
Effects of New Mutations in BMPRIB, GDF9, BMP15, LEPR, and B4GALNT2 Genes on Litter Size in Sheep
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. SNP Genotyping by iPLEX MassARRAY
2.3. Statistical Analyses
3. Results
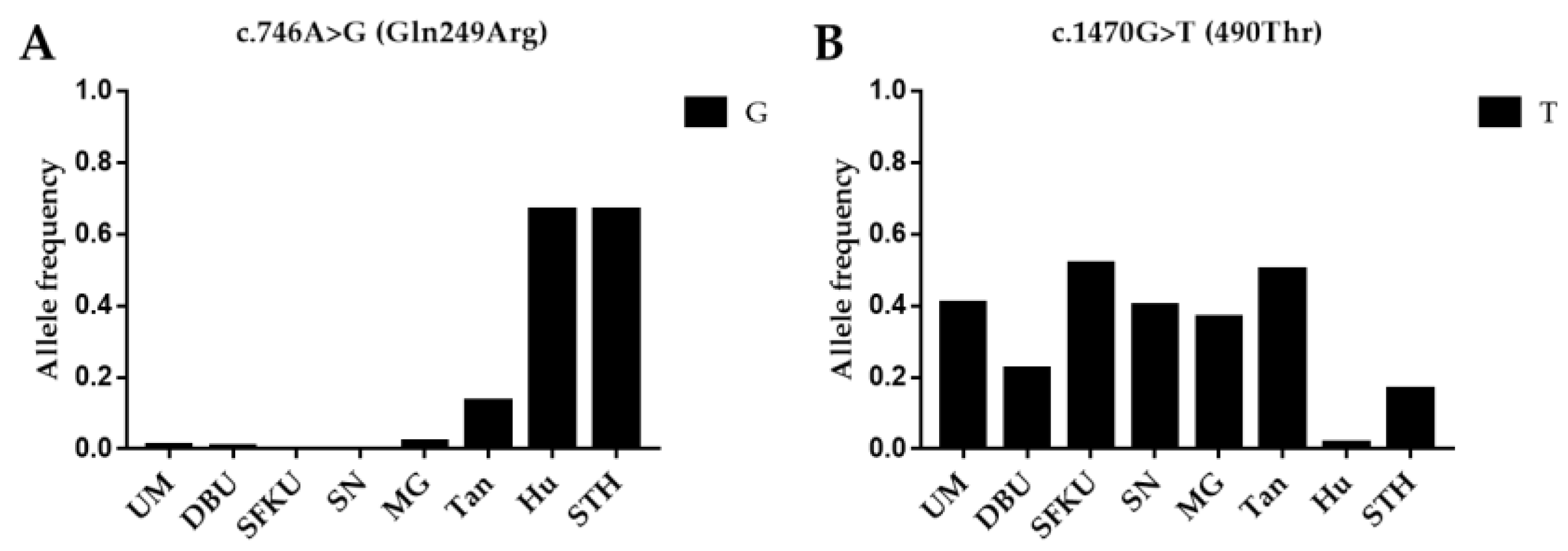
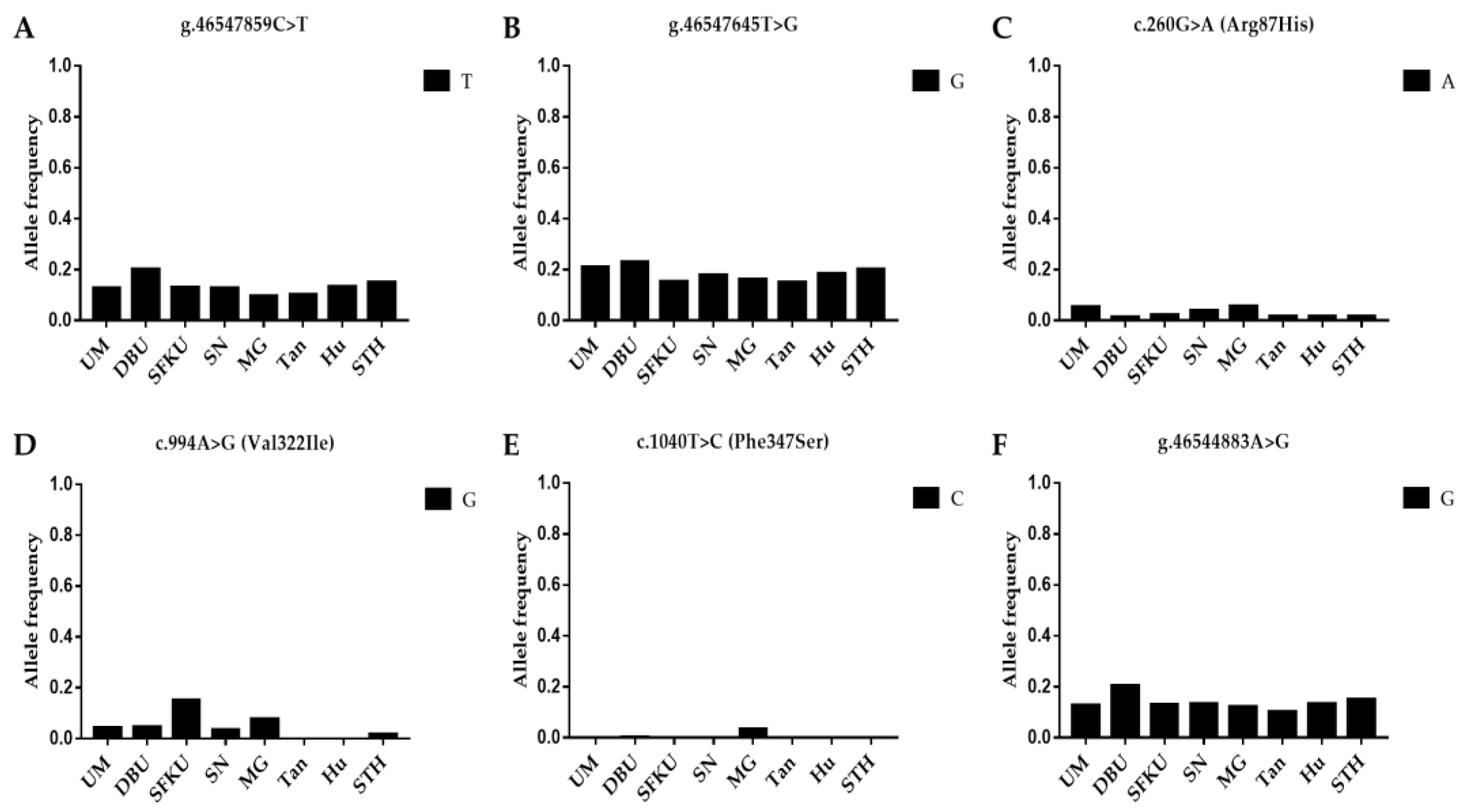
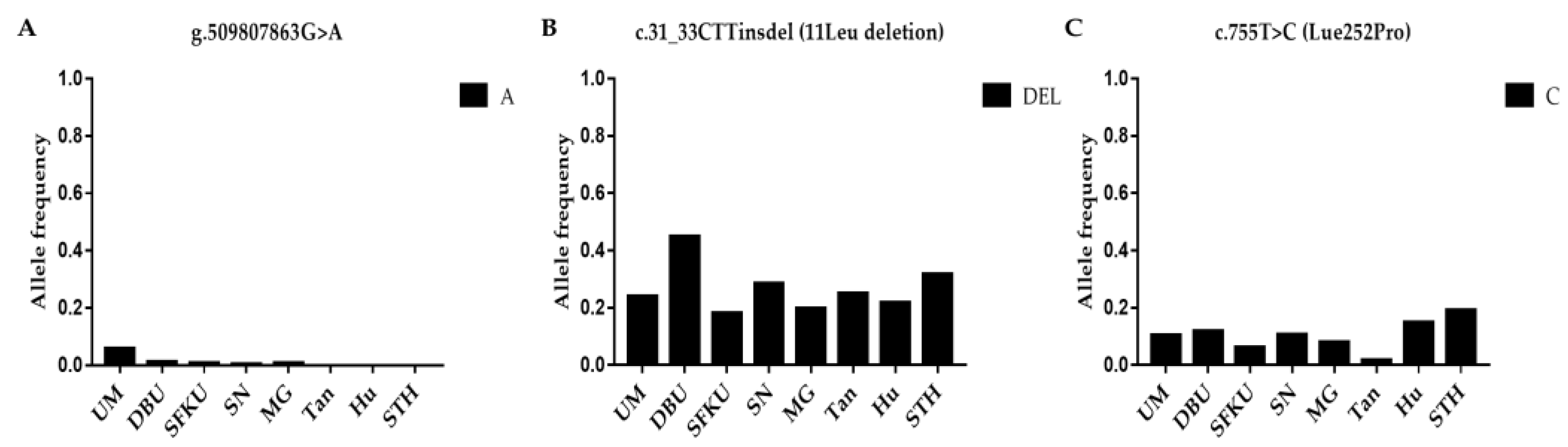
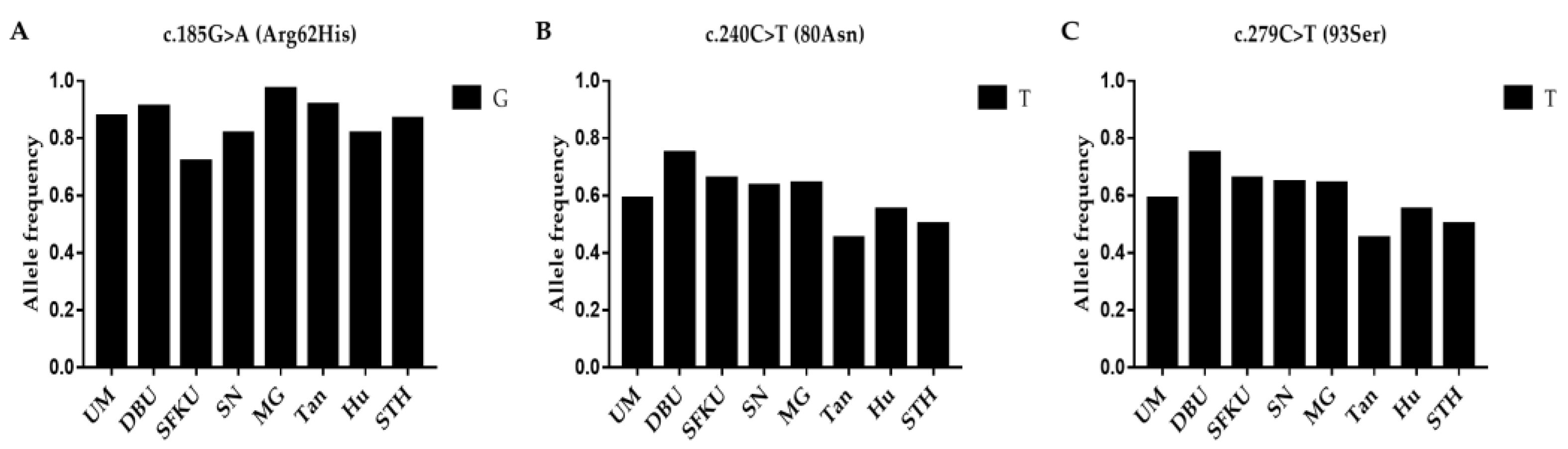
3.1. Genetic Diversity Analysis
3.2. Associations between Mutations in Candidate Genes and Litter Size in Sheep
3.2.1. Associations between Mutations in BMPR1B and Litter Size in Sheep
3.2.2. Associations between Mutations in GDF9 and Litter Size in Sheep
3.2.3. Associations between Mutations in BMP15 and Litter Size in Sheep
3.2.4. Associations between Mutations in LEPR and Litter Size in Sheep
3.2.5. Associations between Mutations in B4GALNT2 and Litter Size in Sheep
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- China National Commission of Animal Genetic Resources (CNCAGR). Sheep and Goats, Animal Genetic Resources in China; China Agriculture Press: Beijing, China, 2011. [Google Scholar]
- Liu, Z.; Ji, Z.; Wang, G.; Chao, T.; Hou, L.; Wang, J. Genome-wide analysis reveals signatures of selection for important traits in domestic sheep from different ecoregions. BMC Genomics 2016, 17, 863. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, W.; Lv, F.; He, S.; Tian, S.; Peng, W.; Sun, Y.; Zhao, Y.; Tu, X.; Zhang, M.; et al. Whole-genome sequencing of native sheep provides insights into rapid adaptations to extreme environments. Mol. Biol. Evol. 2016, 33, 2576–2592. [Google Scholar] [CrossRef] [PubMed]
- Lande, R.; Thompson, R. Efficiency of marker-assisted selection in the improvement of quantitative trait. Genetics 1990, 124, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Tong, B.; Wang, J.; Cheng, Z.; Liu, J.; Wu, Y.; Li, Y.; Bai, C.; Zhao, S.; Yu, H.; Li, G. Novel Variants in GDF9 Gene Affect Promoter Activity and Litter Size in Mongolia Sheep. Genes 2020, 11, 375. [Google Scholar] [CrossRef]
