
Expression and Biochemical Characterization of a Novel Fucoidanase from Flavobacteriumalgicola with the Principal Product of Fucoidan-Derived Disaccharide


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Strains, and Culture Conditions
2.2. Gene Cloning and Plasmid Construction
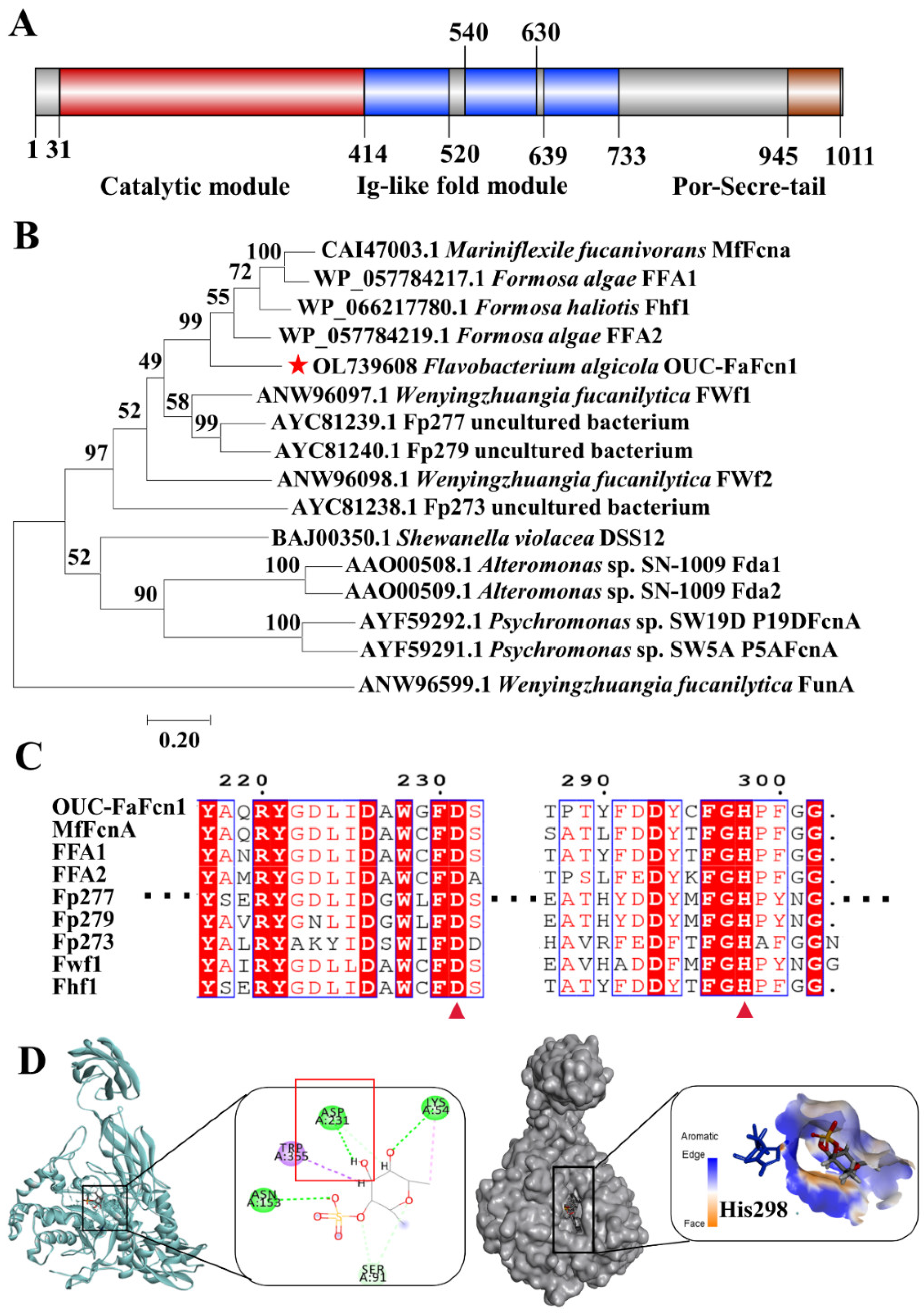
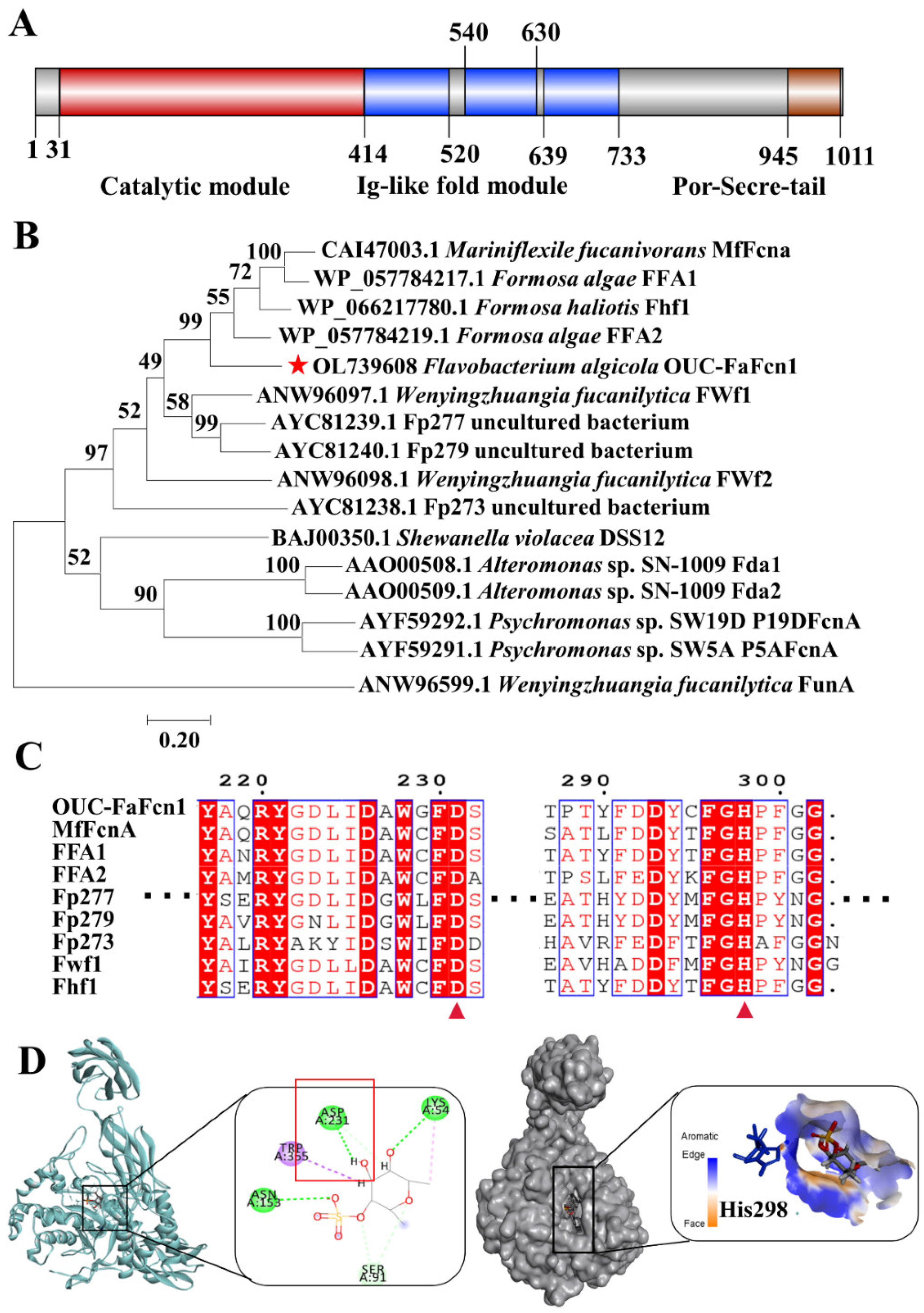
2.3. Sequence Analysis and Homology Dynamics Simulations
2.4. Production and Purification of Enzyme
2.5. Enzyme Activity Assay
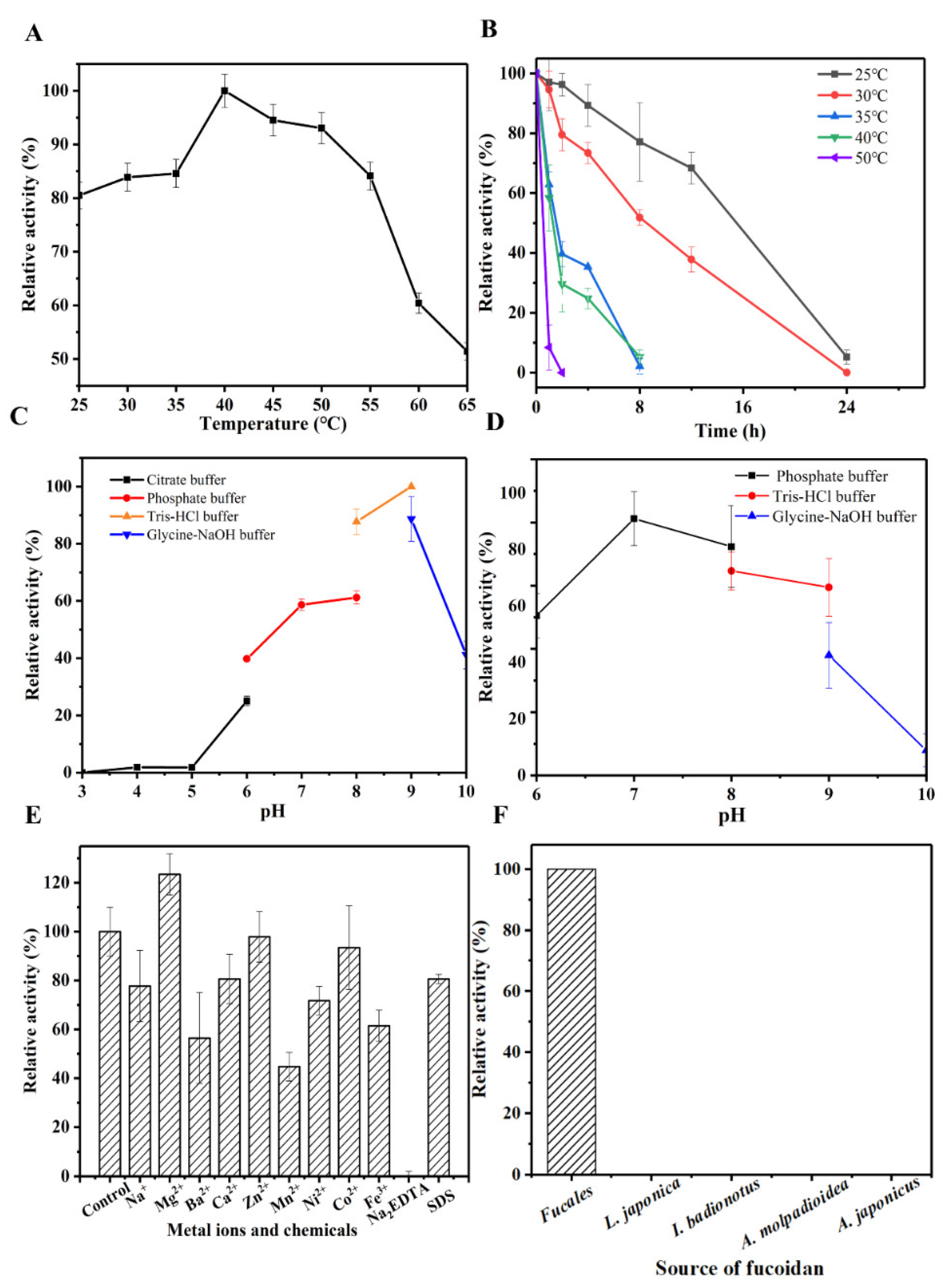
2.6. Characterization of Enzyme
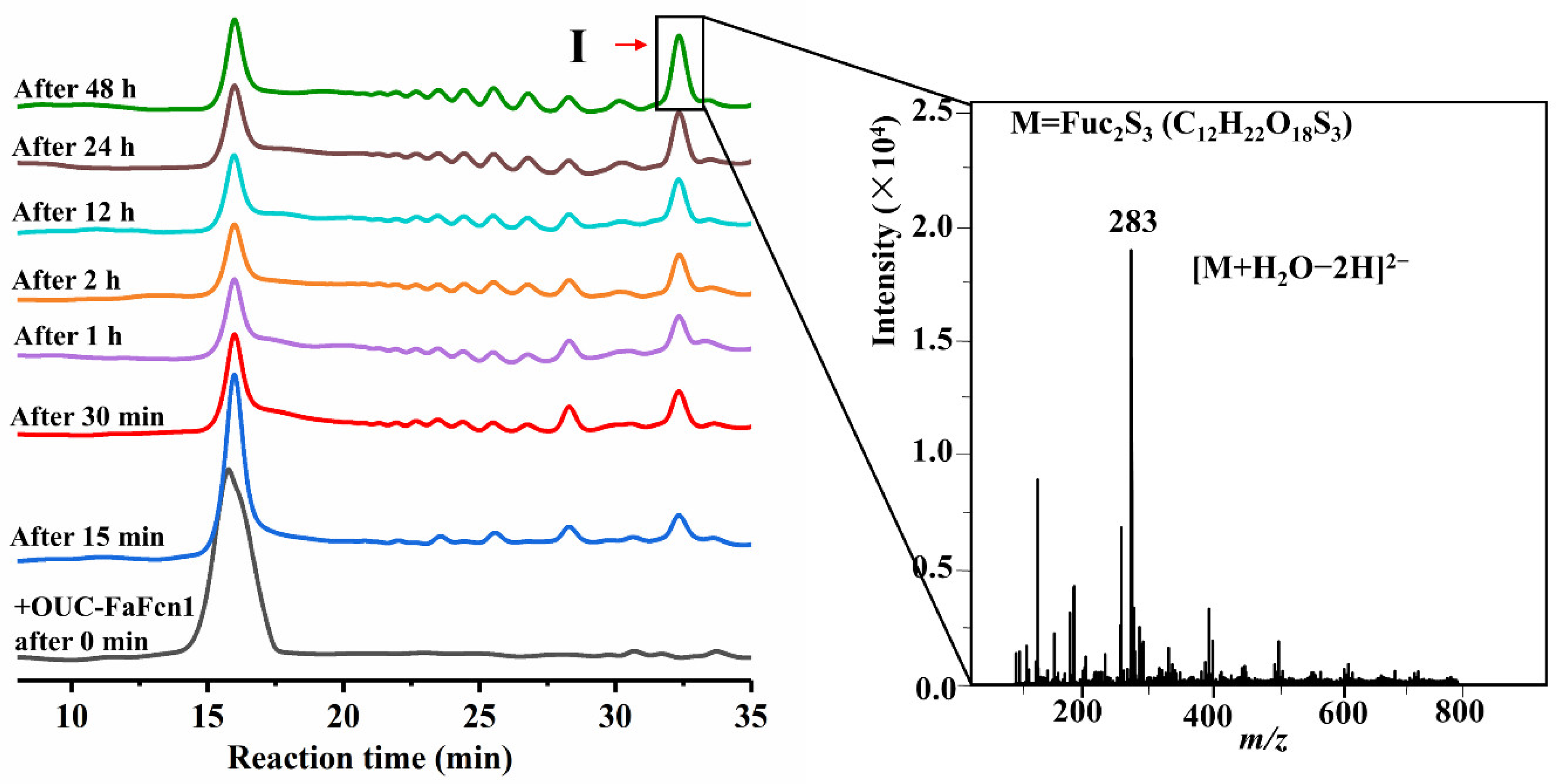
2.7. Analysis of Hydrolysis Products
3. Results and Discussion
3.1. Sequence Analysis of Fucoidanase OUC-FaFcn1
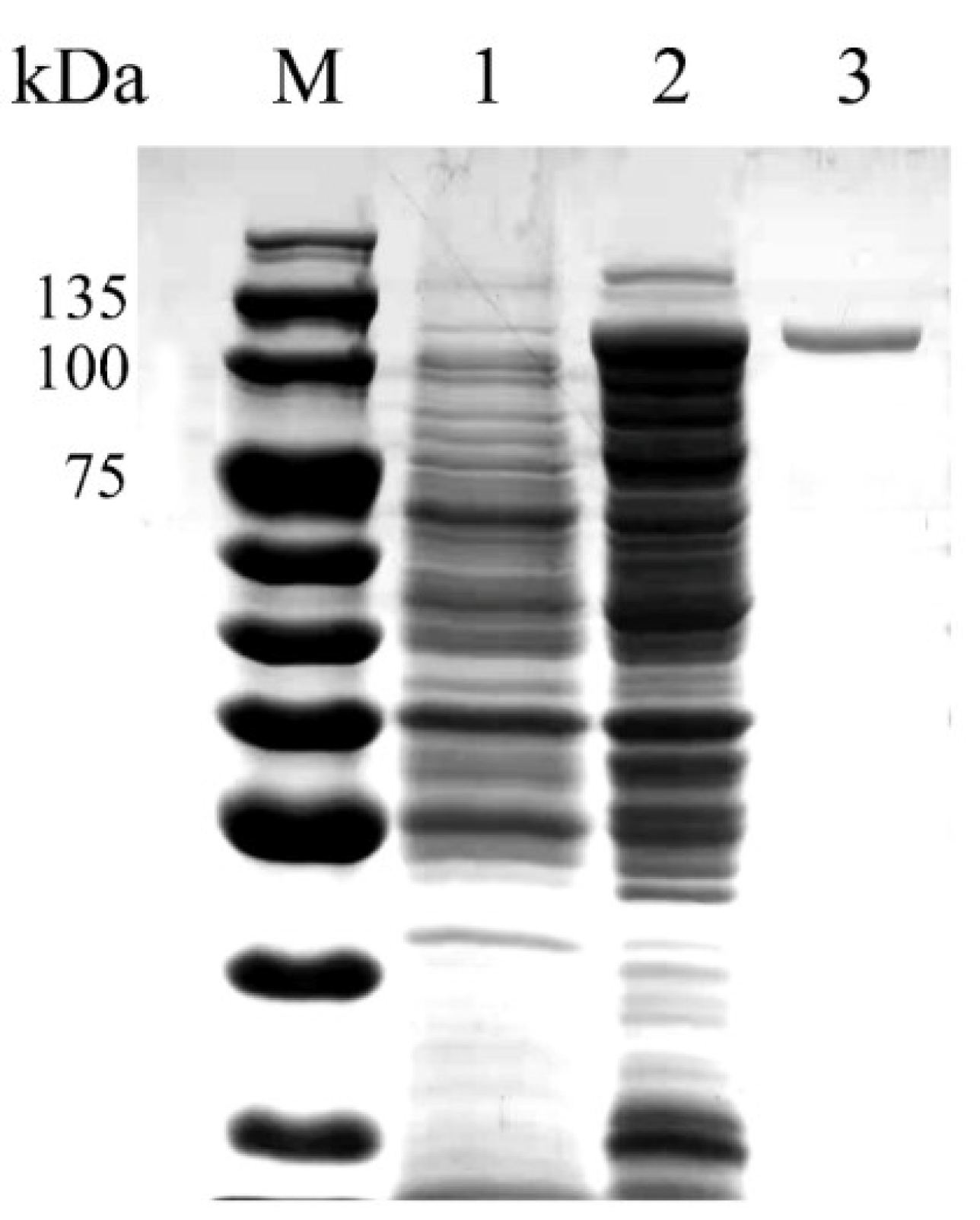
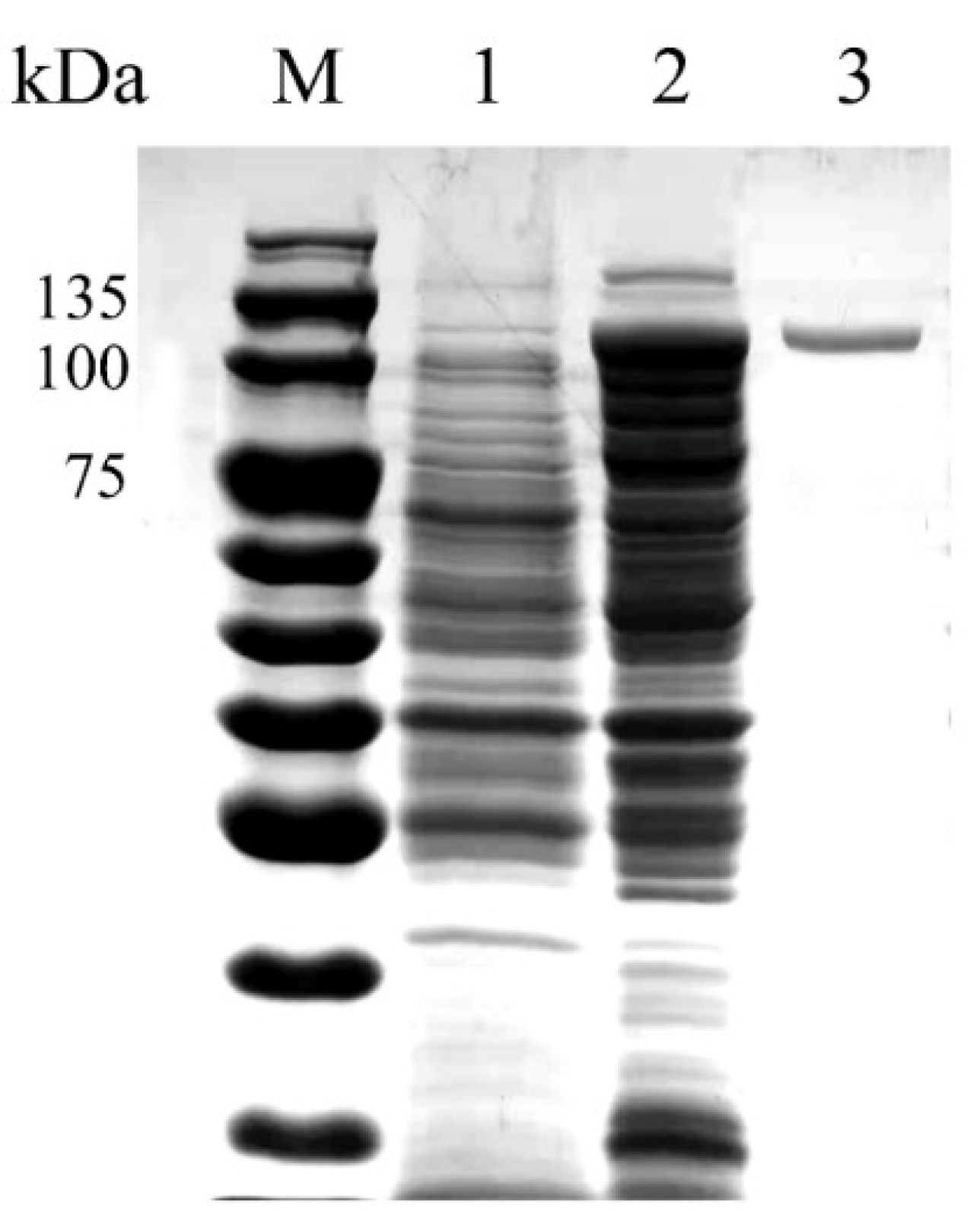
3.2. Heterologous Expression and Purification of Recombinant OUC-FaFcn1
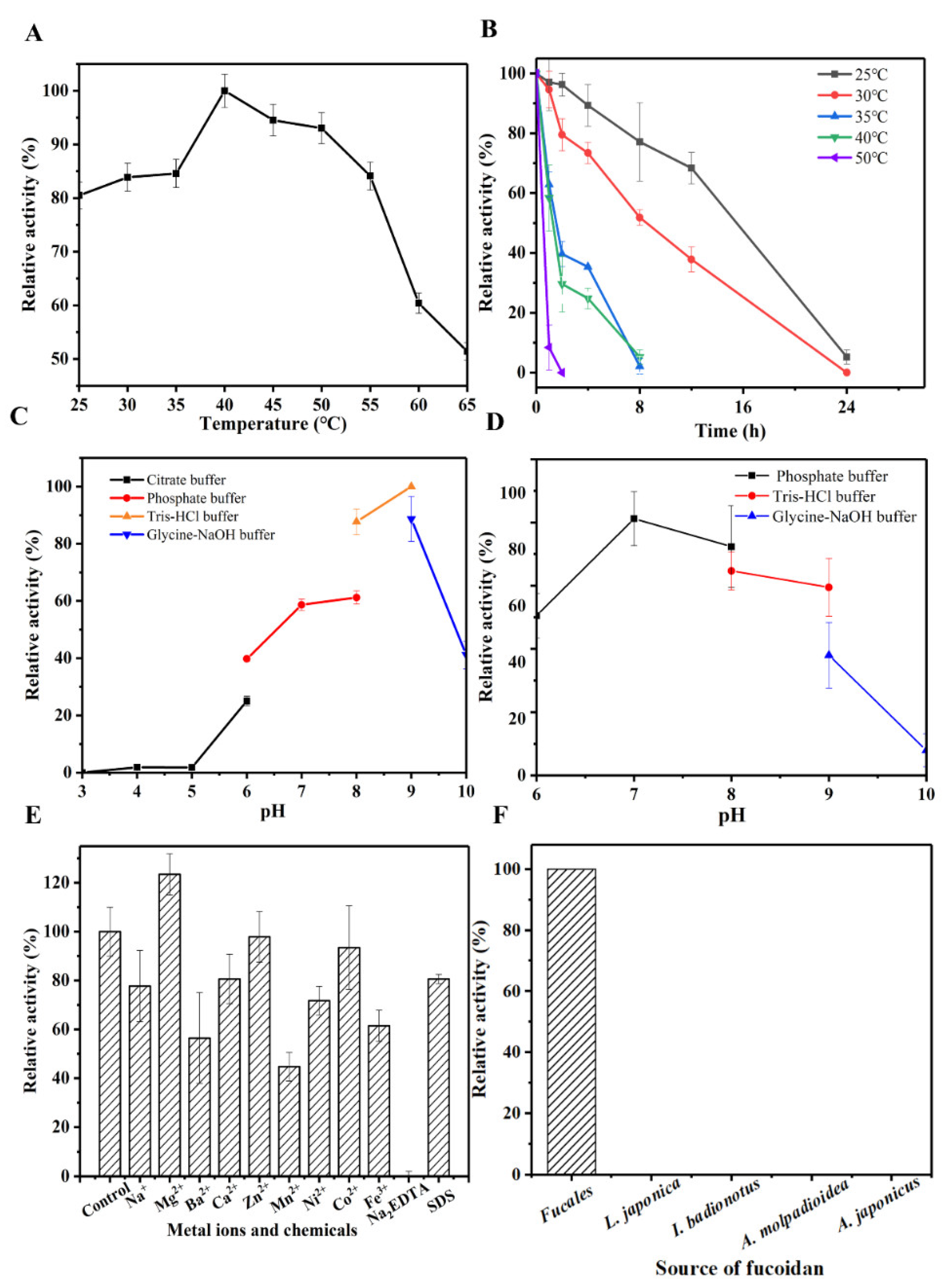
3.3. Characteristics of OUC-FaFcn1
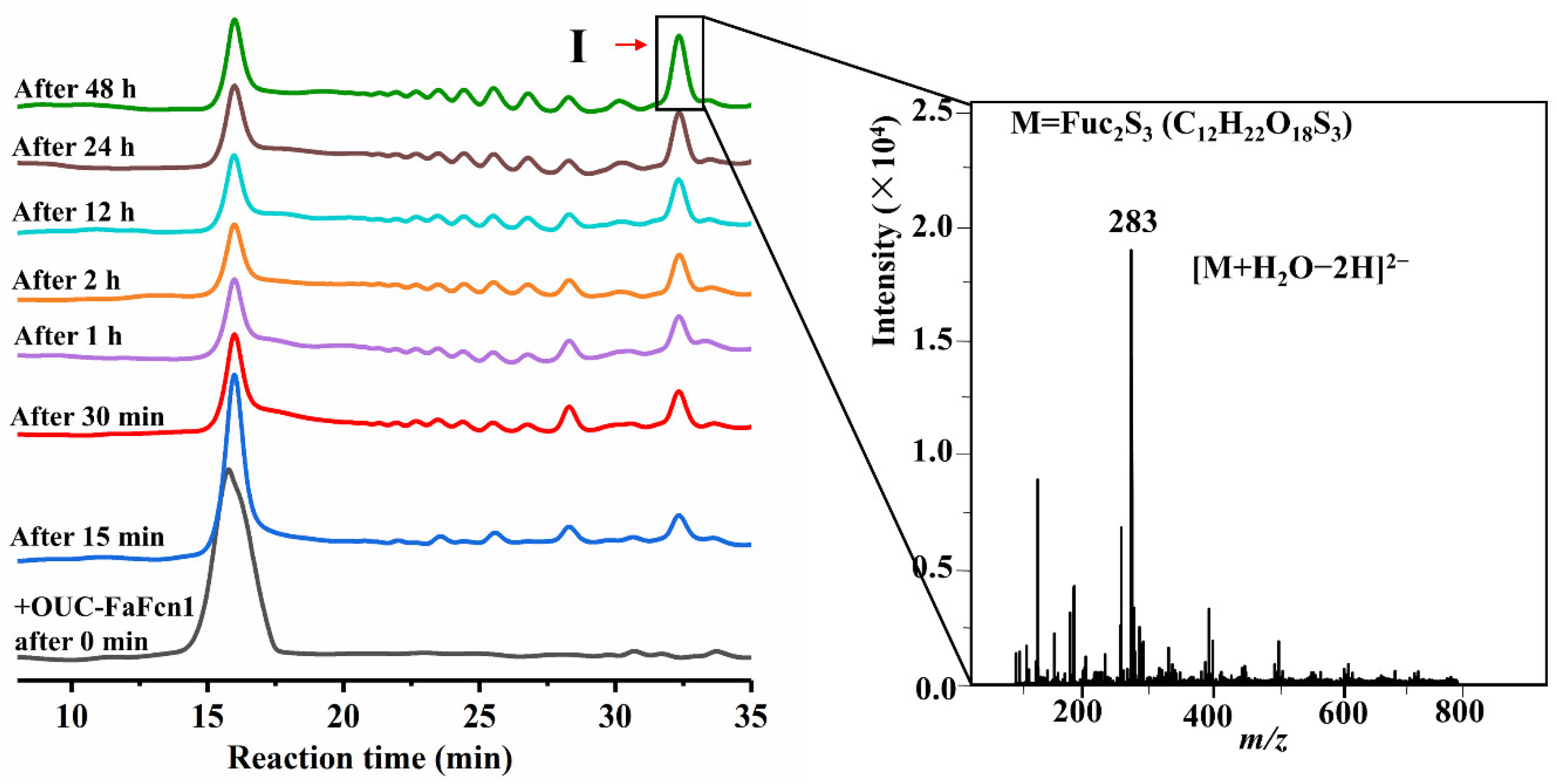
3.4. Hydrolysis Process and Principal Product Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kylin, H. Biochemistry of sea algae. Hoppe-Seyler Z. Physiol. Chem. 1913, 83, 171–197. [Google Scholar] [CrossRef]
- Mourao, P.A.; Bastos, I.G. Highly acidic glycans from sea cucumbers. Isolation and fractionation of fucose-rich sulfated polysaccharides from the body wall of Ludwigothurea grisea. Eur. J. Biochem. 1987, 166, 639–645. [Google Scholar] [CrossRef]
