
Transcriptomic Insights into the Enhanced Aroma of Guangdong Oolong Dry Tea (Camellia sinensis cv. Yashixiang Dancong) in Winter


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of the Volatile Components
2.3. GC-MS Analysis
2.4. GC-MS Data Analysis
2.5. Calculation of Odor Activity Value (OAV) and Terpene Index (TI)
2.6. RNA Extraction and Transcriptome Sequencing
2.7. Transcriptome Analysis
2.8. qRT-PCR
2.9. Statistical Analysis
3. Results
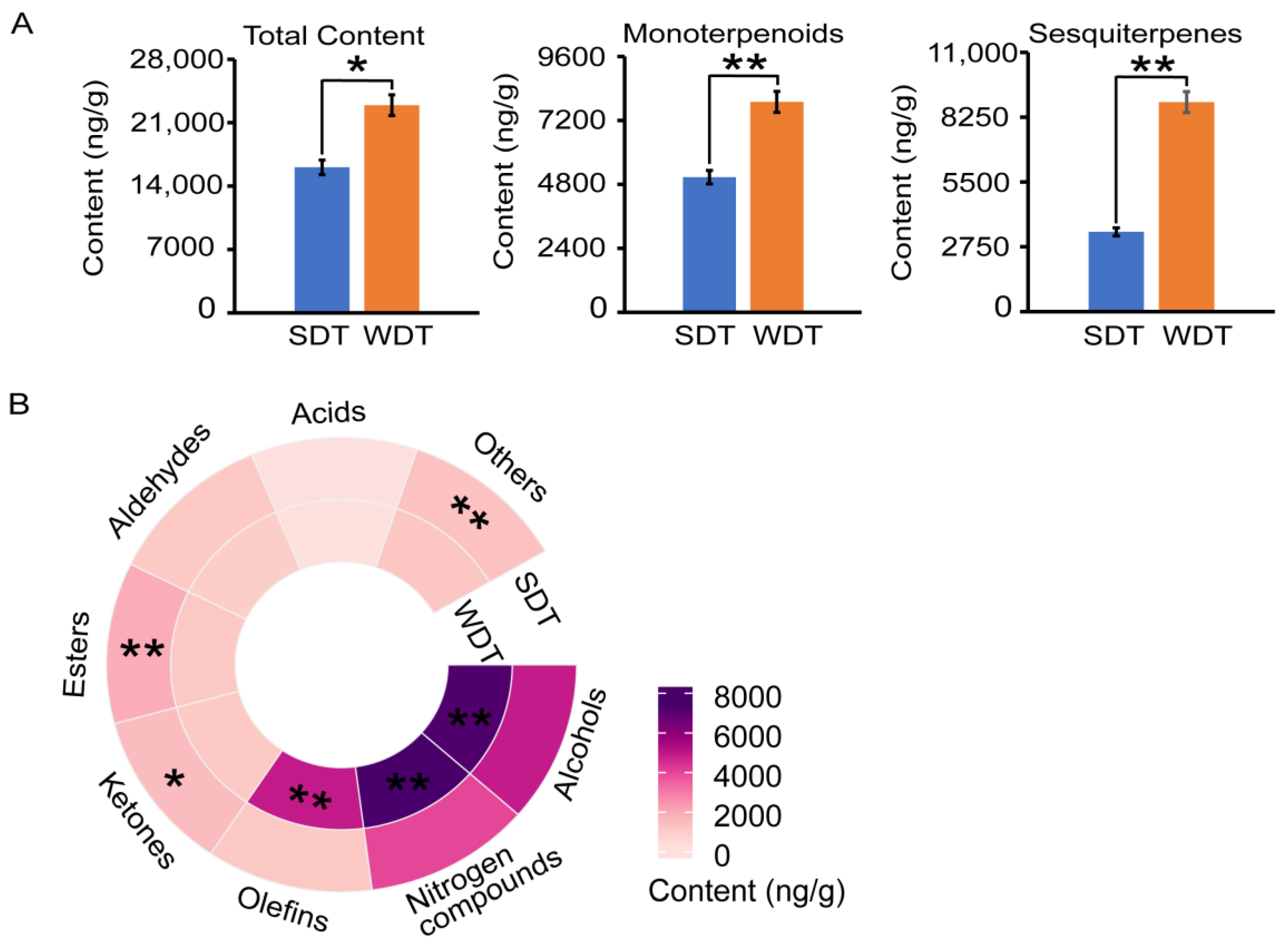
3.1. Volatile Components of DT
3.2. Quality Analysis of DT
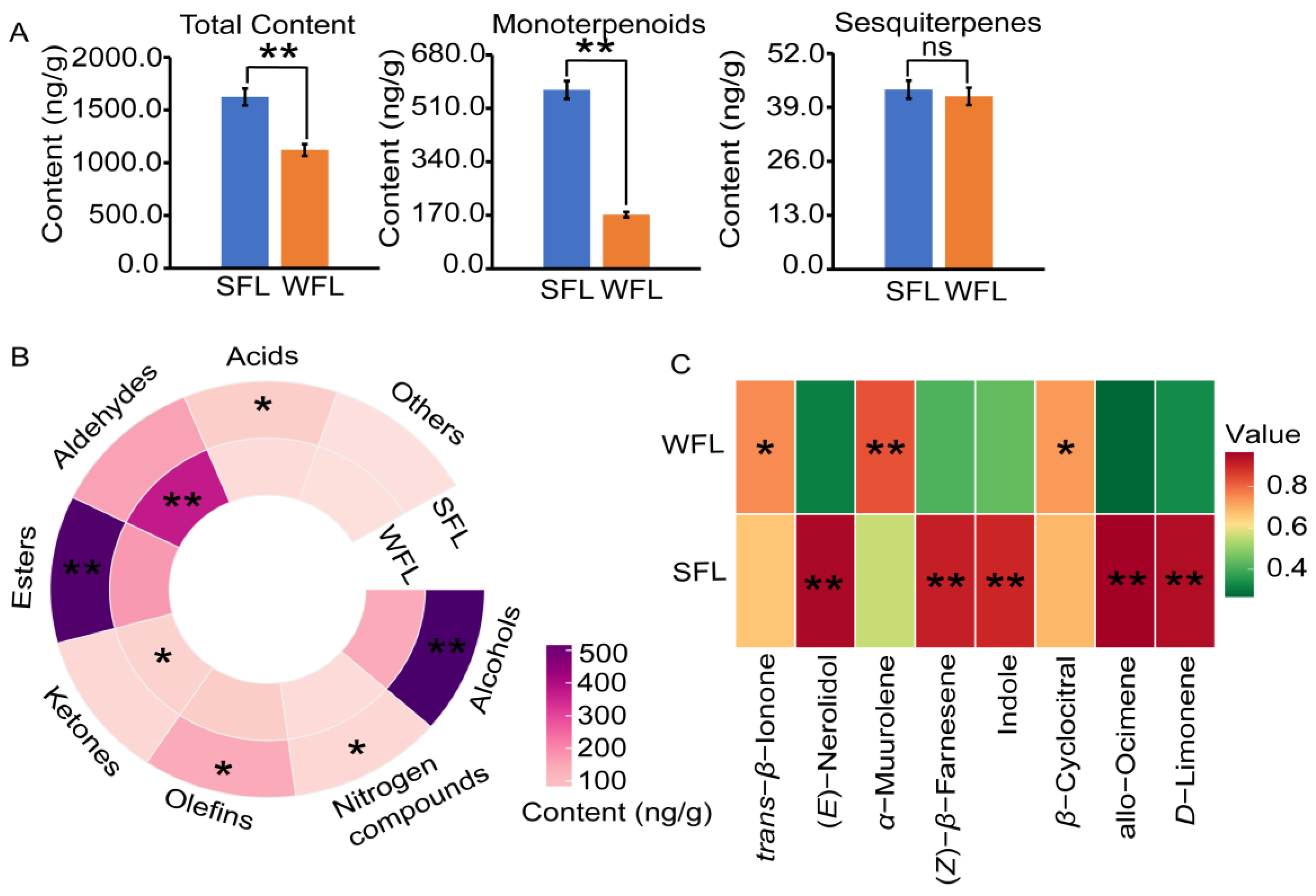
3.3. Volatile Components of FLs
3.4. Screening and Analysis of DEGs
3.5. Quantitative RT-PCR Analysis
4. Discussion
4.1. Superior Aroma Profile of WDT over SDT
4.2. Low Temperatures Influence the Aroma of DT
4.3. Multiple Biological Pathways Involved in Regulating the Formation of Terpene Aroma Compounds in WDT
4.4. Expression of Key Genes Regulates Terpene Aroma Synthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeng, L.; Zhou, X.; Su, X.; Yang, Z. Chinese Oolong Tea: An Aromatic Beverage Produced under Multiple Stresses. Trends Food Sci. Technol. 2020, 106, 242–253. [Google Scholar] [CrossRef]
- Chen, P.; Cai, J.; Zheng, P.; Yuan, Y.; Tsewang, W.; Chen, Y.; Xiao, X.; Liao, J.; Sun, B.; Liu, S. Quantitatively Unravelling the Impact of High Altitude on Oolong Tea Flavor from Camellia sinensis Grown on the Plateaus of Tibet. Horticulture 2022, 8, 539. [Google Scholar] [CrossRef]
- Liu, G.; Yang, M.; Fu, J. Identification and Characterization of Two Sesquiterpene Synthase Genes Involved in Volatile-mediated Defense in Tea Plant (Camellia sinensis). Plant Physiol. Biochem. 2020, 155, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Wang, X.; Xiao, Y.; Gu, D.; Liao, Y.; Xu, X.; Jia, Y.; Deng, R.; Song, C.; Yang, Z. Elucidation of (Z)-3-Hexenyl-beta-glucopyranoside Enhancement Mechanism under Stresses from the Oolong Tea Manufacturing Process. J. Agric. Food Chem. 2019, 67, 6541–6550. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.G.; Lee, Y.R.; Lee, M.S.; Hwang, K.H.; Park, C.Y.; Kim, E.H.; Park, J.S.; Hong, Y.S. Diverse Metabolite Variations in Tea (Camellia sinensis L.) Leaves Grown under Various Shade Conditions Revisited: A Metabolomics Study. J. Agric. Food Chem. 2018, 66, 1889–1897. [Google Scholar] [CrossRef] [PubMed]
