
Antimicrobial Peptide Reduces Cytotoxicity and Inflammation in Canine Epidermal Keratinocyte Progenitor Cells Induced by Pseudomonas aeruginosa Infection


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Reagents
2.2. Peptide Synthesis
2.3. Evaluation of Direct Antibacterial and Antibiofilm Activities of Synthetic Peptides against P. aeruginosa PAO1
2.3.1. Effects of Antimicrobial Peptides on Planktonic P. aeruginosa PAO1
2.3.2. Biofilm Formation Assay
2.4. Effects of AMPs of Keratinocytes
2.4.1. Cell Culture
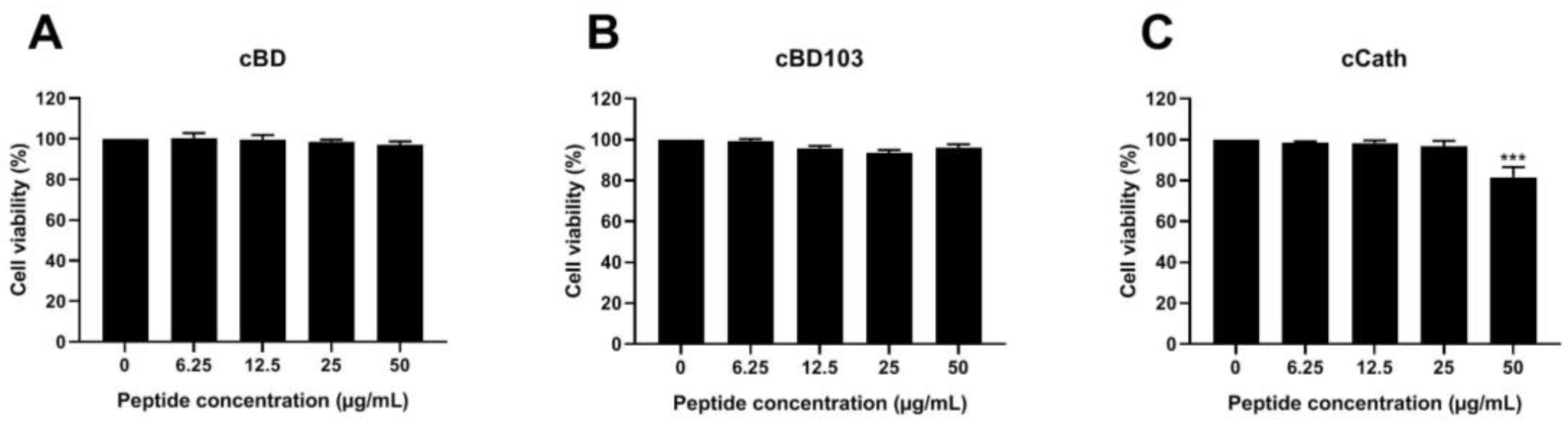
2.4.2. Cytotoxicity of AMPs to CPEK Cells
2.4.3. Effects of AMPs on the Cytotoxicity of P. aeruginosa
2.4.4. Determination of Cytokine Expression
2.5. Statistical Analysis
3. Results
3.1. AMPs Exerted Bactericidal Activity on Planktonic P. aeruginosa
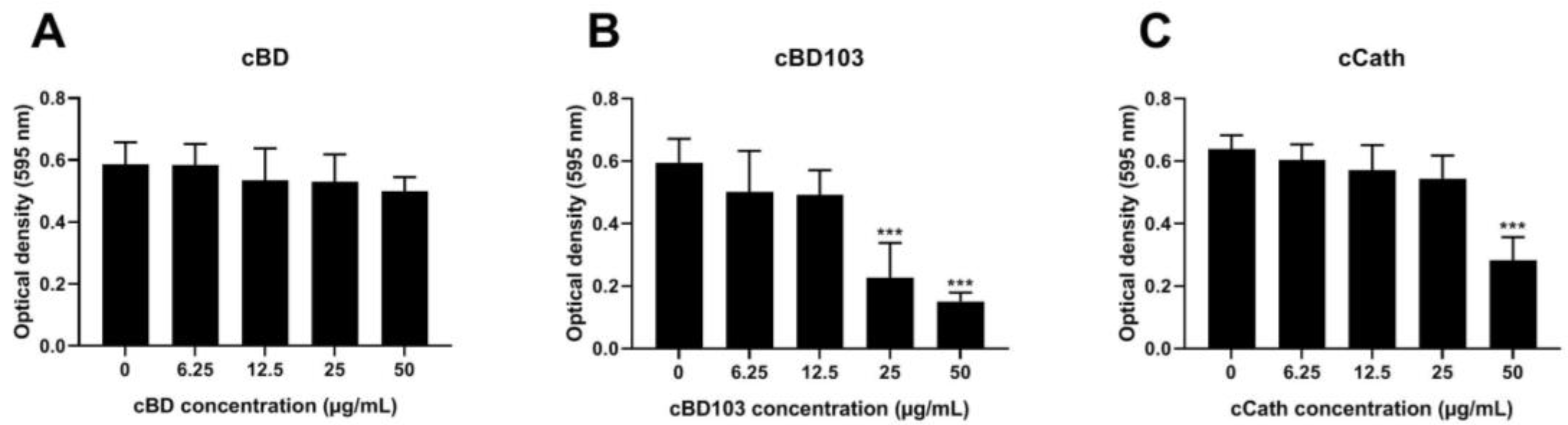
3.2. AMPs Suppressed Pseudomonal Biofilm Formation
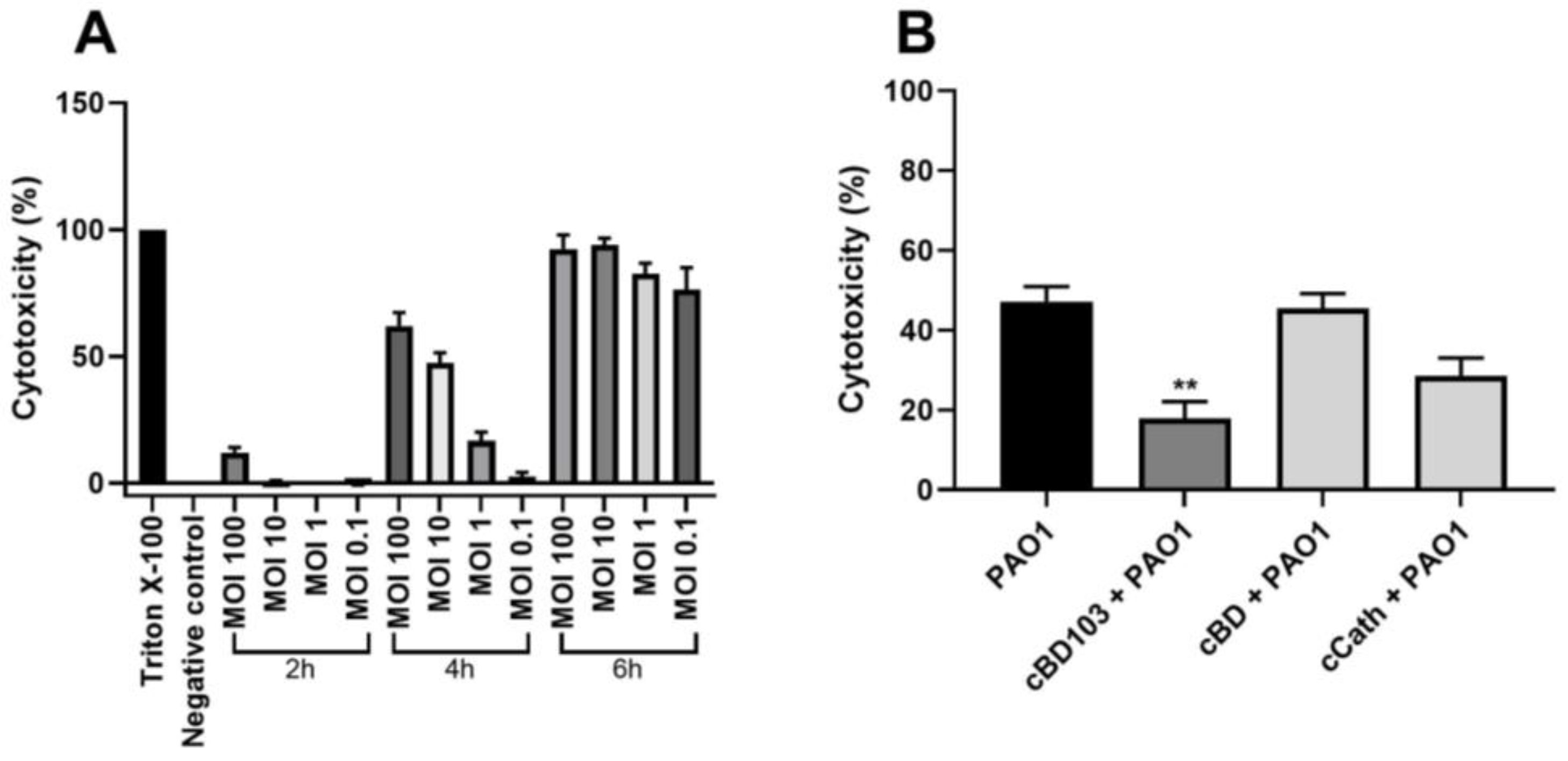
3.3. AMPs Alleviated the Cytotoxicity of Canine Keratinocytes Induced by PAO1
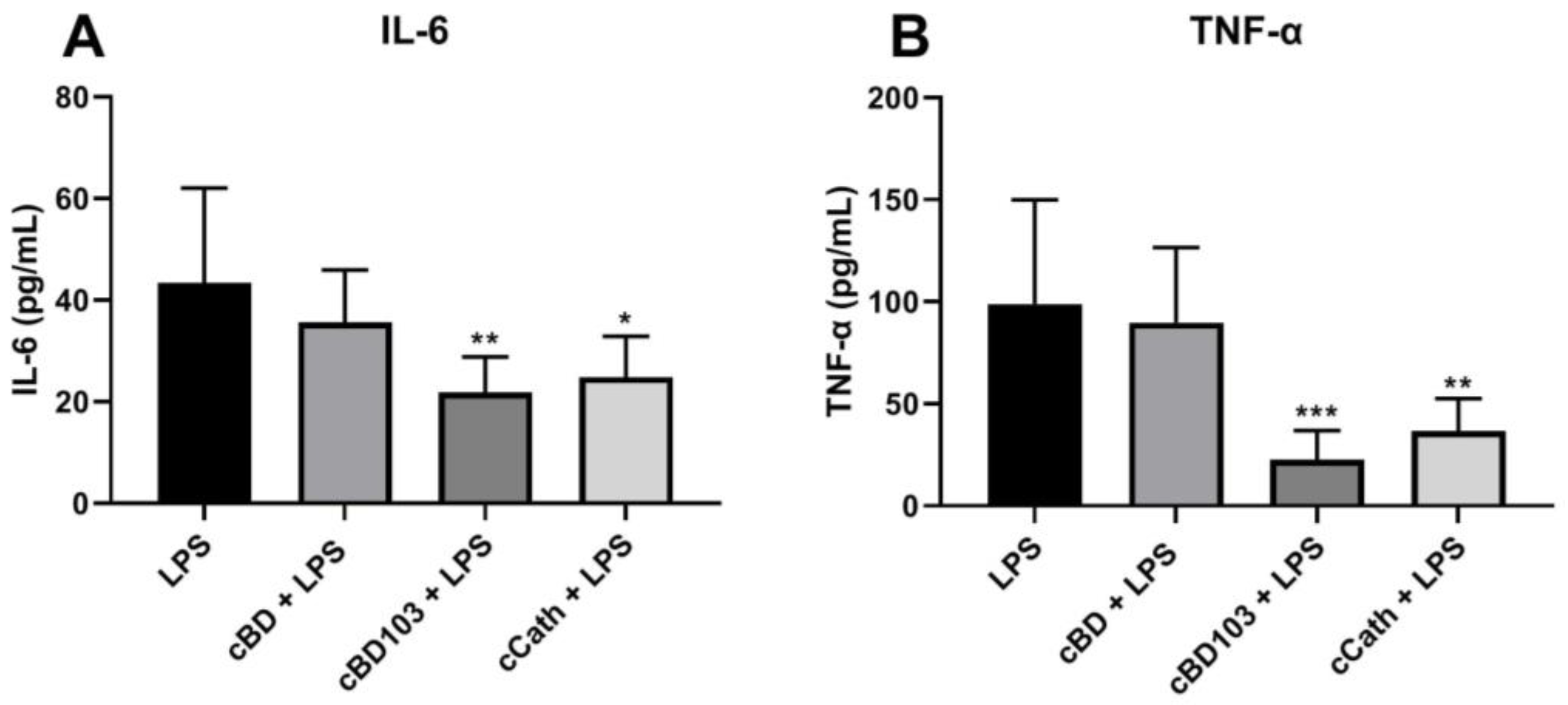
3.4. AMPs Mitigated P. aeruginosa LPS-Induced Inflammation in Canine Keratinocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2008, 19, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Braff, M.H.; Di Nardo, A.; Gallo, R.L. Keratinocytes store the antimicrobial peptide cathelicidin in lamellar bodies. J. Investig. Dermatol. 2005, 124, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Schauber, J.; Gallo, R.L. Antimicrobial peptides and the skin immune defense system. J. Allergy Clin. Immunol. 2008, 122, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.G.; Yan, H.; Hancock, R.E. Biological properties of structurally related α-helical cationic antimicrobial peptides. Infect. Immun. 1999, 67, 2005–2009. [Google Scholar] [CrossRef] [PubMed]