- Cheng, Z.; Lu, D.; Liu, J.; Li, Y.; Zhang, X.; Chao, L.; Cang, M.; Wang, J.; Yu, H.; Li, G.; et al. Effect of g. 46547645T>G Locus of GDF9 Gene on Promoter Activity and Litter Size of Mongolia Sheep (Ovis aries). J. Agric. Biotechnol. 2021, 29, 540–549. [Google Scholar]
- Gao, Y.; Hao, Q.; Cang, M.; Wang, J.; Yu, H.; Liu, Y.; Zhang, W.; Tong, B. Association between novel variants in BMPR1B gene and litter size in Mongolia and Ujimqin sheep breeds. Reprod. Domest. Anim. 2021, 56, 1562–1571. [Google Scholar] [CrossRef]
- Wang, Y.; Chi, Z.; Jia, S.; Zhao, S.; Cao, G.; Purev, C.; Chang, M.; Yu, H.; Li, X.; Bao, S.; et al. Effects of novel variants in BMP15 gene on litter size in Mongolia and Ujimqin sheep breeds. Theriogenology 2023, 198, 1–11. [Google Scholar] [CrossRef]
- Ma, S.; Ji, X.; Cang, M.; Wang, J.; Yu, H.; Liu, Y.; Zhang, W.; Wu, Y.; Zhao, S.; Cao, G.; et al. Association analysis between novel variants in LEPR gene and litter size in Mongolia and Ujimqin sheep breeds. Theriogenology 2022, 183, 79–89. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, J.; Chao, L.; Cang, M.; Yu, H.; Wang, J.; Bao, S.; Liu, Y.; Zhang, W.; Ma, Q.; et al. Association Between Polymorphisms in B4GALNT2 Gene and Litter Size in Mongolia Sheep and Ujimqin Sheep (Ovis aries). J. Agric. Biotechnol. 2022, 30, 1510–1523. [Google Scholar]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mérino ewes. Proc. Natl. Acad. Sci. USA 2001, 98, 5104–5109. [Google Scholar] [CrossRef]
- Souza, C.J.; MacDougall, C.; Campbell, B.K.; McNeilly, A.S.; Baird, D.T. The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1 B (BMPR1B) gene. J. Endocrinol. 2001, 169, R1–R6. [Google Scholar] [CrossRef]
- Wilson, T.; Wu, X.Y.; Juengel, J.L.; Ross, I.K.; Lumsden, J.M.; Lord, E.A.; Dodds, K.G.; Walling, G.A.; McEwan, J.C.; O’Connell, A.R.; et al. Highly prolific Booroola sheep have a mutation in the intracellular kinase domain of bone morphogenetic protein IB receptor (ALK-6) that is expressed in both oocytes and granulosa cells. Biol. Reprod. 