
Fusion Expression and Immune Effect of PCV2 Cap Protein Tandem Multiantigen Epitopes with CD154/GM-CSF

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells Lines, Plasmids and Viruses
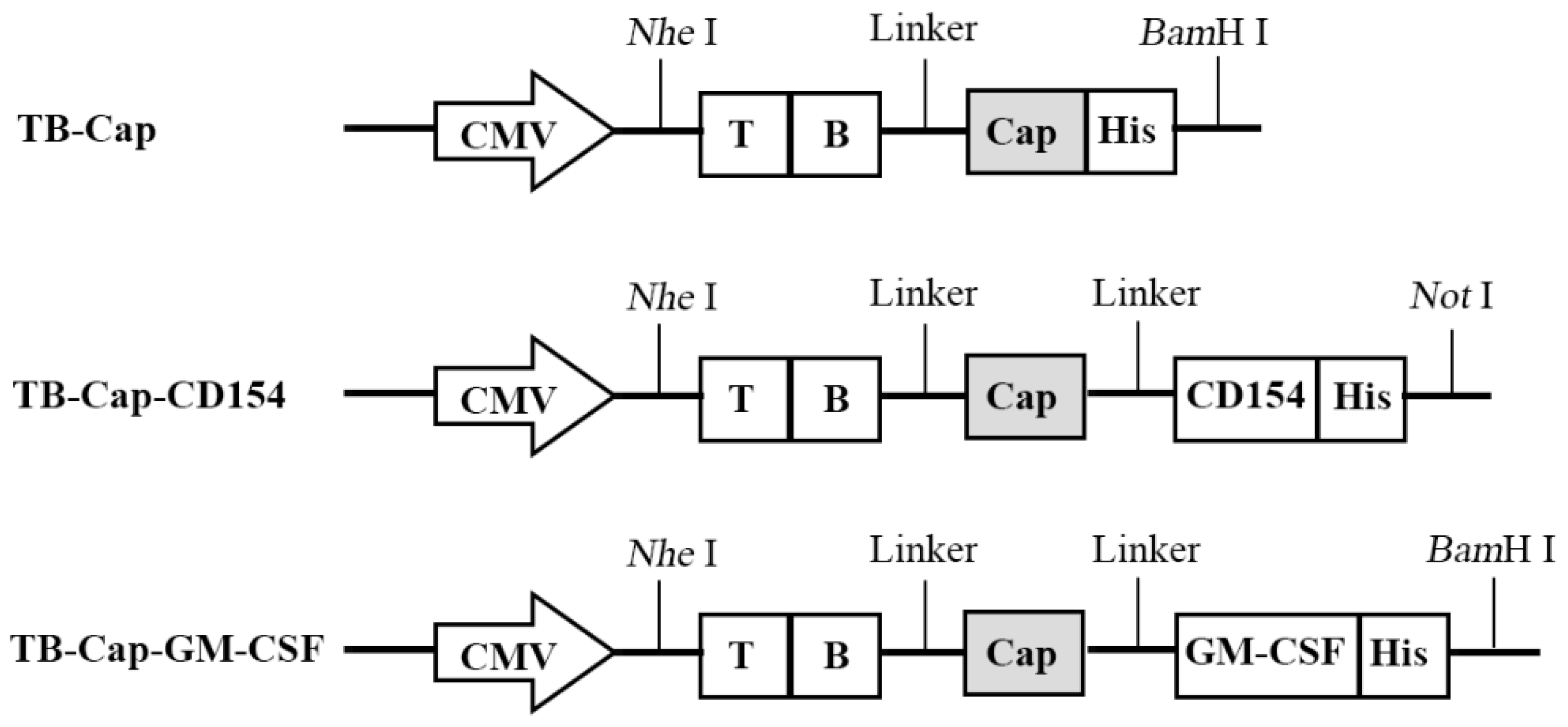
2.2. Construction of Recombinant Lentivirus Plasmids and Production of Lentivirus
2.3. Infection CHO-K1 of Recombinant Lentivirus
2.4. Detection of the Recombinant Proteins in CHO-K1 Cells
2.5. Animals, Vaccination, and Challenge Experiments
2.6. Serology
2.7. Lymphocyte Proliferation Assay
2.8. Detection of CD4+ and CD8+ by Flow Cytometry
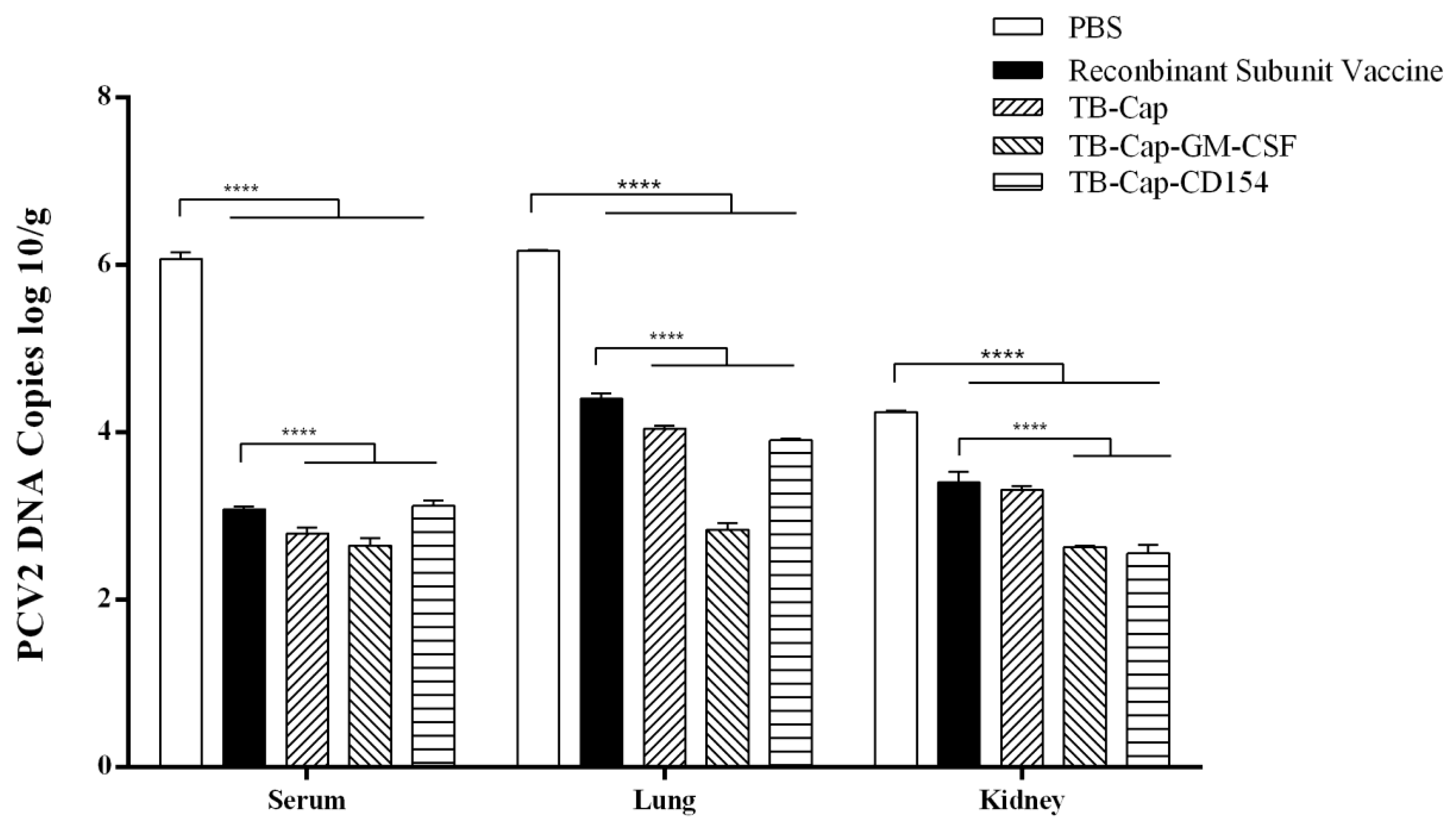
2.9. Detection of the Viremia by PCR
2.10. Real-Time PCR
2.11. IL-2, IFN-Gamma, IL-4 and IL-10 Cytokine Detection
2.12. Statistical Analysis
3. Results
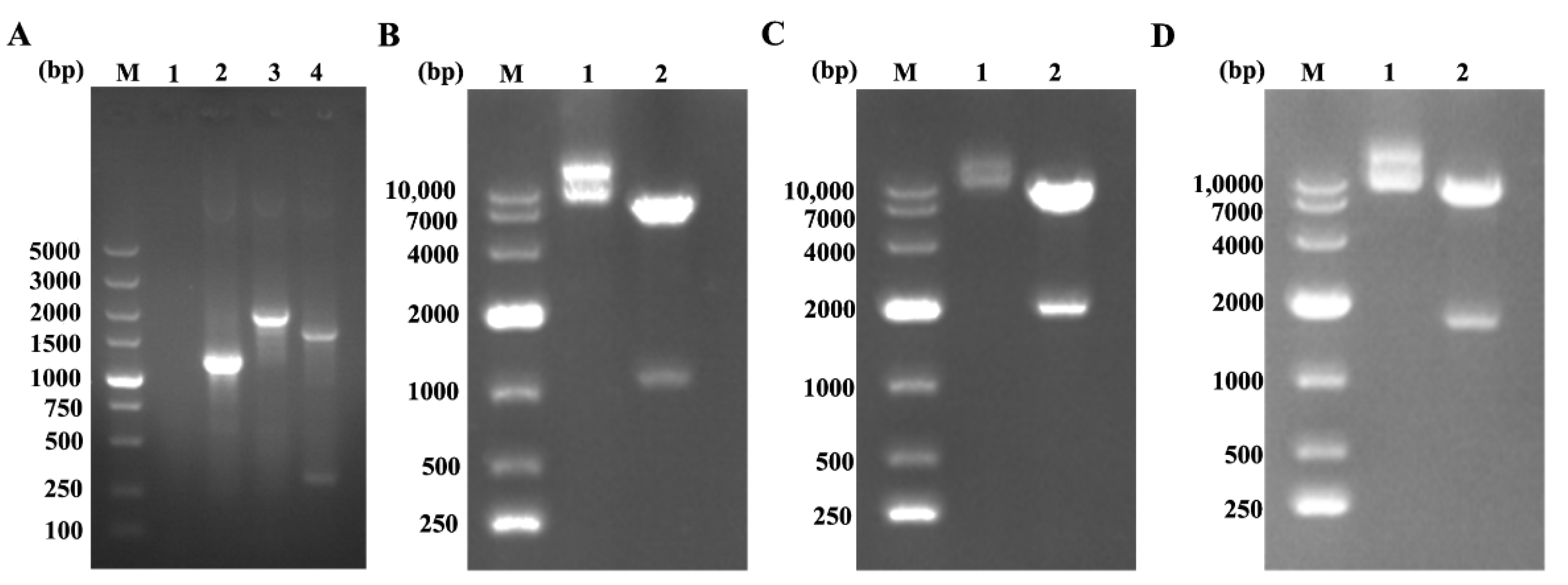
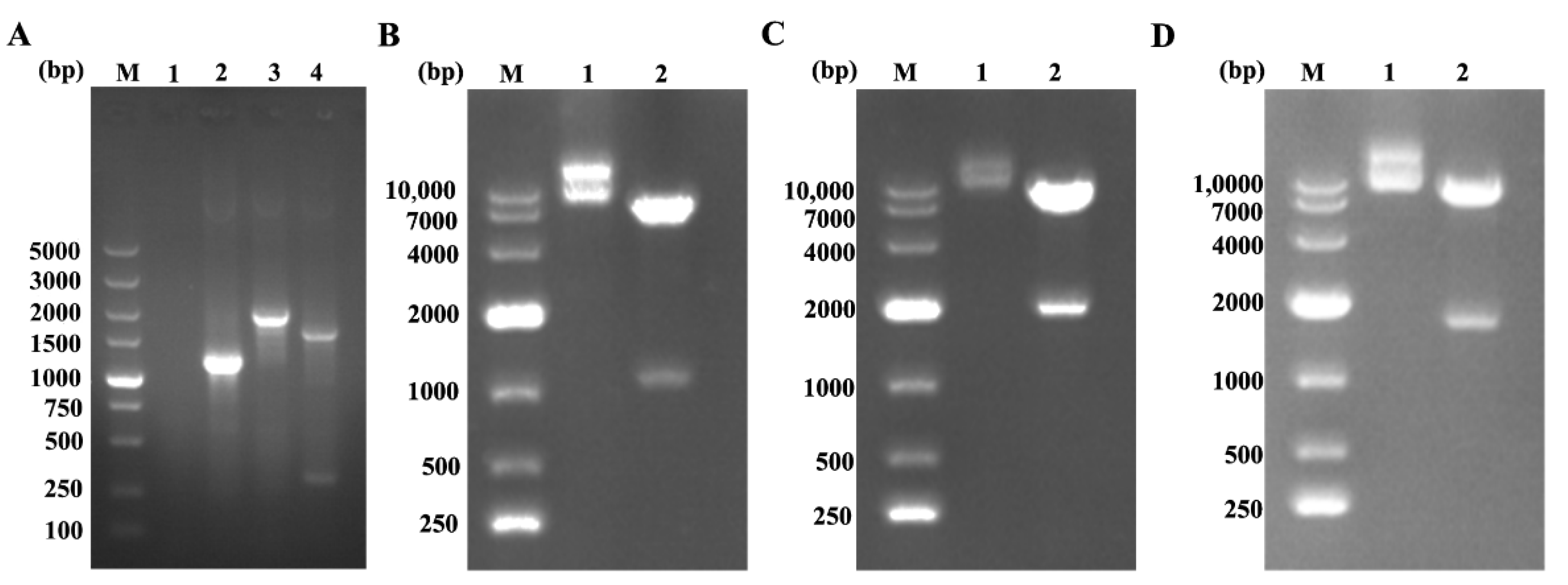
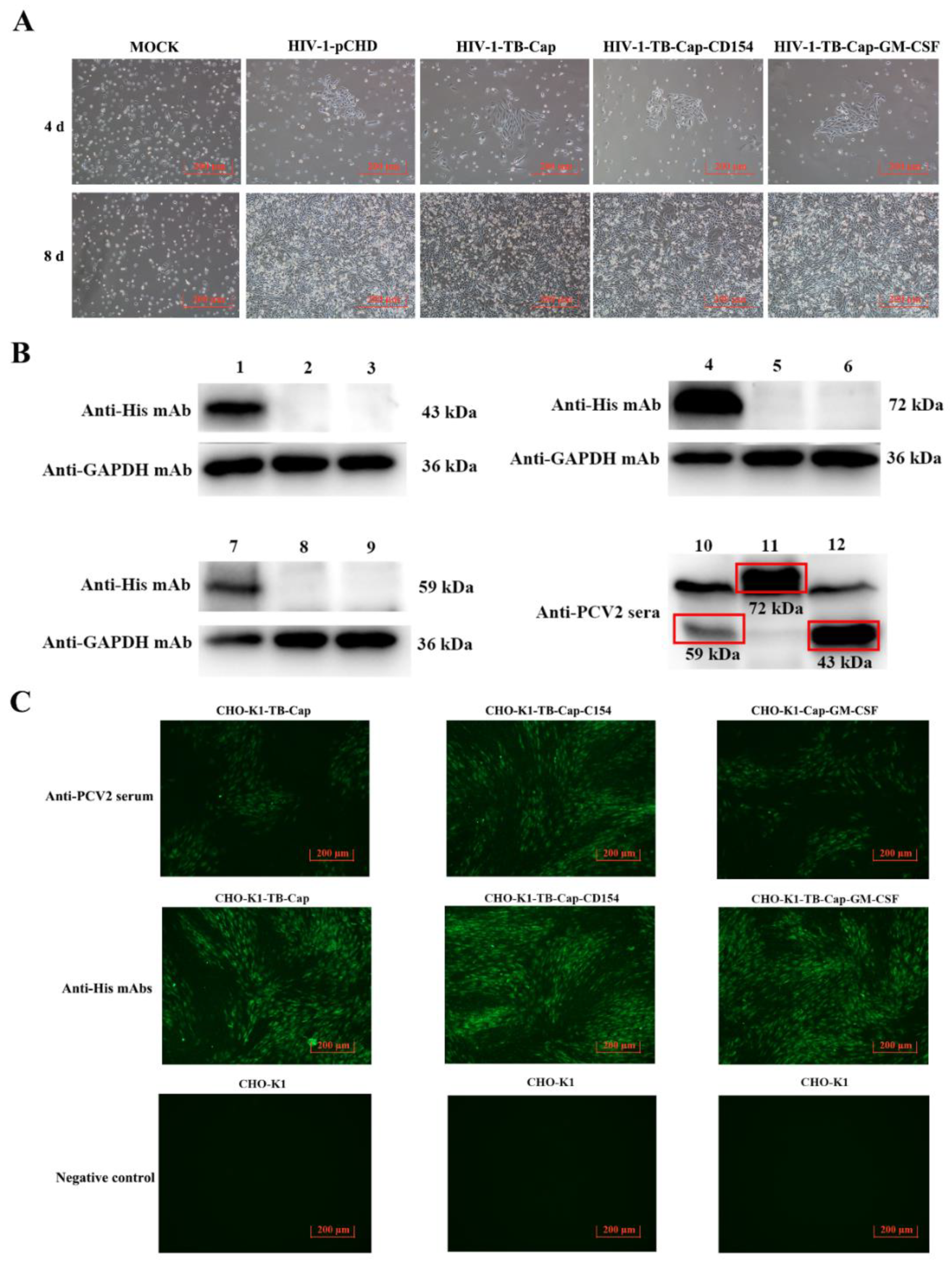
3.1. Dentification of Recombinant Lentivirus Plasmids
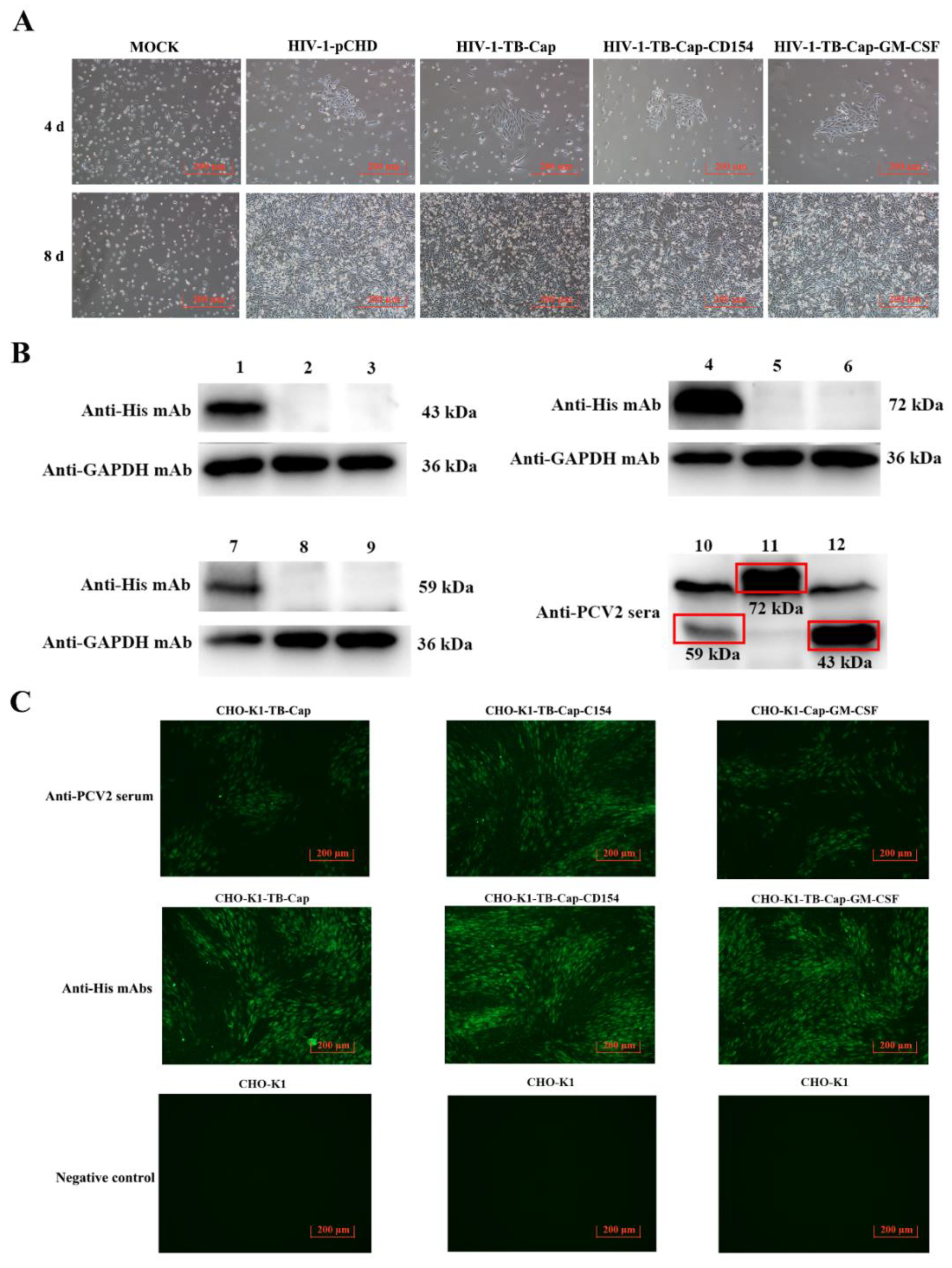
3.2. Expression of the Recombinant Proteins
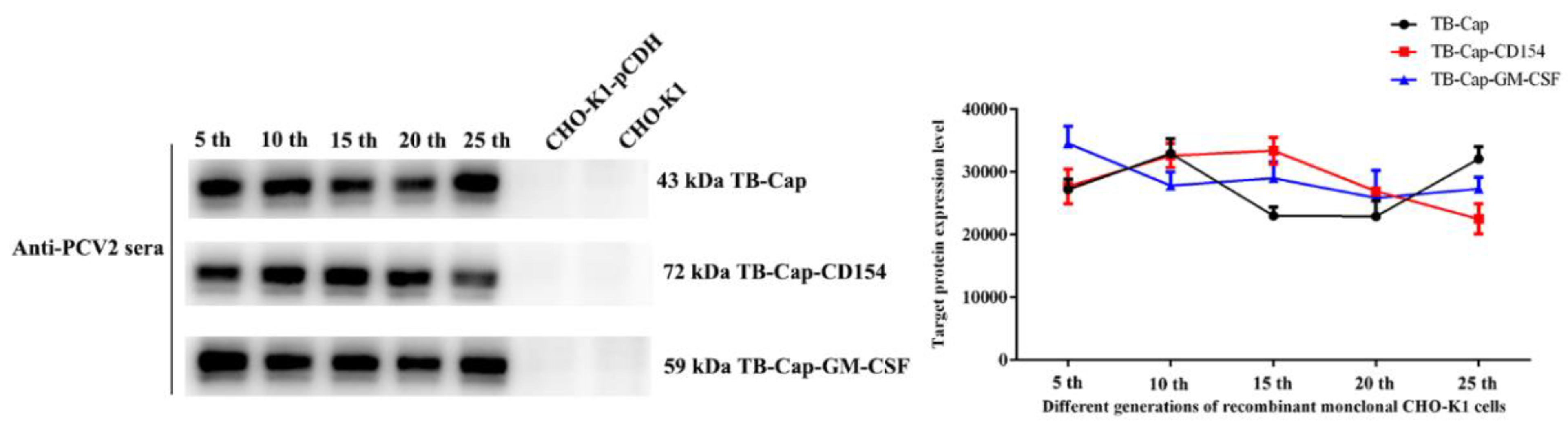
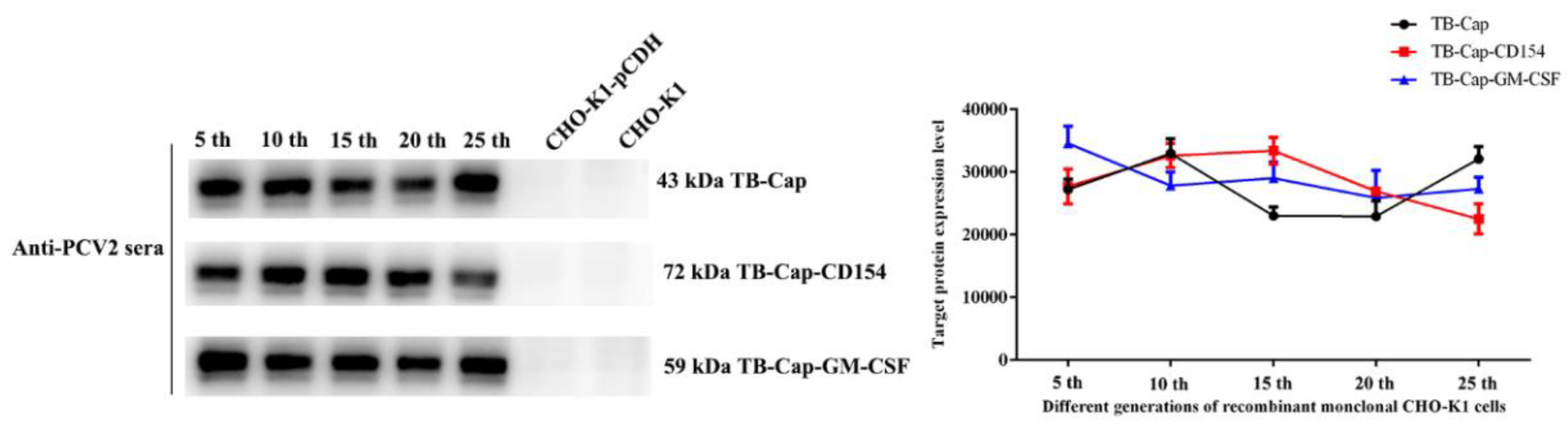
3.3. Screening and Stability Identification of Monoclonal Cells
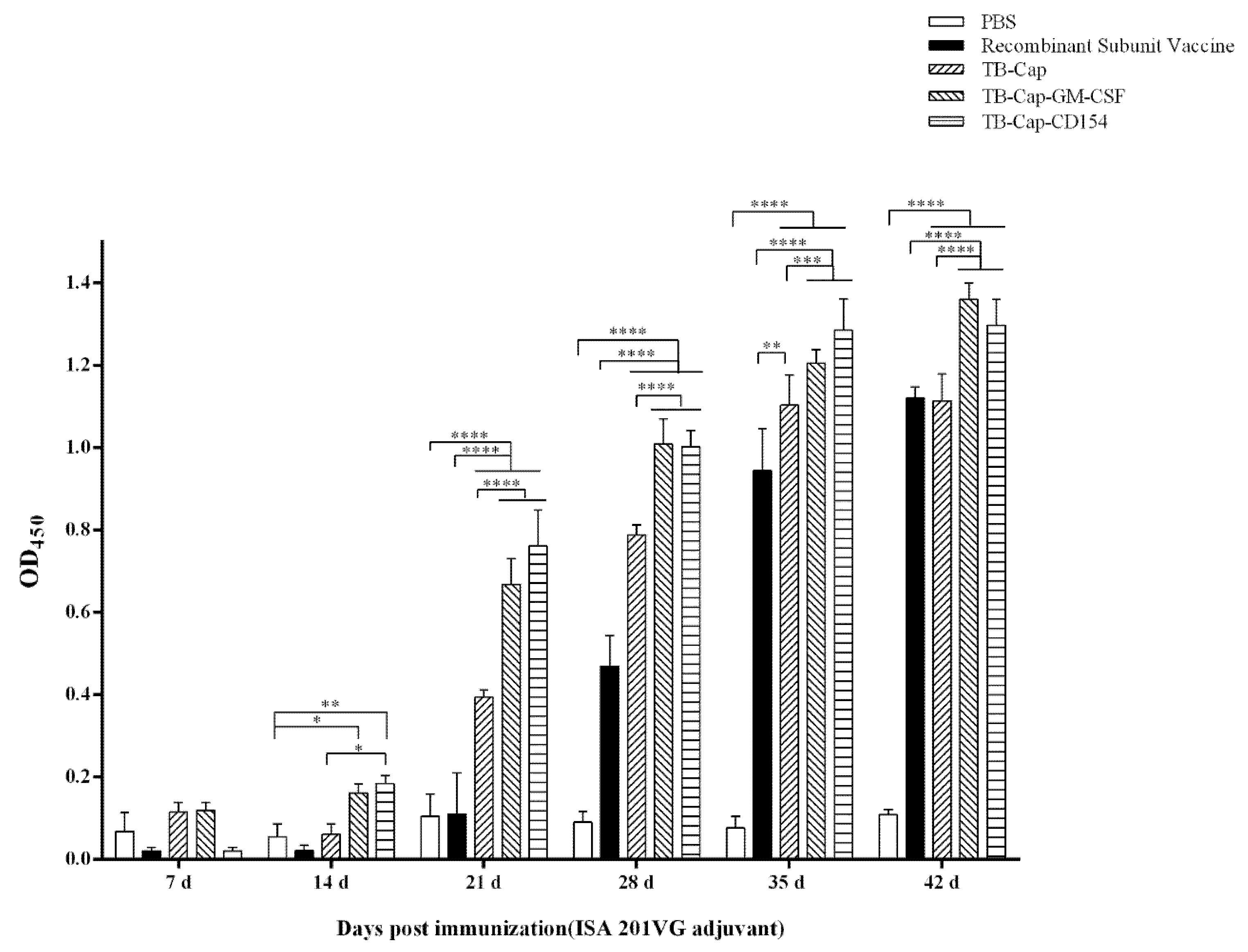
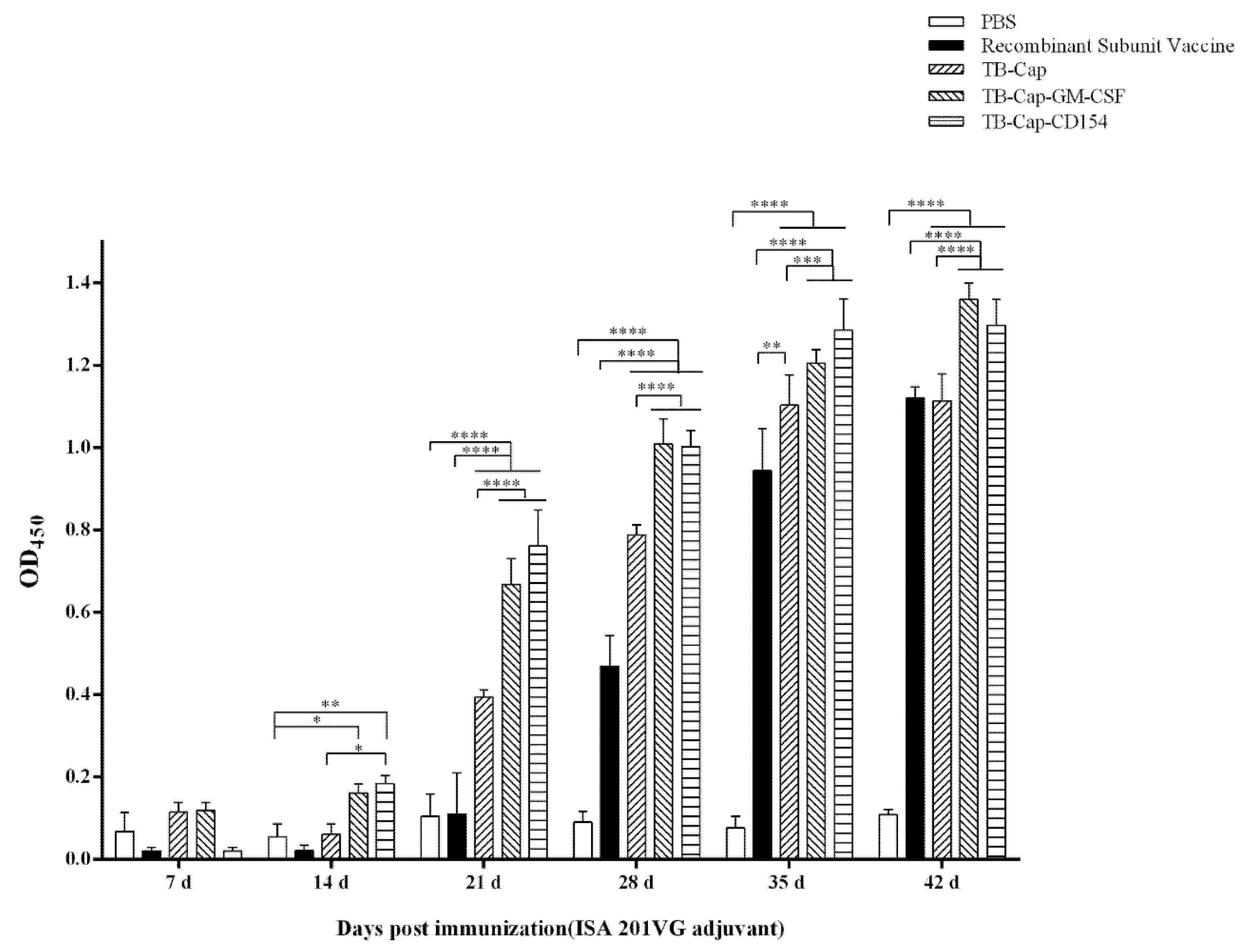
3.4. Humoral Immune Response
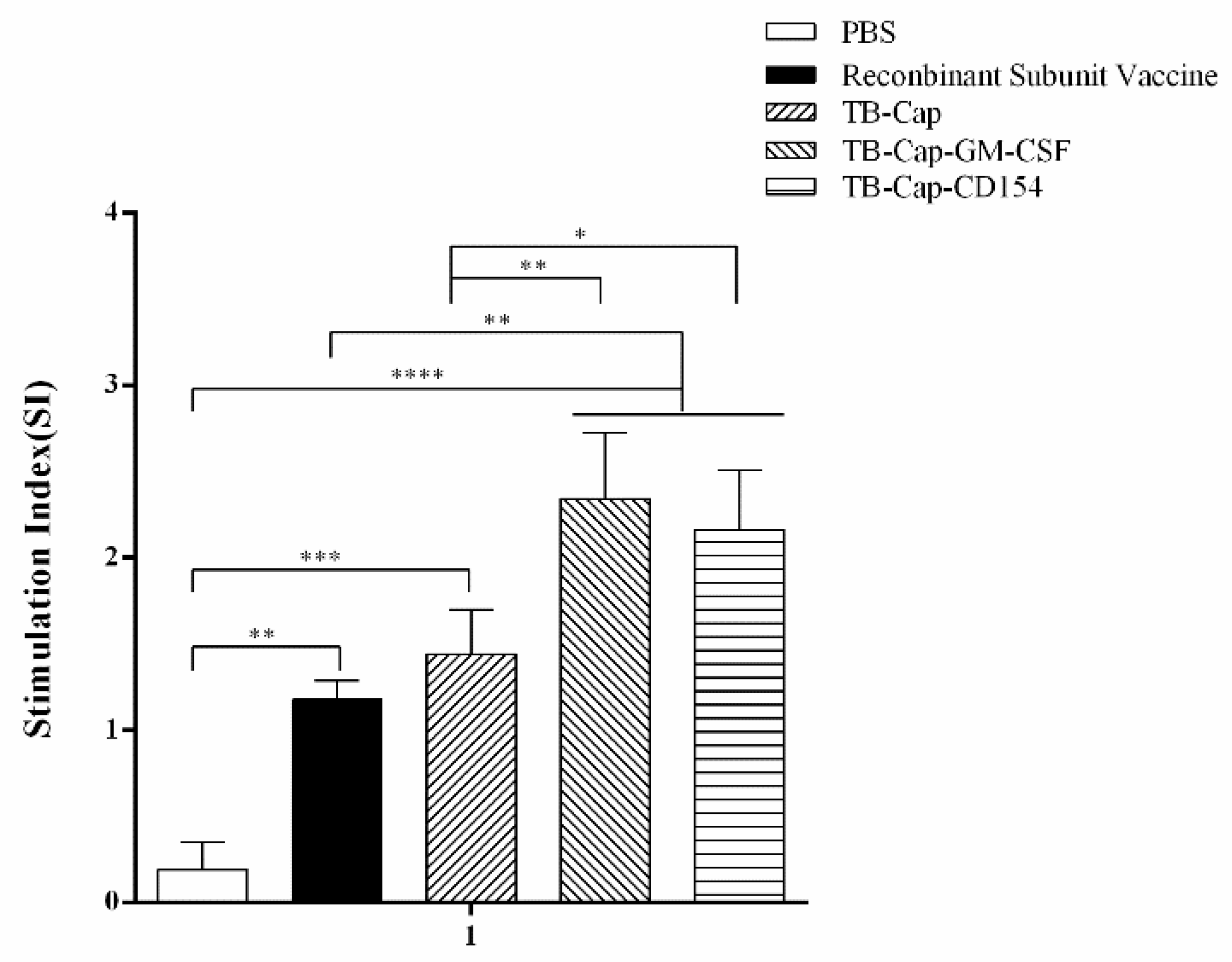
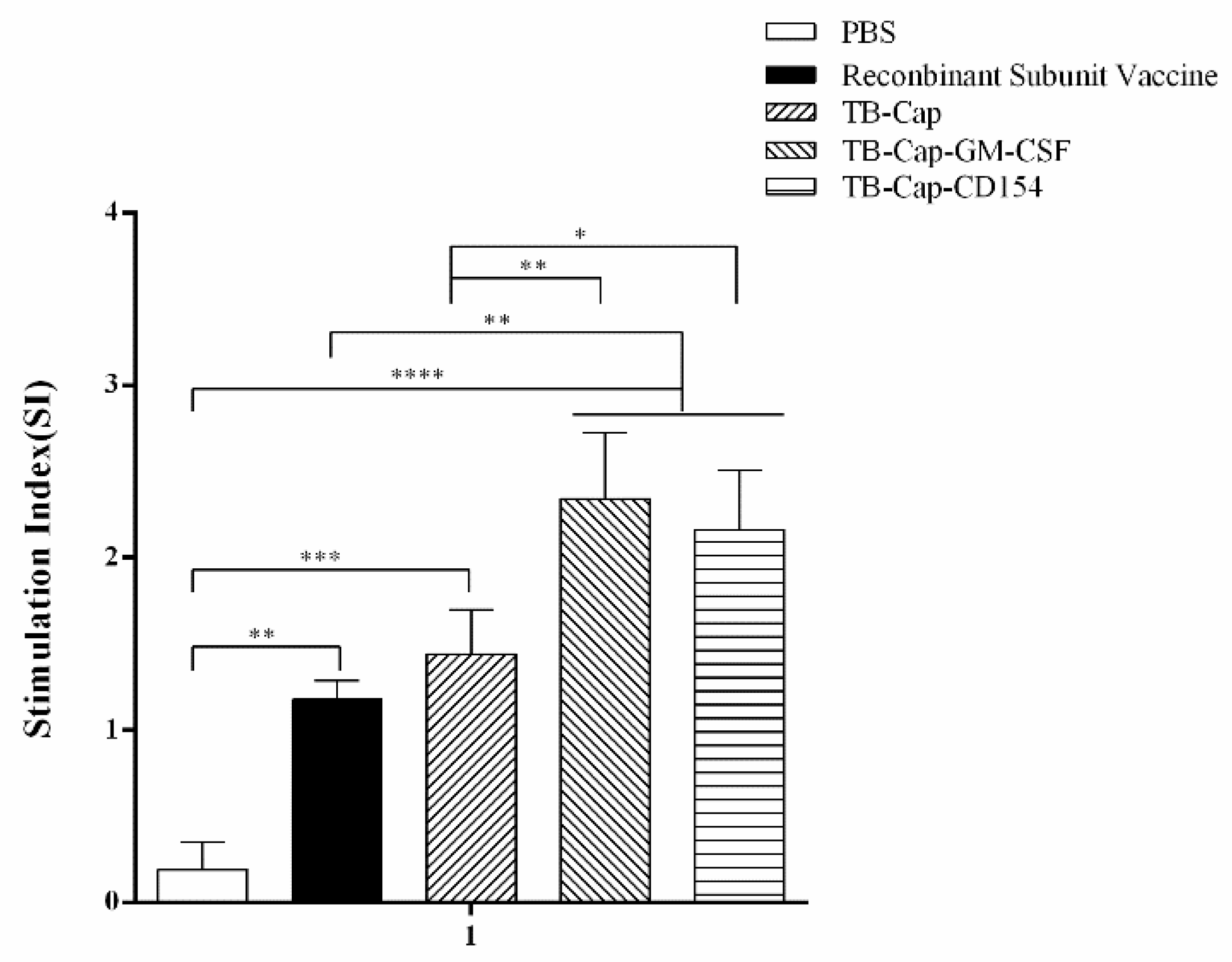
3.5. Lymphocyte Proliferative Activity
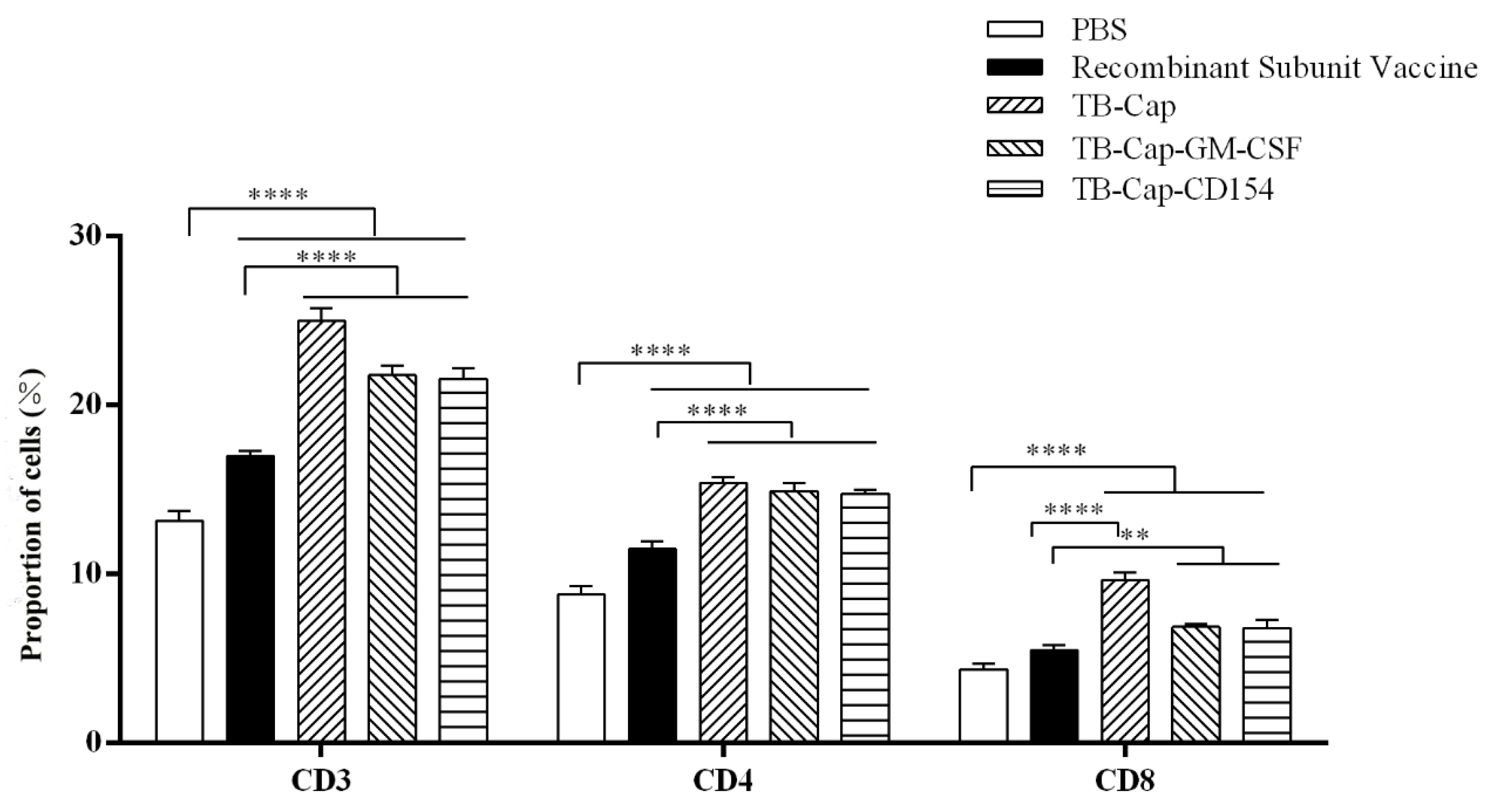
3.6. Detection of CD4+ T and CD8+ T Analysis by Flow Cytometry
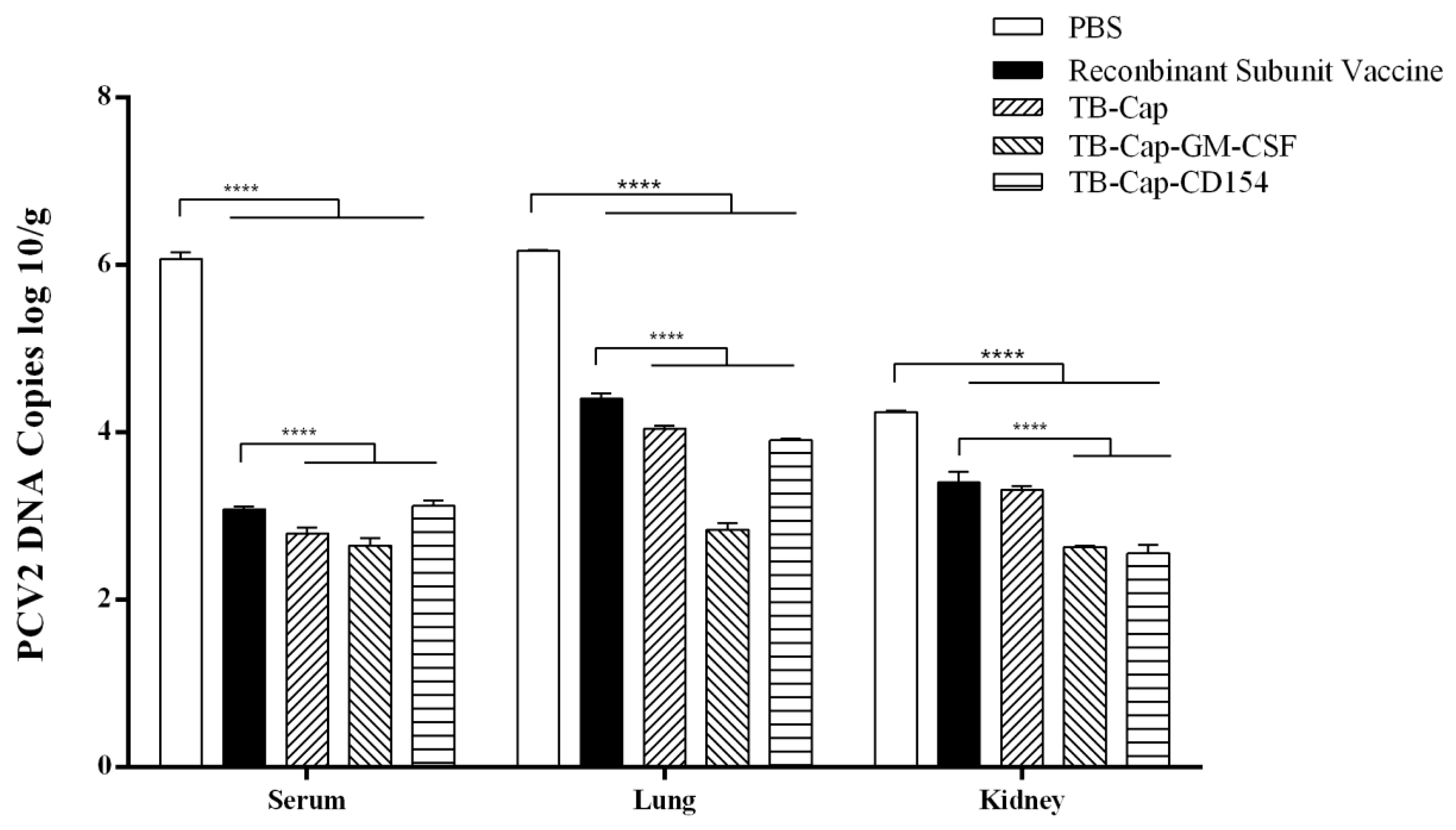
3.7. Immune Protection against PCV2 Challenge
