
Bisphenol A Disrupts Ribosome Function during Ovarian Development of Mice


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Chemicals
2.2. Sample Collection
2.3. Total BPA in Serum Determination
2.4. Immunohistochemistry Staining
2.5. Apoptosis of Ovarian Cells
2.6. RNA Extraction and Quality Determination
2.7. Total RNA Sequencing
2.8. Real-Time PCR
2.9. Statistical Analysis
3. Results
3.1. BPA Exposure Increased BPA Contents in Serum F1 Females
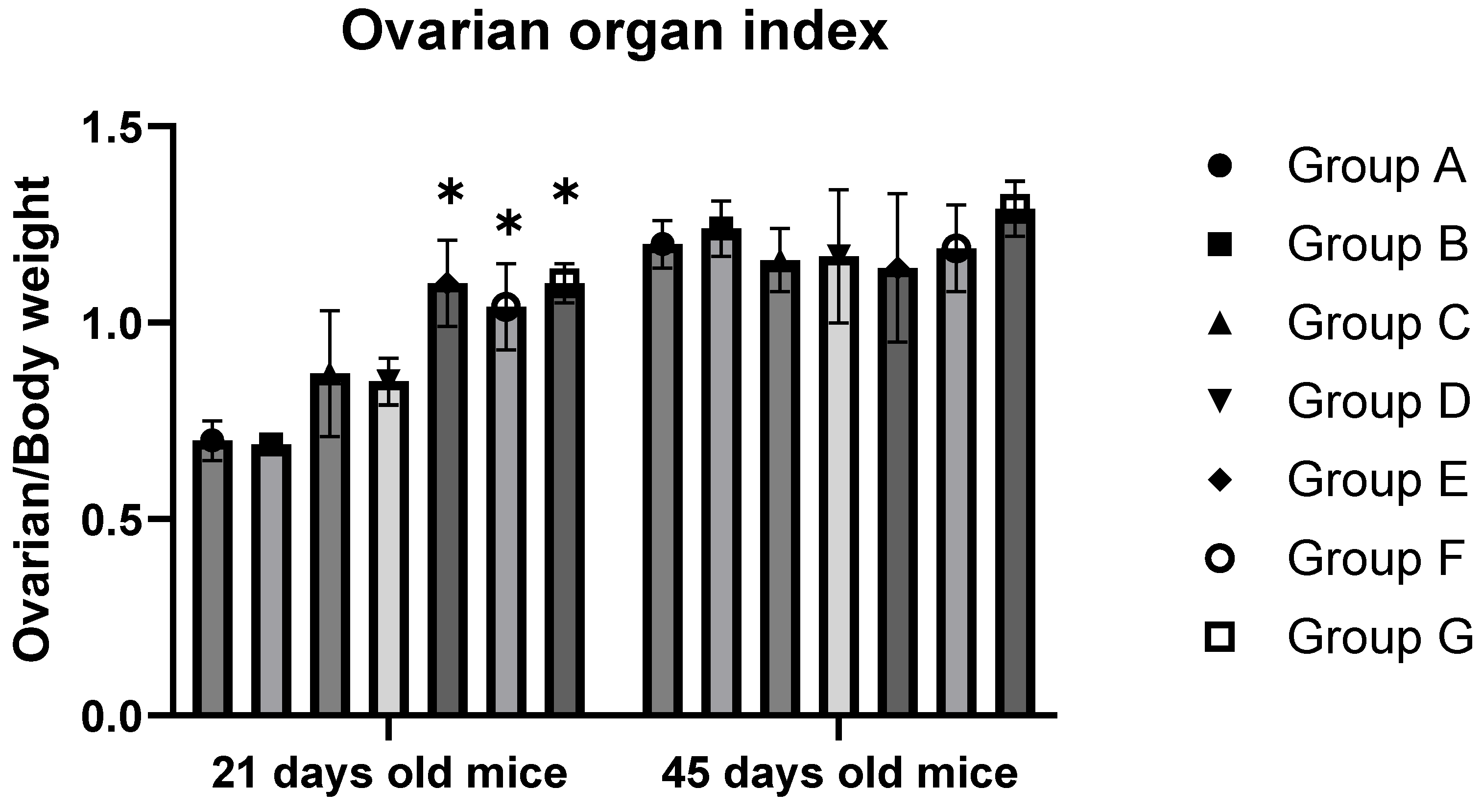
3.2. BPA Exposure Elevates the Body Weight, Ovarian Weight, and Ovarian Organ Index of 21-Day-Old F1 Females
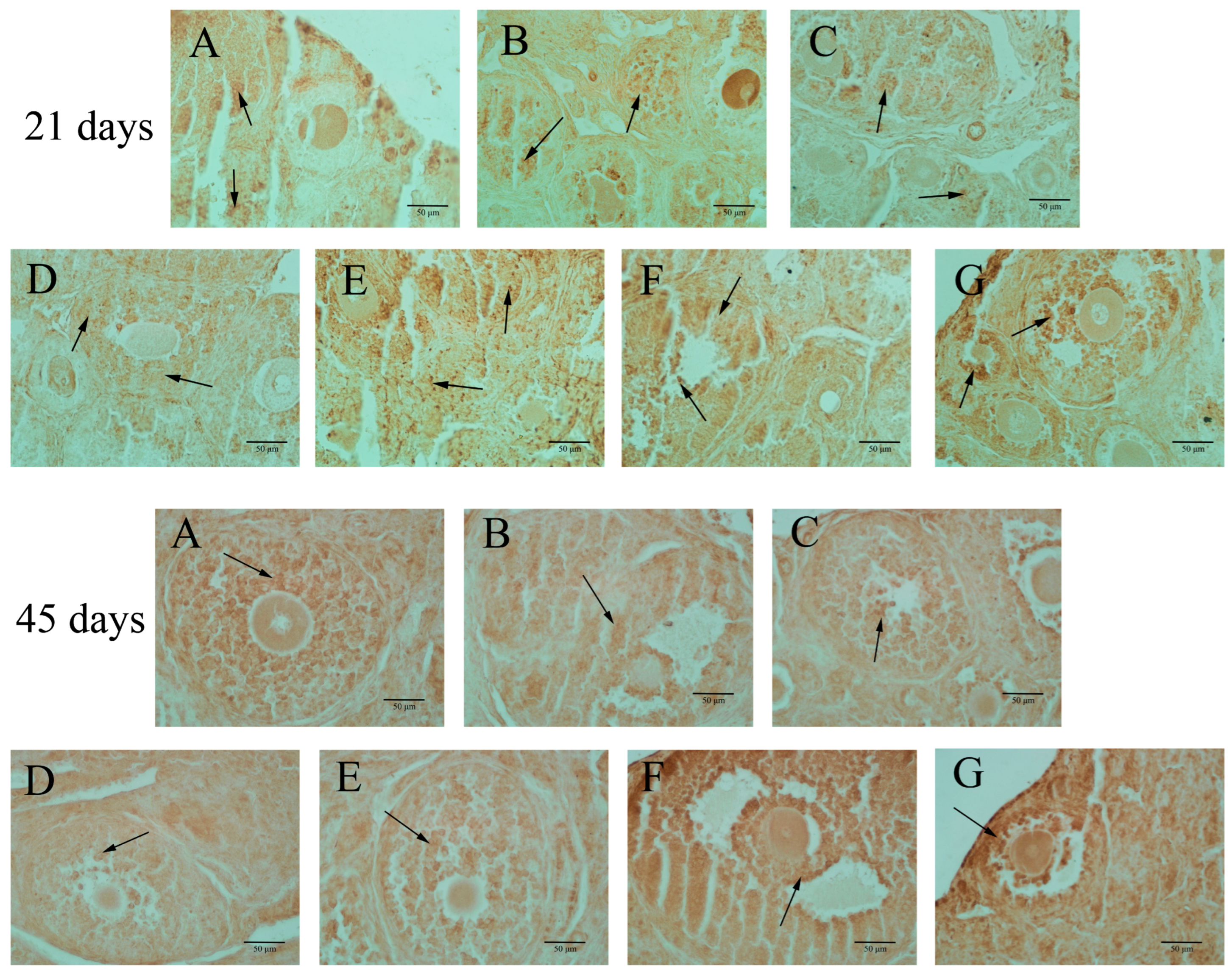
3.3. Apoptosis in Ovarian Granulosa Cells
3.4. ERβ Expressions in the Ovary of F1 Females
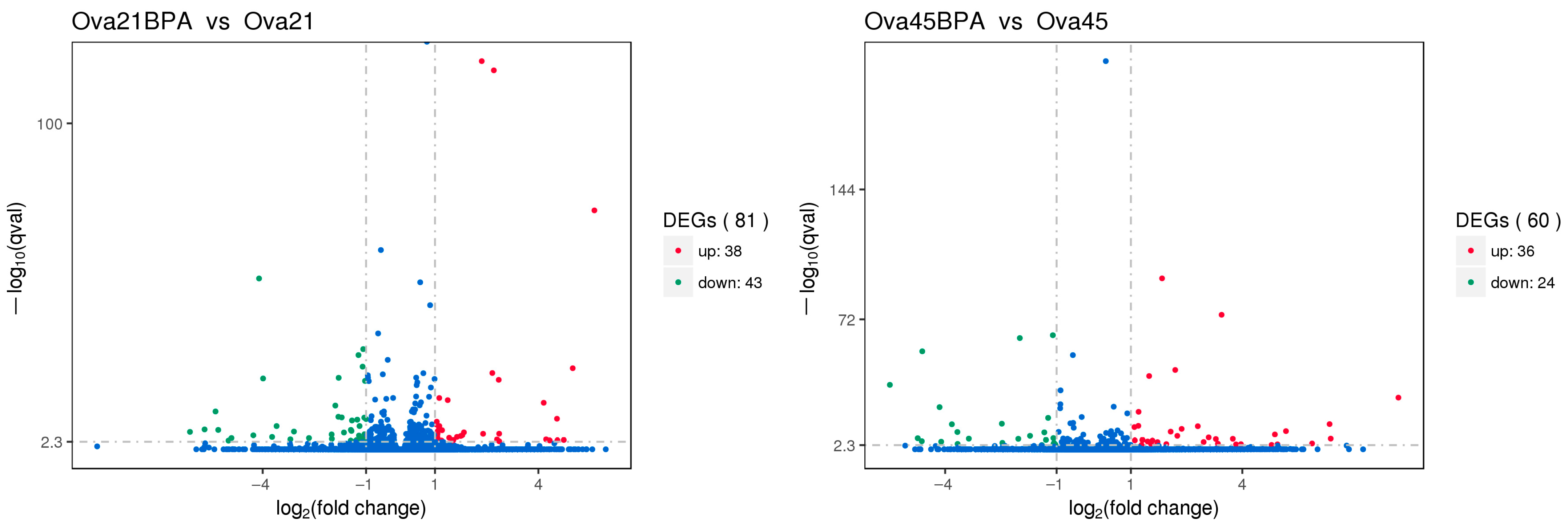
3.5. Differentially Expressed Genes in the Ovary
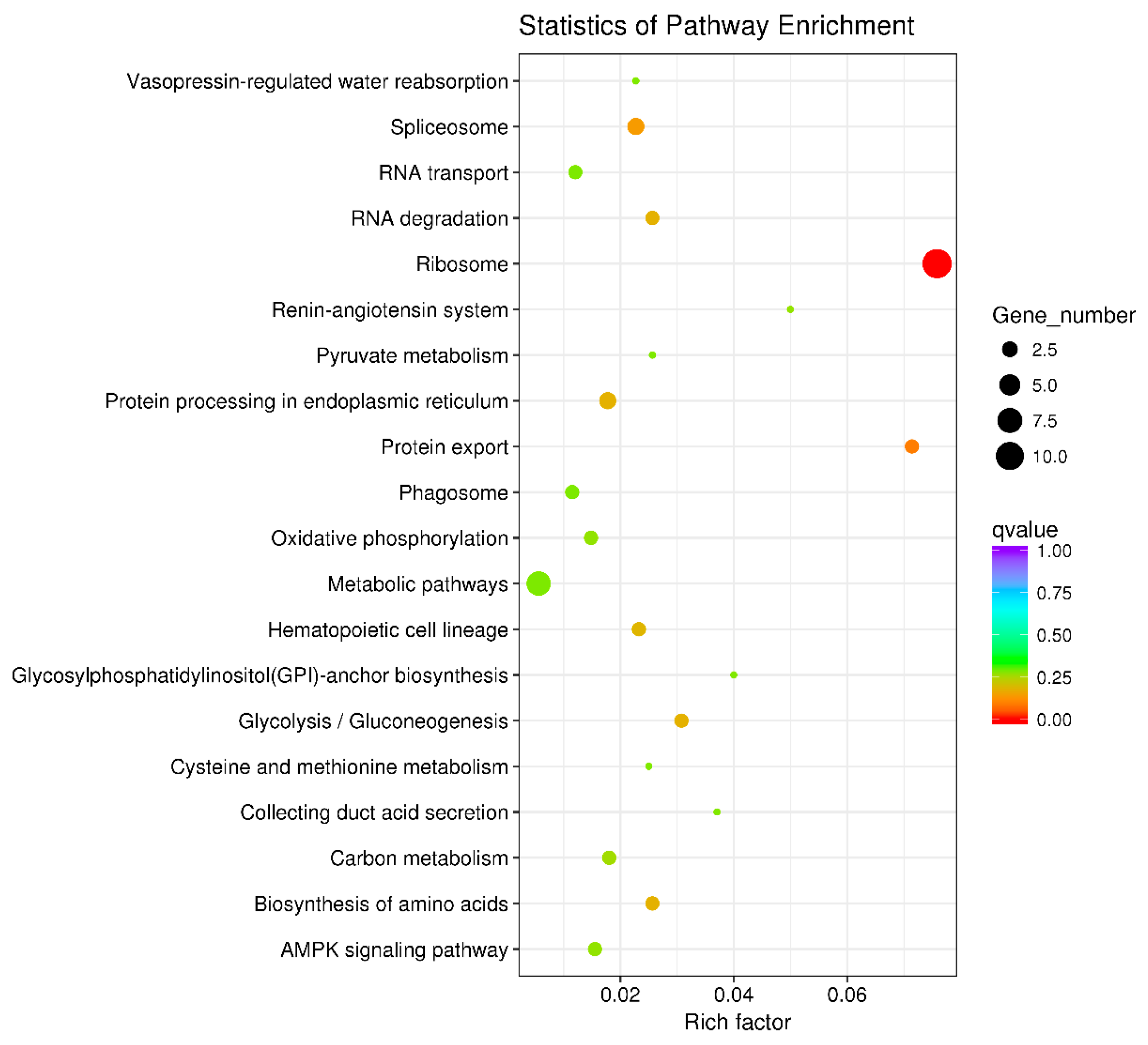
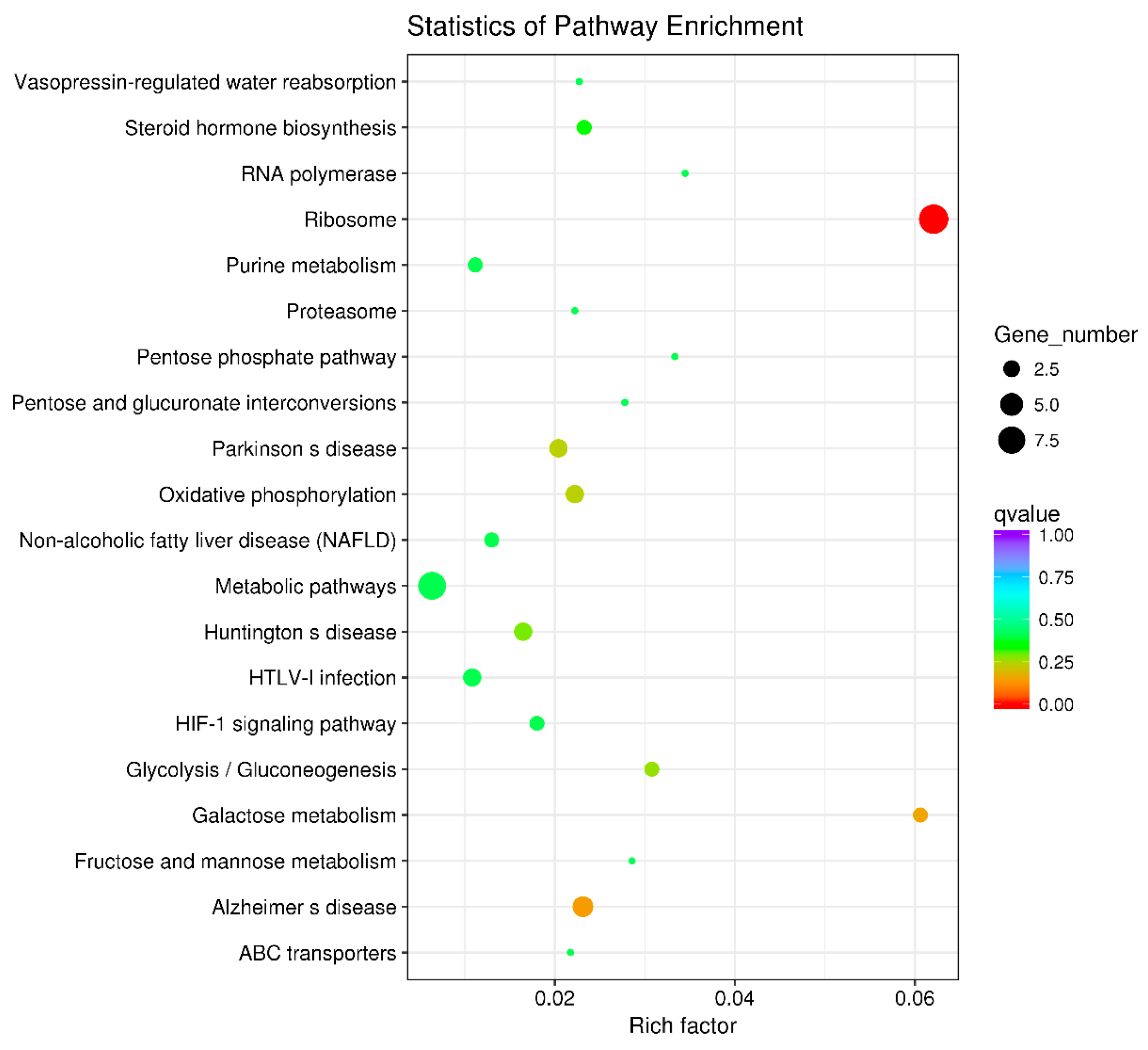
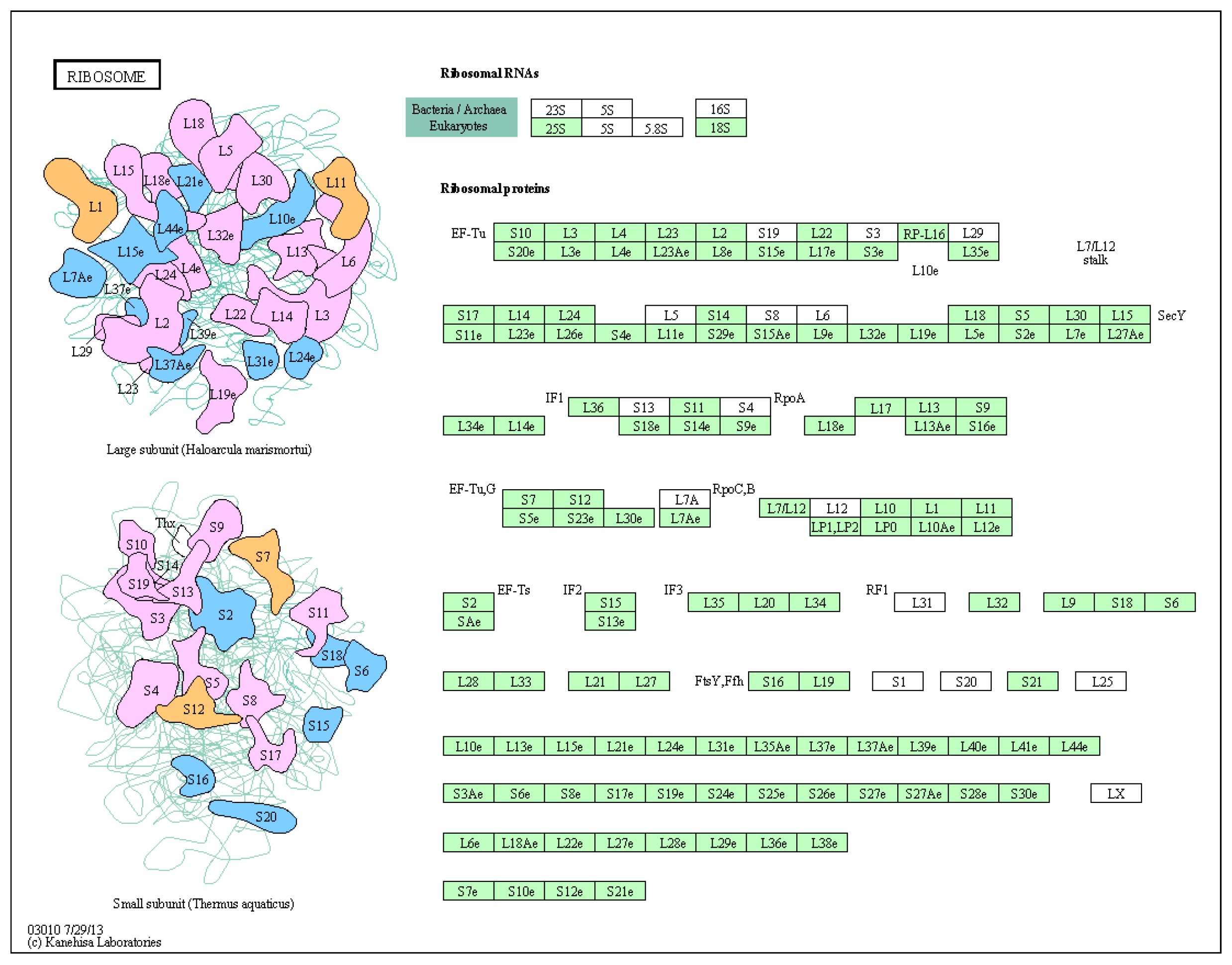
3.6. Key Metabolic Pathways Affected by BPA in Ovarian Development
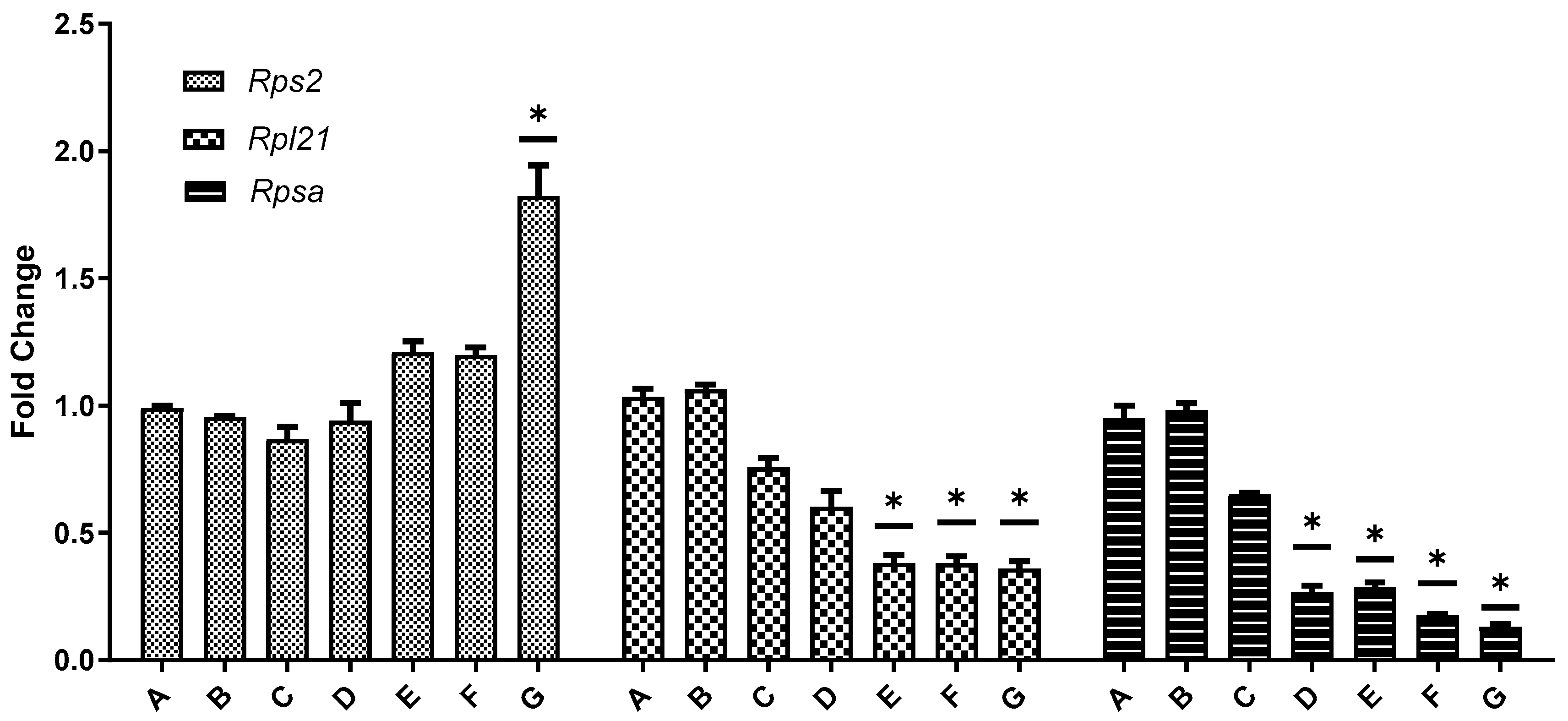
3.7. Gene Expression of Rps2, Rpl21, and Rpsa
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tarafdar, A.; Sirohi, R.; Balakumaran, P.A.; Reshmy, R.; Madhavan, A.; Sindhu, R.; Binod, P.; Kumar, Y.; Kumar, D.; Sim, S.J. The hazardous threat of Bisphenol A: Toxicity, detection and remediation. J. Hazard. Mater. 2022, 423, 127097. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xiao, Y.; Gai, Z.; Li, R.; Zhu, Z.; Bai, C.; Tanguay, R.L.; Xu, X.; Huang, C.; Dong, Q. Reproductive toxicity of low level bisphenol A exposures in a two-generation zebrafish assay: Evidence of male-specific effects. Aquat. Toxicol. 2015, 169, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Vermeirssen, E.L.; Dietschweiler, C.; Werner, I.; Burkhardt, M. Corrosion protection products as a source of bisphenol A and toxicity to the aquatic environment. Water Res. 2017, 123, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Pinto, C.; Hao, R.; Grimaldi, M.; Thrikawala, S.; Boulahtouf, A.; Ait-Aissa, S.; Brion, F.; Gustafsson, J.-Å.; Balaguer, P.; Bondesson, M. Differential activity of BPA, BPAF and BPC on zebrafish estrogen receptors in vitro and in vivo. Toxicol. Appl. Pharmacol. 2019, 380, 114709. [Google Scholar] [CrossRef]