2001, 64, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Hanrahan, P.J.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Amini, H.R.; Ajaki, A.; Farahi, M.; Heidari, M.; Pirali, A.; Forouzanfar, M.; Eghbalsaied, S. The novel T755C mutation in BMP15 is associated with the litter size of Iranian Afshari, Ghezel, and Shal breeds. Arch. Anim. Breed. 2018, 61, 153–160. [Google Scholar] [CrossRef]
- Juengel, J.L.; O’Connell, A.R.; French, M.C.; Proctor, L.E.; Wheeler, R.; Farquhar, P.A.; Dodds, K.G.; Galloway, S.M.; Johnstone, P.D.; Davis, G.H. Identification of a line of sheep carrying a putative autosomal gene increasing ovulation rate in sheep that does not appear to interact with mutations in the transforming growth factor beta superfamily. Biol. Reprod. 2011, 85, 113–120. [Google Scholar] [CrossRef]
- Guo, X.; Wang, X.; Liang, B.; Di, R.; Liu, Q.; Hu, W.; He, X.; Zhang, J.; Zhang, X.; Chu, M. Molecular cloning of the B4GALNT2 gene and its single nucleotide polymorphisms association with litter size in Small Tail Han sheep. Animals 2018, 8, 160. [Google Scholar] [CrossRef]
- Gabriel, S.; Ziaugra, L.; Tabbaa, D. SNP genotyping using the Sequenom MassARRAY iPLEX platform. Curr. Protoc. Hum. Genet. 2009, 60, 2.12.1–2.12.18. [Google Scholar] [CrossRef]
- Nei, M. Mathematical model for studying genetic variation in terms of restriction endonuclease. Proc. Natl. Acad. Sci. USA 1979, 89, 1477–1481. [Google Scholar] [CrossRef]
- Davis, G.H.; Galloway, S.M.; Ross, I.K.; Gregan, S.M.; Ward, J.; Nimbkar, B.V.; Ghalsasi, P.M.; Nimbkar, C.; Gray, G.D.; Subandriyo; et al. DNA tests in prolific sheep from eight countries provide new evidence on origin of the Booroola (FecB) mutation. Biol. Reprod. 2002, 66, 1869–1874. [Google Scholar] [CrossRef]
- Chu, M.X.; Liu, Z.H.; Jiao, C.L.; He, Y.Q.; Fang, L.; Ye, S.C.; Chen, G.H.; Wang, J.Y. Mutations in BMPR-IB and BMP-15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J. Anim. Sci. 2007, 85, 598–603. [Google Scholar] [CrossRef]