- van Damme, C.M.; Willemse, T.; van Dijk, A.; Haagsman, H.P.; Veldhuizen, E.J. Altered cutaneous expression of beta-defensins in dogs with atopic dermatitis. Mol. Immunol. 2009, 46, 2449–2455. [Google Scholar] [CrossRef]
- Wuerth, K.C.; Falsafi, R.; Hancock, R.E.W. Synthetic host defense peptide IDR-1002 reduces inflammation in Pseudomonas aeruginosa lung infection. PLoS ONE 2017, 12, e0187565. [Google Scholar] [CrossRef] [PubMed]
- Dosunmu, E.F.; Emeh, R.O.; Dixit, S.; Bakeer, M.K.; Coats, M.T.; Owen, D.R.; Pillai, S.R.; Singh, S.R.; Dennis, V.A. The anti-microbial peptide TP359 attenuates inflammation in human lung cells infected with Pseudomonas aeruginosa via TLR5 and MAPK pathways. PLoS ONE 2017, 12, e0176640. [Google Scholar] [CrossRef]
- Santoro, D.; Maddox, C.W. Canine antimicrobial peptides are effective against resistant bacteria and yeasts. Vet. Dermatol. 2014, 25, 35-e12. [Google Scholar] [CrossRef] [PubMed]
- Kerr, K.G.; Snelling, A.M. Pseudomonas aeruginosa: A formidable and ever-present adversary. J. Hosp. Infect. 2009, 73, 338–344. [Google Scholar] [CrossRef]
- Golovkine, G.; Reboud, E.; Huber, P. Pseudomonas aeruginosa takes a multi-target approach to achieve junction breach. Front. Cell. Infect. Microbiol. 2017, 7, 532. [Google Scholar]