- Ocharoen, Y.; Boonphakdee, C.; Boonphakdee, T.; Shinn, A.P.; Moonmangmee, S. High levels of the endocrine disruptors bisphenol-A and 17β-estradiol detected in populations of green mussel, Perna viridis, cultured in the Gulf of Thailand. Aquaculture 2018, 497, 348–356. [Google Scholar] [CrossRef]
- Khakimov, B.; Bakhytkyzy, I.; Fauhl-Hassek, C.; Engelsen, S.B. Non-volatile molecular composition and discrimination of single grape white wines of Chardonnay, Riesling, Sauvignon Blanc and Silvaner using untargeted GC-MS analysis. Food Chem. 2022, 369, 130878. [Google Scholar] [CrossRef]
- Cirillo, T.; Esposito, F.; Fasano, E.; Scognamiglio, G.; Di Marco Pisciottano, I.; Mita, G.D.; Gallo, P. BPA, BPB, BPF, BADGE and BFDGE in canned beers from the Italian market. Food Addit. Contam. Part B 2019, 12, 268–274. [Google Scholar] [CrossRef]
- Abulehia, H.F.; Mohd Nor, N.S.; Sheikh Abdul Kadir, S.H. The Current findings on the impact of prenatal BPA exposure on metabolic parameters: In vivo and epidemiological evidence. Nutrients 2022, 14, 2766. [Google Scholar] [CrossRef] [PubMed]
- Kobroob, A.; Peerapanyasut, W.; Chattipakorn, N.; Wongmekiat, O. Damaging effects of bisphenol A on the kidney and the protection by melatonin: Emerging evidences from in vivo and in vitro studies. Oxidative Med. Cell. Longev. 2018, 2018, 3082438. [Google Scholar] [CrossRef]
- Foster, W.G.; Kubwabo, C.; Kosarac, I.; Gregorovich, S.; Aryal, G.; Coleman, K. Free bisphenol A (BPA), BPA-Glucuronide (BPA-G), and total BPA concentrations in maternal serum and urine during pregnancy and umbilical cord blood at delivery. Emerg. Contam. 2019, 5, 279–287. [Google Scholar] [CrossRef]
