
Environmentally Relevant Mixture of Pesticides Affect Mobility and DNA Integrity of Early Life Stages of Rainbow Trout (Oncorhynchus mykiss)


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Chemicals
2.2. Exposure System
2.3. Chemical Analysis of Pollutants in Water
2.4. Embryo-Toxicity Assay
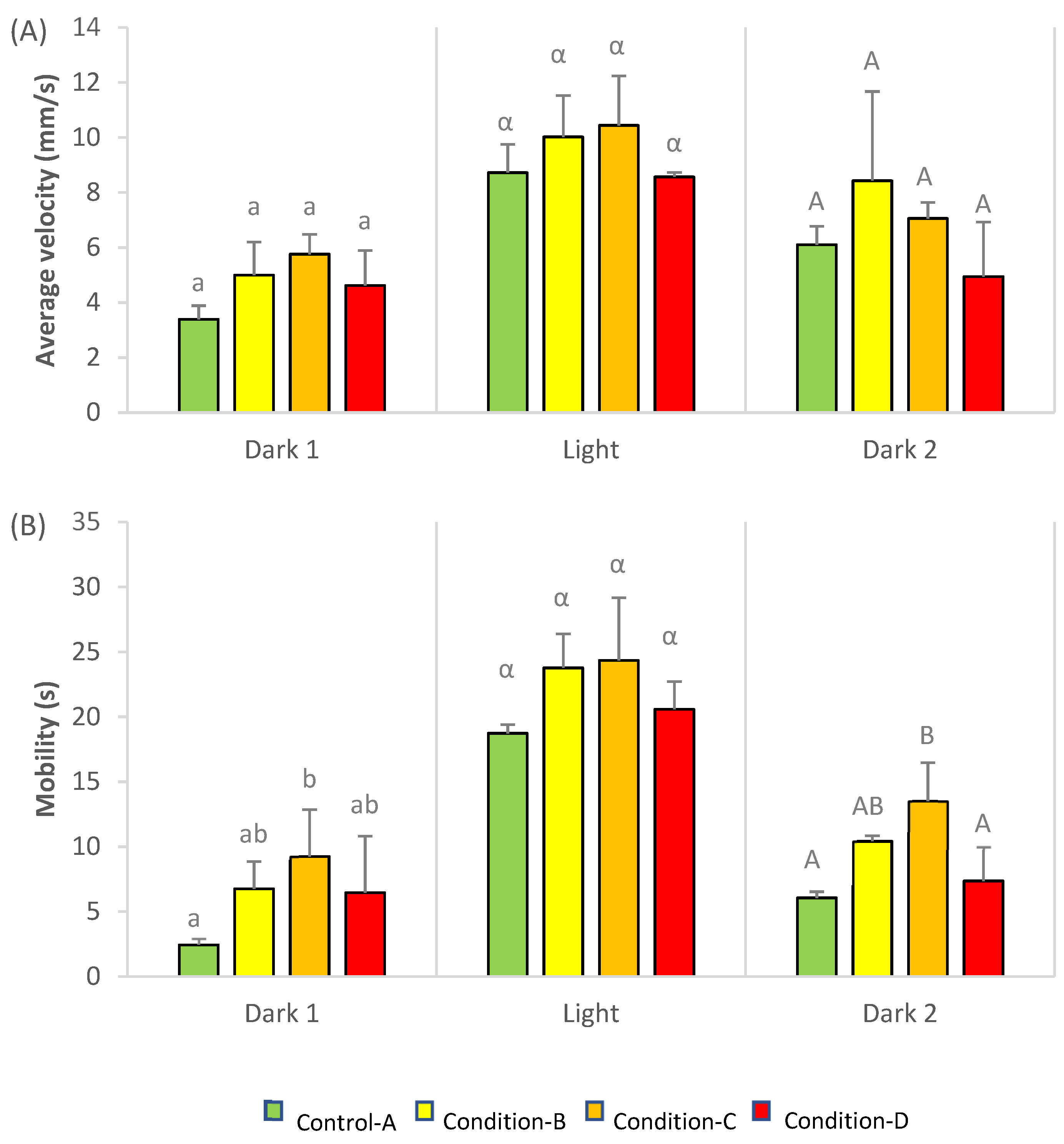
2.5. Swimming Behavior Analyses
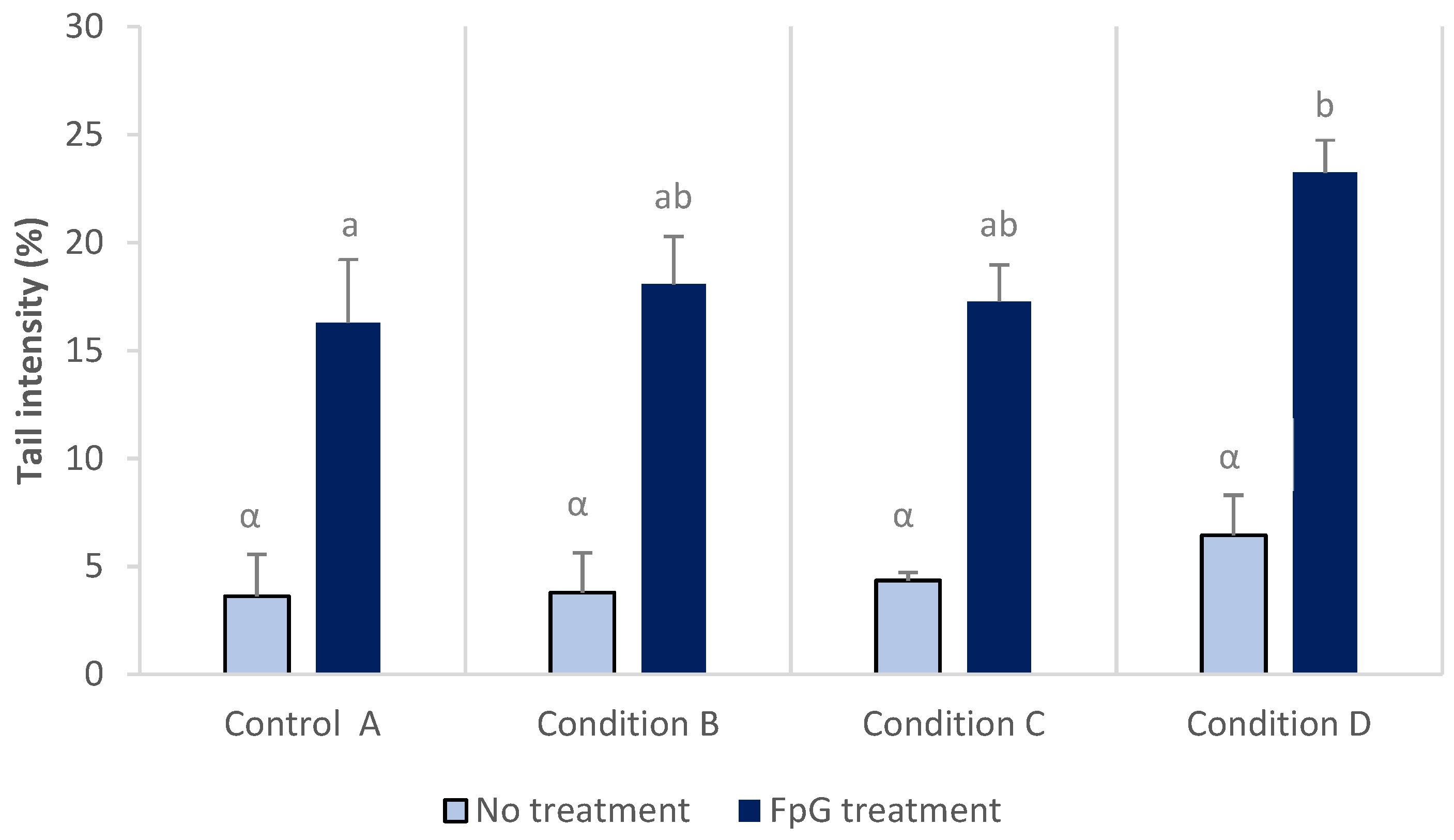
2.6. Comet Assay with FPG Enzyme
2.7. Biochemical Analysis
2.7.1. Preparation of Supernatant
2.7.2. Total Protein
2.7.3. Lipid Peroxidation (TBARS)
2.7.4. Protein Carbonyl Assay
2.8. Gene Expression
2.8.1. RNA Extraction
2.8.2. Retro-Transcription of Total RNA into cDNA
2.8.3. Quantitative Real-Time PCR
2.9. Statistics
3. Results
3.1. Condition of Exposure
3.2. Embryonic and Larval Survival
3.3. Phenotypic Effects
3.4. Swimming Behavior
3.5. DNA Damage
3.6. Lipid Peroxidation and Protein Carbonyls
3.7. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez-Bayo, F. Chapter 4: Impacts of agricultural pesticides on terrestrial ecosystems. Ecolofical Impacts Toxic Chem. 2011, 63–87. [Google Scholar] [CrossRef]
- Stone, W.W.; Gilliom, R.J.; Ryberg, K.R. Pesticides in U.S. streams and rivers: Occurrence and trends during 1992–2011. Environ. Sci. Technol. 2014, 48, 11025–11030. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, F.; Ou, J. Global pesticide consumption and pollution: With China as a focus. Proc. Int. Acad. Ecol. Environ. Sci. 2011, 1, 125–144. [Google Scholar] [CrossRef]
- Ippolito, A.; Kattwinkel, M.; Rasmussen, J.J.; Schäfer, R.B.; Fornaroli, R.; Liess, M. Modeling global distribution of agricultural insecticides in surface waters. Environ. Pollut. 2015, 198, 54–60. [Google Scholar] [CrossRef]
- Krishnan, K.; Brodeur, J. Toxic interactions among environmental pollutants: Corroborating laboratory observations with human experience. Environ. Health Perspect 1994, 102, 11–17. [Google Scholar] [CrossRef]
- Relyea, R.A. A cocktail of contaminants: How mixtures of pesticides at low concentrations affect aquatic communities. Oecologia 2009, 159, 363–376. [Google Scholar] [CrossRef]
- Relyea, R.; Hoverman, J. Assessing the ecology in ecotoxicology: A review and synthesis in freshwater systems. Ecol. Lett. 2006, 9, 1157–1171. [Google Scholar] [CrossRef]
- Schäfer, R.B.; Von Der Ohe, P.C.; Rasmussen, J.; Ke, B.J.; Beketov, M.A.; Schulz, R.; Liess, M. Thresholds for the effects of Pesticides on Invertebrate Communities and Leaf Breakdown in Stream Ecosystems. Environ. Sci. Technol. 2012, 46, 5134–5142. [Google Scholar] [CrossRef]
- Beyer, J.; Petersen, K.; Song, Y.; Ruus, A.; Grung, M.; Bakke, T.; Tollefsen, K.E. Environmental risk assessment of combined effects in aquatic ecotoxicology: A discussion paper. Mar. Environ. Res. 2014, 96, 81–91. [Google Scholar] [CrossRef]
- WFD Water Framework Directive. Available online: http://ec.europa.eu/environment/water/water-framework/index_en.html (accessed on 23 October 2000).