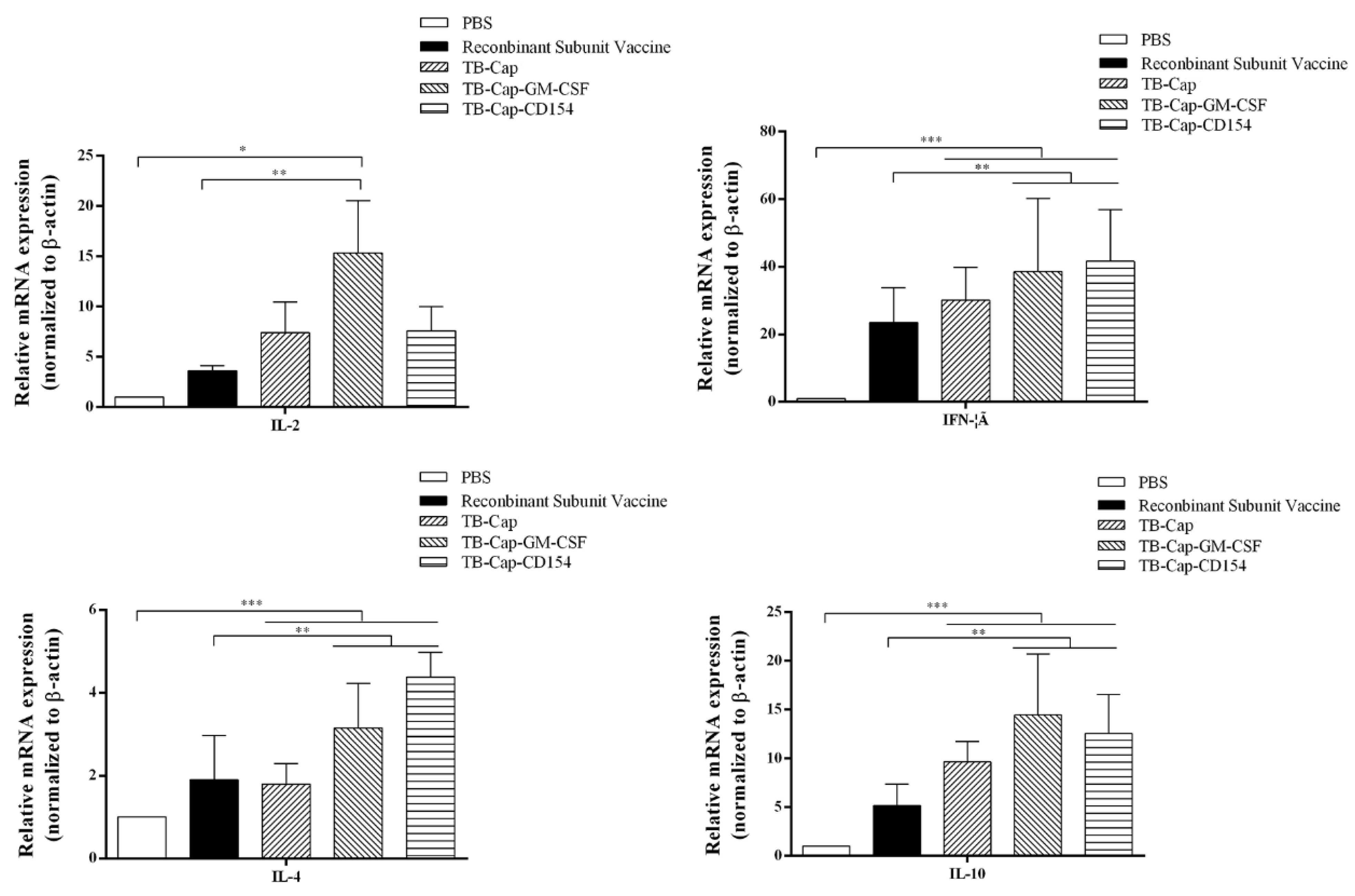
3.8. Analysis of IL-2, IFN-Gamma, IL-4 and IL-10
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-Infection of Swine with Porcine Circovirus Type 2 and Other Swine Viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, T.; Li, J.; Liu, C. A radical form of nitric oxide inhibits porcine circovirus type 2 replication in vitro. BMC Vet. Res. 2019, 15, 47. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Zhao, Y.; Cui, J.; Zheng, H.; Xu, T.; Hou, C.; Wang, Z.; Li, X.; Zheng, L.; Chen, H. Molecular detection and phylogenetic analysis of Porcine circovirus 4 in Henan and Shanxi Provinces of China. Transbound. Emerg. Dis. 2020, 68, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Huang, Y.; Wang, T.; Zhang, X.; Chen, Y.; Cui, B.; Li, D.; Zhao, X.; Zhang, W.; Chang, L.; et al. Porcine circovirus type 2 activates PI3K/Akt and p38 MAPK pathways to promote interleukin-10 production in macrophages via Cap interaction of gC1qR. Oncotarget 2016, 7, 17492–17507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Bermudez, D.S.; Diaz, A.; Mogollon, J.D.; Jaime, J. Longitudinal comparison of the humoral immune response and viral load of Porcine Circovirus Type 2 in pigs with different vaccination schemes under field conditions. F1000Research 2018, 7, 42. [Google Scholar] [CrossRef]
- Kotoucek, J.; Hezova, R.; Vrablikova, A.; Hubatka, F.; Kulich, P.; Macaulay, S.; Roessner, D.; Raska, M.; Psikal, I.; Turanek, J. Characterization and purification of pentameric chimeric protein particles using asymmetric flow field-flow fractionation coupled with multiple detectors. Anal. Bioanal. Chem. 2021, 413, 3749–3761. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, J.; Chen, H.-T.; Zhou, J.-H.; Ma, L.-N.; Ding, Y.-Z.; Liu, Y.-S. Research in advance for FMD Novel Vaccines. Virol. J. 2011, 8, 268. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Li, G.; Zheng, W.; Shu, J.; Chen, J.; Yang, F.; Wu, Y.; He, Y. Development of a Combined Genetic Engineering Vaccine for Porcine Circovirus Type 2 and Mycoplasma Hyopneumoniae by a Baculovirus Expression System. Int. J. Mol. Sci. 2019, 20, 4425. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Luo, Y.; Zhang, Y.; Guo, K. Enhanced protective immune response to PCV2 adenovirus vaccine by fusion expression of Cap protein with InvC in pigs. J. Vet. Sci. 2019, 20, e35. [Google Scholar] [CrossRef]
- Zhang, H.; Qian, P.; Peng, B.; Shi, L.; Chen, H.; Li, X. A novel subunit vaccine co-expressing GM-CSF and PCV2b Cap protein enhances protective immunity against porcine circovirus type 2 in piglets. Vaccine 2015, 33, 2449–2456. [Google Scholar] [CrossRef]
- Liu, J.; Wei, C.; Dai, A.; Lin, Z.; Fan, K.; Fan, J.; Liu, J.; Luo, M.; Yang, X. Detection of PCV2e strains in Southeast China. PeerJ 2018, 6, e4476. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Huang, Y.; Du, Q.; Wang, Z.; Chang, L.; Zhao, X.; Tong, D. CD40 Ligand and GMCSF Coexpression Enhance the Immune Responses and Protective Efficacy of PCV2 Adenovirus Vaccine. Viral Immunol. 2016, 29, 148–158. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Yang, W.-C.; Chang, Y.-K.; Wang, C.-Y.; Huang, W.-R.; Li, J.