- Pan, Y.; Deng, M.; Li, J.; Du, B.; Lan, S.; Liang, X.; Zeng, L. Technology: Occurrence and maternal transfer of multiple bisphenols, including an emerging derivative with unexpectedly high concentrations, in the human maternal–fetal–placental unit. Environ. Sci. Technol. 2020, 54, 3476–3486. [Google Scholar] [CrossRef] [PubMed]
- Nomiri, S.; Hoshyar, R.; Ambrosino, C.; Tyler, C.R.; Mansouri, B. A mini review of bisphenol A (BPA) effects on cancer-related cellular signaling pathways. Environ. Sci. Pollut. Res. 2019, 26, 8459–8467. [Google Scholar] [CrossRef] [PubMed]
- Jalal, N.; Surendranath, A.R.; Pathak, J.L.; Yu, S.; Chung, C. Bisphenol A (BPA) the mighty and the mutagenic. Toxicol. Rep. 2018, 5, 76–84. [Google Scholar] [CrossRef]
- Babiloni-Chust, I.; Dos Santos, R.S.; Medina-Gali, R.M.; Perez-Serna, A.A.; Encinar, J.-A.; Martinez-Pinna, J.; Gustafsson, J.-A.; Marroqui, L.; Nadal, A. G protein-coupled estrogen receptor activation by bisphenol-A disrupts the protection from apoptosis conferred by the estrogen receptors ERα and ERβ in pancreatic beta cells. Environ. Int. 2022, 164, 107250. [Google Scholar] [CrossRef] [PubMed]
- Shelby, M.D. NTP-CERHR monograph on the potential human reproductive and developmental effects of bisphenol A. NTP CERHR MON 2008, vii–ix. [Google Scholar]
- Vom Saal, F.S.; Vandenberg, L.N. Update on the health effects of bisphenol A: Overwhelming evidence of harm. Endocrinology 2021, 162, bqaa171. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, H.; Liu, S. Low-dose bisphenol A exposure: A seemingly instigating carcinogenic effect on breast cancer. Adv. Sci. 2017, 4, 1600248. [Google Scholar] [CrossRef]
- Leung, Y.-K.; Biesiada, J.; Govindarajah, V.; Ying, J.; Kendler, A.; Medvedovic, M.; Ho, S.-M. Low-dose bisphenol a in a rat model of endometrial cancer: A CLARITY-BPA study. Environ. Health Perspect. 2020, 128, 127005. [Google Scholar] [CrossRef]
