Desjardin, D.E.; Halling, R.E.; Henkel, T.W.; Moreau, P.A.; Nagasawa, E.; Soytong, K.; Taylor, A.F.; et al. Molecular phylogenetics of porcini mushrooms (Boletus section Boletus). Mol. Phylogenet. Evol. 2010, 57, 1276–1292. [Google Scholar] [CrossRef]
- Binder, M.; Larsson, K.H.; Matheny, P.B.; Hibbett, D.S. Amylocorticiales ord. nov. and Jaapiales ord. nov.: Early diverging clades of agaricomycetidae dominated by corticioid forms. Mycologia 2010, 102, 865–880. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.T.; Li, Y.C.; Wu, G.; Feng, B.; Zhao, K.; Gelardi, M.; Kost, G.W.; Yang, Z.L. The genus Imleria (Boletaceae) in East Asia. Phytotaxa 2014, 191, 81–98. [Google Scholar] [CrossRef]
- Chuankid, B.; Vadthanarat, S.; Thongbai, B.; Stadler, M.; Lumyong, S.; Hyde, K.D.; Raspe, O. Retiboletus (Boletaceae) in northern Thailand: One novel species and two first records. Mycoscience 2021, 62, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Khmelnitsky, O.; Davoodian, N.; Singh, P.; Raspé, O.; Lee, S.M.; Fechner, N.; Bonito, G.; Lebel, T. Ionosporus: A new genus for Boletus longipes (Boletaceae), with a new species, I. australis, from Australia. Mycol. Prog. 2019, 18, 439–451. [Google Scholar] [CrossRef]
- Halling, R.E.; Fechner, N.A.; Holmes, G.; Davoodian, N. Kgaria (Boletaceae, Boletoideae) gen. nov. in Australia: Neither a Tylopilus nor a Porphyrellus. Fungal Syst. Evol. 2023, 12, 31–45. [Google Scholar] [CrossRef]
- Orihara, T.; Smith, M.; Shimomura, N.; Iwase, K.; Maekawa, N. Diversity and systematics of the sequestrate genus Octaviania in Japan: Two new subgenera and eleven new species. Persoonia 2012, 28, 85–112. [Google Scholar] [CrossRef]
- Jargeat, P.; Chaumeton, J.P.; Navaud, O.; Vizzini, A.; Gryta, H. The Paxillus involutus (Boletales, Paxillaceae) complex in Europe: Genetic diversity and morphological description of the new species Paxillus cuprinus, typification of P. involutus s.s., and synthesis of species boundaries. Fungal Biol. 2014, 118, 12–31. [Google Scholar] [CrossRef]
- Binder, M.; Bresinsky, A. Derivation of a polymorphic lineage of Gasteromycetes from boletoid ancestors. Mycologia 2002, 94, 85–98. [Google Scholar] [CrossRef]
- Xie, H.J.; Zhang, C.X.; He, M.X.; Liang, Z.Q.; Deng, X.H.; Zeng, N.K. Buchwaldoboletus xylophilus and Phlebopus portentosus, two non-ectomycorrhizal boletes from tropical China. Phytotaxa 2021, 520, 137–154. [Google Scholar] [CrossRef]
- Binder, M.; Hibbett, D.S. Molecular systematics and biological diversification of Boletales. Mycologia 2006, 98, 971–981. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, F.; Zeng, N.K.; Cui, Y.Y.; Yang, Z.L. A new genus Pseudoaustroboletus (Boletaceae, Boletales) from Asia as inferred from molecular and morphological data. Mycol. Prog. 2014, 13, 1207–1216. [Google Scholar] [CrossRef]
- Li, Y.C.; Yang, Z.L. The Boletes of China: Tylopilus s.l.; Science Press: Beijing, China, 2021. [Google Scholar] [CrossRef]
- Vadthanarat, S.; Raghoonundon, B.; Lumyong, S.; Raspé, O. Rostrupomyces, a new genus to accommodate Xerocomussisongkhramensis, and a new Hemileccinum species (Xerocomoideae, Boletaceae) from Thailand. MycoKeys 2024, 103, 129–165. [Google Scholar] [CrossRef] [PubMed]
- Raspe, O.; Vadthanarat, S.; De Kesel, A.; Degreef, J.; Hyde, K.D.; Lumyong, S. Pulveroboletus fragrans, a new Boletaceae species from Northern Thailand, with a remarkable aromatic odor. Mycol. Prog. 2016, 15, 1–8. [Google Scholar] [CrossRef]