- Devez, A.; Gomez, E.; Gilbin, R.; Elbaz-Poulichet, F.; Persin, F.; Andrieux, P.; Casellas, C. Assessment of copper bioavailability and toxicity in vineyard runoff waters by DPASV and algal bioassay. Sci. Total Environ. 2005, 348, 82–92. [Google Scholar] [CrossRef]
- Ruyters, S.; Salaets, P.; Oorts, K.; Smolders, E. Copper toxicity in soils under established vineyards in Europe: A survey. Sci. Total Environ. 2013, 443, 470–477. [Google Scholar] [CrossRef]
- Manzl, C.; Enrich, J.; Ebner, H.; Dallinger, R.; Krumschnabel, G. Copper-induced formation of reactive oxygen species causes cell death and disruption of calcium homeostasis in trout hepatocytes. Toxicology 2004, 196, 57–64. [Google Scholar] [CrossRef]
- Prousek, J. Fenton chemistry in biology and medicine. Pure Appl. Chem. 2007, 79, 2325–2338. [Google Scholar] [CrossRef]
- Weeks Santos, S.; Cachot, J.; Gourves, P.Y.; Clérandeau, C.; Morin, B.; Gonzalez, P. Sub-lethal effects of waterborne copper in early developmental stages of rainbow trout (Oncorhynchus mykiss). Ecotoxicol. Environ. Saf. 2019, 170, 778–788. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616–617, 255–268. [Google Scholar] [CrossRef]
- Aparicio, V.C.; De Gerónimo, E.; Marino, D.; Primost, J.; Carriquiriborde, P.; Costa, J.L. Environmental fate of glyphosate and aminomethylphosphonic acid in surface waters and soil of agricultural basins. Chemosphere 2013, 93, 1866–1873. [Google Scholar] [CrossRef]
- Peruzzo, P.J.; Porta, A.A.; Ronco, A.E. Levels of glyphosate in surface waters, sediments and soils associated with direct sowing soybean cultivation in north pampasic region of Argentina. Environ. Pollut. 2008, 156, 61–66. [Google Scholar] [CrossRef]
- Coupe, R.H.; Kalkhoff, S.J.; Capel, P.D.; Gregoire, C. Fate and transport of glyphosate and aminomethylphosphonic acid in surface waters of agricultural basins. Pest. Manag. Sci. 2012, 68, 16–30. [Google Scholar] [CrossRef]
- Çavaş, T.; Könen, S. Detection of cytogenetic and DNA damage in peripheral erythrocytes of goldfish (Carassius auratus) exposed to a glyphosate formulation using the micronucleus test and the comet assay. Mutagenesis 2007, 22, 263–268. [Google Scholar] [CrossRef]
- Guilherme, S.; Gaivão, I.; Santos, M.A.; Pacheco, M. European eel (Anguilla anguilla) genotoxic and pro-oxidant responses following short-term exposure to Roundup®—A glyphosate-based herbicide. Mutagenesis 2010, 25, 523–530. [Google Scholar] [CrossRef]
- Guilherme, S.; Santos, M.A.; Barroso, C.; Gaivão, I.; Pacheco, M. Differential genotoxicity of Roundup® formulation and its constituents in blood cells of fish (Anguilla anguilla): Considerations on chemical interactions and DNA damaging mechanisms. Ecotoxicology 2012, 21, 1381–1390. [Google Scholar] [CrossRef]
