
VEGF-B, VEGF-A, FLT-1, KDR, ERBB2, EGFR, GRB2, RAC1, CDH1 and HYAL-1 Genes Expression Analysis in Canine Mammary Gland Tumors and the Association with Tumor ClinicoPathological Parameters and Dog Breed Assessment

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Histopathology
2.3. Immunohistochemistry and PI Evaluation
2.4. Tissue sample Preparation and RNA Extraction
2.5. Reverse Transcription and Quantitative Real-time Polymerase Chain Reaction (qRT-PCR)
2.6. Statistical Analysis
3. Results
3.1. Patients and Tumors Characteristics
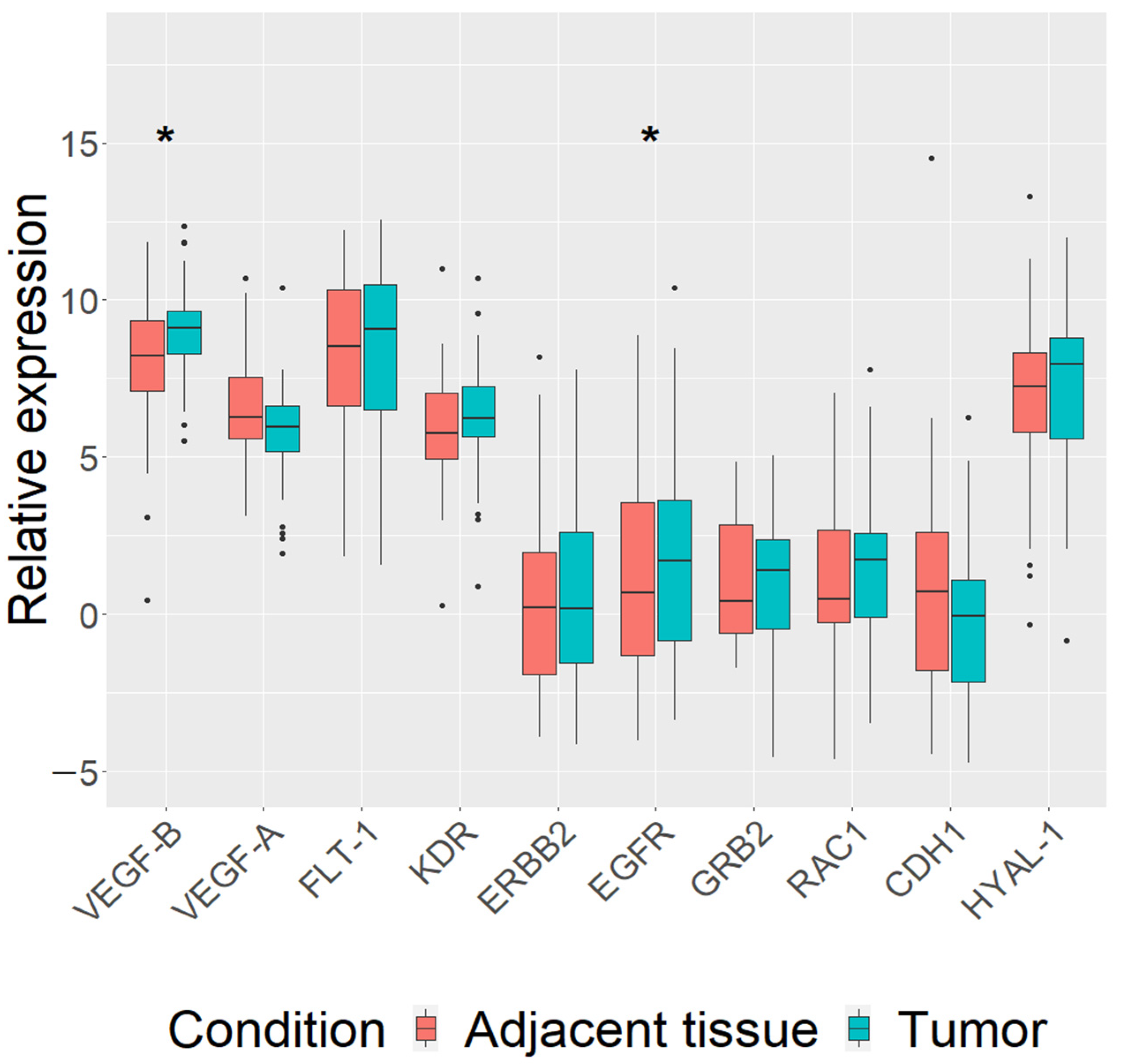
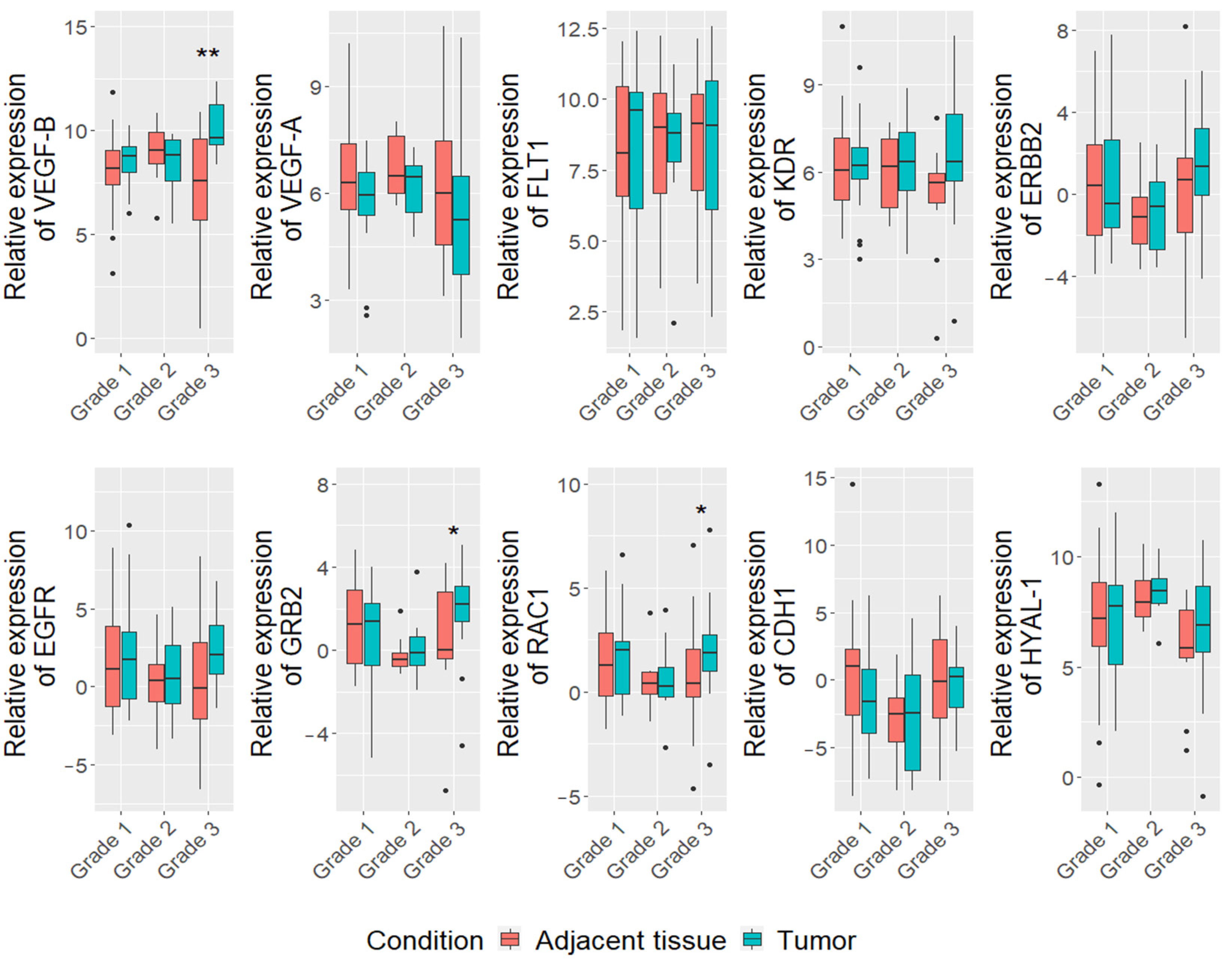
3.2. Differentially Expressed Genes in Mammary Carcinoma Tissue of All Breed Dogs
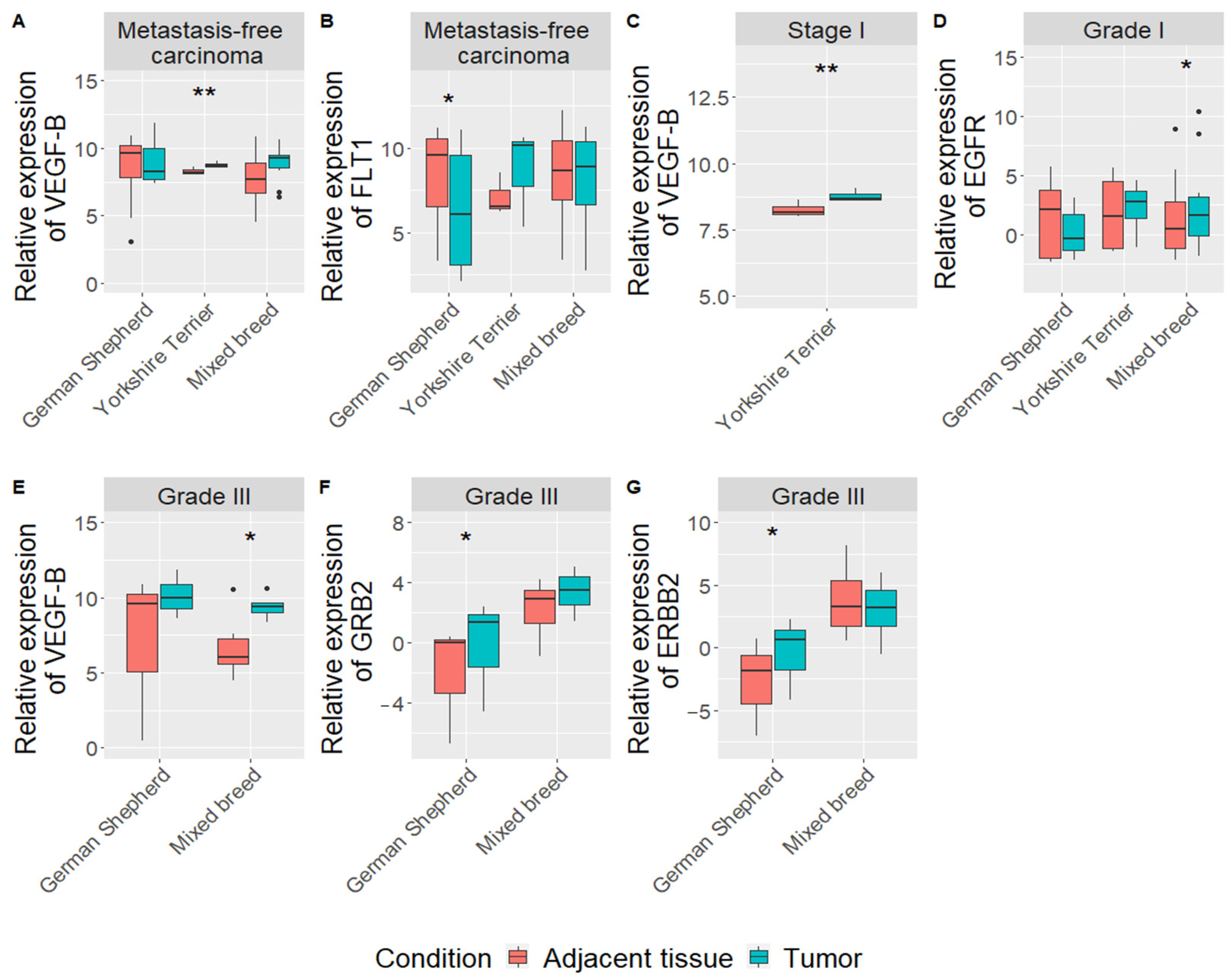
3.3. Gene Expression in Mammary Gland Carcinomas According to the Dog Breed
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Misdorp, W. Tumors of the mammary gland. In Tumors in Domestic Animals, 4th ed.; Meuten, D.J., Ed.; Iowa State Press: Ames, IA, USA, 2002; pp. 575–606. [Google Scholar]
- Gobar, G.M.; Case, J.T.; Kass, P.H. Program for surveillance of causes of death of dogs, using the Internet to survey small animal veterinarians. J. Am. Vet. Med. Assoc. 1998, 213, 251–256. [Google Scholar]
- Rutteman, G.R.; Withrow, S.J.; MacEwen, E.G. Tumors of the mammary gland. In Small Animal Clinical Oncology, 4th ed.; Withrow, S.J., MacEwen, D.R., Eds.; WB Saunders Company: Philadelphia, PA, USA, 2001; pp. 455–477. [Google Scholar]
- Gundim, L.F.; de Araújo, C.P.; Blanca, W.T.; Guimarães, E.C.; Medeiros, A.A. Clinical staging in bitches with mammary tumors: Influence of type and histological grade. Can. J. Vet. Res. 2016, 80, 318–322. [Google Scholar] [PubMed]
- Chang, S.C.; Chang, C.C.; Chang, T.J.; Wong, M.L. Prognostic factors associated with survival two years after surgery in dogs with malignant mammary tumors: 79 cases (1998–2002). J. Am. Vet. Med. Assoc. 2005, 227, 1625–1629. [Google Scholar] [CrossRef] [PubMed]
- Karayannopoulou, M.; Kaldrymidou, E.; Constantinidis, T.C.; Dessiris, A. Histological Grading and Prognosis in Dogs with Mammary Carcinomas: Application of a Human Grading Method. J. Comp. Pathol. 2005, 133, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Rasotto, R.; Berlato, D.; Goldschmidt, M.H.; Zappulli, V. Prognostic significance of canine mammary tumor histologic subtypes: An observational cohort study of 229 cases. Vet. Pathol. 