- Owuor, P.O.; Obaga, S.O.; Othieno, C.O. The Effects of Altitude on the Chemical Composition of Black tea. Sci. Food Agric. 1990, 50, 9–17. [Google Scholar] [CrossRef]
- Hu, C.; Li, D.; Ma, Y.; Zhang, W.; Lin, C.; Zheng, X.; Liang, Y.; Lu, J. Formation Mechanism of the Oolong Tea Characteristic Aroma during Bruising and Withering Treatment. Food Chem. 2018, 269, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Picazo-Aragones, J.; Terrab, A.; Balao, F. Plant Volatile Organic Compounds Evolution: Transcriptional Regulation, Epigenetics and Polyploidy. Int. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef]
- Zeng, L.; Jin, S.; Xu, Y.; Granato, D.; Fu, Y.; Sun, W.; Yin, J.; Xu, Y. Exogenous Stimulation-Induced Biosynthesis of Volatile Compounds: Aroma Formation of Oolong Tea at Postharvest Stage. Crit. Rev. Food Sci. Nutr. 2022, 64, 76–86. [Google Scholar] [CrossRef]
- Wang, P.; Yu, J.; Jin, S.; Chen, S.; Yue, C.; Wang, W.; Gao, S.; Cao, H.; Zheng, Y.; Gu, M.; et al. Genetic Basis of High Aroma and Stress Tolerance in the Oolong Tea Cultivar Genome. Hortic. Res. 2021, 8, 107. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, P.; Chen, X.; Sun, Y.; Yue, C.; Ye, N. Transcriptome and Metabolite Profiling Reveal Novel Insights into Volatile Heterosis in the Tea Plant (Camellia sinensis). Molecules 2019, 24, 3380. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. The 1-deoxy-D-xylulose-5-phosphate Pathway of Isoprenoid Biosynthesis in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Kempinski, C.; Jiang, Z.; Zinck, G.; Sato, S.J.; Ge, Z.; Clemente, T.E.; Chappell, J. Engineering Linear, Branched-chain Triterpene Metabolism in Monocots. Plant Biotechnol. J. 2019, 17, 373–385. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral Volatiles: From Biosynthesis to Function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef] [PubMed]
