


Molecular Toxicity Mechanism Induced by the Antibacterial Agent Triclosan in Freshwater Euglena gracilis Based on the Transcriptome
Abstract

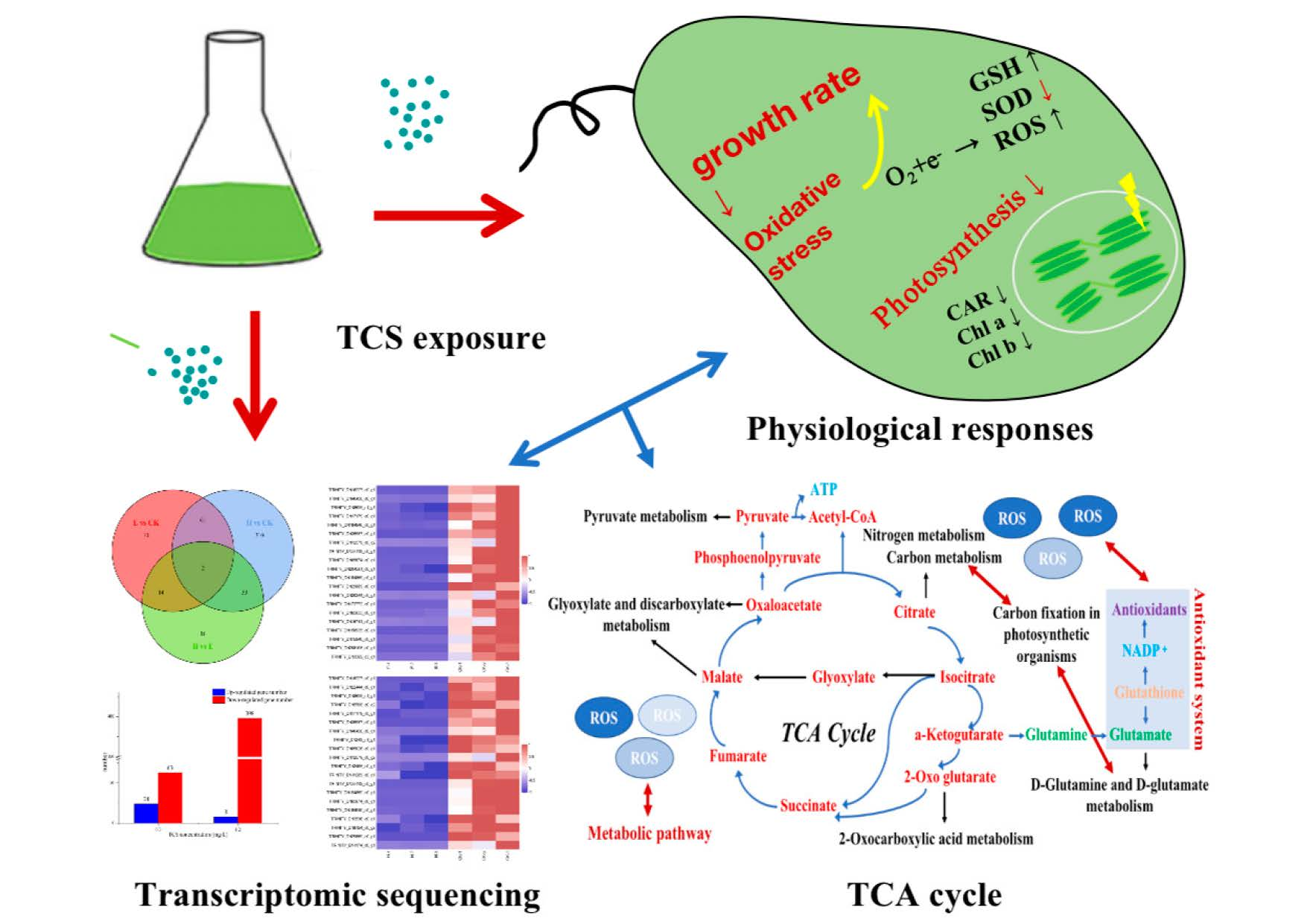{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Triclosan and Euglena Gracilis Cultivation
2.2. Triclosan Exposure
2.3. Transmission Electron Microscopy Analysis
2.4. Pigments Content
2.5. Oxidative Stress
2.6. RNA Extraction and Sequencing
2.7. De Novo Transcriptome Assembly
2.8. Differential Gene Expression
2.9. Statistical Analysis
3. Results
3.1. Dose–Effect Relationship between Triclosan and Euglena Gracilis Growth
3.2. Effects on Cell Morphology and Ultrastructure
3.3. Physiological Index Changes Induced by Triclosan Exposure
3.4. Transcriptome Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eren, G.; Egemen, A.; Emel, T.; Elif, P.-M. Effect of triclosan and its photolysis products on marine bacterium V. fischeri and freshwater alga R. subcapitata. J. Environ. Manag. 2018, 211, 218–224. [Google Scholar]
- Yueh, M.-F.; Tukey, R.-H. Triclosan: A widespread environmental toxicant with many biological effects. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 251–272. [Google Scholar] [CrossRef]
- Li, X.-Q.; Shang, Y.; Yao, W.-W. Comparison of transcriptomics changes induced by TCS and MTCS exposure in human hepatoma HepG2 cells. ACS Omega 2020, 5, 10715–10724. [Google Scholar] [CrossRef]
- Machado, M.-D.; Soares, E.-V. Toxicological effects induced by the biocide triclosan on Pseudokirchneriella subcapitata. Aquat. Toxicol. 2021, 230, 105706. [Google Scholar] [CrossRef]
- Xie, J.-H.; Zhao, N.; Zhang, Y.-Y.; Hu, H.-M.; Zhao, M.-R.; Jin, H.-B. Occurrence and partitioning of bisphenol analogues triclocarban and triclosan in seawater and sediment from East China Sea. Chemosphere 2022, 287, 132218. [Google Scholar] [CrossRef] [PubMed]
- Halden, R.-U. On the need and speed of regulating triclosan and triclocarban in the United States. Environ. Sci. Technol. 2014, 48, 3603–3611. [Google Scholar] [CrossRef]
- Montaseri, H.; Forbes, P.-B.-C. A review of monitoring methods for triclosan and its occurrence in aquatic environments. TrAC Trends Anal. Chem. 2016, 85, 221–231. [Google Scholar] [CrossRef]
- Wei, M.; Yang, X.; Watson, P.; Yang, F.; Liu, H. A cyclodextrin polymer membrane-based passive sampler for measuring triclocarban triclosan and methyl triclosan in rivers. Sci. Total Environ. 2019, 648, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Mao, B.; He, H.; Shang, Y.; Zhong, Y.; Yu, Z.; Yang, Y.; Li, H.; An, J. Comparison of hepatotoxicity and mechanisms induced by triclosan (TCS) and methyl-triclosan (MTCS) in human liver hepatocellular HepG2 cells. Toxicol. Res. 2019, 8, 38–45. [Google Scholar] [CrossRef]
- European Comission. Commission Implementing Decision (EU) 2016/110 of 27 January 2016 Not Approving Triclosan as an Existing Active Substance for Use in Biocidal Products for Product Type 1; European Union: Maastricht, The Netherlands, 2016. [Google Scholar]
- US-FDA, Food and Drug Administration. USA, Safety and Effectiveness of Health Care Antiseptics; Topical Antimicrobial Drug Products for Overthe-Counter Human Use; Final rule; US-FDA, Food and Drug Administration: Silver Spring, MD, USA, 2017. [Google Scholar]