2017, 54, 571–578. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Durham, A.C.; Kristiansen, V.; Pena, L.; Goldschmidt, M.H.; Stefanovski, D. Developing and testing prognostic bio-scoring systems for canine mammary gland carcinomas. Vet. Comp. Oncol. 2019, 17, 479–488. [Google Scholar] [CrossRef]
- Pena, L.L.; Nieto, A.I.; Perez-Alenza, D.; Cuesta, P.; Castano, M. Immunohistochemical detection of Ki-67 and PCNA in canine mammary tumours: Relationship to clinical and pathologic variables. J. Vet. Diagn. Investig. 1998, 10, 237–246. [Google Scholar] [CrossRef]
- Abdelmegeed, S.M.; Mohammed, S. Canine mammary tumors as a model for human disease. Oncol. Lett. 2018, 15, 8195–8205. [Google Scholar] [CrossRef]
- Huskey, A.L.W.; Goebel, K.; Lloveras-Fuentes, C.; McNeely, I.; Merner, N.D. Whole genome sequencing for the investigation of canine mammary tumor inheritance—An initial assessment of high-risk breast cancer genes reveal BRCA2 and STK11 variants potentially associated with risk in purebred dogs. Canine Genet. Epidemiol. 2020, 7, 1–13. [Google Scholar] [CrossRef]
- Goebel, K.; Merner, N.D. A monograph proposing the use of canine mammary tumours as a model for the study of hereditary breast cancer susceptibility genes in humans. Vet. Med. Sci. 2017, 3, 51–62. [Google Scholar] [CrossRef]
- Salas, Y.; Márquez, A.; Diaz, D.; Romero, L. Epidemiological Study of Mammary Tumors in Female Dogs Diagnosed during the Period 2002–2012: A Growing Animal Health Problem. PLoS ONE 2015, 10, e0127381. [Google Scholar] [CrossRef]
- Dobson, J.M. Breed-predispositions to cancer in pedigree dogs. ISRN Vet. Sci. 2013, 17, 941275. [Google Scholar] [CrossRef] [PubMed]
- Ping, J.; Guo, X.; Ye, F.; Long, J.; Lipworth, L.; Cai, Q.; Blot, W.; Shu, X.; Zheng, W. Differences in gene-expression profiles in breast cancer between African and European-ancestry women. Carcinogenesis 2020, 41, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Parada, H.; Sun, X.; Fleming, J.M.; Williams-DeVane, C.R.; Kirk, E.L.; Olsson, L.T.; Perou, C.M.; Olshan, A.F.; Troester, M.A. Race-associated biological differences among luminal A and basal-like breast cancers in the Carolina Breast Cancer Study. Breast Cancer Res. 2017, 19, 29228969. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Lim, H.Y.; Shin, J.I.; Seung, B.J.; Ju, J.H.; Sur, J.H. Breed- and age-related differences in canine mammary tumors. Can. J. Vet. Res. 2016, 80, 146–155. [Google Scholar] [PubMed]