- Flugge, U.I.; Gao, W. Transport of Isoprenoid Intermediates Across Chloroplast Envelope Membranes. Plant Biol. 2005, 7, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Andersson, S.; Orlova, I.; Gatto, N.; Reichelt, M.; Rhodes, D.; Boland, W.; Gershenzon, J. The Nonmevalonate Pathway Supports both Monoterpene and Sesquiterpene Formation in Snapdragon Flowers. Proc. Natl. Acad. Sci. USA 2005, 102, 933–938. [Google Scholar] [CrossRef]
- Vranova, E.; Coman, D.; Gruissem, W. Network Analysis of the MVA and MEP Pathways for Isoprenoid Synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Xie, Z.; Kapteyn, J.; Gang, D.R. A Systems Biology Investigation of the MEP/Terpenoid and Shikimate/Phenylpropanoid Pathways Points to Multiple Levels of Metabolic Control in Sweet Basil Glandular Trichomes. Plant J. 2008, 54, 349–361. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, L.; Wang, J.; Jin, J.; Zhang, N.; Lei, L.; Gao, T.; Jing, T.; Zhang, S.; Wu, Y.; et al. Induction of Priming by Cold Stress via Inducible Volatile Cues in Neighboring Tea Plants. J. Integr. Plant Biol. 2020, 62, 1461–1468. [Google Scholar] [CrossRef]
- Zhao, M.; Zhang, N.; Gao, T.; Jin, J.; Jing, T.; Wang, J.; Wu, Y.; Wan, X.; Schwab, W.; Song, C. Sesquiterpene Glucosylation Mediated by Glucosyltransferase UGT91Q2 is Involved in the Modulation of Cold Stress Tolerance in Tea Plants. New Phytol. 2020, 226, 362–372. [Google Scholar] [CrossRef]
- Ohgami, S.; Ono, E.; Horikawa, M.; Murata, J.; Totsuka, K.; Toyonaga, H.; Ohba, Y.; Dohra, H.; Asai, T.; Matsui, K.; et al. Volatile Glycosylation in Tea Plants: Sequential Glycosylations for the Biosynthesis of Aroma beta-Primeverosides are Catalyzed by Two Camellia sinensis Glycosyltransferases. Plant Physiol. 2015, 168, 464–477. [Google Scholar] [CrossRef]
- Guo, X.; Ho, C.T.; Wan, X.; Zhu, H.; Liu, Q.; Wen, Z. Changes of Volatile Compounds and Odor Profiles in Wuyi Rock Tea during Processing. Food Chem. 2021, 341, 128230. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Song, C.; Ho, C.; Wan, X. Contribution of L-theanine to the Formation of 2,5-Dimethylpyrazine, a Key Roasted Peanutty Flavor in Oolong Tea during Manufacturing Processes. Food Chem. 2018, 263, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Niu, Y.; Xiao, Z. Characterization of the Key Aroma Compounds in Laoshan Green Teas by Application of Odour Activity Value (OAV), Gas Chromatography-mass Spectrometry-olfactometry (GC-MS-O) and Comprehensive Two-dimensional Gas Chromatography Mass Spectrometry (GC × GC-qMS). Food Chem. 2021, 339, 128136. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Lu, C.; Li, M.; Ye, Y.; Wei, X.; Tong, H. Identification of Key Aromatic Compounds in Congou Black Tea by Partial Least-square Regression with Variable Importance of Projection Scores and Gas Chromatography-mass Spectrometry/Gas Chromatography-olfactometry. J. Sci. Food Agric. 2018, 98, 5278–5286. [Google Scholar] [CrossRef] [PubMed]
- Zhaopeng, S. Tea Evaluation and Inspection; China Agriculture Press: Beijing, China, 2010; pp. 88–92. ISBN 978-7-109-14785-0. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-fast All-in-one FASTQ Preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-read Alignment with Bowtie 2. Nat. Methods 2012, 9, 354–357. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 121–357. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved Reconstruction of a Transcriptome from RNA-seq Reads. Nat. Biotechnol. 2015, 33, 290. [Google Scholar] [CrossRef]
- Sun, B.; Zhu, Z.; Cao, P.; Chen, H.; Chen, C.; Zhou, X.; Mao, Y.; Lei, J.; Jiang, Y.; Meng, W.; et al. Purple Foliage Coloration in Tea (Camellia sinensis L.) Arises from Activation of the R2R3-MYB Transcription Factor CsAN1. Sci. Rep. 2016, 6, 32534. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ho, C.; Schwab, W.; Wan, X. Effect of the Roasting Degree on Flavor Quality of Large-leaf Yellow Tea. Food Chem. 2021, 347, 129016. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Zhang, L.; Granvogl, M.; Ho, C.; Wan, X. Flavor of tea (Camellia sinensis): A Review on Odorants and Analytical Techniques. Compr. Rev. Food Sci. Food Saf. 2022, 21, 3867–3909. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Baldermann, S.; Watanabe, N. Recent Studies of the Volatile Compounds in Tea. Food Res. Int. 2013, 53, 585–599. [Google Scholar] [CrossRef]