- Salbego, J.; Pretto, A.; Gioda, C.R.; De Menezes, C.C.; Lazzari, R.; Radünz Neto, J.; Baldisserotto, B.; Loro, V.L. Herbicide formulation with glyphosate affects growth, acetylcholinesterase activity, and metabolic and hematological parameters in Piava (Leporinus obtusidens). Arch. Environ. Contam. Toxicol. 2010, 58, 740–745. [Google Scholar] [CrossRef]
- Sinhorin, V.D.G.; Sinhorin, A.P.; Teixeira, J.M.S.; Miléski, K.M.L.; Hansen, P.C.; Moeller, P.R.; Moreira, P.S.A.; Baviera, A.M.; Loro, V.L. Metabolic and Behavior Changes in Surubim Acutely Exposed to a Glyphosate-Based Herbicide. Arch. Environ. Contam Toxicol. 2014, 659–667. [Google Scholar] [CrossRef]
- Bridi, D.; Altenhofen, S.; Gonzalez, J.B.; Reolon, G.K.; Bonan, C.D. Glyphosate and Roundup®alter morphology and behavior in zebrafish. Toxicology 2017, 392, 32–39. [Google Scholar] [CrossRef]
- de Moura, F.R.; Brentegani, K.R.; Gemelli, A.; Sinhorin, A.P.; Sinhorin, V.D.G. Oxidative stress in the hybrid fish jundiara (Leiarius marmoratus × Pseudoplatystoma reticulatum) exposed to Roundup Original®. Chemosphere 2017, 185, 445–451. [Google Scholar] [CrossRef]
- Glusczak, L.; Loro, V.L.; Pretto, A.; Moraes, B.S.; Raabe, A.; Duarte, M.F.; Da Fonseca, M.B.; De Menezes, C.C.; De Sousa Valladão, D.M. Acute exposure to glyphosate herbicide affects oxidative parameters in piava (Leporinus obtusidens). Arch. Environ. Contam. Toxicol. 2011, 61, 624–630. [Google Scholar] [CrossRef]
- Weeks Santos, S.; Gonzalez, P.; Cormier, B.; Mazzella, N.; Bonnaud, B.; Morin, S.; Clérandeau, C.; Morin, B.; Cachot, J. A glyphosate-based herbicide induces sub-lethal effects in early life stages and liver cell line of rainbow trout, Oncorhynchus mykiss. Aquat. Toxicol. 2019, 216, 105291. [Google Scholar] [CrossRef]
- Marino, D.; Ronco, A. Cypermethrin and Chlorpyrifos Concentration Levels in Surface Water Bodies of the Pampa Ondulada, Argentina. Bull. Environ. Contam. Toxicol. 2005, 75, 820–826. [Google Scholar] [CrossRef]
- Jergentz, S.; Mugni, H.; Bonetto, C.; Schulz, R. Assessment of insecticide contamination in runoff and stream water of small agricultural streams in the main soybean area of Argentina. Chemosphere 2005, 61, 817–826. [Google Scholar] [CrossRef]
- Barron, M.G.; Plakas, S.M.; Wilga, P.C.; Ball, T. Absorption, tissue distribution and metabolism of chlorpyrifos in channel catfish following waterborne exposure. Environ. Toxicol. Chem. 1993, 12, 1469–1476. [Google Scholar] [CrossRef]
- Fulton, M.H.; Key, P.B. Acetylcholinesterase inhibition in estuarine fish and invertebrates as an indicator of organophosphorus insecticide exposure and effects. Environ. Toxicol. Chem. 2001, 20, 37–45. [Google Scholar] [CrossRef]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Weeks Santos, S.; Gonzalez, P.; Cormier, B.; Mazzella, N.; Moreira, A.; Clérandeau, C.; Morin, B.; Cachot, J. Sub-lethal exposure to environmental concentrations of chlorpyrifos affect swimming activity of rainbow trout larvae. Environ. Toxicol. Chem. Rev. 2021. In Press. [Google Scholar]
