Nano-Curcumin Prevents Copper Reproductive Toxicity by Attenuating Oxidative Stress and Inflammation and Improving Nrf2/HO-1 Signaling and Pituitary-Gonadal Axis in Male Rats

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Experimental Animals and Treatments
2.3. Assay of Testosterone, Gonadotropins, and TNF-α
2.4. Assay of MDA, NO and Antioxidants
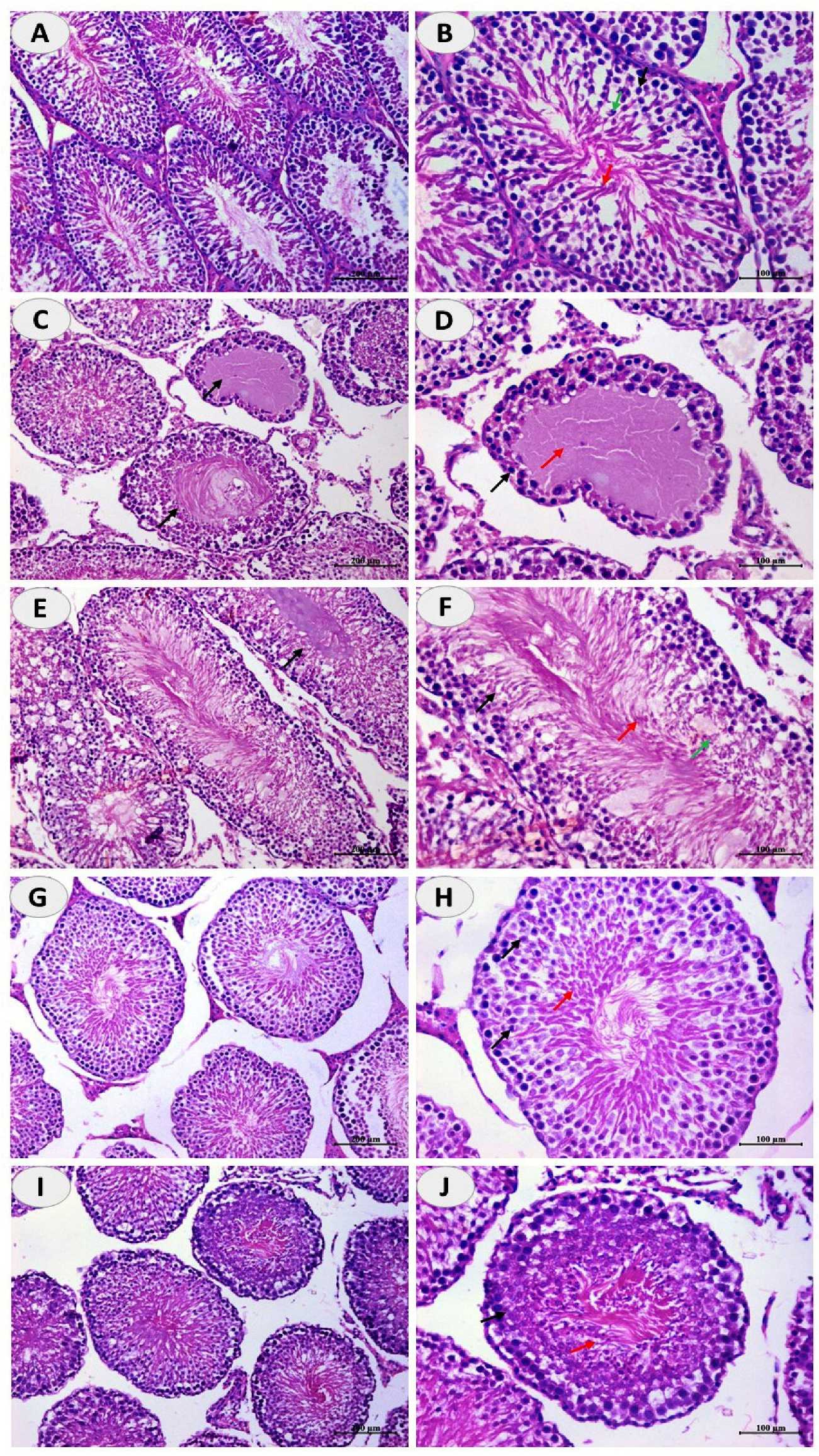
2.5. Histopathological Examination
2.6. Gene Expression
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. N-CUR and CUR Enhance the Pituitary-Gonadal Axis and Prevent Testicular Injury in CuSO4-Administered Rats
3.2. N-CUR and CUR Upregulate Steroidogenesis and AR in CuSO4-Administered Rats
3.3. N-CUR and CUR Attenuate Testicular Oxidative Stress in CuSO4-Administered Rats
3.4. N-CUR and CUR Upregulate Nrf2/HO-1 Signaling in CuSO4-Administered Rats
3.5. N-CUR and CUR Mitigate Testicular Inflammation in CuSO4-Administered Rats
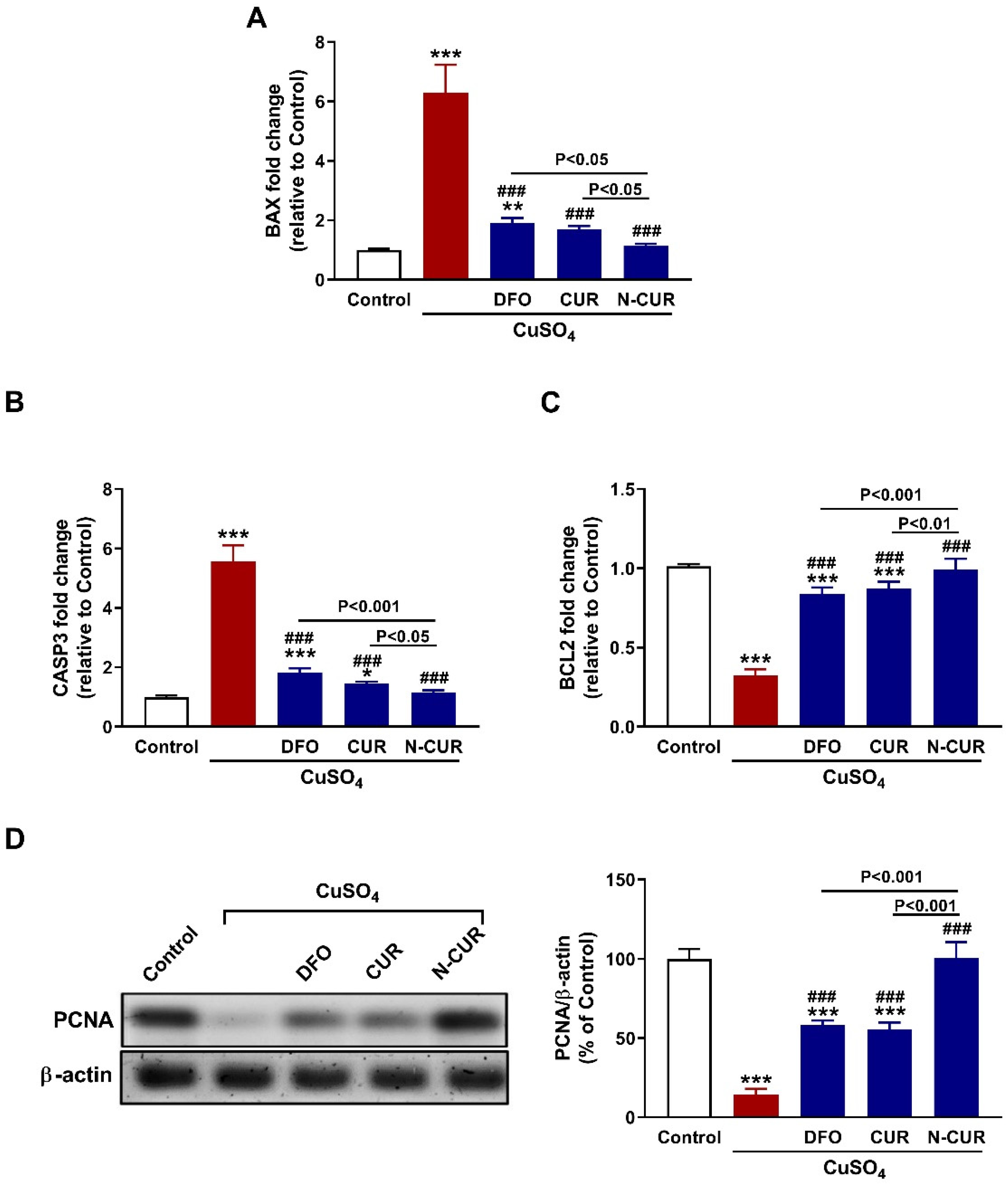
3.6. N-CUR and CUR Prevent Testicular Apoptosis in CuSO4-Administered Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheiber, I.F.; Mercer, J.F.; Dringen, R. Metabolism and functions of copper in brain. Prog. Neurobiol. 2014, 116, 33–57. [Google Scholar] [CrossRef]
- Uriu-Adams, J.Y.; Keen, C.L. Copper, oxidative stress, and human health. Mol. Asp. Med. 2005, 26, 268–298. [Google Scholar] [CrossRef]
- Bost, M.; Houdart, S.; Oberli, M.; Kalonji, E.; Huneau, J.-F.; Margaritis, I. Dietary copper and human health: Current evidence and unresolved issues. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. (GMS) 2016, 35, 107–115. [Google Scholar] [CrossRef]
- Denoyer, D.; Masaldan, S.; La Fontaine, S.; Cater, M.A. Targeting copper in cancer therapy: ‘Copper that cancer’. Met. Integr. Biometal. Sci. 2015, 7, 1459–1476. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Nath, S.; Massanyi, P.; Stawarz, R.; Kačániová, M.; Kolesarova, A. Copper-Induced Changes in Reproductive Functions: In Vivo and In Vitro Effects. Physiol. Res. 2016, 65, 11–22. [Google Scholar] [CrossRef]
- Zhang, L.; Yuan, Z.; Bi, J. Estimation of Copper In-use Stocks in Nanjing, China. J. Ind. Ecol. 2012, 16, 191–202. [Google Scholar] [CrossRef]
- Committee on Copper in Drinking. Copper in Drinking Water; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Lamtai, M.; Zghari, O.; Ouakki, S.; Marmouzi, I.; Mesfioui, A.; El Hessni, A.; Ouichou, A. Chronic copper exposure leads to hippocampus oxidative stress and impaired learning and memory in male and female rats. Toxicol. Res. 2020, 36, 359–366. [Google Scholar] [CrossRef]