- Ortega, M.A.; Fraile-Martínez, O.; Asúnsolo, Á.; Buján, J.; García-Honduvilla, N.; Coca, S. Signal Transduction Pathways in Breast Cancer: The Important Role of PI3K/Akt/mTOR. J. Oncol. 2020, 2020, 9258396. [Google Scholar] [CrossRef] [PubMed]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. PI3K pathway in human disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef]
- Jiang, B.H.; Liu, L.Z. PI3K/PTEN signaling in angiogenesis and tumorigenesis. Adv. Cancer Res. 2009, 102, 19–65. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.; Niu, X.; Yin, L.; Wang, Y.; Gu, Y.; Li, D.; Zhang, H.; Lu, M.; Zhang, F.; et al. Sodium-Dependent Glucose Transporter 1 (SGLT1) Stabled by HER2 Promotes Breast Cancer Cell Proliferation by Activation of the PI3K/Akt/mTOR Signaling Pathway in HER2+ Breast Cancer. Dis. Markers 2020, 2020, 32377271. [Google Scholar] [CrossRef]
- Ke, K.; Lou, T. MicroRNA-10a suppresses breast cancer progression via PI3K/Akt/mTOR pathway. Oncol. Lett. 2017, 14, 5994–6000. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Zhang, W.; Lu, S.; Ren, H.; Guo, Y. NRSN2 promotes breast cancer metastasis by activating PI3K/AKT/mTOR and NF-κB signaling pathways. Oncol. Lett. 2020, 19, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Zhou, J.; Luo, L.P.; Han, B.; Li, F.; Chen, J.Y.; Zhu, Y.F.; Chen, W.; Yu, X.P. Black rice anthocyanins suppress metastasis of breast cancer cells by targeting RAS/RAF/MAPK pathway. Biomed. Res. Int. 2015, 2015, 414250. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zheng, Z.; Chen, L.; Zheng, H. MAPK, NFKB, and VEGF signaling pathways regulate breast cancer liver metastasis. Oncotarget 2017, 8, 101452–101460. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Mai, H.; Zhu, Y.; Li, G.; Sun, J.; Li, G.; Liang, B.; Chen, S. MicroRNA-4500 inhibits migration, invasion, and angiogenesis of breast cancer cells via RRM2-dependent MAPK signaling pathway. Mol. Ther. Nucleic Acids 2020, 21, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Asproni, P.; Millanta, F.; Ressel, L.; Podestá, F.; Parisi, F.; Vannozzi, I.; Poli., A. An Immunohistochemical Study of the PTEN/AKT Pathway Involvement in Canine and Feline Mammary Tumors. Animals 2021, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Uva, P.; Aurisicchio, L.; Watters, J.; Loboda, A.; Kulkarni, A.; Castle, J.; Palombo, F.; Viti, V.; Mesiti, G.; Zappulli, V.; et al. Comparative expression pathway analysis of human and canine mammary tumors. BMC Genom. 2009, 10, 135. [Google Scholar] [CrossRef]