-Y.; Chuang, K.-P.; Wu, H.-Y.; Chang, C.-D.; Nielsen, B.L.; et al. Construction of polycistronic baculovirus surface display vectors to express the PCV2 Cap(d41) protein and analysis of its immunogenicity in mice and swine. Vet. Res. 2020, 51, 112. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.A.; Mendoza, R.B.; Kipps, T.J. Plasmids encoding granulocyte–macrophage colony-stimulating factor and CD154 enhance the immune response to genetic vaccines. Vaccine 2001, 19, 2181–2189. [Google Scholar] [CrossRef]
- Noh, K.H.; Park, Y.M.; Kim, H.S.; Kang, T.H.; Song, K.-H.; Lee, Y.-H.; Byeon, Y.; Jeon, H.N.; Jung, I.D.; Shin, B.C.; et al. GM-CSF-loaded chitosan hydrogel as an immunoadjuvant enhances antigen-specific immune responses with reduced toxicity. BMC Immunol. 2014, 15, 48. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.A.; Aleyas, A.; George, J.A.; Park, S.O.; Han, Y.W.; Lee, J.H.; Cho, J.-G.; Eo, S.K. Cytokine GM-CSF Genetic Adjuvant Facilitates Prophylactic DNA Vaccine against Pseudorabies Virus through Enhanced Immune Responses. Microbiol. Immunol. 2006, 50, 83–92. [Google Scholar] [CrossRef]
- Bracci, L.; Fragale, A.; Gabriele, L.; Moschella, F. Towards a Systems Immunology Approach to Unravel Responses to Cancer Immunotherapy. Front. Immunol. 2020, 11, 582744. [Google Scholar] [CrossRef]
- Chen, X.; Bai, Y.; Zaro, J.L.; Shen, W.-C. Design of an in vivo cleavable disulfide linker in recombinant fusion proteins. BioTechniques 2010, 49, 513–518. [Google Scholar] [CrossRef] [Green Version]