- Brewer, G.J. Alzheimer’s disease causation by copper toxicity and treatment with zinc. Front. Aging Neurosci. 2014, 6, 92. [Google Scholar] [CrossRef]
- Montes, S.; Rivera-Mancia, S.; Diaz-Ruiz, A.; Tristan-Lopez, L.; Rios, C. Copper and Copper Proteins in Parkinson’s Disease. Oxidative Med. Cell. Longev. 2014, 2014, 147251. [Google Scholar] [CrossRef]
- Zietz, B.; De Vergara, J.D.; Schneider, H.; Kessler, B.; Dunkelberg, H. Prevalence of elevated copper concentrations in tap water in two areas of Germany used for infant feeding and possible health implications. Eur. J. Med. Res. 1999, 4, 298. [Google Scholar]
- Hsiao, C.-D.; Wu, H.-H.; Malhotra, N.; Liu, Y.-C.; Wu, Y.-H.; Lin, Y.-N.; Saputra, F.; Santoso, F.; Chen, K. Expression and Purification of Recombinant GHK Tripeptides Are Able to Protect against Acute Cardiotoxicity from Exposure to Waterborne-Copper in Zebrafish. Biomolecules 2020, 10, 1202. [Google Scholar] [CrossRef]
- Hashish, E.A.; Elgaml, S.A. Hepatoprotective and Nephroprotective Effect of Curcumin Against Copper Toxicity in Rats. Indian J. Clin. Biochem. 2016, 31, 270–277. [Google Scholar] [CrossRef]
- Kumar, V.; Kalita, J.; Misra, U.; Bora, H. A study of dose response and organ susceptibility of copper toxicity in a rat model. J. Trace Elem. Med. Biol. 2015, 29, 269–274. [Google Scholar] [CrossRef]
- Sarawi, W.S.; Alhusaini, A.M.; Fadda, L.M.; Alomar, H.A.; Albaker, A.B.; Aljrboa, A.S.; Alotaibi, A.M.; Hasan, I.H.; Mahmoud, A.M. Curcumin and Nano-Curcumin Mitigate Copper Neurotoxicity by Modulating Oxidative Stress, Inflammation, and Akt/GSK-3β Signaling. Molecules 2021, 26, 5591. [Google Scholar] [CrossRef]
- Sarawi, W.S.; Alhusaini, A.M.; Fadda, L.M.; Alomar, H.A.; Albaker, A.B.; Aljrboa, A.S.; Alotaibi, A.M.; Hasan, I.H.; Mahmoud, A.M. Nano-curcumin prevents cardiac injury, oxidative stress and inflammation, and modulates TLR4/NF-κB and MAPK signaling in copper sulfate-intoxicated rats. Antioxidants 2021, 10, 1414. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, X.; Li, X.; Ru, S.; Wang, Y.; Yin, J.; Liu, D. Oxidative damage induced by copper in testis of the red swamp crayfish Procambarus clarkii and its underlying mechanisms. Aquat. Toxicol. 2019, 207, 120–131. [Google Scholar] [CrossRef]
- Lin, S.; Qiao, N.; Chen, H.; Tang, Z.; Han, Q.; Mehmood, K.; Fazlani, S.A.; Hameed, S.; Li, Y.; Zhang, H. Integration of transcriptomic and metabolomic data reveals metabolic pathway alteration in mouse spermatogonia with the effect of copper exposure. Chemosphere 2020, 256, 126974. [Google Scholar] [CrossRef]