- Fauvelle, V.; Nhu-Trang, T.T.; Feret, T.; Madarassou, K.; Randon, J.; Mazzella, N. Evaluation of Titanium Dioxide as a Binding Phase for the Passive Sampling of Glyphosate and Aminomethyl Phosphonic Acid in an Aquatic Environment. Anal. Chem. 2015, 87, 6004–6009. [Google Scholar] [CrossRef]
- Le Bihanic, F.; Morin, B.; Cousin, X.; Le Menach, K.; Budzinski, H.; Cachot, J. Developmental toxicity of PAH mixtures in fish early life stages. Part I: Adverse effects in rainbow trout. Environ. Sci. Pollut. Res. 2014, 21, 13720–13731. [Google Scholar] [CrossRef]
- Kienzler, A.; Tronchère, X.; Devaux, A.; Bony, S. Assessment of RTG-W1, RTL-W1, and PLHC-1 fish cell lines for genotoxicity testing of environmental pollutants by means of a Fpg-modified comet assay. Toxicol. Vitr. 2012, 26, 500–510. [Google Scholar] [CrossRef]
- Lowry, O.; Rosebrough, N.; Farr, L.; Randall, R. Protein measurement with the folin phenol reagent. Readings 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal Lipid Peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Augustyniak, E.; Adam, A.; Wojdyla, K.; Rogowska-Wrzesinska, A.; Willetts, R.; Korkmaz, A.; Atalay, M.; Weber, D.; Grune, T.; Borsa, C.; et al. Validation of protein carbonyl measurement: A multi-centre study. Redox Biol. 2015, 4, 149–157. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Dorsch, D.E.; Bay, S.M.; Mays, M.A.; Greenstein, D.J.; Young, D.; Wolf, J.; Pham, D.; Loguinov, A.V.; Vulpe, C. Using Gene Expression to Assess the Status of Fish from Anthropogenically Influenced Estuarine Wetlands. Environ. Sci. Technol. 2011, 46, 69–77. [Google Scholar] [CrossRef]
- Davis, A.P.; Shokouhian, M.; Ni, S. Loading estimates of lead, copper, cadmium, and zinc in urban runoff from specific sources. Chemosphere 2001, 44, 997–1009. [Google Scholar] [CrossRef]
- Phillips, P.J.; Bode, R.W. Pesticides in surface water runoff in south-eastern New York State, USA: Seasonal and stormflow effects on concentrations. Pest. Manag. Sci. 2004, 60, 531–543. [Google Scholar] [CrossRef]
- Subramaniam, V.; Hoggard, P.E. Metal Complexes of Glyphosate. J. Agric. Food Chem. 1988, 36, 1326–1329. [Google Scholar] [CrossRef]
- Glass, R.L. Metal Complex Formation by Glyphosate. J. Agric. Food Chem. 1984, 32, 1249–1253. [Google Scholar] [CrossRef]
- Zhou, C.F.; Wang, Y.J.; Yu, Y.C.; Sun, R.J.; Zhu, X.D.; Zhang, H.L.; Zhou, D.M. Does glyphosate impact on Cu uptake by, and toxicity to, the earthworm Eisenia fetida. Ecotoxicology 2012, 21, 2297–2305. [Google Scholar] [CrossRef]
- Tsui, M.T.K.; Wang, W.X.; Chu, L.M. Influence of glyphosate and its formulation (Roundup®) on the toxicity and bioavailability of metals to Ceriodaphnia dubia. Environ. Pollut. 2005, 138, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Ayoola, S.O. Toxicity of glyphosate herbicide on Nile tilapia (Oreochromis niloticus) juvenile. Afr. J. Agric. Res. 2008, 3, 825–834. [Google Scholar]
- Zhang, S.; Xu, J.; Kuang, X.; Li, S.; Li, X.; Chen, D.; Zhao, X.; Feng, X. Biological impacts of glyphosate on morphology, embryo biomechanics and larval behavior in zebrafish (Danio rerio). Chemosphere 2017, 181, 270–280. [Google Scholar] [CrossRef]