- Vadthanarat, S.; Raspé, O.; Lumyong, S. Rubinosporus auriporus gen. et sp. nov. (Boletaceae: Xerocomoideae) from tropical forests of Thailand, producing unusual dark ruby spore deposits. J. Fungi 2022, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Wu, G.; Yang, Z.L. A new genus, Rubroboletus, to accommodate Boletus sinicus and its allies. Phytotaxa 2014, 188, 61–77. [Google Scholar] [CrossRef]
- Liang, Z.Q.; An, D.Y.; Jiang, S.; Su, M.S.; Zeng, N.K. Butyriboletus hainanensis (Boletaceae, Boletales), a new species from tropical China. Phytotaxa 2016, 267, 256–262. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Binder, M.; Roekring, S.; Flegel, T. Spongiforma, a new genus of gasteroid boletes from Thailand. Fungal Divers. 2009, 37, 1–8. [Google Scholar]
- Orihara, T.; Lebel, T.; Ge, Z.W.; Smith, M.; Maekawa, N. Evolutionary history of the sequestrate genus Rossbeevera (Boletaceae) reveals a new genus Turmalinea and highlights the utility of ITS minisatellite–like insertions for molecular identification. Persoonia 2016, 37, 173–198. [Google Scholar] [CrossRef]
- Li, Y.C.; Ortiz-Santana, B.; Zeng, N.K.; Feng, B.; Yang, Z.L. Molecular phylogeny and taxonomy of the genus Veloporphyrellus. Mycol. 2014, 106, 291–306. [Google Scholar] [CrossRef]
- Li, Y.C.; Feng, B.; Yang, Z.L. Zangia, a new genus of Boletaceae supported by molecular and morphological evidence. Fungal Divers. 2011, 49, 125–143. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. Bayesian Analysis of Molecular Evolution Using MrBayes; Springer: New York, NY, USA, 2005; pp. 183–226. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest 2.3. Program Distributed by the Author; Evolutionary Biology Center, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Singer, R. Das System der Agaricales. II. Ann. Mycol. 1942, 40, 1–132. [Google Scholar]
- Watling, R. A Manual and Source Book of the Boletes and Their Allies; Fungiflora: Oslo, Norway, 2008. [Google Scholar]
- Bandala, V.M.; Montoya, L.; Jarvio, D. Two interesting records of boletes found in coffee plantations in eastern Mexico. Persoonia-Mol. Phylogeny Evol. Fungi 2004, 18, 365–380. [Google Scholar]
- Šutara, J. Xerocomus s. l. in the light of the present state of knowledge. Czech Mycol. 2008, 60, 29–62. [Google Scholar] [CrossRef]
- Li, M.X.; Wu, G.; Yang, Z.L. Four new species of Hemileccinum (Xerocomoideae, Boletaceae) from southwestern China. J. Fungi 2021, 7, 823. [Google Scholar] [CrossRef]
- Bresinsky, A.; Besl, H. Schlüssel zur Gattungsbestimmung der Blätter-, Leisten-und Röhrenpilze mit Literaturhinweisen zur Artbestimmung. Regensbg. Mykol. Schriften 2003, 11, 5–236. [Google Scholar]
- Xue, R.; Wu, L.L.; Jiang, S.; Hao, Y.J.; Chai, H.; Liang, Z.Q.; Zeng, N.K.; Su, M.S. Two new species of the genus Leccinellum (Boletaceae, Boletales) from the south of China. Phytotaxa 2019, 175, 133–140. [Google Scholar] [CrossRef]
- Binder, M.; Besl, H. 28S rDNA sequence data and chemotaxonomical analyses on the generic concept of Leccinum (Boletales). In Micologia; Associazone Micologica Bresadola: Trento, Italy, 2000; pp. 71–82. [Google Scholar]
- Bakker, H.C.; Noordeloos, M.E. A revision of European species of Leccinum Gray and notes on extralimital species. Persoonia 2005, 18, 511–574. [Google Scholar]
- Halling, R.E.; Mueller, G.M. Leccinum (Boletaceae) in Costa Rica. Mycologia 2003, 95, 488–499. [Google Scholar] [CrossRef]
- Meng, X.; Wang, G.S.; Wu, G.; Wang, P.M.; Yang, Z.L.; Li, Y.C. The genus Leccinum (Boletaceae, Boletales) from China based on morphological and molecular data. J. Fungi 2021, 7, 732. [Google Scholar] [CrossRef]
- Halling, R.E.; Nuhn, M.; Fechner, N.A.; Osmundson, T.W.; Soytong, K.; Arora, D.; Hibbett, D.S.; Binder, M. Sutorius: A new genus for Boletus eximius. Mycologia 2012, 104, 951–961. [Google Scholar] [CrossRef]