- Khushboo, M.; Murthy, M.K.; Devi, M.S.; Sanjeev, S.; Ibrahim, K.S.; Kumar, N.S.; Roy, V.K.; Gurusubramanian, G. Testicular toxicity and sperm quality following copper exposure in Wistar albino rats: Ameliorative potentials of L-carnitine. Environ. Sci. Pollut. Res. 2018, 25, 1837–1862. [Google Scholar] [CrossRef]
- Chen, H.; Kang, Z.; Qiao, N.; Liu, G.; Huang, K.; Wang, X.; Pang, C.; Zeng, Q.; Tang, Z.; Li, Y. Chronic Copper Exposure Induces Hypospermatogenesis in Mice by Increasing Apoptosis Without Affecting Testosterone Secretion. Biol. Trace Elem. Res. 2020, 195, 472–480. [Google Scholar] [CrossRef]
- Sakhaee, E.; Emadi, L.; Abshenas, J.; Kheirandish, R.; Azari, O.; Amiri, E. Evaluation of epididymal sperm quality following experimentally induced copper poisoning in male rats. Andrologia 2012, 44, 110–116. [Google Scholar] [CrossRef]
- Babaei, H.; Roshangar, L.; Sakhaee, E.; Abshenas, J.; Kheirandish, R.; Dehghani, R. Ultrastructural and morphometrical changes of mice ovaries following experimentally induced copper poisoning. Iran. Red Crescent Med. J. 2012, 14, 558–568. [Google Scholar]
- Chattopadhyay, A.; Sarkar, M.; Sengupta, R.; Roychowdhury, G.; Biswas, N.M. Antitesticular effect of copper chloride in albino rats. J. Toxicol. Sci. 1999, 24, 393–397. [Google Scholar] [CrossRef][Green Version]
- Li, Y.; Chen, H.; Liao, J.; Chen, K.; Javed, M.T.; Qiao, N.; Zeng, Q.; Liu, B.; Yi, J.; Tang, Z.; et al. Long-term copper exposure promotes apoptosis and autophagy by inducing oxidative stress in pig testis. Environ. Sci. Pollut. Res. 2021, 28, 55140–55153. [Google Scholar] [CrossRef]
- Aitken, R.J.; Roman, S. Antioxidant Systems and Oxidative Stress in the Testes. Oxidative Med. Cell. Longev. 2008, 1, 616821. [Google Scholar] [CrossRef]
- Saghir, S.A.M.; Alharbi, S.A.; Al-Garadi, M.A.; Al-Gabri, N.; Rady, H.Y.; Olama, N.K.; Abdulghani, M.A.M.; Al Hroob, A.M.; Almaiman, A.A.; Bin-Jumah, M.; et al. Curcumin Prevents Cyclophosphamide-Induced Lung Injury in Rats by Suppressing Oxidative Stress and Apoptosis. Processes 2020, 8, 127. [Google Scholar] [CrossRef]
- Pizzo, P.; Scapin, C.; Vitadello, M.; Florean, C.; Gorza, L. Grp94 acts as a mediator of curcumin-induced antioxidant defence in myogenic cells. J. Cell. Mol. Med. 2010, 14, 970–981. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Huebbe, P.; Ernst, I.M.A.; Chin, D.; Wagner, A.E.; Rimbach, G. Curcumin-From Molecule to Biological Function. Angew. Chem. Int. Ed. 2012, 51, 5308–5332. [Google Scholar] [CrossRef]
- De, R.; Kundu, P.; Swarnakar, S.; Ramamurthy, T.; Chowdhury, A.; Nair, G.B.; Mukhopadhyay, A.K. Antimicrobial Activity of Curcumin against Helicobacter pylori Isolates from India and during Infections in Mice. Antimicrob. Agents Chemother. 2009, 53, 1592–1597. [Google Scholar] [CrossRef]