- Lau, G.W.; Hassett, D.J.; Britigan, B.E. Modulation of lung epithelial functions by Pseudomonas aeruginosa. Trends. Microbiol. 2005, 13, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa biofilms in cystic fibrosis. Future Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Hillier, A.; Alcorn, J.R.; Cole, L.K.; Kowalski, J.J. Pyoderma caused by Pseudomonas aeruginosa infection in dogs: 20 cases. Vet. Dermatol. 2006, 17, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, T.; Cole, L.K. Evidence-based veterinary dermatology: A systematic review of interventions for treatment of Pseudomonas otitis in dogs. Vet. Dermatol. 2007, 18, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.E.; Chung, T.H.; Hwang, C.Y. Identification of VIM-2 metallo-β-lactamase-producing Pseudomonas aeruginosa isolated from dogs with pyoderma and otitis in Korea. Vet. Dermatol. 2018, 29, 186-e68. [Google Scholar] [CrossRef] [PubMed]
- Cabassi, C.S.; Sala, A.; Santospirito, D.; Alborali, G.L.; Carretto, E.; Ghibaudo, G.; Taddei, S. Activity of AMP2041 against human and animal multidrug resistant Pseudomonas aeruginosa clinical isolates. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 17. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005, 1, 1B.1.1–1B.1.18. [Google Scholar]
- Fazakerley, J.; Crossley, J.; McEwan, N.; Carter, S.; Nuttall, T. In vitro antimicrobial efficacy of β-defensin 3 against Staphylococcus pseudintermedius isolates from healthy and atopic canine skin. Vet. Dermatol. 2010, 21, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Fritz, P.; Beck-Jendroschek, V.; Brasch, J. Inhibition of dermatophytes by the antimicrobial peptides human β-defensin-2, ribonuclease 7 and psoriasin. Med. Mycol. 2012, 50, 579–584. [Google Scholar] [CrossRef]