- Guan, F.; Liu, S.R.; Shi, G.Q.; Yang, L.G. Polymorphism of FecB gene in nine sheep breeds or strains and its effects on litter size, lamb growth and development. Anim. Reprod. Sci. 2007, 99, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, N.M. A review of the effects of the Booroola gene (FecB) on sheep production. Small. Ruminant. Res. 2009, 85, 75–84. [Google Scholar] [CrossRef]
- Mahdavi, M.; Nanekarani, S.; Hosseini, S.D. Mutation in BMPR-IB gene is associated with litter size in Iranian Kalehkoohi sheep. Anim. Reprod. Sci. 2014, 147, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.S.; Geng, L.Y.; Du, L.X.; Liu, Z.Z.; Fu, Z.X.; Feng, M.S.; Gong, F.Y. Polymorphic study of FecX(G), FecG(H) and Fec(B) mutations in four domestic sheep breeds in the Lower Yellow River Valley of China. J. Anim. Vet. Adv. 2011, 10, 2198–2201. [Google Scholar]
- Abdoli, R.; Zamani, P.; Deljou, A.; Rezvan, H. Association of BMPR-1B and GDF9 genes polymorphisms and secondary protein structure changes with reproduction traits in Mehraban ewes. Gene 2013, 524, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Chen, Q.; Gui, L.; Jin, J.; Li, Y.; Ru, Q.; Hou, S. Association of polymorphisms in bone morphogenetic protein receptor-1B gene exon-9 with litter size in Dorset, Mongolian, and Small Tail Han ewes. Asian-Australas. J. Anim. Sci. 2019, 32, 949–955. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, X.; Liu, G.; Deng, M.; Sun, B.; Guo, Y.; Liu, D.; Li, Y. Polymorphisms in BMPR-IB gene and their association with litter size trait in Chinese Hu sheep. Anim. Biotechnol. 2022, 33, 250–259. [Google Scholar] [CrossRef]
- Li, H.; Xu, H.; Akhatayeva, Z.; Liu, H.; Lin, C.; Han, X.; Lu, X.; Lan, X.; Zhang, Q.; Pan, C. Novel indel variations of the sheep FecB gene and their effects on litter size. Gene 2021, 767, 145176. [Google Scholar] [CrossRef]
- Hua, G.H.; Yang, L.G. A review of research progress of FecB gene in Chinese breeds of sheep. Anim. Reprod. Sci. 2009, 116, 1–9. [Google Scholar] [CrossRef]