- Vadthanarat, S.; Halling, R.E.; Amalfi, M.; Lumyong, S.; Raspé, O. An unexpectedly high number of new Sutorius (Boletaceae) species from northern and northeastern Thailand. Front. Microbiol. 2021, 12, 643505. [Google Scholar] [CrossRef] [PubMed]
- Su, L.H.; Geng, C.A.; Li, T.Z.; Huang, X.Y.; Ma, Y.B.; Zhang, X.M.; Wu, G.; Yang, Z.L.; Chen, J.J. Spiroseoflosterol, a Rearranged Ergostane-Steroid from the Fruiting Bodies of Butyriboletus roseoflavus. J. Nat. Prod. 2020, 83, 1706–1710. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Di, Y.T.; Zhang, Y.; Hu, X.J. Four new compounds from Neoboletus magnificus. Nat. Prod. Res. 2020, 34, 1152–1157. [Google Scholar] [CrossRef]
- Bahram, M.; Netherway, T. Fungi as mediators linking organisms and ecosystems. FEMS Microbiol. Rev. 2022, 46, fuab058. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Miyauchi, S.; Morin, E.; Kuo, A.; Drula, E.; Varga, T.; Kohler, A.; Feng, B.; Cao, Y.; Lipzen, A.; et al. Evolutionary innovations through gain and loss of genes in the ectomycorrhizal Boletales. New Phytol. 2022, 233, 1383–1400. [Google Scholar] [CrossRef]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher | Locality | GenBank Accession Nos. | Reference | ||
|---|---|---|---|---|---|---|
| 28S | TEF1 | RPB2 | ||||
| Abtylopilus scabrosus | HKAS50211 | Yunnan, SW China | KT990552 | KT990752 | KT990389 | [39] |
| Acyanoboletus controversus | HKAS126560 | Yunnan, SW China | OQ888714 | OQ873451 | OQ873490 | [3] |
| Acyanoboletus dissimilis | ZT14030 | Malaysia | OQ888716 | OQ873453 | OQ873492 | [3] |
| Afrocastellanoa ivoryana | Arora 126 | Zimbabwe | KX685721 | KX685715 | — | [40] |
| Alessioporus ichnusanus | KJ729504 | Italy | KJ729504 | KJ729513 | — | [41] |
| Amoenoboletus miraculosus | ZT14046 | Malaysia | MW520188 | MW566745 | — | [18] |
| Amoenoboletusgranulopunctatus | HKAS56280 | Yunnan, SW China | KF112418 | KF112265 | KF112708 | [15] |
| Amylotrama clelandii | MEL2432546 | Australia | MT459235 | MN413630 | — | [42] |
| Anthracoporus holophaeus | HKAS59407 | China | KT990708 | KT990888 | KT990506 | [39] |
| Aureoboletus catenarius | HKAS54467 | Yunnan, SW China | KT990510 | KT990711 | KT990349 | [39] |
| Aureoboletus duplicatoporus | HKAS83115 | Yunnan, SW China | KT990512 | KT990713 | KT990351 | [39] |
| Austroboletus aff. fusisporus | HKAS52683 | Fujian, SE China | KF112484 | KF112213 | KF112766 | [15] |
| Baorangia rufomaculata | 4414 | USA | KF030248 | KF030406 | — | [43] |
| Baorangia pseudocalopus | HKAS75739 | Yunnan, SW China | KJ184558 | KJ184570 | KM605179 | [44] |
| Boletellus aff. emodensis | HKAS 52678 | Fujian, SE China | KF112426 | KF112305 | KF112757 | [15] |
| Boletellus indistinctus | HKAS77623 | Guangdong, southern China | KT990531 | KT990733 | KT990371 | [39] |
| Boletinellus merulioides | AFTOL-ID 575 | USA | AY684153 | DQ056287 | — | [45] |
| Boletus bainiugan | HKAS52235 | Yunnan, SW China | KF112457 | KF112203 | KF112705 | [15] |
| Boletus morrisii | 8206 | USA | KF030326 | KF030433 | — | [43] |
| Borofutus dhakanus | HKAS73789 | Bangladesh | JQ928616 | JQ928576 | JQ928597 | [46] |
| Bothia castanella | MB03_053 | USA | DQ867117 | KF030421 | — | [43,47] |
| Bothia fujianensis | HKAS82694 | Fujian, SE China | KM269193 | KM272860 | — | [48] |
| Buchwaldoboletus lignicola | HKAS76674 | Heilongjiang, NE China | KF112350 | KF112277 | KF112819 | [15] |
| Buchwaldoboletus lignicola | HKAS84904 | Germany | KT990538 | KT990740 | KT990377 | [39] |
| Butyriboletus pseudospeciosus | HKAS63513 | Yunnan, SW China | KT990541 | KT990743 | KT990380 | [39] |