- Baroni, A.; Buommino, E.; De Gregorio, V.; Ruocco, E.; Ruocco, V.; Wolf, R. Structure and function of the epidermis related to barrier properties. Clin. Dermatol. 2012, 30, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Santoro, D.; Bunick, D.; Graves, T.K.; Campbell, K.L. Expression and distribution of antimicrobial peptides in the skin of healthy beagles. Vet. Dermatol. 2011, 22, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Jindal, H.M.; Le, C.F.; Mohd Yusof, M.Y.; Velayuthan, R.D.; Lee, V.S.; Zain, S.M.; Isa, D.M.; Sekaran, S.D. Antimicrobial activity of novel synthetic peptides derived from indolicidin and ranalexin against Streptococcus pneumoniae. PLoS ONE 2015, 10, e0128532. [Google Scholar] [CrossRef] [PubMed]
- Le, C.F.; Yusof, M.Y.; Hassan, H.; Sekaran, S.D. In vitro properties of designed antimicrobial peptides that exhibit potent antipneumococcal activity and produces synergism in combination with penicillin. Sci. Rep. 2015, 5, 9761. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sahoo, N.; Bhunia, A. Antimicrobial peptides and their pore/ion channel properties in neutralization of pathogenic microbes. Curr. Top. Med. Chem. 2016, 16, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Rad, H.I.; Arzanlou, M.; Omid, M.R.; Ravaji, S.; Doghaheh, H.P. Effect of culture media on chemical stability and antibacterial activity of allicin. J. Funct. Foods 2017, 28, 321–325. [Google Scholar]