- Zheng, X.; Li, Q.; Xiang, L.; Liang, Y. Recent Advances in Volatiles of Teas. Molecules 2016, 21, 338. [Google Scholar] [CrossRef] [PubMed]
- Ting, G.; Yucheng, Z.; Pengjie, W.; Mengya, G.; Xuejin, C.; Yaping, H.; Binghao, H.; Naixiang, Y. Genome-wide Identification and Expression Analysis of TPS Gene Family in Camellia sinensis. Chin. J. Appl. Environ. Biol. 2022, 28, 1485–1495. (In Chinese) [Google Scholar]
- Zhu, C.; Zhang, S.; Zhou, C.; Chen, L.; Zaripov, T.; Zhan, D.; Weng, J.; Lin, Y.; Lai, Z.; Guo, Y. Integrated Transcriptome, MicroRNA, and Phytochemical Analyses Reveal Roles of Phytohormone Signal Transduction and ABC Transporters in Flavor Formation of Oolong Tea (Camellia sinensis) during Solar Withering. J. Agric. Food Chem. 2020, 68, 12749–12767. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zeng, L.; Hou, X.; Liao, Y.; Yang, Z. Low Temperature Synergistically promotes Wounding-induced Indole Accumulation by Inducer of CBF Expression-mediated Alterations of Jasmonic Acid Signaling in Camellia sinensis. J. Exp. Bot. 2020, 71, 2172–2185. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, L.; Liu, X.; Gui, J.; Mei, X.; Fu, X.; Dong, F.; Tang, J.; Zhang, L.; Yang, Z. Formation of (E)-Nerolidol in Tea (Camellia sinensis) Leaves Exposed to Multiple Stresses during Tea Manufacturing. Food Chem. 2017, 231, 78–86. [Google Scholar] [CrossRef]
- Chen, H.; Yue, Y.; Yu, R.; Fan, Y. A Hedychium Coronarium Short Chain Alcohol Dehydrogenase is a Player in allo-Ocimene Biosynthesis. Plant Mol. Biol. 2019, 101, 297–313. [Google Scholar] [CrossRef]
- Havaux, M. beta-Cyclocitral and Derivatives: Emerging Molecular Signals Serving Multiple Biological Functions. Plant Physiol. Biochem. 2020, 155, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Steele, C.L.; Crock, J.; Bohlmann, J.; Croteau, R. Sesquiterpene synthases from grand fir (Abies grandis). Comparison of Constitutive and Wound-induced Activities, and cDNA Isolation, Characterization, and Bacterial Expression of Delta-selinene Synthase and Gamma-humulene Synthase. J. Biol. Chem. 1998, 273, 2078–2089. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; He, F.; Zhu, B.; Lan, Y.; Pan, Q.; Li, C.; Reeves, M.J.; Wang, J. Free and glycosidically bound aroma compounds in cherry (Prunus avium L.). Food Chem. 2014, 152, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ma, W.; Shi, J.; Zhu, Y.; Lin, Z.; Lv, H. Characterization of the key aroma compounds in Longjing tea using stir bar sorptive extraction (SBSE) combined with gas chromatography-mass spectrometry (GC–MS), gas chromatography-olfactometry (GC-O), odor activity value (OAV), and aroma recombination. Food Res. Int. 2020, 130, 108908. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volatile Compounds | Odor Type | Odor Threshold (ng/g) | OAV d | |
|---|---|---|---|---|
| SDT | WDT | |||
| (E)-Nerolidol | Floral | 10.00 c | 219.08 ± 35.33 | 427.05 ± 71.72 |
| (Z)-β-Farnesene | Floral | 87.00 a | 1.25 ± 0.17 | 3.07 ± 0.50 |
| Indole | Floral | 40.00 a | 75.51 ± 8.96 | 136.34 ± 10.74 |
| Linalool | Floral | 0.22 a | 3887.77 ± 660.68 | 4616.14 ± 762.64 |
| trans-β-Ionone | Floral | 0.01 a | 48,204.00 ± 7662.00 | 25,580.00 ± 4925.00 |
| α-Muurolene | Woody | 0.10 b | 439.50 ± 65.80 | 33,902.70 ± 2223.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Chen, P.; Zheng, P.; Chen, J.; Sun, B.; Liu, S. Transcriptomic Insights into the Enhanced Aroma of Guangdong Oolong Dry Tea (Camellia sinensis cv. Yashixiang Dancong) in Winter. Foods 2024, 13, 160. https://doi.org/10.3390/foods13010160
Zheng Y, Chen P, Zheng P, Chen J, Sun B, Liu S. Transcriptomic Insights into the Enhanced Aroma of Guangdong Oolong Dry Tea (Camellia sinensis cv. Yashixiang Dancong) in Winter. Foods. 2024; 13(1):160. https://doi.org/10.3390/foods13010160
Chicago/Turabian StyleZheng, Yanchun, Peifen Chen, Peng Zheng, Jiahao Chen, Binmei Sun, and Shaoqun Liu. 2024. "Transcriptomic Insights into the Enhanced Aroma of Guangdong Oolong Dry Tea (Camellia sinensis cv. Yashixiang Dancong) in Winter" Foods 13, no. 1: 160. https://doi.org/10.3390/foods13010160
APA StyleZheng, Y., Chen, P., Zheng, P., Chen, J., Sun, B., & Liu, S. (2024). Transcriptomic Insights into the Enhanced Aroma of Guangdong Oolong Dry Tea (Camellia sinensis cv. Yashixiang Dancong) in Winter. Foods, 13(1), 160. https://doi.org/10.3390/foods13010160