- Huang, M.; Huang, M.; Li, X.; Liu, S.; Fu, L.; Jiang, X.; Yang, M. Bisphenol A induces apoptosis through GPER-dependent activation of the ROS/Ca2+-ASK1-JNK pathway in human granulosa cell line KGN. Ecotoxicol. Environ. Saf. 2021, 208, 111429. [Google Scholar] [CrossRef]
- Tang, Z.-R.; Xu, X.-L.; Deng, S.-L.; Lian, Z.-X.; Yu, K. Oestrogenic endocrine disruptors in the placenta and the fetus. Int. J. Mol. Sci. 2020, 21, 1519. [Google Scholar] [CrossRef]
- Martínez-Peña, A.A.; Peña-Castillo, A.; Parra-Forero, L.Y.; Hernández-Ochoa, I.; Hernández-Barrientos, L.R.; Morimoto, S.; Mendoza-Rodríguez, C.A. Parental perinatal exposure to bisphenol A reduces the threshold to disrupt blastocyst implantation via decreasing talin, occudin and E-cadherin levels. Reprod. Toxicol. 2019, 86, 86–97. [Google Scholar] [CrossRef]
- Collins, T.J. ImageJ for microscopy. Biotechniques 2007, 43, S25–S30. [Google Scholar] [CrossRef] [PubMed]
- Garcia, T.I.; Shen, Y.; Crawford, D.; Oleksiak, M.F.; Whitehead, A.; Walter, R.B. RNA-Seq reveals complex genetic response to deepwater horizon oil release in Fundulus grandis. BMC Genom. 2012, 13, 474. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Leng, N.; Dawson, J.A.; Thomson, J.A.; Ruotti, V.; Rissman, A.I.; Smits, B.M.; Haag, J.D.; Gould, M.N.; Stewart, R.M.; Kendziorski, C. EBSeq: An empirical Bayes hierarchical model for inference in RNA-seq experiments. Bioinformatics 2013, 29, 1035–1043. [Google Scholar] [CrossRef]
- Stahlhut, R.W.; Welshons, W.V.; Swan, S.H. Bisphenol A data in NHANES suggest longer than expected half-life, substantial nonfood exposure, or both. Environ. Health Perspect. 2009, 117, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Cousins, I.; Staples, C.; Kleĉka, G.; Mackay, D. A multimedia assessment of the environmental fate of bisphenol A. Hum. Ecol. Risk Assess. 2002, 8, 1107–1135. [Google Scholar] [CrossRef]
- Agostinho, B.; Silvestre, A.J.; Coutinho, J.A.; Sousa, A.F. Synthetic (bio) degradable polymers–when does recycling fail? Green Chem. 2023, 25, 13–31. [Google Scholar] [CrossRef]