- Wong, A.S.; Gumbiner, B.M. Adhesion-independent mechanism for suppression of tumor cell invasion by E-cadherin. J. Cell Biol. 2003, 161, 191–1203. [Google Scholar] [CrossRef]
- Huang, R.; Ding, P.; Yang, F. Clinicopathological significance and potential drug target of CDH1 in breast cancer: A meta-analysis and literature review. Drug Des. Dev. Ther. 2015, 9, 5277–5285. [Google Scholar]
- Matos, A.J.; Lopes, C.; Carvalheira, J.; Santos, M.; Rutteman, G.R.; Gartner, F. E-cadherin expression in canine malignant mammary tumours: Relationship to other clinico-pathological variables. J. Comp. Pathol. 2006, 134, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.J.; Lopes, C.C.; Faustino, A.M.; Carvalheira, J.G.; Rutteman, G.R.; Gärtner Mde, F. E-cadherin, beta-catenin, invasion and lymph node metastases in canine malignant mammary tumours. Apmis 2007, 115, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Yoshida, S.; Choisunirachon, N.; Saito, T.; Matsumoto, K.; Saeki, K.; Mochizuki, M.; Nishimura, R.; Sasaki, N.; Nakagawa, T. The relationship between clinicopathological features and expression of epithelial and mesenchymal markers in spontaneous canine mammary gland tumors. J. Vet. Sci. 2014, 76, 1321–1327. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.X.; Wang, X.Y.; Li, H.Y.; Su, X.L.; Wang, L.; Ran, L.; Zheng, K.; Ren, G.S. HYAL1 overexpression is correlated with the malignant behavior of human breast cancer. Int. J. Cancer 2011, 128, 1303–1315. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.X.; Wang, X.Y.; Su, X.L.; Li, H.Y.; Shi, Y.; Wang, L.; Ren, G.S. Upregulation of HYAL1 expression in breast cancer promoted tumor cell proliferation, migration, invasion and angiogenesis. PLoS ONE 2011, 6, e22836. [Google Scholar] [CrossRef] [PubMed]
- McAtee, C.O.; Barycki, J.J.; Simpson, M.A. Emerging roles for hyaluronidase in cancer metastasis and therapy. Adv. Cancer Res. 2014, 123, 1–34. [Google Scholar]