- Kannan, R.R.R.; Arumugam, R.; Anantharaman, P. Pharmaceutical potential of a fucoidan-like sulphated polysaccharide isolated from Halodule pinifolia. Int. J. Biol. Macromol. 2013, 62, 30–34. [Google Scholar] [CrossRef]
- Pereira, M.S.; Mulloy, B.; Mourao, P. Structure and anticoagulant activity of sulfated fucans. Comparison between the regular, repetitive, and linear fucans from echinoderms with the more heterogeneous and branched polymers from brown algae. Int. J. Biol. Macromol. 1999, 274, 7656–7667. [Google Scholar]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Mar. Drugs. 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deniaud-Bouet, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Herve, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef]
- Skriptsova, A.V. Seasonal variations in the fucoidan content of brown algae from Peter the Great Bay, Sea of Japan. Russ. J. Mar. Biol. 2016, 42, 351–356. [Google Scholar] [CrossRef]
- Usol’tseva, R.V.; Zhao, P.; Kusaikin, M.I.; Jia, A.; Yuan, W.; Zhang, M.; Liu, C.; Ermakova, S.P. Structural characteristics and antitumor activity of fucoidans from the brown alga Sargassum muticum. Chem. Nat. Compd. 2017, 53, 219–223. [Google Scholar] [CrossRef]