- Qi, C.; Huang, J.; Wang, B.; Deng, S.; Wang, Y.; Yu, G. Contaminants of emerging concern in landfill leachate in China: A review. Emerg. Contam. 2018, 4, 1–10. [Google Scholar] [CrossRef]
- Bousoumah, R.; Leso, V.; Iavicoli, I.; Huuskonen, P.; Viegas, S.; Porras, S.P.; Santonen, T.; Frery, N.; Robert, A.; Ndaw, S. Biomonitoring of occupational exposure to bisphenol A, bisphenol S and bisphenol F: A systematic review. Sci. Total Environ. 2021, 783, 146905. [Google Scholar] [CrossRef]
- Lucarini, F.; Krasniqi, T.; Bailat Rosset, G.; Roth, N.; Hopf, N.B.; Broillet, M.-C.; Staedler, D. Exposure to new emerging bisphenols among young children in Switzerland. Int. J. Environ. Res. Public Health 2020, 17, 4793. [Google Scholar] [CrossRef]
- Nesan, D.; Feighan, K.M.; Antle, M.C.; Kurrasch, D.M. Gestational low-dose BPA exposure impacts suprachiasmatic nucleus neurogenesis and circadian activity with transgenerational effects. Sci. Adv. 2021, 7, eabd1159. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, S.-W.; Wang, L.; Sun, Y.; Xu, F.; He, H.; Wang, S.; Zhang, Z.; Pan, X. Interfering effects of bisphenol A on in vitro growth of preantral follicles and maturation of oocyes. Clin. Chim. Acta 2018, 485, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Kechagias, K.S.; Semertzidou, A.; Athanasiou, A.; Paraskevaidi, M.; Kyrgiou, M.J. Bisphenol-A and polycystic ovary syndrome: A review of the literature. Rev. Environ. Health 2020, 35, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Kawa, I.A.; Fatima, Q.; Mir, S.A.; Jeelani, H.; Manzoor, S.; Rashid, F. Endocrine disrupting chemical Bisphenol A and its potential effects on female health. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 803–811. [Google Scholar] [CrossRef]
- Liu, Y.; Qu, K.; Hai, Y.; Zhao, C. Bisphenol A (BPA) binding on full-length architectures of estrogen receptor. J. Cell. Biochem. 2018, 119, 6784–6794. [Google Scholar] [CrossRef]