- Kienle, C.; Köhler, H.R.; Gerhardt, A. Behavioural and developmental toxicity of chlorpyrifos and nickel chloride to zebrafish (Danio rerio) embryos and larvae. Ecotoxicol. Environ. Saf. 2009, 72, 1740–1747. [Google Scholar] [CrossRef]
- Richendrfer, H.; Pelkowski, S.D.; Colwill, R.M.; Créton, R. Developmental sub-chronic exposure to chlorpyrifos reduces anxiety-related behavior in zebrafish larvae. Neurotoxicol. Teratol. 2012, 34, 458–465. [Google Scholar] [CrossRef]
- Bonansea, R.I.; Wunderlin, D.A.; Amé, M.V. Behavioral swimming effects and acetylcholinesterase activity changes in Jenynsia multidentata exposed to chlorpyrifos and cypermethrin individually and in mixtures. Ecotoxicol. Environ. Saf. 2016, 129, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Sandahl, J.F.; Baldwin, D.H.; Jenkins, J.J.; Scholz, N.L. Comparative thresholds for acetylcholinesterase inhibition and behavioral impairment in coho salmon exposed to chlorpyrifos. Environ. Toxicol. Chem. 2005, 24, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.; Donerly, S.; Levin, E.D.; Linney, E.A. Differential acetylcholinesterase inhibition of chlorpyrifos, diazinon and parathion in larval zebrafish. Neurotoxicol. Teratol. 2011, 33, 735–741. [Google Scholar] [CrossRef]
- Bonifacio, A.F.; Cazenave, J.; Bacchetta, C.; Ballesteros, M.L.; Bistoni, M.D.L.A.; Amé, M.V.; Bertrand, L.; Hued, A.C. Alterations in the general condition, biochemical parameters and locomotor activity in Cnesterodon decemmaculatus exposed to commercial formulations of chlorpyrifos, glyphosate and their mixtures. Ecol. Indic. 2016, 67, 88–97. [Google Scholar] [CrossRef]
- Tilton, F.A.; Tilton, S.C.; Bammler, T.K.; Beyer, R.P.; Stapleton, P.L.; Scholz, N.L.; Gallagher, E.P. Transcriptional impact of organophosphate and metal mixtures on olfaction: Copper dominates the chlorpyrifos-induced response in adult zebrafish. Aquat. Toxicol. 2011, 102, 205–215. [Google Scholar] [CrossRef]
- Faria, M.; Garcia-Reyero, N.; Padrós, F.; Babin, P.J.; Sebastián, D.; Cachot, J.; Prats, E.; Arick, M.; Rial, E.; Knoll-Gellida, A.; et al. Zebrafish Models for Human Acute Organophosphorus Poisoning. Sci. Rep. 2015, 5, 15591. [Google Scholar] [CrossRef]
- Kavitha, P.; Venkateswara Rao, J. Toxic effects of chlorpyrifos on antioxidant enzymes and target enzyme acetylcholinesterase interaction in mosquito fish, Gambusia affinis. Environ. Toxicol. Pharmacol. 2008, 26, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Fuentes, G.; Rubio-Escalante, F.J.; Noreña-Barroso, E.; Escalante-Herrera, K.S.; Schlenk, D. Impacts of oxidative stress on acetylcholinesterase transcription, and activity in embryos of zebra fish (Danio rerio) following Chlorpyrifos exposure. Comp. Biochem. Physiol. 2015, 172–173, 19–25. [Google Scholar] [CrossRef]