- Bravo, S.; Larama, G.; Paz, E.; Inostroza, K.; Montaldo, H.H.; Sepúlveda, N. Polymorphism of the GDF9 gene associated with litter size in Araucana creole sheep. Anim. Genet. 2016, 47, 390–391. [Google Scholar] [CrossRef]
- Niu, Z.; Qin, J.; Jiang, Y.; Ding, X.; Ding, Y.; Tang, S.; Shi, H. The Identification of Mutation in BMP15 Gene Associated with Litter Size in Xinjiang Cele Black Sheep. Animals 2021, 11, 668. [Google Scholar] [CrossRef]
- Najafabadi, H.A.; Khansefid, M.; Mahmoud, G.G.; Haruna, I.L.; Zhou, H.; Hickford, J.G. Identification of sequence variation in the oocyte-derived bone morphogenetic protein 15 (BMP15) gene (BMP15) associated with litter size in New Zealand sheep (Ovis aries) breeds. Mol. Biol. Rep. 2021, 48, 6335–6342. [Google Scholar] [CrossRef] [PubMed]
- Di, R.; Wang, F.; Yu, P.; Wang, X.; He, X.; Mwacharo, J.M.; Pan, L.; Chu, M. Detection of Novel Variations Related to Litter Size in BMP15 Gene of Luzhong Mutton Sheep (Ovis aries). Animals 2021, 11, 3528. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, R.; Zamani, P.; Mirhoseini, S.Z.; Hossein-Zadeh, N.G.; Nadri, S. A review on prolificacy genes in sheep. Reprod. Domest. Anim. 2016, 51, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Vinet, A.; Drouilhet, L.; Bodin, L.; Mulsant, P.; Fabre, S.; Phocas, F. Genetic control of multiple births in low ovulating mammalian species. Mamm. Genome 2012, 23, 727–740. [Google Scholar] [CrossRef]
- Drouilhet, L.; Lecerf, F.; Bodin, L.; Fabre, S.; Mulsant, P. Fine mapping of the FecL locus influencing prolificacy in Lacaune sheep. Anim. Genet. 2009, 40, 804–812. [Google Scholar] [CrossRef]
- Ricordeau, G.; Thimonier, J.; Poivey, J.; Driancourt, M.; Hochereau-DeReviers, M.; Tchamitchian, L.I.N.R.A. Research on the Romanov sheep breed in France: A review. Livest. Prod. Sci. 1990, 24, 305–332. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
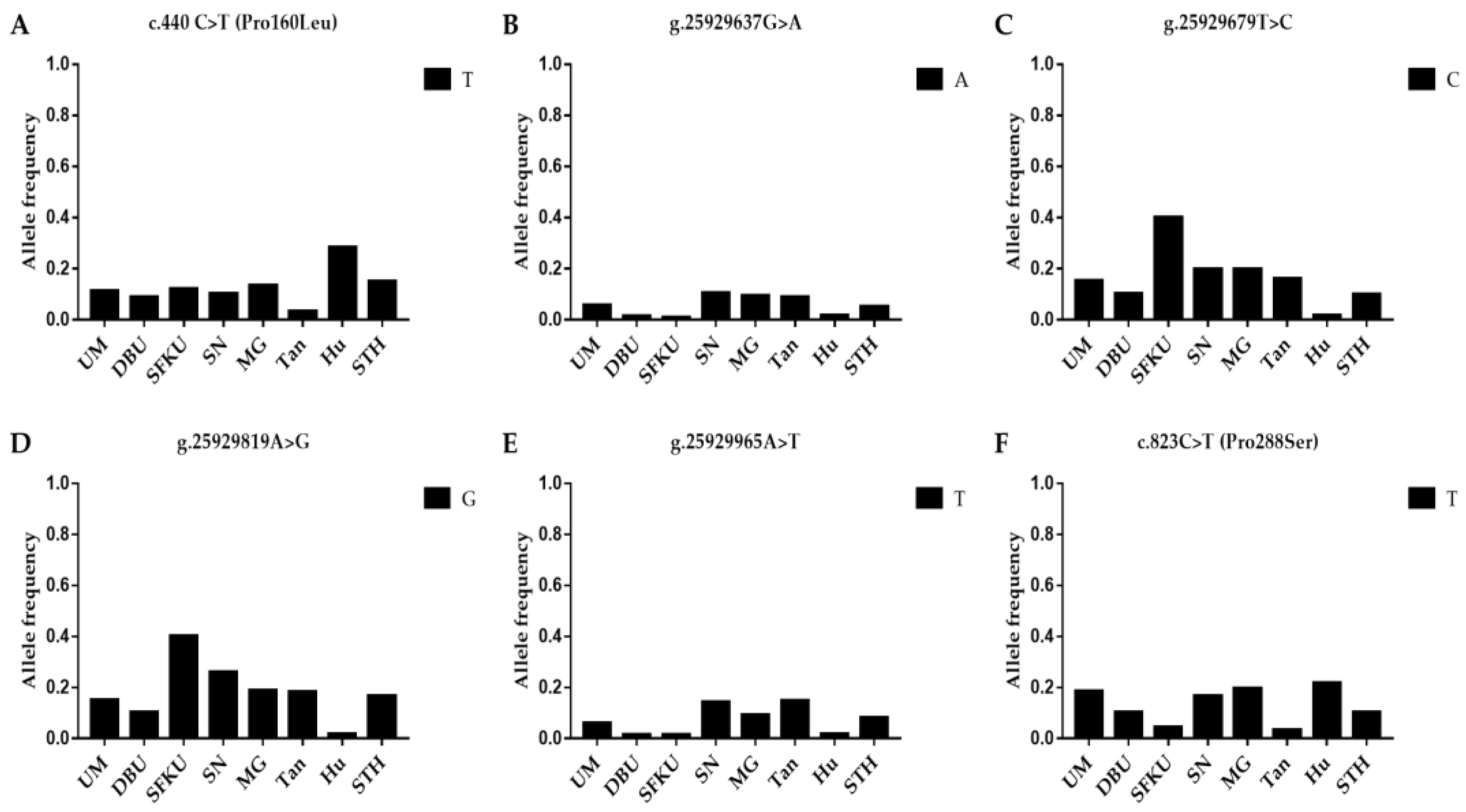{kind=link}
| Breed | Abbreviation | Number of Ewes | Type |
|---|---|---|---|
| Ujimqin | UM | 325 (single lamb 264 + twin lambs 61) | Single birth |