- Tsuda, T. Curcumin as a functional food-derived factor: Degradation products, metabolites, bioactivity, and future perspectives. Food Funct. 2018, 9, 705–714. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Ahmed, O.M.; Galaly, S.R. Thymoquinone and curcumin attenuate gentamicin-induced renal oxidative stress, inflammation and apoptosis in rats. EXCLI J. 2014, 13, 98–110. [Google Scholar]
- Alhusaini, A.; Fadda, L.; Hasan, I.H.; Zakaria, E.; Alenazi, A.M.; Mahmoud, A.M. Curcumin Ameliorates Lead-Induced Hepatotoxicity by Suppressing Oxidative Stress and Inflammation, and Modulating Akt/GSK-3β Signaling Pathway. Biomolecules 2019, 9, 703. [Google Scholar] [CrossRef]
- Al-Dossari, M.H.; Fadda, L.M.; Attia, H.A.; Hasan, I.H.; Mahmoud, A.M. Curcumin and Selenium Prevent Lipopolysaccharide/Diclofenac-Induced Liver Injury by Suppressing Inflammation and Oxidative Stress. Biol. Trace Elem. Res. 2019, 196, 173–183. [Google Scholar] [CrossRef]
- Galaly, S.R.; Ahmed, O.M.; Mahmoud, A.M. Thymoquinone and curcumin prevent gentamicin-induced liver injury by attenuating oxidative stress, inflammation and apoptosis. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2014, 65, 823–832. [Google Scholar]
- Damiano, S.; Longobardi, C.; Andretta, E.; Prisco, F.; Piegari, G.; Squillacioti, C.; Montagnaro, S.; Pagnini, F.; Badino, P.; Florio, S.; et al. Antioxidative Effects of Curcumin on the Hepatotoxicity Induced by Ochratoxin A in Rats. Antioxidants 2021, 10, 125. [Google Scholar] [CrossRef]
- Longobardi, C.; Damiano, S.; Andretta, E.; Prisco, F.; Russo, V.; Pagnini, F.; Florio, S.; Ciarcia, R. Curcumin Modulates Nitrosative Stress, Inflammation, and DNA Damage and Protects against Ochratoxin A-Induced Hepatotoxicity and Nephrotoxicity in Rats. Antioxidants 2021, 10, 1239. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Harikumar, K.B. Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases. Int. J. Biochem. Cell Biol. 2009, 41, 40–59. [Google Scholar] [CrossRef]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef]
- Mirzaei, H.; Masoudifar, A.; Sahebkar, A.; Zare, N.; Sadri Nahand, J.; Rashidi, B.; Mehrabian, E.; Mohammadi, M.; Mirzaei, H.R.; Jaafari, M.R. MicroRNA: A novel target of curcumin in cancer therapy. J. Cell. Physiol. 2018, 233, 3004–3015. [Google Scholar] [CrossRef]
- Carlos-Reyes, Á.; López-González, J.S.; Meneses-Flores, M.; Gallardo-Rincón, D.; Ruíz-García, E.; Marchat, L.; La Vega, H.A.-D.; De La Cruz, O.N.H.; López-Camarillo, C. Dietary Compounds as Epigenetic Modulating Agents in Cancer. Front. Genet. 2019, 10, 79. [Google Scholar] [CrossRef]