| Butyriboletus regius | HKAS84878 | Germany | MT264910 | MT269659 | MT269661 | Unpublished |
| Butyriboletus subsplendidus | HKAS50444 | Yunnan, SW China | KT990540 | KT990742 | KT990379 | [39] |
| Cacaoporus pallidicarneus | OR1306 | Thailand | — | MK372272 | MK372285 | [10] |
| Caloboletus calopus | HKAS74739 | Yunnan, SW China | KF112335 | KF112166 | KF112667 | [15] |
| Caloboletus yunnanensis | HKAS63040 | Yunnan, SW China | KJ605676 | KJ619471 | KT990395 | [39,49] |
| Chalciporus piperatus | HKAS84882 | Germany | KT990562 | KT990758 | KT990397 | [39] |
| Chalciporus rubinelloides | HKAS74952 | Yunnan, SW China | KT990565 | KT990761 | KT990400 | [39] |
| Chamonixia caespitosa | OSC-117571 | USA | MK601731 | MK721085 | MK766293 | [50] |
| Chiua olivaceoreticulata | HKAS59706 | Yunnan, SW China | KT990593 | KT990787 | KT990428 | [39] |
| Chiua virens | HKAS76678 | Sichuan, SW China | KF112438 | KF112272 | KF112793 | [15] |
| Crocinoboletus laetissimus | HKAS50232 | China | KT990567 | KT990762 | — | [39] |
| Crocinoboletus rufoaureus | HKAS59820 | Hainan, southern China | KF112434 | — | KF112709 | [15] |
| Cyanoboletus brunneoruber | HKAS80579-1 | Yunnan, SW China | KT990568 | KT990763 | KT990401 | [39] |
| Cyanoboletus sinopulverulentus | HKAS59609 | Yunnan, SW China | KF112366 | KF112193 | KF112700 | [15] |
| Erythrophylloporus cinnabarinus | GDGM70536 | Hainan, southern China | MH374045 | MH378802 | MH374035 | [51] |
| Exsudoporus floridanus | FLAS-F-59069 | USA | OL960488 | OL960496 | OL960503 | [52] |
| Fistulinella prunicolor | REH9502 | Australia | JX889648 | JX889690 | MG212630 | [53,54] |
| Guyanaporus albipodus | TH8848 | Guyana | HQ161868 | — | LC043083 | [7,55] |
| Gyrodon lividus | REG Gl1 | Germany | — | GU187701 | GU187786 | [56] |
| Gyrodon sp. | HKAS59448 | Yunnan, SW China | KF112349 | KF112276 | KF112818 | [15] |
| Harrya atrogrisea | HKAS50542 | Yunnan, SW China | KT990694 | KT990880 | KT990499 | [39] |
| Heimioporus japonicus | HKAS52237 | Yunnan, SW China | KF112347 | KF112228 | KF112806 | [15] |
| Heimioporus subretisporus | HKAS80582 | Yunnan, SW China | KT990574 | KT990770 | KT990409 | [39] |
| Heliogaster columellifer | KPM-NC 23012 | Japan | KX685724 | KX685718 | — | [40] |
| Hemilanmaoa retistipitatus | HMJAU 60052 (H3624) | Guizhou, SW China | OP380695 | OP495816 | OP495814 | [1] |
| Hemilanmaoa retistipitatus | HMJAU 60053 (H3633) | Guizhou, SW China | OP380696 | OP495817 | OP495815 | [1] |
| Hemileccinum impolitum | HKAS84869 | Germany | KT990575 | KT990771 | KT990410 | [39] |
| Hemileccinum rugosum | HKAS84355 | Yunnan, SW China | KT990578 | KT990774 | KT990413 | [39] |
| Hongoboletus ventricosus | HKAS59660 | Yunnan, SW China | KF112358 | KF112153 | KF112664 | [15] |
| Hongoboletus ventricosus | HKAS63598 | Yunnan, SW China | OQ888735 | KF112152 | KF112663 | [3,15] |
| Hourangia cheoi | HKAS52269 | Yunnan, SW China | KF112385 | KF112286 | KF112773 | [15] |
| Hourangia cheoi | HKAS74774 | Yunnan, SW China | KF112384 | KF112285 | KF112772 | [15] |
| Hymenoboletus luteopurpureus | HKAS46334 | Yunnan, SW China | KF112471 | KF112271 | KF112795 | [15] |
| Imleria badia | HKAS74714 | Germany | KC215212 | KC215242 | — | [57] |
| Imleria obscurebrunnea | HKAS52557 | Yunnan, SW China | KC215220 | KC215243 | KC215234 | [57] |
| Imperator torosus | MB000258 | Germany | — | MW566748 | MW560082 | [18] |
| Ionosporus longipes | Lee1180 | Singapore | — | MT085471 | MH712031 | [58,59] |
| Kgaria cyanogranulifer | REH9508 | Australia | JX889646 | JX889688 | OR263680 | [53,60] |
| Lanmaoa angustispora | HKAS74752 | Yunnan, SW China | KM605139 | KM605154 | KM605177 | [44] |