- Varallo, G.R.; Jardim-Perassi, B.V.; Alexandre, P.A.; Fukumasu, H.; Zuccari, D.A.P.C. Global gene expression profile in canine mammary carcinomas. Vet. J. 2019, 254, 105393. [Google Scholar] [CrossRef]
- Goldschmidt, M.; Peña, L.; Rasotto, R.; Zappulli, V. Classification and grading of canine mammary tumors. Vet. Pathol. 2011, 48, 117–131. [Google Scholar] [CrossRef]
- Elston, C.W.; Ellis, I.O. Pathological prognostic factors in breast cancer, I: The value of histological grade in breast cancer: Experience from a large study with long-term follow-up. Histopathology 1991, 19, 403–410. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 10 August 2021).
- Sorenmo, K.U.; Worley, D.R.; Goldschmidt, M.H. Mammary gland tumors in dogs. In Withrow & MacEwen’s Small Animal Clinical Oncology, 5th ed.; Withrow, S.J., Vail, D.M., Eds.; Saunders Elsevier: St. Louis, MO, USA, 2013; pp. 538–556. [Google Scholar]
- Varallo, G.R.; Gelaleti, G.B.; Maschio-Signorini, L.B.; Moschetta, M.G.; Lopes, J.R.; De Nardi, A.B.; Tinucci-Costa, M.; Rocha, R.M.; De Campos Zuccari, D.A.P. Prognostic phenotypic classification for canine mammary tumors. Oncol. Lett. 2019, 18, 6545–6553. [Google Scholar] [CrossRef]
- Carvalho, M.I.; Guimarães, M.J.; Pires, I.; Prada, J.; Silva-Carvalho, R.; Lopes, C.; Queiroga, F.L. EGFR and microvessel density in canine malignant mammary tumours. Res. Vet. Sci. 2013, 95, 1094–1099. [Google Scholar] [CrossRef]
- Gama, A.; Gartner, F.; Alves, A. Immunohistochemical expression of epidermal growth factor receptor (EGFR) in canine mammary tissues. Res. Vet. Sci. 2009, 87, 432–437. [Google Scholar] [CrossRef]
- Damasceno, K.A.; Ferreira, E.; Estrela-Lima, A.; Gamba, C.D.O.; Miranda, F.F.; Alves, M.R.; Rocha, R.M.; de Barros, A.L.; Cassali, G.D. HER-2 and EGFR mRNA Expression and Its Relationship with Versican in Malignant Matrix-Producing Tumors of the Canine Mammary Gland. PLoS ONE 2016, 11, e0160419. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, F.N.; Mendes, H.N.; Bacchi, C.E.; Carvalho, F.M. Comparison of nuclear grade and immunohistochemical features in situ and invasive components of ductal carcinoma of breast. Rev. Bras. Ginecol. Obstet. 2013, 35, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, Y.; Hosaka, K.; Andersson, P.; Wang, J.; Tholander, F.; Cao, Z.; Morikawa, H.; Tegnér, J.; Yang, Y.; et al. VEGF-B promotes cancer metastasis. Proc. Natl. Acad. Sci. USA 2015, 112, E2900–E2909. [Google Scholar] [CrossRef] [PubMed]