- Dhillon, G.-S.; Kaur, S.; Pulicharla, R.; Brar, S.-K.; Cledón, M.; Verma, M. Triclosan: Current status occurrence environmental risks and bioaccumulation potential. Int. J. Environ. Res. Public. Health 2015, 12, 5657–5684. [Google Scholar] [CrossRef]
- Ramires, P.-F.; Tavella, R.-A.; Escarrone, A.-L.; Volcão, L.-M.; Honscha, L.-C. Ecotoxicity of triclosan in soil: An approach using different species. Environ. Sci. Pollut. Res. 2021, 28, 41233–41241. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-J.; Zhang, T.-L.; Zhu, P.; Cai, H.-T.; Jin, Y.; Gao, K.-G.; Li, J. Fabrication of Ag3PO4/polyaniline-activated biochar photocatalyst for efficient triclosan degradation process and toxicity assessment. Sci. Total Environ. 2022, 821, 153453. [Google Scholar] [CrossRef]
- Kookana, R.-S.; Shareef, A.; Fernandes, M.-B.; Hoare, S.; Gaylard, S.; Kumar, A. Bioconcentration of triclosan and methyl-triclosan in marine mussels (Mytilus gallo-provincialis) under laboratory conditions and in metropolitan waters of Gulf St Vin-cent South Australia. Mar. Pollut. Bull. 2013, 74, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Solá-Gutiérrez, C.; Schröder, S.; San-Román, M.-F.; Ortiz, I. Critical review on the mechanistic photolytic and photocatalytic degradation of triclosan. J. Environ. Manag. 2020, 260, 110101. [Google Scholar] [CrossRef] [PubMed]
- Schebb, N.-H.; Flores, I.; Kurobe, T.; Franze, B.; Ranganathan, A.; Hammock, B.-D. Bioconcentration metabolism and excretion of triclocarban in larval Qurt medaka (Oryzias latipes). Aquat. Toxicol. 2011, 105, 448–454. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, T.; Cheng, P.; Bai, Q.; Zhou, C.; Wang, W. Effects of triclosan and triclocarban on the growth inhibition cell viability genotoxicity and multixenobiotic resistance responses of Tetrahymena thermophila. Chemosphere 2015, 139, 434–440. [Google Scholar] [CrossRef]
- Martínez-Paz, P. Response of detoxification system genes on Chironomus riparius aquatic larvae after antibacterial agent triclosan exposures. Sci. Total Environ. 2018, 624, 1–8. [Google Scholar] [CrossRef]
- Rowett, C.-J.; Hutchinson, T.-H.; Comber, S.-D.-W. The impact of natural and anthropogenic dissolved organic carbon (DOC) and pH on the toxicity of triclosan to the crustacean Gammarus pulex (L.). Sci. Total Environ. 2016, 565, 222–231. [Google Scholar] [CrossRef]
- Dar, O.I.; Sharma, S.; Singh, K.; Sharma, A.; Bhardwaj, R.; Kaur, A. Biochemical markers for prolongation of the acute stress of triclosan in the early life stages of four food fishes. Chemosphere 2020, 247, 125914. [Google Scholar] [CrossRef]
- Paul, T.; Kumar, S.; Shukla, S.-P.; Pal, P.; Kumar, K.; Poojary, N. A multibiomarker approach using integrated biomarker response to assess the effect of pH on triclosan toxicity in Pangasianodon hypophthalmus (Sauvage 1878). Environ. Pollut. 2020, 260, 114001. [Google Scholar] [CrossRef]
- Martins, D.; Monteiro, M.-S.; Soares, A.-M.-V.-M.; Quintaneiro, C. Effects of 4-MBC and triclosan in embryos of the frog Pelophylax perezi. Chemosphere 2017, 178, 325–332. [Google Scholar] [CrossRef]