| Lanmaoa asiatica | HKAS54094 | Yunnan, SW China | KF112353 | KF112161 | KF112682 | [15] |
| Leccinellum corsicum | Buf4507 | USA | KF030347 | KF030435 | — | [43] |
| Leccinellum sp. | HKAS53427 | Hunan, central China | KF112488 | KF112253 | — | [15] |
| Leccinum quercinum | HKAS63502 | Yunnan, SW China | KF112444 | KF112250 | KF112724 | [15] |
| Leccinum scabrum | HKAS56371 | Yunnan, SW China | KT990587 | KT990782 | KT990423 | [39] |
| Leccinum variicolor | HKAS57758 | Yunnan, SW China | KF112445 | KF112251 | KF112725 | [15] |
| Mucilopilus castaneiceps | HKAS71039 | Japan | KT990547 | KT990748 | KT990385 | [39] |
| Mucilopilus castaneiceps | HKAS50338 | Yunnan, SW China | KT990555 | KT990755 | KT990391 | [39] |
| Neoboletus thibetanus | HKAS57093 | Tibet, SW China | KF112326 | — | KF112655 | [15] |
| Nevesoporus nigrostipitatus | VIES 9901383 | Brazil | OM068918 | OM160562 | — | [11] |
| Nevesoporus nigrostipitatus | VIES 9901384 | Brazil | OM068919 | — | — | [11] |
| Octaviania japonimontana | KPM-NC-0017812 | Japan | JN378486 | JN378428 | – | [61] |
| Octaviania tasmanica | OSC132097 | Australia | JN378494 | JN378435 | – | [61] |
| Paxilloboletus africanus | SAB0716 | Guinea | MZ702479 | MZ707865 | MZ707869 | [13] |
| Paxillus rubicundulus | Ve08.2h10 | France | — | KF261553 | — | [62] |
| Paxillus ammoniavirescens | Pou09.1 | France | — | KF261533 | — | [62] |
| Paxillus involutus | SCV11.1 | France | — | KF261544 | — | [62] |
| Phlebopus portentosus | Php1 | Africa | AF336260 | GU187735 | GU187801 | [56,63] |
| Phlebopus portentosus | FHMU5935 | Yunnan, SW China | MW783432 | MW897345 | — | [64] |
| Phylloboletellus chloephorus | 3388 | Mexico | DQ534658 | — | — | [65] |
| Phylloporopsis boletinoides | JBSD127411 | Dominican Republic | MH571711 | MH588312 | — | [12] |
| Phylloporus maculatus | HKAS56683 | Yunnan, SW China | JQ967210 | JQ967167 | — | [20] |
| Phylloporus rubrosquamosus | HKAS 54559 | Yunnan, SW China | JQ967219 | JQ967175 | — | [20] |
| Porphyrellus castaneus | HKAS52554 | Yunnan, SW China | KT990697 | KT990883 | KT990502 | [39] |
| Porphyrellus porphyrosporus | HKAS76671 | Jilin, NE China | KF112482 | KF112243 | KF112718 | [15] |
| Pseudoaustroboletus valens | HKAS52603 | Yunnan, SW China | KM274869 | KM274877 | — | [66] |
| Pseudoaustroboletus valens | HKAS82644 | China | — | MT110359 | MT110431 | [67] |
| Pseudophylloporus baishanzuensis | N.K. Zeng7702 (FHMU7694) | Zhejiang, eastern China | PQ330210 | PQ330110 | PQ330114 | This study |
| Pseudophylloporus baishanzuensis | N.K. Zeng7703 (FHMU7695) | Zhejiang, eastern China | PQ330211 | PQ330111 | PQ330115 | This study |
| Pseudophylloporus baishanzuensis | N.K. Zeng7705 (FHMU7696) | Zhejiang, eastern China | PQ330212 | PQ330112 | PQ330116 | This study |
| Pseudophylloporus baishanzuensis | N.K. Zeng7746 (FHMU7697) | Zhejiang, eastern China | PQ330213 | PQ330113 | PQ330117 | This study |
| Pulchroboletus roseoalbidus | AMB 12757 | Italy | KJ729499 | KJ729512 | — | [41] |
| Pulveroboletus brunneopunctatus | HKAS55369 | Yunnan, SW China | KT990620 | KT990814 | KT990455 | [39] |
| Pulveroboletus mirus | HKAS57628 | Yunnan, SW China | KT990618 | KT990812 | KT990453 | [39] |
| Retiboletus brunneolus | HKAS52680 | Fujian, SE China | KF112424 | KF112179 | KF112690 | [15] |
| Retiboletus fuscus | HKAS63590 | Yunnan, SW China | KF112417 | KF112178 | KF112691 | [15] |
| Rhodactina rostratispora | SV208 | Thailand | — | MG212606 | MG212646 | [54] |
| Rossbeevera vittatispora | OSC61484 | Australia | JN378506 | JN378446 | — | [61] |
| Rostrupomyces sisongkhramensis | SV0155 | Thailand | — | OP358324 | OP358316 | [68] |