- Glusczak, L.; Dos Santos Miron, D.; Moraes Silveira, B.; Rodrigues Simões, R.; Chitolina Schetinger, M.R.; Morsch, V.M.; Loro, V.L. Acute effects of glyphosate herbicide on metabolic and enzymatic parameters of silver catfish (Rhamdia quelen). Comp. Biochem. Physiol. Toxicol. Pharmacol. 2007, 146, 519–524. [Google Scholar] [CrossRef]
- Topal, A.; Atamanalp, M.; Uçar, A.; Oruç, E.; Kocaman, E.M.; Sulukan, E.; Akdemir, F.; Beydemir, Ş.; Kilinç, N.; Erdoğan, O. Effects of glyphosate on juvenile rainbow trout (Oncorhynchus mykiss): Transcriptional and enzymatic analyses of antioxidant defence system, histopathological liver damage and swimming performance. Ecotoxicol. Environ. Saf. 2015, 111, 206–214. [Google Scholar] [CrossRef]
- Guecheva, T.; Henriques, J.A.P.; Erdtmann, B. Genotoxic effects of copper sulphate in freshwater planarian in vivo, studied with the single-cell gel test (comet assay). Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2001, 497, 19–27. [Google Scholar] [CrossRef]
- Ali, D.; Nagpure, N.S.; Kumar, S.; Kumar, R.; Kushwaha, B.; Lakra, W.S. Assessment of genotoxic and mutagenic effects of chlorpyrifos in freshwater fish Channa punctatus (Bloch) using micronucleus assay and alkaline single-cell gel electrophoresis. Food Chem. Toxicol. 2009, 47, 650–656. [Google Scholar] [CrossRef]
- dos Santos, K.C.; Martinez, C.B.R. Genotoxic and biochemical effects of atrazine and Roundup®, alone and in combination, on the Asian clam Corbicula fluminea. Ecotoxicol. Environ. Saf. 2014, 100, 7–14. [Google Scholar] [CrossRef]
- Ambreen, F.; Javed, M. Pesticide mixture induced DNA damage in peripheral blood erythrocytes of freshwater fish, Oreochromis niloticus. Pak. J. Zool. 2018, 50, 339–346. [Google Scholar] [CrossRef]
- Obiakor, M.O.; Okonkwo, J.C.; Ezeonyejiaku, C.D.; Ezenwelu, C.O. Genotoxicology: Single and Joint Action of Copper and Zinc to Synodontis clarias and Tilapia nilotica. J. Appl. Sci. Environ. Manag. 2010, 14, 59–64. [Google Scholar] [CrossRef][Green Version]
- Ahmad, I.; Hamid, T.; Fatima, M.; Chand, H.S.; Jain, S.K.; Athar, M.; Raisuddin, S. Induction of hepatic antioxidants in freshwater catfish (Channa punctatus Bloch) is a biomarker of paper mill effluent exposure. Biochim. Biophys. 2000, 1523, 37–48. [Google Scholar] [CrossRef]
- Marques, A.; Guilherme, S.; Gaivão, I.; Santos, M.A.; Pacheco, M. Progression of DNA damage induced by a glyphosate-based herbicide in fish (Anguilla anguilla) upon exposure and post-exposure periods—Insights into the mechanisms of genotoxicity and DNA repair. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2014, 166, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.; Yum, S.; Park, H.S.; Lee, T.K.; Ryu, J.C. Effects of heavy metals on antioxidants and stress-responsive gene expression in Javanese medaka (Oryzias javanicus). Comp. Biochem. Physiol. Toxicol. Pharmacol. 2009, 149, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Casanova, F.M.; Honda, R.T.; Ferreira-nozawa, M.S. Effects of Copper and Cadmium Exposure on mRNA Expression of Catalase, Glutamine Synthetase, Cytochrome P450 and Heat Shock Protein 70 in Tambaqui Fish (Colossoma macropomum). Gene Expr. Genet. Genom. 2013, 6, 1. [Google Scholar] [CrossRef]