- Mah, T.F.; Pitts, B.; Pellock, B.; Walker, G.C.; Stewart, P.S.; O’Toole, G.A. A genetic basis for Pseudomonas aeruginosa biofilm antibiotic resistance. Nature 2003, 426, 306–310. [Google Scholar] [CrossRef]
- Roland, P.S.; Stroman, D.W. Microbiology of acute otitis externa. Laryngoscope 2002, 112, 1166–1177. [Google Scholar] [CrossRef]
- Post, J.C.; Hiller, N.L.; Nistico, L.; Stoodley, P.; Ehrlich, G.D. The role of biofilms in otolaryngologic infections: Update 2007. Curr. Opin. Otolaryngol. Head Neck Surg. 2007, 15, 347–351. [Google Scholar] [CrossRef]
- Nuti, R.; Goud, N.S.; Saraswati, A.P.; Alvala, R.; Alvala, M. Antimicrobial Peptides: A Promising Therapeutic Strategy in Tackling Antimicrobial Resistance. Curr. Med. Chem. 2017, 24, 4303–4314. [Google Scholar] [CrossRef]
- Dunzendorfer, S.; Lee, H.-K.; Soldau, K.; Tobias, P.S. TLR4 is the signaling but not the lipopolysaccharide uptake receptor. J. Immunol. 2004, 173, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.M.; Pereira, M.O. Pseudomonas aeruginosa diversification during infection development in cystic fibrosis lungs—A review. Pathogens 2014, 3, 680–703. [Google Scholar] [CrossRef]
- Malhotra, S.; Limoli, D.H.; English, A.E.; Parsek, M.R.; Wozniak, D.J. Mixed communities of mucoid and nonmucoid Pseudomonas aeruginosa exhibit enhanced resistance to host antimicrobials. mBio 2018, 9, e00275-18. [Google Scholar] [CrossRef] [PubMed]
- Milivojevic, D.; Šumonja, N.; Medić, S.; Pavic, A.; Moric, I.; Vasiljevic, B.; Senerovic, L.; Nikodinovic-Runic, J. Biofilm-forming ability and infection potential of Pseudomonas aeruginosa strains isolated from animals and humans. Pathog. Dis. 2018, 76, fty041. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Name | Sequence | Molecular Weight * | NCBI Reference Sequence |