- Karimi, S.; Khorsandi, L.; Nejaddehbashi, F. Protective effects of Curcumin on testicular toxicity induced by titanium dioxide nanoparticles in mice. JBRA Assist. Reprod. 2019, 23, 344–351. [Google Scholar] [CrossRef]
- Sudjarwo, S.A.; Sudjarwo, G.W. Koerniasari Protective effect of curcumin on lead acetate-induced testicular toxicity in Wistar rats. Res. Pharm. Sci. 2017, 12, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Huyut, Z.; Alp, H.H.; Yaman, T.; Keleş, Ö.F.; Yener, Z.; Türkan, F.; Ayengin, K. Comparison of the protective effects of curcumin and caffeic acid phenethyl ester against doxorubicin-induced testicular toxicity. Andrologia 2020, 53, e13919. [Google Scholar] [CrossRef] [PubMed]
- Flora, G.; Gupta, D.; Tiwari, A. Nanocurcumin: A Promising Therapeutic Advancement over Native Curcumin. Crit. Rev. Ther. Drug Carrier Syst. 2013, 30, 331–368. [Google Scholar] [CrossRef] [PubMed]
- Gera, M.; Sharma, N.; Ghosh, M.; Huynh, D.L.; Lee, S.J.; Min, T.; Kwon, T.; Jeong, D.K. Nanoformulations of curcumin: An emerging paradigm for improved remedial application. Oncotarget 2017, 8, 66680–66698. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Wei, Y.; Lee, R.J.; Zhao, L. Liposomal curcumin and its application in cancer. Int. J. Nanomed. 2017, 12, 6027–6044. [Google Scholar] [CrossRef]
- Nisini, R.; Poerio, N.; Mariotti, S.; De Santis, F.; Fraziano, M. The Multirole of Liposomes in Therapy and Prevention of Infectious Diseases. Front. Immunol. 2018, 9, 155. [Google Scholar] [CrossRef]
- Satta, S.; Mahmoud, A.M.; Wilkinson, F.L.; Alexander, M.Y.; White, S.J. The role of Nrf2 in cardiovascular function and disease. Oxid. Med. Cell. Longev. 2017, 2017, 9237263. [Google Scholar] [CrossRef]
- Alhusaini, A.; Fadda, L.; Hassan, I.; Ali, H.M.; Alsaadan, N.; Aldowsari, N.; Aldosari, A.; Alharbi, B. Liposomal curcumin attenuates the incidence of oxidative stress, inflammation, and DNA damage induced by copper sulfate in rat liver. Dose Response A Publ. Int. Hormesis Soc. 2018, 16, 1559325818790869. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Grisham, M.B.; Johnson, G.G.; Lancaster, J.R., Jr. Quantitation of nitrate and nitrite in extracellular fluids. Methods Enzymol. 1996, 268, 237–246. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Marklund, S.L. Superoxide dismutase isoenzymes in tissues and plasma from New Zealand black mice, nude mice and normal BALB/c mice. Mutat. Res. 1985, 148, 129–134. [Google Scholar] [CrossRef]
- Cohen, G.; Dembiec, D.; Marcus, J. Measurement of catalase activity in tissue extracts. Anal. Biochem. 1970, 34, 30–38. [Google Scholar] [CrossRef]