| Rostrupomyces sisongkhramensis | SV0219 | Thailand | — | OP358325 | OP358317 | [68] |
| Royoungia boletoides | Trappe 27456 | Australia | JX889655 | JX889696 | — | [53] |
| Royoungia palumanus | REH9421 | Australia | JX889675 | JX889685 | — | [53] |
| Rubinoboletus rubinus | AF2835 | Belgium | — | KT824028 | KT823995 | [69] |
| Rubinosporus auriporus | SV0101 | Thailand | — | MZ355902 | MZ355904 | [70] |
| Rubroboletus latisporus | HKAS80358 | Chongqing, SW China | KP055023 | KP055020 | KP055029 | [71] |
| Rubroboletus sinicus | HKAS68620 | Yunnan, SW China | KF112319 | KF112146 | KF112661 | [15] |
| Rubroleccinum latisporus | N.K. Zeng7988 (FHMU7698) | Fujian, SE China | PQ325253 | PQ330106 | PQ330108 | This study |
| Rubroleccinum latisporus | N.K. Zeng8006 (FHMU7699) | Fujian, SE China | PQ325254 | PQ330107 | PQ330109 | This study |
| Rufoboletus hainanensis | KUN-HKAS 59814 | Hainan, southern China | KF112336 | KF112199 | KF112699 | [15] |
| Rufoboletus hainanensis | N.K. Zeng2418 (FHMU2437) | Hainan, southern China | KU961652 | KU961656 | KX453856 | [72] |
| Rugiboletus brunneiporus | HKAS83209 | Tibet, SW China | KM605134 | KM605144 | KM605168 | [44] |
| Rugiboletus extremiorientalis | HKAS74754 | China | KT990639 | KT990832 | KT990469 | [39] |
| Singerocomus rubriflavus | Henkel 9585 | Guyana | LC043093 | MH645597 | — | [7,10] |
| Spongiforma thailandica | DED7873 | Thailand | EU685108 | KF030436 | — | [43,73] |
| Strobilomyces atrosquamosus | HKAS55368 | Yunnan, SW China | KT990648 | KT990839 | KT990476 | [39] |
| Strobilomyces seminudus | HKAS59461 | Yunnan, SW China | KF112479 | KF112260 | KF112815 | [15] |
| Suillellus amygdalinus | NY00035656 | USA | KT990650 | KT990840 | KT990477 | [39] |
| Suillellus subamygdalinus | HKAS57953 | Tibet, SW China | KT990652 | KT990842 | — | [39] |
| Sutorius brunneissimus | HKAS57451 | Yunnan, SW China | KM605137 | KM605149 | KM605172 | [44] |
| Sutorius brunneissimus | HKAS50538 | Yunnan, SW China | KM605138 | KM605150 | KM605173 | [44] |
| Sutorius hainanensis | HKAS59469 | Yunnan, SW China | KF112359 | KF112175 | KF112669 | [15] |
| Tengioboletus glutinosus | HKAS53425 | Hunan, central China | KF112341 | KF112204 | KF112800 | [15] |
| Tengioboletus reticulatus | HKAS53426 | Hunan, central China | KF112491 | KF112313 | KF112828 | [15] |
| Turmalinea yuwanensis | KPM-NC-0018011 | Japan | KC552046 | KC552089 | — | [74] |
| Tylocinum griseolum | HKAS52612 | Yunnan, SW China | KT990631 | KT990825 | — | [39] |
| Tylocinum griseolum | HKAS50281 | Yunnan, SW China | KF112451 | KF112284 | KF112730 | [15] |
| Tylopilus aff. chromapes | 01-513 | Zambia | JX889672 | JX889682 | — | [53] |
| Tylopilus aff. virens | 01-541 | Zambia | JX889677 | JX889687 | — | [53] |
| Tylopilus otsuensis | HKAS50240 | Yunnan, SW China | KT990553 | KT990753 | MT110417 | [39,67] |
| Tylopilus sp. | HKAS50229 | Yunnan, SW China | KF112423 | KF112216 | KF112769 | [15] |
| Tylopilus violaceobrunneus | HKAS89443 | Shandong, eastern China | KT990702 | KT990886 | KT990504 | [39] |
| Veloboletus limbatus | REH9228 | Australia | — | MN413636 | MT747397 | Unpublished |
| Veloporphyrellus pseudovelatus | HKAS52258 | Yunnan, SW China | JX984540 | JX984551 | MT110439 | [67,75] |
| Veloporphyrellus velatus | HKAS63668 | Hainan, southern China | JX984546 | JX984554 | — | [75] |
| Xanthoconium affine | NY00815399 | USA | KT990661 | KT990850 | KT990486 | [39] |
| Xanthoconium porophyllum | HKAS90217 | Guangdong, southern China | KT990662 | KT990851 | KT990487 | [39] |
| Xerocomellus communis | HKAS68204 | Yunnan, SW China | KT990671 | KT990859 | KT990495 | [39] |