| Dorper × Ujimqin F1 | DPU | 304 (single lamb 180 + twin lambs 124) | Single birth |
| Suffolk × Ujimqin F1 | SFKU | 66 (single lamb 28 + twin lambs 38) | Single birth |
| Mongolia 1 | MG | 250 | Single birth |
| Sonid | SN | 184 | Single birth |
| Tan | Tan | 30 | Single birth |
| Hu | Hu | 30 | Multiple births |
| Small-tailed Han | STH | 30 | Multiple births |
| Sheep Population | Mutation | Genotype | Number | Litter Size |
|---|---|---|---|---|
| Ujimqin | c.746A>G (Gln249Arg) | AA | 320 | 1.18 ± 0.02 a |
| AG | 5 | 1.60 ± 0.24 b | ||
| c.1470G>T (490Thr) | AA | 45 | 1.20 ± 0.05 | |
| AC | 155 | 1.17 ± 0.03 | ||
| CC | 116 | 1.21 ± 0.04 | ||
| Dorper × Ujimqin F1 | c.746A>G (Gln249Arg) | AA | 300 | 1.40 ± 0.03 a |
| AG | 4 | 2.00 ± 0.00 b | ||
| c.1470G>T (490Thr) | AA | 12 | 1.25 ± 0.13 | |
| AC | 111 | 1.41± 0.05 | ||
| CC | 181 | 1.42 ± 0.04 | ||
| Suffolk × Ujimqin F1 | c.1470G>T (490Thr) | AA | 14 | 1.50 ± 0.14 |
| AC | 40 | 1.63 ± 0.08 | ||
| CC | 12 | 1.28 ± 0.06 |
| Breed | Mutation | Genotype | Number | Litter Size |
|---|---|---|---|---|
| Ujimqin | g.46547859C>T | CC | 244 | 1.19 ± 0.03 |
| CT | 79 | 1.18 ± 0.04 | ||
| g.46547645T>G | TT | 198 | 1.20 ± 0.03 | |
| TG | 118 | 1.16 ± 0.03 | ||
| c.260G>A (Arg87His) | GG | 292 | 1.19 ± 0.02 | |
| GA | 32 | 1.16 ± 0.07 | ||
| c.994A>G (Val322Ile) | AA | 299 | 1.19 ± 0.02 | |
| AG | 24 | 1.21 ± 0.08 | ||
| g.46544883A>G | AA | 244 | 1.19 ± 0.03 | |
| AG | 79 | 1.18 ± 0.04 | ||
| Dorper × Ujimqin F1 | g.46547859C>T | CC | 189 | 1.40 ± 0.04 |
| CT | 107 | 1.43 ± 0.05 | ||
| g.46547645T>G | TT | 178 | 1.40 ± 0.04 | |
| GT | 113 | 1.42 ± 0.05 | ||
| c.994A>G (Val322Ile) | AA | 279 | 1.41 ± 0.03 | |
| AG | 28 | 1.43 ± 0.09 | ||
| g.46544883A>G | AA | 189 | 1.40 ± 0.04 | |
| AG | 106 | 1.43 ± 0.05 | ||
| Suffolk × Ujimqin F1 | g.46547859C>T | CC | 51 | 1.59 ± 0.07 |
| CT | 13 | 1.61 ± 0.14 | ||
| g.46547645T>G | TT | 48 | 1.60 ± 0.07 | |
| TG | 16 | 1.56 ± 0.13 | ||
| c.994A>G (Val322Ile) | AA | 46 | 1.48 ± 0.07 a | |
| AG | 20 | 1.80 ± 0.09 b | ||