- Benedetto, A.; Brizio, P.; Squadrone, S.; Scanzio, T.; Righetti, M.; Gasco, L.; Prearo, M.; Abete, M.C. Oxidative stress related to chlorpyrifos exposure in rainbow trout: Acute and medium term effects on genetic biomarkers. Pestic. Biochem. Physiol. 2016, 129, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, D.; Chattopadhyay, A. Induction of oxidative stress and related transcriptional effects of sodium fluoride in female zebrafish liver. Bull. Environ. Contam. Toxicol. 2014, 93, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The role of metallothionein in oxidative stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef]
- Chen, Q.; Vazquez, E.J.; Moghaddas, S.; Hoppel, C.L.; Lesnefsky, E.J. Production of reactive oxygen species by mitochondria: Central role of complex III. J. Biol. Chem. 2003, 278, 36027–36031. [Google Scholar] [CrossRef] [PubMed]
- Achard-Joris, M.; Gonzalez, P.; Marie, V.; Baudrimont, M.; Bourdineaud, J.P. Cytochrome c oxydase subunit I gene is up-regulated by cadmium in freshwater and marine bivalves. BioMetals 2006, 19, 237–244. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | Accession Number | Primer (5′–3′) |
|---|---|---|
| rpl7 | NM_001160672.2 | GGTCGCTCTCACAGACAACA a TTATGTCCGTCCCTCTGGGT b |
| cat | FJ226382.1 | CAGGTGTCTTTCTTGTTCAG a GTCCAGGATGGGAAGTTGC b |
| sod Cu/Zn | NM_001124329.1 | TGATTGGGGAGATCTCGGGT a CGGGTCCAGTGAGAGTCAAC b |
| gst | BT073173.1 | ATTTTGGGACGGGCTGACA a CCTGGTGCTCTGCTCCAGT b |
| cox1 | KP013084.1 | TCGTTTGAGCCGTGCTAGTT a CTTCTGGGTGGCCGAAGAAT b |
| 12S | KY798500.1 | GCGCCAGCTTAAAACCCAAA a GCCCATTTCTTCCCACCTCA b |
| ogg1 | XR_002474791.1 | CTGATGGACAAGGCCAGTGT a GTAAGGACCCCATGGCTGTC b |
| rad51 | XM_021612309.1 | AGGCTGGAGGAGGACATCAT a GTATTTGAGGGTGGCAGCCT b |
| bax | BT074328.1 | CAGAAAACCCAGGGAGGCAT a AGAACACATCCTGGGCACAG b |
| mt1 | M18104.1 | GTGGATCCTGCAAGTGCTCA a GTAATGCACCAGGCCTCACT b |
| ache | XM_021577686 | AGGAGGGTTCTACAGCGGAT a TATCCTGGACCCACTGGAGG b |
| CPF (µg/L) 1 | Cu2+ (µg/L) 2 | GLY (µg/L) 3 | |||||
|---|---|---|---|---|---|---|---|
| Nominal Concentration | Measured Concentration | Nominal Concentration | Measured Concentration | Nominal Concentration | Measured Concentration | ||
| Acidified (HNO3) | No EDTA | EDTA | |||||
| Condition B | 0.1 | T0 < 0.04 | 0.67 | T0 < 1.1 | 33.3 | T0 63.3 | 64.7 |
| T24 < 0.04 | T24 < 1.1 | T24 67.5 | 49.2 | ||||
| T48 < 0.04 | T48 < 1.1 | T48 55.4 | 42.2 | ||||
| Condition C | 0.3 | T0 0.18 | 2.0 | T0 2.03 | 100 | T0 160.0 | 175.8 |
| T24 < 0.04 | T24 4.16 | T24 140.0 | 168.0 | ||||
| T48 < 0.04 | T48 4.81 | T48 154.0 | 137.25 | ||||
| Condition D | 3.0 | T0 1.63 | 20.0 | T0 19.63 | 1000 | T0 1345.0 | 1785.0 |
| T24 0.1 | T24 26.32 | T24 1645.0 | 2022.5 | ||||