- Melonivici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)–Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Raposo-Ferreira, T.M.; Salvador, R.C.; Terra, E.M.; Ferreira, J.H.; Vechetti-Junior, I.J.; Tinucci-Costa, M.; Rogatto, S.R.; Laufer-Amorim, R. Evaluation of vascular endothelial growth factor gene and protein expression in canine metastatic mammary carcinomas. Microsc. Res. Tech. 2016, 79, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.W.; Lin, D.G.; Wang, J.Q.; Li, C.Y.; Deng, G.Z. Expression and significance of PTEN and VEGF in canine mammary gland tumours. Vet. Res. Commun. 2008, 32, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.A.Y.; Sayad, S.; Shahsavar, F.; Nekouian, R.; Panahi, M.; Sayad, S.; Boroujeni, M.B.; Akbari, S. Expression of Angiogenesis-related Genes in a Group of Iranian Cases of Breast Cancer. Curr. Pharm. Pers. Med. 2020, 17, 197–205. [Google Scholar]
- Gunningham, S.P.; Currie, M.J.; Han, C.; Robinson, B.A.; Scott, P.A.; Harris, A.L.; Fox, S.B. VEGF-B expression in human primary breast cancers is associated with lymph node metastasis but not angiogenesis. J. Pathol. 2001, 193, 325–332. [Google Scholar] [CrossRef]
- Rakha, E.A.; Abd El Rehim, D.; Pinder, S.E.; Lewis, S.A.; Ellis, I.O. E-cadherin expression in invasive non-lobular carcinoma of the breast and its prognostic significance. Histopathology 2005, 46, 685–693. [Google Scholar] [CrossRef]
- Canadas, A.; Santos, M.; Medeiros, R.; Dias-Pereira, P. Influence of E-cadherin genetic variation in canine mammary tumour risk, clinicopathological features and prognosis. Vet. Comp. Oncol. 2019, 17, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Timsah, Z.; Suen, K.M.; Cook, N.P.; Lee, G.R., IV; Lin, C.; Gagea, M.; Marti, A.A.; Ladbury, J.E. Grb2 monomer–dimer equilibrium determines normal versus oncogenic function. Nat. Commun. 2015, 6, 7354. [Google Scholar] [CrossRef] [PubMed]
- Melzer, C.; Hass, R.; von der Ohe, J.; Lehnert, H.; Ungefroren, H. The role of TGF-β and its crosstalk with RAC1/RAC1b signaling in breast and pancreas carcinoma. Cell Commun. Signal. 2017, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Dantas Cassali, G.; Cavalheiro Bertagnolli, A.; Ferreira, E.; Araújo Damasceno, K.; de Oliveira Gamba, C.; Bonolo de Campos, C. Canine mammary mixed tumours: A review. Vet. Med. Int. 2012, 2012, 274608. [Google Scholar] [CrossRef]