| g.46544883A>G | AA | 51 | 1.59 ± 0.07 | |
| AG | 13 | 1.62 ± 0.14 |
| Breed | Mutation | Genotype | Number | Litter Size |
|---|---|---|---|---|
| Ujimqin | g.509807863G>A | GG | 287 | 1.19 ± 0.02 |
| GA | 38 | 1.18 ± 0.06 | ||
| c.31_33CTTinsdel (11Leu deletion) | CCT.CCT | 191 | 1.13 ± 0.02 A | |
| CCT.DEL | 112 | 1.27 ± 0.04 B | ||
| DEL.DEL | 22 | 1.27 ± 0.10 | ||
| c.755T>C (Lue252Pro) | TT | 259 | 1.19 ± 0.02 | |
| TC | 64 | 1.20 ± 0.05 | ||
| Dorper × Ujimqin F1 | c.31_33CTTinsdel (11Leu deletion) | CCT.CCT | 84 | 1.38 ± 0.05 |
| CCT.DEL | 167 | 1.41 ± 0.04 | ||
| DEL.DEL | 53 | 1.43 ± 0.07 | ||
| c.755T>C (Lue252Pro) | TT | 235 | 1.41 ± 0.03 | |
| TC | 66 | 1.41 ± 0.06 | ||
| Suffolk × Ujimqin F1 | c.31_33CTTinsdel (11Leu_deletion) | CCT.CCT | 44 | 1.59 ± 0.07 |
| CCT.DEL | 20 | 1.60 ± 0.11 |
| Breed | Mutation | Genotype | Number | Litter Size |
|---|---|---|---|---|
| Ujimqin | c.185G>A (Arg62His) | GG | 249 | 1.21 ± 0.03 |
| GA | 71 | 1.11 ± 0.04 | ||
| c.240C>T (80Asn) | CC | 60 | 1.10 ± 0.04 | |
| CT | 147 | 1.18 ± 0.03 | ||
| TT | 118 | 1.24 ± 0.04 | ||
| c.279C>T (93Ser) | CC | 60 | 1.10 ± 0.04 | |
| CT | 147 | 1.18 ± 0.03 | ||
| TT | 118 | 1.24 ± 0.04 | ||
| Dorper × Ujimqin F1 | c.185G>A (Arg62His) | GG | 249 | 1.41 ± 0.03 |
| GA | 55 | 1.40 ± 0.07 | ||
| c.240C>T (80Asn) | CC | 16 | 1.38 ± 0.13 | |
| CT | 121 | 1.43 ± 0.06 | ||
| TT | 167 | 1.40 ± 0.04 | ||
| c.279C>T (93Ser) | CC | 16 | 1.38 ± 0.13 | |
| CT | 121 | 1.43 ± 0.06 | ||
| TT | 167 | 1.40 ± 0.04 | ||
| Suffolk × Ujimqin F1 | c.185G>A (Arg62His) | GG | 35 | 1.51 ± 0.09 |
| GA | 25 | 1.56 ± 0.10 | ||
| c.240C>T (80Asn) | CT | 35 | 1.66 ± 0.08 | |
| TT | 26 | 1.46 ± 0.10 | ||
| c.279C>T (93Ser) | CT | 35 | 1.66 ± 0.08 | |
| TT | 26 | 1.46 ± 0.10 |
| Breed | Mutation | Genotype | Number | Litter Size |
|---|---|---|---|---|
| Ujimqin | c.440C>T (Pro160Leu) | CC | 255 | 1.18 ± 0.02 |
| CT | 66 | 1.20 ± 0.05 | ||
| g.25929637G>A | GG | 289 | 1.19 ± 0.02 | |
| GA | 35 | 1.17 ± 0.06 | ||
| g.25929679T>C | TT | 229 | 1.18 ± 0.03 | |