- Alotaibi, M.F.; Al-Joufi, F.; Seif, H.S.A.; Alzoghaibi, M.A.; Djouhri, L.; Ahmeda, A.F.; Mahmoud, A.M. Umbelliferone Inhibits Spermatogenic Defects and Testicular Injury in Lead-Intoxicated Rats by Suppressing Oxidative Stress and Inflammation, and Improving Nrf2/HO-1 Signaling. Drug Des. Dev. Ther. 2020, 14, 4003–4019. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vázquez-Blanco, R.; Arias-Estévez, M.; Bååth, E.; Fernández-Calviño, D. Comparison of Cu salts and commercial Cu based fungicides on toxicity towards microorganisms in soil. Environ. Pollut. 2020, 257, 113585. [Google Scholar] [CrossRef]
- Oduwole, O.O.; Peltoketo, H.; Huhtaniemi, I.T. Role of Follicle-Stimulating Hormone in Spermatogenesis. Front. Endocrinol. 2018, 9, 763. [Google Scholar] [CrossRef]
- Miller, W.L.; Bose, H.S. Early steps in steroidogenesis: Intracellular cholesterol trafficking. J. Lipid Res. 2011, 52, 2111–2135. [Google Scholar] [CrossRef] [PubMed]
- Caron, K.M.; Soo, S.-C.; Wetsel, W.C.; Stocco, D.M.; Clark, B.J.; Parker, K.L. Targeted disruption of the mouse gene encoding steroidogenic acute regulatory protein provides insights into congenital lipoid adrenal hyperplasia. Proc. Natl. Acad. Sci. USA 1997, 94, 11540–11545. [Google Scholar] [CrossRef]
- Bremer, A.A.; Miller, W.L. Chapter 13–Regulation of steroidogenesis. In Cellular Endocrinology in Health and Disease; Ulloa-Aguirre, A., Conn, P.M., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 207–227. [Google Scholar]
- Dušková, M.; Pospíšilová, H. The Role of Non-Aromatizable Testosterone Metabolite in Metabolic Pathways. Physiol. Res. 2011, 60, 253–261. [Google Scholar] [CrossRef]
- Wang, R.-S.; Yeh, S.; Tzeng, C.-R.; Chang, C. Androgen Receptor Roles in Spermatogenesis and Fertility: Lessons from Testicular Cell-Specific Androgen Receptor Knockout Mice. Endocr. Rev. 2009, 30, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, K. Quantum chemical insights into alzheimer’s disease: Curcumin’s chelation with Cu(ii), Zn(ii), and Pd(ii) as a mechanism for its prevention. Int. J. Quantum Chem. 2016, 116, 1107–1119. [Google Scholar] [CrossRef]
- Gunther, M.R.; Hanna, P.; Mason, R.P.; Cohen, M. Hydroxyl Radical Formation from Cuprous Ion and Hydrogen Peroxide: A Spin-Trapping Study. Arch. Biochem. Biophys. 1995, 316, 515–522. [Google Scholar] [CrossRef]
- Gaetke, L.M.; Chow-Johnson, H.S.; Chow, C.K. Copper: Toxicological relevance and mechanisms. Arch. Toxicol. 2014, 88, 1929–1938. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Sayed, A.M.; Ahmed, O.S.; Abdel-Daim, M.M.; Hassanein, E.H.M. The role of flavonoids in inhibiting IL-6 and inflammatory arthritis. Curr. Top. Med. Chem. 2022, 22, 746–768. [Google Scholar] [CrossRef]