- Marei, H.; Malliri, A. Rac1 in human diseases: The therapeutic potential of targeting Rac1 signaling regulatory mechanisms. Small GTPases 2017, 8, 139–163. [Google Scholar] [CrossRef]
- Zhou, Y.; Liao, Q.; Han, Y.; Chen, J.; Liu, Z.; Ling, H.; Zhang, J.; Yang, W.; Oyang, L.; Xia, L.; et al. Rac1 overexpression is correlated with epithelial mesenchymal transition and predicts poor prognosis in non-small cell lung cancer. J. Cancer 2016, 7, 2100–2109. [Google Scholar] [CrossRef]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current biomarkers of canine mammary tumors. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef]
- Behelgardi, M.F.; Saber, Z.; Farhad, M.; Kamran, M.; Mohsen, A.S. A peptide mimicking the binding sites of VEGF-A and VEGF-B inhibits VEGFR-1/-2 driven angiogenesis, tumor growth and metastasis. Sci. Rep. 2018, 8, 17924. [Google Scholar] [CrossRef]
- Tari, A.; Hung, M.C.; Li, K.; Lopez-Berestein, G. Growth inhibition of breast cancer cells by Grb2 downregulation is correlated with inactivation of mitogen-activated protein kinase in EGFR, but not in ErbB2, cells. Oncogene 1999, 18, 1325–1332. [Google Scholar] [CrossRef][Green Version]
- Sareyeldin, R.M.; Gupta, I.; Al-Hashimi, I.; Al-Thawadi, H.A.; Al Farsi, H.F.; Vranic, S.; Al-Moustafa, A. Gene Expression and miRNAs Profiling: Function and Regulation in Human Epidermal Growth Factor Receptor 2 (HER2)-Positive Breast Cancer. Cancers 2019, 11, 646. [Google Scholar] [CrossRef]
- de Las Mulas, J.M.; Ordás, J.; Millán, Y.; Fernández-Soria, V.; Cajal, S.R. Oncogene HER-2 in canine mammary gland carcinomas: An immunohistochemical and chromogenic in situ hybridization study. Breast Cancer Res. Treat. 2003, 80, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Dutra, A.P.; Granja, N.V.M.; Schmitt, F.; Cassali, G.D. cerbB-2 expression and nuclear pleomorphism in canine mammary tumors. Braz. J. Med. Biol. Res. 2004, 37, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Copley, B.; Niu, Q.; Liu, F.; Johnson, J.A.; Sutton, T.; Khramtsova, G.; Sveen, E.; Yoshimatsu, T.F.; Zeng, Y.; et al. Racial disparities in survival outcomes among breast cancer patients by molecular subtypes. Breast Cancer Res. Treat. 2021, 185, 841–849. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Parameters | Number of Cases |