| TC | 93 | 1.18 ± 0.04 | ||
| g.25929819A>G | AA | 229 | 1.18 ± 0.03 | |
| AG | 93 | 1.18 ± 0.04 | ||
| g.25929965A>T | AA | 289 | 1.19 ± 0.02 | |
| AT | 33 | 1.12 ± 0.06 | ||
| c.823C>T (Pro288Ser) | CC | 218 | 1.22 ± 0.03 | |
| CT | 93 | 1.14 ± 0.04 | ||
| TT | 14 | 1.07 ± 0.07 | ||
| Dorper × Ujimqin F1 | c.440C>T (Pro160Leu) | CC | 251 | 1.43 ± 0.03 |
| CT | 52 | 1.33 ± 0.07 | ||
| g.25929679T>C | TT | 243 | 1.39 ± 0.03 | |
| TC | 59 | 1.44 ± 0.07 | ||
| g.25929819A>G | AA | 243 | 1.40 ± 0.03 | |
| AG | 59 | 1.44 ± 0.07 | ||
| c.823C>T (Pro288Ser) | CC | 244 | 1.39 ± 0.03 | |
| CT | 57 | 1.49 ± 0.07 | ||
| Suffolk × Ujimqin F1 | c.440C>T (Pro160Leu) | CC | 51 | 1.59 ± 0.07 |
| CT | 14 | 1.57 ± 0.14 | ||
| g.25929679T>C | TT | 20 | 1.40 ± 0.11 | |
| TC | 39 | 1.64 ± 0.08 | ||
| g.25929819A>G | AA | 20 | 1.40 ± 0.11 | |
| AG | 39 | 1.64 ± 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, X.; Cao, Z.; Hao, Q.; He, M.; Cang, M.; Yu, H.; Ma, Q.; Li, X.; Bao, S.; Wang, J.; et al. Effects of New Mutations in BMPRIB, GDF9, BMP15, LEPR, and B4GALNT2 Genes on Litter Size in Sheep. Vet. Sci. 2023, 10, 258. https://doi.org/10.3390/vetsci10040258
Ji X, Cao Z, Hao Q, He M, Cang M, Yu H, Ma Q, Li X, Bao S, Wang J, et al. Effects of New Mutations in BMPRIB, GDF9, BMP15, LEPR, and B4GALNT2 Genes on Litter Size in Sheep. Veterinary Sciences. 2023; 10(4):258. https://doi.org/10.3390/vetsci10040258
Chicago/Turabian StyleJi, Xuewen, Ziwei Cao, Qi Hao, Mei He, Ming Cang, Haiquan Yu, Qing Ma, Xihe Li, Siqin Bao, Jianguo Wang, and et al. 2023. "Effects of New Mutations in BMPRIB, GDF9, BMP15, LEPR, and B4GALNT2 Genes on Litter Size in Sheep" Veterinary Sciences 10, no. 4: 258. https://doi.org/10.3390/vetsci10040258
APA StyleJi, X., Cao, Z., Hao, Q., He, M., Cang, M., Yu, H., Ma, Q., Li, X., Bao, S., Wang, J., & Tong, B. (2023). Effects of New Mutations in BMPRIB, GDF9, BMP15, LEPR, and B4GALNT2 Genes on Litter Size in Sheep. Veterinary Sciences, 10(4), 258. https://doi.org/10.3390/vetsci10040258