- Dobrincic, A.; Balbino, S.; Zoric, Z.; Pedisic, S.; Kovacevic, D.B.; Garofulic, I.E.; Dragovic-Uzelac, V. Advanced technologies for the extraction of marine brown algal polysaccharides. Mar. Drugs 2020, 18, 168. [Google Scholar] [CrossRef] [Green Version]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus serratus L. Carbohyd Res. 2006, 341, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Rasin, A.B.; Zueva, A.O.; Kusaykin, M.I.; Zvyagintseva, T.N.; Rubtsov, N.K.; Ermakova, S.P. Discovery of a fucoidan endo-4O-sulfatase: Regioselective 4O-desulfation of fucoidans and its effect on anticancer activity in vitro. Carbohyd. Polym. 2021, 271, 118449. [Google Scholar] [CrossRef] [PubMed]
- Kasai, A.; Arafuka, S.; Koshiba, N.; Takahashi, D.; Toshima, K. Systematic synthesis of low-molecular weight fucoidan derivatives and their effect on cancer cells. Org. Biomol. Chem. 2015, 13, 10556–10568. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, Y.; Cao, M.-J.; Liu, G.-M.; Chen, Q.; Sun, L.; Chen, H. Antibacterial activity and mechanisms of depolymerized fucoidans isolated from Laminaria japonica. Carbohyd. Polym. 2017, 172, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Kou, L.; Wang, F.; Wang, Y. Size-dependent whitening activity of enzyme-degraded fucoidan from Laminaria japonica. Carbohyd. Polym. 2019, 225, 115211. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Gao, L.; Zhou, H.; Ai, C.; Huang, X.; Wang, M.; Zhang, Y.; Zhao, C. Opportunities and challenges of algal fucoidan for diabetes management. Trends Food Sci Technol. 2021, 111, 628–641. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Shevchenko, L.S.; Nedashkovskaya, O.I.; Shevchenko, N.M.; Alekseeva, S.A.; Mikhailov, V.V.; Zvyagintseva, T.N. Screening of marine bacteria for fucoidanases. Mikrobiologiya 2000, 69, 370–376. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Nedashkovskaya, O.I.; Alekseeva, S.A.; Ivanova, E.P.; Romanenko, L.A.; Gorshkova, N.M.; Isakov, V.V.; Zvyagintseva, T.N.; Mikhailov, V.V. Degradation of fucoidan by the marine proteobacterium Pseudoalteromonas citrea. Microbiology 2002, 71, 41–47. [Google Scholar] [CrossRef]
- Sakai, T.; Ishizuka, K.; Kato, I. Isolation and characterization of a fucoidan-degrading marine bacterium. Mar. Biotechnol. 2003, 5, 409–416. [Google Scholar] [CrossRef]
- Sakai, T.; Kawai, T.; Kato, I. Isolation and characterization of a fucoidan-degrading marine bacterial strain and its fucoidanase. Mar. Biotechnol. 2004, 6, 335–346. [Google Scholar] [CrossRef]
- Colin, S.; Deniaud, E.; Jam, M.; Descamps, V.; Chevolot, Y.; Kervarec, N.; Yvin, J.C.; Barbeyron, T.; Michel, G.; Kloareg, B. Cloning and biochemical characterization of the fucanase FcnA: Definition of a novel glycoside hydrolase family specific for sulfated fucans. Glycobiology 2006, 16, 1021–1032. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Rasin, A.B.; Kusaykin, M.I.; Kalinovsky, A.I.; Miansong, Z.; Changheng, L.; Malyarenko, O.; Zueva, A.O.; Zvyagintseva, T.N.; Ermakova, S.P. Structure, enzymatic transformation, anticancer activity of fucoidan and sulphated fucooligosaccharides from Sargassum horneri. Carbohyd. Polym. 2017, 175, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Ustyuzhanina, N.E.; Kusaykin, M.I.; Krylov, V.B.; Shashkov, A.S.; Dmitrenok, A.S.; Usoltseva, R.V.; Zueva, A.O.; Nifantiev, N.E.; Zvyagintseva, T.N. Expression and biochemical characterization and substrate specificity of the fucoidanase from Formosa algae. Glycobiology 2017, 27, 254–263. [Google Scholar] [PubMed] [Green Version]
- Vickers, C.; Liu, F.; Abe, K.; Salama-Alber, O.; Jenkins, M.; Springate, C.M.K.; Burke, J.E.; Withers, S.G.; Boraston, A.B. Endo-fucoidan hydrolases from glycoside hydrolase family 107 (GH107) display structural and mechanistic similarities to -l-fucosidases from GH29. J. Biol. Chem. 2018, 293, 18296–18308. [Google Scholar] [CrossRef] [Green Version]
- Vuillemin, M.; Silchenko, A.S.; Cao, H.T.T.; Kokoulin, M.S.; Trang, V.T.D.; Holck, J.; Ermakova, S.P.; Meyer, A.S.; Mikkelsen, M.D. Functional characterization of a new GH107 endo-alpha-(1,4)-fucoidanase from the marine bacterium Formosa haliotis. Mar. Drugs. 2020, 18, 562. [Google Scholar] [CrossRef] [PubMed]