|---|---|---|---|
| cBD | KCWNLRGSCREKCIKNEKLYIFCTSGKLCCLKPK | 3994.92 | NM_001313788.1 (202–303, 102 bp) |
| cBD103 | GIINTLQRYYCRIRSGRCALLSCLPKEEQIGRCSSTGRKCCRRKK | 5206.23 | NM_001129980.1 (180–314, 135 bp) |
| cCath | RLKELITTGGQKIGEKIRRIGQRIKDFFKNLQPREEKS | 4512.29 | NM_001003359.1 (443–556, 114 bp) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyun, J.-E.; Hwang, C.-Y. Antimicrobial Peptide Reduces Cytotoxicity and Inflammation in Canine Epidermal Keratinocyte Progenitor Cells Induced by Pseudomonas aeruginosa Infection. Vet. Sci. 2024, 11, 235. https://doi.org/10.3390/vetsci11060235
Hyun J-E, Hwang C-Y. Antimicrobial Peptide Reduces Cytotoxicity and Inflammation in Canine Epidermal Keratinocyte Progenitor Cells Induced by Pseudomonas aeruginosa Infection. Veterinary Sciences. 2024; 11(6):235. https://doi.org/10.3390/vetsci11060235
Chicago/Turabian StyleHyun, Jae-Eun, and Cheol-Yong Hwang. 2024. "Antimicrobial Peptide Reduces Cytotoxicity and Inflammation in Canine Epidermal Keratinocyte Progenitor Cells Induced by Pseudomonas aeruginosa Infection" Veterinary Sciences 11, no. 6: 235. https://doi.org/10.3390/vetsci11060235
APA StyleHyun, J.-E., & Hwang, C.-Y. (2024). Antimicrobial Peptide Reduces Cytotoxicity and Inflammation in Canine Epidermal Keratinocyte Progenitor Cells Induced by Pseudomonas aeruginosa Infection. Veterinary Sciences, 11(6), 235. https://doi.org/10.3390/vetsci11060235