- Erickson, A.; Fisher, M.; Furukawa-Stoffer, T.; Ambagala, A.; Hodko, D.; Pasick, J.; King, D.; Nfon, C.; Polo, R.O.; Lung, O. A multiplex reverse transcription PCR and automated electronic microarray assay for detection and differentiation of seven viruses affecting swine. Transbound. Emerg. Dis. 2017, 65, e272–e283. [Google Scholar] [CrossRef] [PubMed]
- Papatsiros, V. Ear necrosis syndrome in weaning pigs associated with PCV2 infection: A case report. Vet. Res. Forum 2012, 3, 217–220. [Google Scholar] [PubMed]
- Perlman, S.; Vijay, R. Middle East respiratory syndrome vaccines. Int. J. Infect. Dis. 2016, 47, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Brandsma, J.L.; Shlyankevich, M.; Su, Y.; Zelterman, D.; Rose, J.K.; Buonocore, L. Reversal of papilloma growth in rabbits therapeutically vaccinated against E6 with naked DNA and/or vesicular stomatitis virus vectors. Vaccine 2010, 28, 8345–8351. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Zhang, C.; Wang, J.; Wei, L.; Quan, R.; Yang, J.; Yan, X.; Li, Z.; She, R.; Hu, F.; et al. Immunity Elicited by an Experimental Vaccine Based on Recombinant Flagellin-Porcine Circovirus Type 2 Cap Fusion Protein in Piglets. PLoS ONE 2016, 11, e0147432. [Google Scholar] [CrossRef]
- Ouyang, T.; Liu, X.-H.; Ouyang, H.-S.; Ren, L.-Z. Mouse models of porcine circovirus 2 infection. Anim. Model. Exp. Med. 2018, 1, 23–28. [Google Scholar] [CrossRef]
- Meillón-García, L.A.; García-Chávez, J.; Gómez-Almaguer, D.; Gutiérrez-Espíndola, G.R.; Martínez-Murillo, C. Primary immune thrombocytopenia in adults in Mexico: National characteristics and the relation to international literature. Gac. Med. Mex. 2014, 150, 279–288. [Google Scholar]