- Trang, V.T.D.; Mikkelsen, M.D.; Vuillemin, M.; Meier, S.; Cao, H.T.T.; Muschiol, J.; Perna, V.; Nguyen, T.T.; Tran, V.H.N.; Holck, J.; et al. The endo-α(1,4) specific fucoidanase Fhf2 from Formosa haliotis releases highly sulfated fucoidan oligosaccharides. Front. Plant Sci. 2022, 13, 823668. [Google Scholar] [CrossRef]
- Zueva, A.O.; Silchenko, A.S.; Rasin, A.B.; Kusaykin, M.I.; Usoltseva, R.V.; Kalinovsky, A.I.; Kurilenko, V.V.; Zvyagintseva, T.N.; Thinh, P.D.; Ermakova, S.P. Expression and biochemical characterization of two recombinant fucoidanases from the marine bacterium Wenyingzhuangia fucanilytica CZ1127(T). Int. J. Biol. Macromol. 2020, 164, 3025–3037. [Google Scholar] [CrossRef]
- Schultz-Johansen, M.; Cueff, M.; Hardouin, K.; Jam, M.; Larocque, R.; Glaring, M.A.; Herve, C.; Czjzek, M.; Stougaard, P. Discovery and screening of novel metagenome-derived GH107 enzymes targeting sulfated fucans from brown algae. FEBS J. 2018, 285, 4281–4295. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Chang, Y.; Zhang, Y.; Mei, X.; Xue, C. Discovery and characterization of an endo-1,3-Fucanase from marine bacterium Wenyingzhuangia fucanilytica: A novel glycoside hydrolase family. Front. Microbiol. 2020, 11, 1674. [Google Scholar] [CrossRef]