- Birgersson, M.; Indukuri, R.; Antonson, P.; Nalvarte, I.; Archer, A.; Williams, C. ERβ in granulosa cell tumors and its clinical potential. Endocrinology 2023, 164, bqad063. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, J.; Zhang, C. Synthesis, regulatory factors, and signaling pathways of estrogen in the ovary. Reprod. Sci. 2023, 30, 350–360. [Google Scholar] [CrossRef]
- Chakravarthi, V.P.; Ghosh, S.; Housami, S.M.; Wang, H.; Roby, K.F.; Wolfe, M.W.; Kinsey, W.H.; Rumi, M.K. ERβ regulated ovarian kisspeptin plays an important role in oocyte maturation. Mol. Cell. Endocrinol. 2021, 527, 111208. [Google Scholar] [CrossRef]
- Zhang, S.-X.; Ding, Z.-M.; Ahmad, M.J.; Wang, Y.-S.; Duan, Z.-Q.; Miao, Y.-L.; Xiong, J.-J.; Huo, L.-J. Bisphenol B exposure disrupts mouse oocyte meiotic maturation in vitro through affecting spindle assembly and chromosome alignment. Front. Cell Dev. Biol. 2020, 8, 616771. [Google Scholar] [CrossRef]
- Mesnage, R.; Phedonos, A.; Arno, M.; Balu, S.; Corton, J.C.; Antoniou, M.N. Transcriptome profiling reveals bisphenol A alternatives activate estrogen receptor alpha in human breast cancer cells. bioRxiv 2017. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Masuya, T.; Hosose, M.; Tagawa, K.; Ishibashi, T.; Suyama, K.; Nose, T.; Yoshihara, E.; Downes, M.; Evans, R.M.; et al. Bisphenol A derivatives act as novel coactivator-binding inhibitors for estrogen receptor β. J. Biol. Chem. 2021, 297, 101173. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodriguez, D.; Franssen, D.; Heger, S.; Parent, A.-S. Endocrine-disrupting chemicals and their effects on puberty. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101579. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, J.; Zhang, E.; Wu, Q.; Wu, X.; Zhang, D.; Liu, Y.; Wang, R.; Li, W. Bisphenol A affects ovarian development in adolescent mice caused by genes expression change. Gene 2020, 740, 144535. [Google Scholar] [CrossRef]