- Sager, E.; Scarcia, P.; Marino, D.; Mac, L.-T.; Rossi, A. Oxidative stress responses after exposure to triclosan sublethal concentrations: An integrated biomarker approach with a native (Corydoras paleatus) and a model fish species (Danio rerio). J. Toxicol. Environ. Health 2021, 85, 291–306. [Google Scholar] [CrossRef]
- Machado, M.-D.; Soares, E.-V. Sensitivity of freshwater and marine green algae to three compounds of emerging concern. J. Appl. Phycol. 2019, 31, 399–408. [Google Scholar] [CrossRef]
- Pan, C.-G.; Peng, F.-J.; Shi, W.-J.; Hu, L.-X.; Wei, X.-D.; Ying, G.-G. Triclosan-induced transcriptional and biochemical alterations in the freshwater green algae Chlamydomonas reinhardtii. Ecotoxicol. Environ. Saf. 2018, 148, 393–401. [Google Scholar] [CrossRef]
- Gonzalez, P.-M.; Rioboo, C.; Reguera, M.; Abreu, I.; Leganes, F.; Cid, A. Calcium mediates the cellular response of Chlamydomonas reinhardtii to the emerging aquatic pollutant triclosan. Aquat. Toxicol. 2017, 186, 50–66. [Google Scholar] [CrossRef]
- Xin, X.; Huang, G.; An, C.; Raina-Fulton, R.; Weger, H. Insights into long-term toxicity of triclosan to freshwater green algae in Lake Erie. Environ. Sci. Technol. 2019, 53, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zou, Y.; Zhang, F.-R.; Zhang, S.-H.; Huang, X.-Y.; Benoit, G. Growth ROS accumulation site and photosynthesis inhibition mechanism of Chlorella vulgaris by triclosan. Environ. Sci. Pollut. Res. 2022, 30, 12125–12137. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.-D.; Wei, L.-Y.; Hou, Z.-M.; Lin, S.-Q.; Li, J.-Y. Biological responses of alga Euglena gracilis to triclosan and galaxolide and the regulation of humic acid. Chemosphere 2022, 307, P1135667. [Google Scholar] [CrossRef]
- Fernando, P.-S.; Silvia, D.; Vanessa, P.; Angeles, A.; Sanna, O. Basis of genetic adaptation to heavy metal stress in the acidophilic green alga Chlamydomonas acidophila. Aquat. Toxicol. 2018, 200, 62–72. [Google Scholar]
- Wang, W.-J.; Sheng, Y.-Q. Effects and mechanisms of decabromodiphenyl ethane on Chlorella sorokiniana: Transcriptomics proteins and fatty acid production. Mar. Environ. Res. 2022, 181, 105764. [Google Scholar] [CrossRef]
- Gao, L.; Xie, Y.; Su, Y.-Y.; Mehmood, T.; Bao, R.-Q.; Fan, H.-J. Elucidating the negatively influential and potentially toxic mechanism of single and combined micro-sized polyethylene and petroleum to Chlorella vulgaris at the cellular and molecular levels. Ecotoxicol. Environ. Saf. 2022, 245, 245114102. [Google Scholar] [CrossRef]
- Sharma, S.; Dar, O.-I.; Singh, K.; Kaur, A.; Faggio, C. Triclosan elicited biochemical and transcriptomic alterations in Labeo rohita larvae. Environ. Toxicol. Pharmacol. 2021, 88, 103748. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Dar, O.-I.; Singh, K.; Thakur, S.; Kesavan, A.-K.; Kaur, A. Genomic markers for the biological responses of Triclosan stressed hatchlings of Labeo rohita. Environ. Sci. Pollut. Res. 2021, 28, 67370–67384. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.-Z.; Qi, Q.-J.; Hao, Y.-G.; Lei, Y.; Guo, J.-H. Transcriptional response of a green alga (Raphidocelis subcapitata) exposed to triclosan: Photosynthetic systems and DNA repair. J. Environ. Sci. 2022, 111, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, J.; Perez, E.; Van, V.-M.; Bertrand, A.-R.; Lupo, V.; Cardol, P. De Novo transcriptome meta-assembly of the mixotrophic freshwater microalga Euglena gracilis. Genes 2021, 12, 842. [Google Scholar] [CrossRef]