| Xerocomus subparvus | HKAS53387 | Fujian, SE China | KF112397 | KF112297 | KF112788 | [15] |
| Xerocomus yunnanensis | HKAS68420 | Yunnan, SW China | KT990690 | KT990877 | — | [39] |
| Zangia olivaceobrunnea | HKAS52275 | Yunnan, SW China | HQ326947 | HQ326875 | — | [76] |
| Zangia roseola | HKAS75046 | Yunnan, SW China | KF112414 | KF112269 | KF112791 | [15] |
| Genus | Pileal Surface | Context | Hymenophore | Stipe Surface | Basidiospores | Clamp Connections |
|---|---|---|---|---|---|---|
| Buchwaldoboletus | Tomentose or pulverulent, yellow to brownish. | Changing blue when injured. | Poroid, light yellow to ochraceous yellow, changing bluish to dark blue when injured. | Yellow to brown tones, without squamules. | Subfusiform, smooth. | Absence. |
| Chalciporus | Glabrous to obscurely, subtomentose, pinkish-red to reddish-brown. | Unchanging or turning bluish when injured. | Poroid, pinkish red to reddish brown, unchanging or staining bluish to dull blue slowly when injured. | Yellow to brown tones, without squamules. | Subfusiform, smooth. | Absence. |
| Erythrophylloporus | Pruinose or velutinous, subtomentose to faintly squamulose, orange, reddish-orange to yellowish-red. | Changing dark violet to blackish blue when injured. | Lamellate, red, usually not forked, changing grayish blue when injured. | Reddish-orange to yellowish-red, covered with reddish-orange to orange red pruinose scales. | Broadly ellipsoid, ellipsoid to nearly ovoid, smooth. | Absence. |
| Paxilloboletus | Tomentose, white, cream to yellowish. | Changing blue when injured. | Lamellate, yellow to yellowish-brown, regularly bifurcating and anastomosing, unchanging in color when injured. | White to yellowish-white tomentose, with or without ridges or reticulation on its uppermost part. | Ellipsoid to fusiform, smooth. | Absence. |
| Phylloboletellus | Glabrous, yellow-orange to orange-reddish or brown. | Changing blue when injured. | Lamellate, yellowish-green to olive-brown, usually forked, changing blue when injured. | Yellow to mustard yellow, covered with brownish clumps or fibrils. | Broadly ellipsoid, with longitudinal, continuous or bifurcate ribs. | Sometimes scarce. |
| Phyllobolites | Glabrous to tomentose, olivaceous brown or reddish-brown. | Unchanging or changing blue when injured. | Lamellate, sometimes forked, yellowish-cream, yellow to luteous, turning sienna or rust-color to chestnut when injured. | Yellow tone, pulverulent to tomentose. | Fusiform, verrucose. | Absence. |
| Phylloporopsis | Velvety-tomentose to fibrillose, carmin red, dull red to reddish-brown. | Changing blue when injured. | Lamellate, sometimes sub-boletinoid, usually not forked, beige to olive cream, changing blue when injured. | Yellowish-brown to reddish-brown, pruinose to longitudinally fibrillose. | Ellipsoid to fusiform, smooth. | Absence. |
| Phylloporus | Tomentose, yellowish-brown to reddish-brown. | Unchanging in color when injured. | Lamellate, usually not forked, yellow to golden yellow, unchanging or changing blue to greenish-blue when injured. | Yellowish-brown to reddish-brown, tomentose. | Fusoid to fusiform, with bacillate ornamentation. | Usually absent. |
| Pseudophylloporus | Nearly smooth, yellowish-brown to earthy yellow. | Turning blue, then changing red, and finally black when injured. | Lamellate, yellow to yellowish-brown, usually forked, turning blue, then changing red, and finally black when injured. | Tawny to pale brown, densely covered with pale brown scales. | Fusoid to elongate, smooth. | Present. |
| Genus | Pileal Surface | Context | Hymenophore | Stipe Surface | Basidiospores | Pileipellis |