- Mika, M.; Shuki, F.; Yasuyoshi, N.; Makoto, N.; Noboru, T.; Tomoyuki, N.; Junichi, N. Flavobacterium algicola sp. nov., isolated from marine algae. Int. J. Syst. Evol. Microbiol. 2010, 60, 344–348. [Google Scholar]
- Jiang, C.; Jiang, H.; Zhang, T.; Lu, Z.; Mao, X. Enzymatic verification and comparative analysis of carrageenan metabolism pathways in marine bacterium Flavobacterium algicola. Appl. Environ. Microbiol. 2022, e00256-22. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.T.; Jiang, H.; Liang, X.X.; Qiu, Y.J.; Wang, L.L.; Mao, X.Z. Discovery and characterization of a novel alpha-L-fucosidase from the marine-derived Flavobacterium algicola and its application in 2 '-fucosyllactose production. Food Chem. 2022, 369, 130942. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Bichem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lever, M. A new reaction for colorimetric determination of carbohydrates. Anal. Bichem. 1972, 47, 273–279. [Google Scholar] [CrossRef]
- Nagao, T.; Arai, Y.; Yamaoka, M.; Komatsu, F.; Yagi, H.; Suzuki, H.; Ohshiro, T. Identification and characterization of the fucoidanase gene from Luteolibacter algae H18. J. Biosci. Bioeng. 2018, 126, 567–572. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Purification Fold | Yield (%) |
|---|---|---|---|---|---|
| Crude enzyme | 117.77 | 94.2 | 1.25 | 1 | 100 |