| T48 0.05 | T48 26.65 | T48 1775.0 | 1790.0 | ||||
| Control A | Condition B | Condition C | Condition D | |
|---|---|---|---|---|
| Acute toxicity | ||||
| Embryo viability (%) | 98.7 ± 2.3 | 96.8 ± 3.4 | 96.4 ± 1.5 | 94.2 ± 4.1 |
| Larval viability (%) | 97.7 ± 2.1 | 94.5 ± 4.8 | 96.0 ± 3.2 | 95.2 ± 4.9 |
| Cumulative viability (%) | 96.4 ± 0.8 | 91.7 ± 7.3 | 92.6 ± 3.1 | 89.5 ± 3.5 |
| Hatching rate (%) | 95.6 ± 2.0 | 92.0 ± 4.6 | 92.0 ± 4.8 | 90.2 ± 4.1 |
| Sub-lethal toxicity | ||||
| Duration of development (DD) | 301.1 ± 2.2 | 294.9 ± 12.2 | 305.7 ± 1.1 | 309.9 ± 8.3 |
| Total length (mm) | 19.5 ± 0.3 | 19.6 ± 0.2 | 19.8 ± 0.2 | 19.1 ± 0.2 |
| Head length (mm) | 4.5 ± 0.0 | 4.5 ± 0.1 | 4.6 ± 0.0 | 4.3 ± 0.2 |
| Ratio of head/body length (%) | 23.2 ± 0.1 | 23.1 ± 0.2 | 23.1 ± 0.1 | 22.4 ± 0.7 |
| Area of yolk sac (mm2) | 10.8 ± 0.2 | 10.1 ± 0.4 | 10.4 ± 0.5 | 10.5 ± 0.1 |
| Abnormalities (%) | ||||
| Total | 15.0 ± 2.8 | 23.7 ± 3.6 | 15.0 ± 7.8 | 22.8 ± 11.3 |
| Oedemas | 7.5 ± 8.4 | 12.7 ± 6.2 | 9.3 ± 5.6 | 11.9 ± 6.5 |
| Spinal | 7.5 ± 8.4 | 16.6 ± 11.2 | 12.9 ± 11.3 | 16.5 ± 14.8 |
| Craniofacial | 1.8 ± 3.2 | 5.4 ± 5.3 | 11.3 ± 4.5 | 14.0 ± 4.5 |
| Haemorrhages | 7.6 ± 6.6 | 0.0 ± 0.0 | 9.3 ± 5.6 | 11.9 ± 6.5 |
| Gene | Condition B | Condition C | Condition D |
|---|---|---|---|
| cat | / | / | / |
| sod | / | / | / |
| gst | 2.5 * | / | / |
| cox1 | 5.3 * | 3.8 * | 23.8 * |
| 12s | 4.9 * | 5.2 * | 32.4 * |
| ogg1 | 4.4 * | 6.3 * | 34.8 * |
| rad51 | / | / | / |
| bax | / | / | / |
| mt1 | 9.4 * | / | 12.7 * |
| ache | 4.5 * | 7.0 * | 36.7 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weeks Santos, S.; Cachot, J.; Cormier, B.; Mazzella, N.; Gourves, P.-Y.; Clérandeau, C.; Morin, B.; Gonzalez, P. Environmentally Relevant Mixture of Pesticides Affect Mobility and DNA Integrity of Early Life Stages of Rainbow Trout (Oncorhynchus mykiss). Toxics 2021, 9, 174. https://doi.org/10.3390/toxics9080174
Weeks Santos S, Cachot J, Cormier B, Mazzella N, Gourves P-Y, Clérandeau C, Morin B, Gonzalez P. Environmentally Relevant Mixture of Pesticides Affect Mobility and DNA Integrity of Early Life Stages of Rainbow Trout (Oncorhynchus mykiss). Toxics. 2021; 9(8):174. https://doi.org/10.3390/toxics9080174
Chicago/Turabian StyleWeeks Santos, Shannon, Jérôme Cachot, Bettie Cormier, Nicolas Mazzella, Pierre-Yves Gourves, Christelle Clérandeau, Bénédicte Morin, and Patrice Gonzalez. 2021. "Environmentally Relevant Mixture of Pesticides Affect Mobility and DNA Integrity of Early Life Stages of Rainbow Trout (Oncorhynchus mykiss)" Toxics 9, no. 8: 174. https://doi.org/10.3390/toxics9080174
APA StyleWeeks Santos, S., Cachot, J., Cormier, B., Mazzella, N., Gourves, P.-Y., Clérandeau, C., Morin, B., & Gonzalez, P. (2021). Environmentally Relevant Mixture of Pesticides Affect Mobility and DNA Integrity of Early Life Stages of Rainbow Trout (Oncorhynchus mykiss). Toxics, 9(8), 174. https://doi.org/10.3390/toxics9080174