|---|---|---|---|---|---|---|
| Hemileccinum | Smooth to subtomentose, or rugose, yellowish-brown to reddish-brown. | Unchanging in color when injured. | Poroid, light yellow to olive yellow, unchanging in color when injured. | Whitish, to cream, pale yellow-brown to dark brown, ornamented with white to yellow, or brown scales. | Subfusiform, irregularly warty. | Hyphoepithelioid type. |
| Leccinellum | Glabrous to subtomentose, rugulose or pitted brown, reddish-brown, brown to dark brown. | Changing red when injured. | Poroid, whitish or yellow, unchanging or staining brownish to ferruginous, or at first reddish then blackish when injured. | White, grayish-brown to yellowish-brown, covered with scabrous, brown to blackish squamules. | Subfusiform to ellipsoid, smooth. | Epithelioid type. |
| Leccinum | Glabrous to subtomentose, grayish-white, yellowish-brown to dark brown. | Unchanging or staining blue or red when injured. | Poroid, whitish or yellow, unchanging or staining blue or red when injured. | White, grayish to blackish-brown covered with scabrous to dotted and brown to blackish squamules. | Subfusiform, smooth. | Trichodermal type. |
| Rubroleccinum | Nearly smooth, orange to reddish-orange when young, then grayish-yellow to reddish-brown. | Changing blue, then turning red when injured. | Poroid, brilliant yellow to yellow, changing blue, then turning red when injured. | Yellow to brown, punctuated with red to reddish-brown scabers. | Cylindrical to fusoid, smooth. | Trichodermal type. |
| Singerocomus | Tomentose, pinkish red to red. | Unchanging in color when injured. | Poroid, yellow, unchanging in color when injured. | Yellow to brown, glabrous or with dull yellow squamules. | Ellipsoidal, smooth. | Trichodermal type. |
| Sutorius | Glabrous to subtomentose, chocolate brown to reddish brown or purplish brown. | Unchanging or staining blue to dark blue when injured. | Poroid, dark purple, purplish red or purplish brown, unchanging or turning red when injured. | Pinkish, yellow to reddish-brown, covered with pale brown to brown, or pale light purple squamules. | Subcylindrical to subfusiform, smooth. | Trichodermal type. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, H.-Z.; Wang, Y.; Lin, W.-F.; Zeng, H.; Hu, L.-G.; Ke, B.-R.; Zeng, Z.-H.; Liang, Z.-Q.; Zeng, N.-K. Pseudophylloporus Gen. nov. and Rubroleccinum Gen. nov., Two New Genera Revealed by Morphological and Phylogenetic Evidences in the Family Boletaceae from Subtropical China. J. Fungi 2024, 10, 817. https://doi.org/10.3390/jof10120817
Qin H-Z, Wang Y, Lin W-F, Zeng H, Hu L-G, Ke B-R, Zeng Z-H, Liang Z-Q, Zeng N-K. Pseudophylloporus Gen. nov. and Rubroleccinum Gen. nov., Two New Genera Revealed by Morphological and Phylogenetic Evidences in the Family Boletaceae from Subtropical China. Journal of Fungi. 2024; 10(12):817. https://doi.org/10.3390/jof10120817
Chicago/Turabian StyleQin, Hua-Zhi, Yi Wang, Wen-Fei Lin, Hui Zeng, Li-Gui Hu, Bin-Rong Ke, Zhi-Heng Zeng, Zhi-Qun Liang, and Nian-Kai Zeng. 2024. "Pseudophylloporus Gen. nov. and Rubroleccinum Gen. nov., Two New Genera Revealed by Morphological and Phylogenetic Evidences in the Family Boletaceae from Subtropical China" Journal of Fungi 10, no. 12: 817. https://doi.org/10.3390/jof10120817
APA StyleQin, H.-Z., Wang, Y., Lin, W.-F., Zeng, H., Hu, L.-G., Ke, B.-R., Zeng, Z.-H., Liang, Z.-Q., & Zeng, N.-K. (2024). Pseudophylloporus Gen. nov. and Rubroleccinum Gen. nov., Two New Genera Revealed by Morphological and Phylogenetic Evidences in the Family Boletaceae from Subtropical China. Journal of Fungi, 10(12), 817. https://doi.org/10.3390/jof10120817