|---|---|
| Histological type | |
| Carcinoma-simple | 6 |
| Carcinoma-solid | 12 |
| Comedocarcinoma | 1 |
| Carcinoma-complex type | 11 |
| Carcinoma-mixed type | 9 |
| Intraductal papillary carcinoma | 3 |
| Squamous cell carcinoma | 1 |
| Malignant myoepithelioma | 2 |
| Inflammatory carcinoma | 3 |
| Grade of malignancy | |
| Grade I | 27 |
| Grade II | 8 |
| Grade III | 13 |
| Stage | |
| Stage I | 8 |
| Stage II | 5 |
| Stage III | 24 |
| Stage IV | 9 |
| Stage V | 2 |
| Metastasis | |
| Yes | 9 |
| No | 37 |
| Proliferation index | |
| Low (<14%) | 27 |
| High (≥14%) | 21 |
| Inflammation | |
| Absent | 38 |
| Present | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakalauskaitė, S.; Šaltenienė, V.; Nikitina, D.; Ugenskienė, R.; Riškevičienė, V.; Karvelienė, B.; Juodžiukynienė, N. VEGF-B, VEGF-A, FLT-1, KDR, ERBB2, EGFR, GRB2, RAC1, CDH1 and HYAL-1 Genes Expression Analysis in Canine Mammary Gland Tumors and the Association with Tumor ClinicoPathological Parameters and Dog Breed Assessment. Vet. Sci. 2021, 8, 212. https://doi.org/10.3390/vetsci8100212
Sakalauskaitė S, Šaltenienė V, Nikitina D, Ugenskienė R, Riškevičienė V, Karvelienė B, Juodžiukynienė N. VEGF-B, VEGF-A, FLT-1, KDR, ERBB2, EGFR, GRB2, RAC1, CDH1 and HYAL-1 Genes Expression Analysis in Canine Mammary Gland Tumors and the Association with Tumor ClinicoPathological Parameters and Dog Breed Assessment. Veterinary Sciences. 2021; 8(10):212. https://doi.org/10.3390/vetsci8100212
Chicago/Turabian StyleSakalauskaitė, Simona, Violeta Šaltenienė, Darja Nikitina, Rasa Ugenskienė, Vita Riškevičienė, Birutė Karvelienė, and Nomeda Juodžiukynienė. 2021. "VEGF-B, VEGF-A, FLT-1, KDR, ERBB2, EGFR, GRB2, RAC1, CDH1 and HYAL-1 Genes Expression Analysis in Canine Mammary Gland Tumors and the Association with Tumor ClinicoPathological Parameters and Dog Breed Assessment" Veterinary Sciences 8, no. 10: 212. https://doi.org/10.3390/vetsci8100212
APA StyleSakalauskaitė, S., Šaltenienė, V., Nikitina, D., Ugenskienė, R., Riškevičienė, V., Karvelienė, B., & Juodžiukynienė, N. (2021). VEGF-B, VEGF-A, FLT-1, KDR, ERBB2, EGFR, GRB2, RAC1, CDH1 and HYAL-1 Genes Expression Analysis in Canine Mammary Gland Tumors and the Association with Tumor ClinicoPathological Parameters and Dog Breed Assessment. Veterinary Sciences, 8(10), 212. https://doi.org/10.3390/vetsci8100212