- Marchingo, J.M.; Cantrell, D.A. Protein synthesis, degradation, and energy metabolism in T cell immunity. Cell. Mol. Immunol. 2022, 19, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Rayinda, T.; McSweeney, S.M.; Fassihi, H.; Fenton, D.; Liu, L.; Stefanato, C.M.; Dand, N.; McGrath, J.A.; Tziotzios, C. A novel heterozygous missense variant in ribosomal protein L21 associated with familial hypotrichosis simplex. Clin. Exp. Dermatol. 2023, 48, 840–843. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Long, T.; Gao, L.; Zhong, Y.; Wang, P.; Wang, X.; Li, Z.; Hu, Z. RPL21 interacts with LAMP3 to promote colorectal cancer invasion and metastasis by regulating focal adhesion formation. Cell. Mol. Biol. Lett. 2023, 28, 31. [Google Scholar] [CrossRef]
- Landry-Voyer, A.-M.; Bergeron, D.; Yague-Sanz, C.; Baker, B.; Bachand, F. PDCD2 functions as an evolutionarily conserved chaperone dedicated for the 40S ribosomal protein uS5 (RPS2). Nucleic Acids Res. 2020, 48, 12900–12916. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Sequence (5′-3′) | GC% | Tm | Length (bp) | |
|---|---|---|---|---|---|
| Rps2 | F | CCTGATGATGGCCGGTATAGATG | 52.2 | 64.7 | 133 |
| R | TTTCCAGAGGTCGGGAGTCAG | 57.1 | 63.9 | ||
| Rpl21 | F | CCGTGGGCATCATTGTCAAC | 55 | 64.6 | 71 |
| R | CTCAATCCGCACATTGATCCTC | 50 | 64 | ||
| Rpsa | F | ACTTAGGTGGCACCAACCTTGAC | 52.2 | 63.7 | 117 |
| R | GAGCTGCGAGCAACAGCTTC | 60 | 63.8 | ||
| β-Actin | F | TCCTTCCTGGGCATGGAGT | 57.9 | 63 | 104 |
| R | AGCACTGTGTTGGCGTACAG | 55 | 60 | ||
| Group | 21 Days Old Mice | 45 Days Old Mice | ||||
|---|---|---|---|---|---|---|
| Body Weight (g) | Bilateral Ovarian Weight (mg) | Ovarian Organ Index (mg/g) | Body Weight (g) | Bilateral Ovarian Weight (mg) | Ovarian Organ Index (mg/g) | |
| Group A | 16.67 ± 0.53 bc | 11.57 ± 0.58 a | 0.70 ± 0.05 a | 30.39 ± 2.54 a | 36.80 ± 4.31 a | 1.20 ± 0.06 a |
| Group B | 19.03 ± 2.21 c | 13.11 ± 1.39 ab | 0.69 ± 0.01 a | 32.94 ± 1.87 a | 40.83 ± 2.52 a | 1.24 ± 0.07 a |
| Group C | 17.83 ± 1.46 b | 15.60 ± 2.97 b | 0.87 ± 0.16 ab | 31.54 ± 0.19 a | 36.57 ± 2.54 a | 1.16 ± 0.08 a |
| Group D | 18.40 ± 1.60 c | 15.70 ± 1.70 b | 0.85 ± 0.06 ab | 29.44 ± 2.56 a | 34.87 ± 7.29 a | 1.17 ± 0.17 a |