- Swinkels, M.; Rijkers, M.; Voorberg, J.; Vidarsson, G.; Leebeek, F.W.G.; Jansen, A.J.G. Emerging Concepts in Immune Thrombocytopenia. Front. Immunol. 2018, 9, 880. [Google Scholar] [CrossRef] [PubMed]
- Senhaji, N.; Kojok, K.; Darif, Y.; Fadainia, C.; Zaid, Y. The Contribution of CD40/CD40L Axis in Inflammatory Bowel Disease: An Update. Front. Immunol. 2015, 6, 529. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.R.; Suzuki, M. Recent advances in oncolytic adenovirus therapies for cancer. Curr. Opin. Virol. 2016, 21, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babai, I.; Samira, S.; Barenholz, Y.; Zakay-Rones, Z.; Kedar, E. A novel influenza subunit vaccine composed of liposome-encapsulated haemagglutinin/neuraminidase and IL-2 or GM-CSF. I. Vaccine characterization and efficacy studies in mice. Vaccine 1999, 17, 1223–1238. [Google Scholar] [CrossRef]
- Liu, X.; Ouyang, T.; Ma, T.; Ouyang, H.; Pang, D.; Ren, L. Immunogenicity evaluation of inactivated virus and purified proteins of porcine circovirus type 2 in mice. BMC Vet. Res. 2018, 14, 137. [Google Scholar] [CrossRef]
- Kar, T.; Narsaria, U.; Basak, S.; Deb, D.; Castiglione, F.; Mueller, D.M.; Srivastava, A.P. A candidate multi-epitope vaccine against SARS-CoV-2. Sci. Rep. 2020, 10, 10895. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.-F.; Shi, J.-R.; Huang, Y.; Wang, S.-N. A20 inhibits lipopolysaccharide-induced inflammation in enterocytes. World J. Gastrointest. Pharmacol. Ther. 2016, 7, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cao, T.