- Yang, S.-H.; He, J.-B.; Yu, L.-H.; Li, L.; Long, M.; Liu, M.-D.; Li, P. Protective role of curcumin in cadmium-induced testicular injury in mice by attenuating oxidative stress via Nrf2/ARE pathway. Environ. Sci. Pollut. Res. 2019, 26, 34575–34583. [Google Scholar] [CrossRef]
- Shahcheraghi, S.H.; Salemi, F.; Peirovi, N.; Ayatollahi, J.; Alam, W.; Khan, H.; Saso, L. Nrf2 Regulation by Curcumin: Molecular Aspects for Therapeutic Prospects. Molecules 2021, 27, 167. [Google Scholar] [CrossRef]
- Bonizzi, G.; Karin, M. The two NF-κB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004, 25, 280–288. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, J.; Weng, C.; Chen, R.; Zheng, Y.; Chen, Q.; Tang, H. Identification of the protein–protein contact site and interaction mode of human VDAC1 with Bcl-2 family proteins. Biochem. Biophys. Res. Commun. 2003, 305, 989–996. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Wrobel, K.-H.; Bickel, D.; Kujat, R. Immunohistochemical study of seminiferous epithelium in adult bovine testis using monoclonal antibodies against Ki-67 protein and proliferating cell nuclear antigen (PCNA). Cell Tissue Res. 1996, 283, 191–201. [Google Scholar] [CrossRef]
- Chen, Z.; Wen, D.; Wang, F.; Wang, C.; Yang, L. Curcumin protects against palmitic acid-induced apoptosis via the inhibition of endoplasmic reticulum stress in testicular Leydig cells. Reprod. Biol. Endocrinol. 2019, 17, 71. [Google Scholar] [CrossRef]
- Kilinc, L.; Uz, Y.H. Protective effects of curcumin against methotrexate-induced testicular damage in rats by suppression of the p38-MAPK and nuclear factor-kappa B pathways. Clin. Exp. Reprod. Med. 2021, 48, 211–220. [Google Scholar] [CrossRef]
- Boyanapalli, S.S.S.; Paredes-Gonzalez, X.; Fuentes, F.; Zhang, C.; Guo, Y.; Pung, D.; Saw, C.L.L.; Kong, A.-N.T. Nrf2 Knockout Attenuates the Anti-Inflammatory Effects of Phenethyl Isothiocyanate and Curcumin. Chem. Res. Toxicol. 2014, 27, 2036–2043. [Google Scholar] [CrossRef]
- Yen, F.-L.; Wu, T.-H.; Tzeng, C.-W.; Lin, L.-T.; Lin, C.-C. Curcumin Nanoparticles Improve the Physicochemical Properties of Curcumin and Effectively Enhance Its Antioxidant and Antihepatoma Activities. J. Agric. Food Chem. 2010, 58, 7376–7382. [Google Scholar] [CrossRef]
- Nahar, P.P.; Slitt, A.L.; Seeram, N.P. Anti-Inflammatory Effects of Novel Standardized Solid Lipid Curcumin Formulations. J. Med. Food 2015, 18, 786–792. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession Number | Primers (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|