- Zakryś, B.; Milanowski, R.; Karnkowska, A. Evolutionary origin of Euglena. Adv. Exp. Med. Biol. 2017, 979. [Google Scholar] [CrossRef]
- Yu, S.-P.; Cole, M.; Chan, B. Review: Effects of microplastic on zooplankton survival and sublethal responses. Oceanogr. Mar. Biol. 2020, 58, 351–394. [Google Scholar]
- He, J.-Y.; Liu, C.-C.; Du, M.-Z.; Zhou, X.-Y.; Hu, Z.-L.; Lei, A. Metabolic responses of a model green microalga Euglena gracilis to different environmental stresses. Front. Bioeng. Biotechnol. 2021, 9, 662655. [Google Scholar] [CrossRef]
- Li, M.; Gao, X.-Y.; Wu, B.; Qian, X.; Giesy, J.-P.; Cui, Y.-B. Microalga Euglena as a bioindicator for testing genotoxic potentials of organic pollutants in Taihu Lake China. Ecotoxicology 2014, 23, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-C.; Jiang, X.-F.; Xiao, Y. Exposure of microalgae Euglena gracilis to polystyrene microbeads and cadmium: Perspective from the physiological and transcriptional responses. Aquat. Toxicol. 2020, 228, 105650. [Google Scholar] [CrossRef]
- Deng, X.-Y.; Cheng, J.; Hu, X.-L.; Wang, L.; Li, D.; Gao, K. Biological effects of TiO2 and CeO2 nanoparticles on the growth photosynthetic activity and cellular components of a marine diatom Phaeodactylum tricornutum. Sci. Total Environ. 2017, 575, 87–96. [Google Scholar] [CrossRef]
- Pertea, G.; Huang, X.-Q.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.-F.; Gish, W.; Miller, W.; Myers, E.-W.; Lipman, D.-J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Prado, R.; Rioboo, C.; Herrero, C. The herbicide paraquat induces alterations in the elemental and biochemical composition of nontarget microalgal species. Chemosphere 2009, 76, 1440–1444. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-M.; Seppänen-Laakso, T.; Rischer, H.; Wiebe, M.a.-G. Euglena gracilis growth and cell composition under different temperature, light and trophic conditions. PLoS ONE 2018, 13, e0195329. [Google Scholar] [CrossRef] [PubMed]
- Azizullah, A.; Richter, P.; Haeder, D.-P. Comparative toxicity of the pesticides carbofuran and malathion to the freshwater flagellate Euglena gracilis. Ecotoxicology 2011, 20, 1442–1454. [Google Scholar] [CrossRef]
- Mao, Y.; Ai, H.; Chen, Y. Phytoplankton response to polystyrene microplastics: Perspective from an entire growth period. Chemosphere 2018, 208, 59–68. [Google Scholar] [CrossRef]
- Xin, X.; Huang, G.; An, C.; Feng, R. Interactive toxicity of triclosan and nanoTiO2 to green alga Eremosphaera viridis in Lake Erie: A new perspective based on Fourier Transform Infrared spectromicroscopy and synchrotron-based X-ray fluorescence imaging. Environ. Sci. Technol. 2019, 53, 9884–9894. [Google Scholar] [CrossRef]