-T.; Wei, Z.-G.; Zhang, Y.-Q. Silk Sericin Hydrolysate is a Potential Candidate as a Serum-Substitute in the Culture of Chinese Hamster Ovary and Henrietta Lacks Cells. J. Insect Sci. 2019, 19, 10. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epitopes | At the Cap/Rep Protein Position | Amino Acid Sequence |
|---|---|---|
| B 1 | C61~85 | TVRTPSWAVDMMRFNIDDFVPPGGG |
| B 2 | C133~131 | QGDRGVGSTAVILDDNFVT |
| B 3 | C169~180 | STIDYFQPNNKR |
| B 4 | C192~202 | NVDHVGLGTAF |
| R 1 | R81~100 | CHIEKAKGTDQQNKEYCSKE |
| R 2 | R201~220 | KWWDGYHGEEVVVZDDFYGW |
| Gene | Sequence (5′–3′) | Size (bp) |
|---|---|---|
| β-actin | F: TGCTGTCCCTGTATGCCTCTG | 100 |
| R: CTTTGATGTCACGCACGATTTC | ||
| IL-2 | F: GCTCTACAGCGGAAGCACAG | 382 |
| R: CATCTCCTCAGAAAGTCCACCAC | ||
| IFN-gamma | F: CTCAAGTGGCATAGATGTGGAAG | 268 |
| R: CTGGACCTGTGGGTTGTTGAC | ||
| IL-4 | F: GTCATCCTGCTCTTCTTTCTCG | 379 |
| R: TGATGCTCTTTAGGCTTTCCAG | ||
| IL-10 | F: ACAACATACTGCTAACCGACTCC | 296 |
| R: TTCATTCATGGCCTTGTAGACAC |
| Group | CD3 (%) | CD4+ (%) | CD8+ (%) | CD4+/CD8+ Ratio |
|---|---|---|---|---|
| PBS | 13.17 (±0.34) | 8.81 (±0.28) | 4.36 (±0.21) | 2.02 (±0.03) |
| Recombinant subunit vaccine | 16.99 (±0.17) | 11.50 (±0.27) | 5.49 (±0.17) | 2.09 (±0.01) |
| TB-Cap | 25.02 (±0.43) | 15.38 (±0.21) | 9.64 (±0.26) | 1.60 (±0.03) |
| TB-Cap-CD154 | 21.56 (±0.37) | 14.76 (±0.13) | 6.80 (±0.29) | 2.17 (±0.07) |
| TB-Cap-GM-CSF | 21.78 (±0.33) | 14.9 (±0.29) | 6.88 (±0.09) | 2.17 (±0.01) |
| Group | Day 14 after the PCV2 Challenge | Day 28 after the PCV2 Challenge | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | Positive rate | 1 | 2 | 3 | 4 | 5 | Positive rate | |