| Ni-NTA Superflow purification | 69.87 | 17 | 4.11 | 3.29 | 59.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Y.; Jiang, H.; Dong, Y.; Wang, Y.; Hamouda, H.I.; Balah, M.A.; Mao, X. Expression and Biochemical Characterization of a Novel Fucoidanase from Flavobacteriumalgicola with the Principal Product of Fucoidan-Derived Disaccharide. Foods 2022, 11, 1025. https://doi.org/10.3390/foods11071025
Qiu Y, Jiang H, Dong Y, Wang Y, Hamouda HI, Balah MA, Mao X. Expression and Biochemical Characterization of a Novel Fucoidanase from Flavobacteriumalgicola with the Principal Product of Fucoidan-Derived Disaccharide. Foods. 2022; 11(7):1025. https://doi.org/10.3390/foods11071025
Chicago/Turabian StyleQiu, Yanjun, Hong Jiang, Yueyang Dong, Yongzhen Wang, Hamed I. Hamouda, Mohamed A. Balah, and Xiangzhao Mao. 2022. "Expression and Biochemical Characterization of a Novel Fucoidanase from Flavobacteriumalgicola with the Principal Product of Fucoidan-Derived Disaccharide" Foods 11, no. 7: 1025. https://doi.org/10.3390/foods11071025
APA StyleQiu, Y., Jiang, H., Dong, Y., Wang, Y., Hamouda, H. I., Balah, M. A., & Mao, X. (2022). Expression and Biochemical Characterization of a Novel Fucoidanase from Flavobacteriumalgicola with the Principal Product of Fucoidan-Derived Disaccharide. Foods, 11(7), 1025. https://doi.org/10.3390/foods11071025