| StAR | NM_031558.3 | F: CCCAGATAGAGTTCGCCAGC R: TGGTGGGCAGTCCTTAACAC | 84 |
| CYP17A1 | XM_006231434.3 | F: TCAGTGACTGTGACCTGGGA R: GTGGAGCGGAGCAACTTCAA | 122 |
| 3β-HSD | NM_001007719.3 | F: TGTGCCAGCCTTCATCTAC R: CTTCTCGGCCATCCTTTT | 145 |
| 17β-HSD | NM_054007.1 | F: GACCGCCGATGAGTTTGT R: TTTGGGTGGTGCTGCTGT | 140 |
| NF-κB p65 | NM_199267.2 | F: CCTCATCTTTCCCTCAGAGCC R: GGTCCCGTGTAGCCATTGAT | 189 |
| iNOS | NM_012611.3 | F: ATTCCCAGCCCAACAACACA R: GCAGCTTGTCCAGGGATTCT | 112 |
| TNF-α | NM_012675.3 | F: AAATGGGCTCCCTCTCATCAGTTC R: TCTGCTTGGTGGTTTGCTACGAC | 111 |
| Caspase-3 | NM_012922.2 | F: GGAGCTTGGAACGCGAAGAA R: ACACAAGCCCATTTCAGGGT | 169 |
| BAX | NM_017059.2 | F: AGGACGCATCCACCAAGAAG R: CAGTTGAAGTTGCCGTCTGC | 166 |
| BCL-2 | NM_016993.1 | F: ACTCTTCAGGGATGGGGTGA R: TGACATCTCCCTGTTGACGC | 94 |
| β-actin | NM_031144.3 | F: AGGAGTACGATGAGTCCGGC R: CGCAGCTCAGTAACAGTCCG | 71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarawi, W.S.; Alhusaini, A.M.; Fadda, L.M.; Alomar, H.A.; Albaker, A.B.; Alghibiwi, H.K.; Aljrboa, A.S.; Alotaibi, A.M.; Hasan, I.H.; Mahmoud, A.M. Nano-Curcumin Prevents Copper Reproductive Toxicity by Attenuating Oxidative Stress and Inflammation and Improving Nrf2/HO-1 Signaling and Pituitary-Gonadal Axis in Male Rats. Toxics 2022, 10, 356. https://doi.org/10.3390/toxics10070356
Sarawi WS, Alhusaini AM, Fadda LM, Alomar HA, Albaker AB, Alghibiwi HK, Aljrboa AS, Alotaibi AM, Hasan IH, Mahmoud AM. Nano-Curcumin Prevents Copper Reproductive Toxicity by Attenuating Oxidative Stress and Inflammation and Improving Nrf2/HO-1 Signaling and Pituitary-Gonadal Axis in Male Rats. Toxics. 2022; 10(7):356. https://doi.org/10.3390/toxics10070356
Chicago/Turabian StyleSarawi, Wedad S., Ahlam M. Alhusaini, Laila M. Fadda, Hatun A. Alomar, Awatif B. Albaker, Hanan K. Alghibiwi, Amjad S. Aljrboa, Areej M. Alotaibi, Iman H. Hasan, and Ayman M. Mahmoud. 2022. "Nano-Curcumin Prevents Copper Reproductive Toxicity by Attenuating Oxidative Stress and Inflammation and Improving Nrf2/HO-1 Signaling and Pituitary-Gonadal Axis in Male Rats" Toxics 10, no. 7: 356. https://doi.org/10.3390/toxics10070356
APA StyleSarawi, W. S., Alhusaini, A. M., Fadda, L. M., Alomar, H. A., Albaker, A. B., Alghibiwi, H. K., Aljrboa, A. S., Alotaibi, A. M., Hasan, I. H., & Mahmoud, A. M. (2022). Nano-Curcumin Prevents Copper Reproductive Toxicity by Attenuating Oxidative Stress and Inflammation and Improving Nrf2/HO-1 Signaling and Pituitary-Gonadal Axis in Male Rats. Toxics, 10(7), 356. https://doi.org/10.3390/toxics10070356