| PBS | + | + | + | + | + | 100 | + | + | + | + | + | 100 |
| Recombinant subunit vaccine | − | − | − | + | − | 20 | − | − | − | + | − | 20 |
| TB-Cap | − | − | − | − | − | 0 | − | − | − | − | − | 0 |
| TB-Cap-CD154 | − | − | − | − | − | 0 | − | − | − | − | − | 0 |
| TB-Cap-GM-CSF | − | − | − | − | − | 0 | − | − | − | − | − | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, Q.; Zhang, W.; Ma, S.; Qiu, Z.; Li, B.; Xu, C.; He, H.; Fan, S.; Wu, K.; Chen, J.; et al. Fusion Expression and Immune Effect of PCV2 Cap Protein Tandem Multiantigen Epitopes with CD154/GM-CSF. Vet. Sci. 2021, 8, 211. https://doi.org/10.3390/vetsci8100211
Mao Q, Zhang W, Ma S, Qiu Z, Li B, Xu C, He H, Fan S, Wu K, Chen J, et al. Fusion Expression and Immune Effect of PCV2 Cap Protein Tandem Multiantigen Epitopes with CD154/GM-CSF. Veterinary Sciences. 2021; 8(10):211. https://doi.org/10.3390/vetsci8100211
Chicago/Turabian StyleMao, Qian, Weijian Zhang, Shengming Ma, Zilong Qiu, Bingke Li, Chen Xu, Huangyu He, Shuangqi Fan, Keke Wu, Jinding Chen, and et al. 2021. "Fusion Expression and Immune Effect of PCV2 Cap Protein Tandem Multiantigen Epitopes with CD154/GM-CSF" Veterinary Sciences 8, no. 10: 211. https://doi.org/10.3390/vetsci8100211
APA StyleMao, Q., Zhang, W., Ma, S., Qiu, Z., Li, B., Xu, C., He, H., Fan, S., Wu, K., Chen, J., & Zhao, M. (2021). Fusion Expression and Immune Effect of PCV2 Cap Protein Tandem Multiantigen Epitopes with CD154/GM-CSF. Veterinary Sciences, 8(10), 211. https://doi.org/10.3390/vetsci8100211