| Group E | 14.23 ± 0.67 a | 15.67 ± 1.10 b | 1.10 ± 0.11 b | 32.02 ± 1.45 a | 36.90 ± 7.26 a | 1.14 ± 0.19 a |
| Group F | 14.47 ± 0.59 a | 15.07 ± 2.16 b | 1.04 ± 0.11 b | 33.15 ± 1.34 a | 39.8 ± 4.88 a | 1.19 ± 0.11 a |
| Group G | 14.87 ± 0.67 a | 16.32 ± 1.26 b | 1.10 ± 0.05 b | 35.71 ± 1.93 a | 45.93 ± 3.26 a | 1.29 ± 0.07 a |
| Groups | Ovary at PND 21 (Gray Value) | Ovary at PND 45 (Gray Value) |
|---|---|---|
| Group A | 72.61 ± 3.50 a | 10.22 ± 1.35 a |
| Group B | 71.22 ± 5.57 a | 7.69 ± 2.33 a |
| Group C | 90.00 ± 4.82 ab | 12.56 ± 6.77 a |
| Group D | 81.72 ± 7.62 a | 20.91 ± 6.97 ab |
| Group E | 77.17 ± 3.28 a | 27.14 ± 4.74 ab |
| Group F | 106.13 ± 10.52 b | 31.83 ± 12.13 ab |
| Group G | 131.71 ± 6.27 c | 38.83 ± 12.25 b |
| Groups | PND 21 ERβ (Gray Value) | PND 45 ERβ (Gray Value) |
|---|---|---|
| Group A | 13.05 ± 3.21 a | 8.61 ± 0.72 a |
| Group B | 13.37 ± 4.46 a | 9.40 ± 2.64 a |
| Group C | 8.81 ± 1.51 a | 7.45 ± 1.92 a |
| Group D | 8.25 ± 0.13 a | 4.74 ± 1.44 a |
| Group E | 15.90 ± 2.32 a | 8.72 ± 1.63 a |
| Group F | 20.02 ± 7.60 a | 30.94 ± 14.90 b |
| Group G | 20.67 ± 7.09 a | 42.12 ± 8.38 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Wang, Y.; Li, W.; Wang, K.; Zhang, S. Bisphenol A Disrupts Ribosome Function during Ovarian Development of Mice. Toxics 2024, 12, 627. https://doi.org/10.3390/toxics12090627
Ma X, Wang Y, Li W, Wang K, Zhang S. Bisphenol A Disrupts Ribosome Function during Ovarian Development of Mice. Toxics. 2024; 12(9):627. https://doi.org/10.3390/toxics12090627
Chicago/Turabian StyleMa, Xin, Yongjie Wang, Weiqi Li, Kaiyue Wang, and Shilei Zhang. 2024. "Bisphenol A Disrupts Ribosome Function during Ovarian Development of Mice" Toxics 12, no. 9: 627. https://doi.org/10.3390/toxics12090627
APA StyleMa, X., Wang, Y., Li, W., Wang, K., & Zhang, S. (2024). Bisphenol A Disrupts Ribosome Function during Ovarian Development of Mice. Toxics, 12(9), 627. https://doi.org/10.3390/toxics12090627