- Kim, H.; Wang, H.; Ki, J.-S. Chloroacetanilides inhibit photosynthesis and disrupt the thylakoid membranes of the dinoflagellate Prorocentrum minimum as revealed with metazachlor treatment. Ecotoxicol. Environ. Saf. 2021, 211, 111928. [Google Scholar] [CrossRef]
- Peng, D.-L.; Wang, W.-J.; Liu, A.-R.; Zhang, Y.; Li, X.-Z.; Wang, G. Comparative transcriptome combined with transgenic analysis reveal the involvement of salicylic acid pathway in the response of Nicotiana tabacum to triclosan stress. Chemosphere 2021, 270, 129456. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.-P. Antioxidant responses of wheat plants under stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef]
- VanEtten, S.-L.; Bonner, M.-R.; Ren, X.-F.; Birnbaum, L.-S.; Kostyniak, P.-J.; Wang, J.; Olson, J.-R. Effect of Exposure to 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) and Polychlorinated Biphenyls (PCBs) on Mitochondrial DNA (mtDNA) Copy Number in Rats. Toxicology 2021, 454, 152744. [Google Scholar] [CrossRef] [PubMed]
- Peinado, F.-M.; Artacho-Cordón, F.; Barrios-Rodríguez, R.; Arrebola, J.-P. Influence of polychlorinated biphenyls and organochlorine pesticides on the inflammatory milieu. A systematic review of in vitro, in vivo and epidemiological studies. Environ. Res. 2020, 186, 109561. [Google Scholar] [CrossRef]
- Du, C.-L.; Zhang, B.; He, Y.-L.; Hu, C.-Y. Biological effect of aqueous C 60 aggregates on Scenedesmus obliquus revealed by transcriptomics and non-targeted metabolomics. J. Hazard. Mater. 2017, 324, 221–229. [Google Scholar] [CrossRef]
- Matthijs, M.; Fabris, M.; Obata, T.; Foubert, I. The transcription factor bZIP14 regulates the TCA cycle in the diatom Phaeodactylum tricornutum. EMBO J. 2017, 36, 1559–1576. [Google Scholar] [CrossRef]
- Jin, Y.-X.; Zheng, S.-S.; Fu, Z.-W. Embryonic exposure to cypermethrin induces apoptosis and immunotoxicity in zebrafish (Danio rerio). Fish. Shellfish. Immunol. 2011, 30, 1049–1054. [Google Scholar] [CrossRef]
- Chae, Y.; Kim, D.; Kim, S.-W.; An, Y. Trophic transfer and individual impact of nano-sized polystyrene in a four-species freshwater food chain. Sci. Rep. 2018, 8, 284. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, T.; Zhang, T.; Yang, W.; Yang, B.; Cao, J.; Yang, Y.; Li, M. Molecular Toxicity Mechanism Induced by the Antibacterial Agent Triclosan in Freshwater Euglena gracilis Based on the Transcriptome. Toxics 2023, 11, 414. https://doi.org/10.3390/toxics11050414
Lu T, Zhang T, Yang W, Yang B, Cao J, Yang Y, Li M. Molecular Toxicity Mechanism Induced by the Antibacterial Agent Triclosan in Freshwater Euglena gracilis Based on the Transcriptome. Toxics. 2023; 11(5):414. https://doi.org/10.3390/toxics11050414
Chicago/Turabian StyleLu, Ting, Tong Zhang, Weishu Yang, Bin Yang, Jing Cao, Yang Yang, and Mei Li. 2023. "Molecular Toxicity Mechanism Induced by the Antibacterial Agent Triclosan in Freshwater Euglena gracilis Based on the Transcriptome" Toxics 11, no. 5: 414. https://doi.org/10.3390/toxics11050414
APA StyleLu, T., Zhang, T., Yang, W., Yang, B., Cao, J., Yang, Y., & Li, M. (2023). Molecular Toxicity Mechanism Induced by the Antibacterial Agent Triclosan in Freshwater Euglena gracilis Based on the Transcriptome. Toxics, 11(5), 414. https://doi.org/10.3390/toxics11050414