
Effects of Moderate Enzymatic Hydrolysis on Structure and Functional Properties of Pea Protein



Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Enzymatic Hydrolysis of PP
2.3. Structure Properties
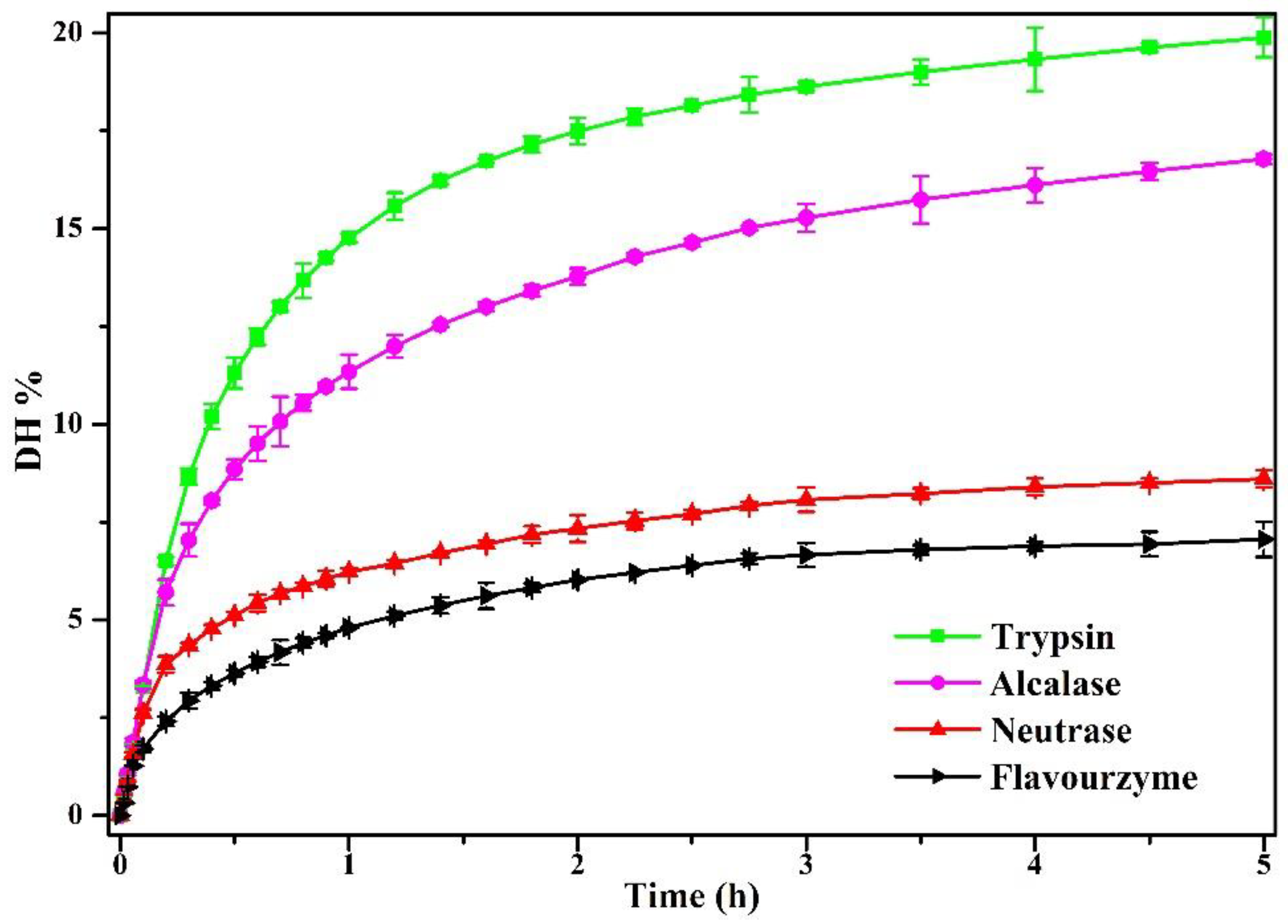
2.3.1. Degree of Enzymolysis (DH)
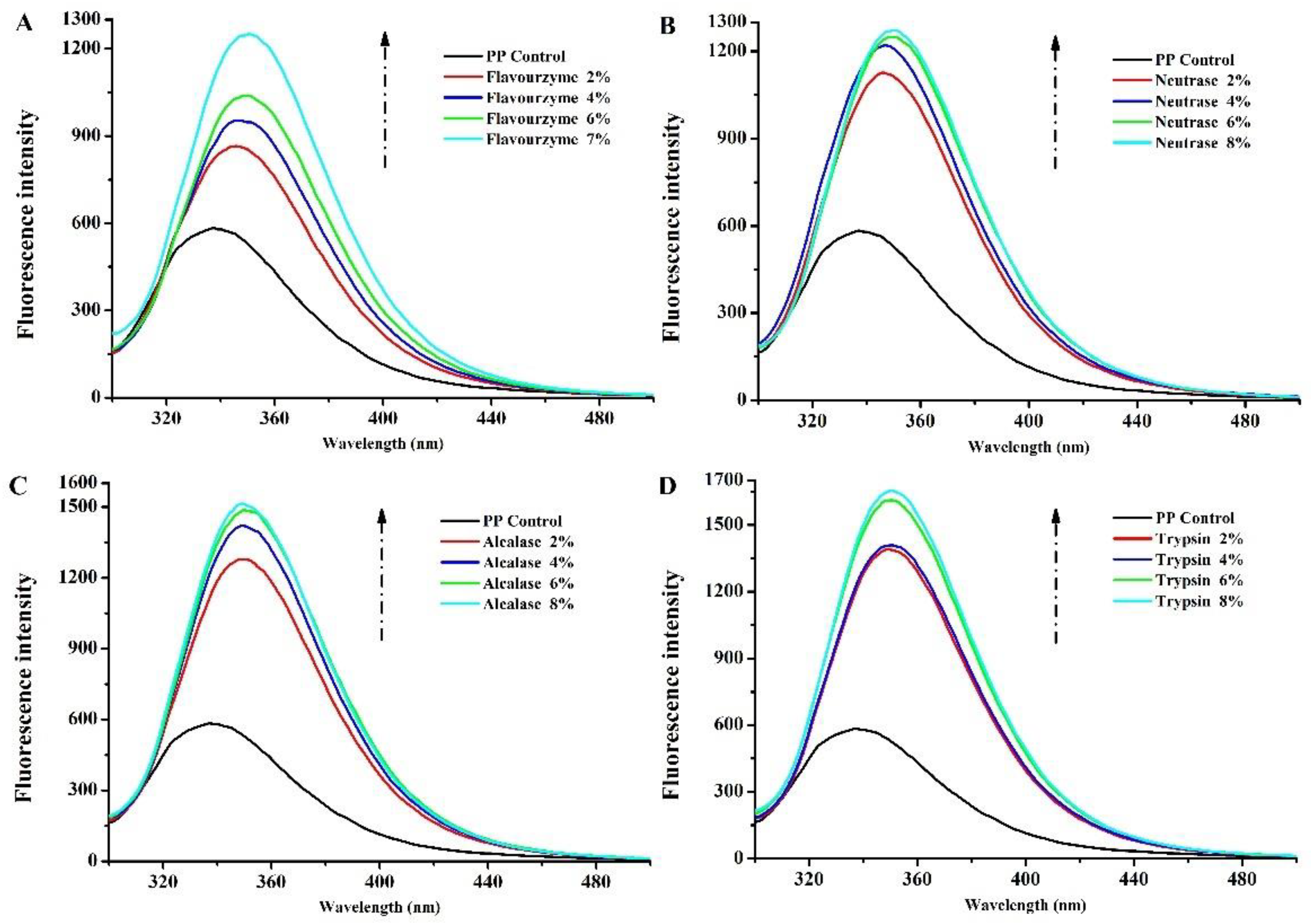
2.3.2. Intrinsic Fluorescence Spectroscopy
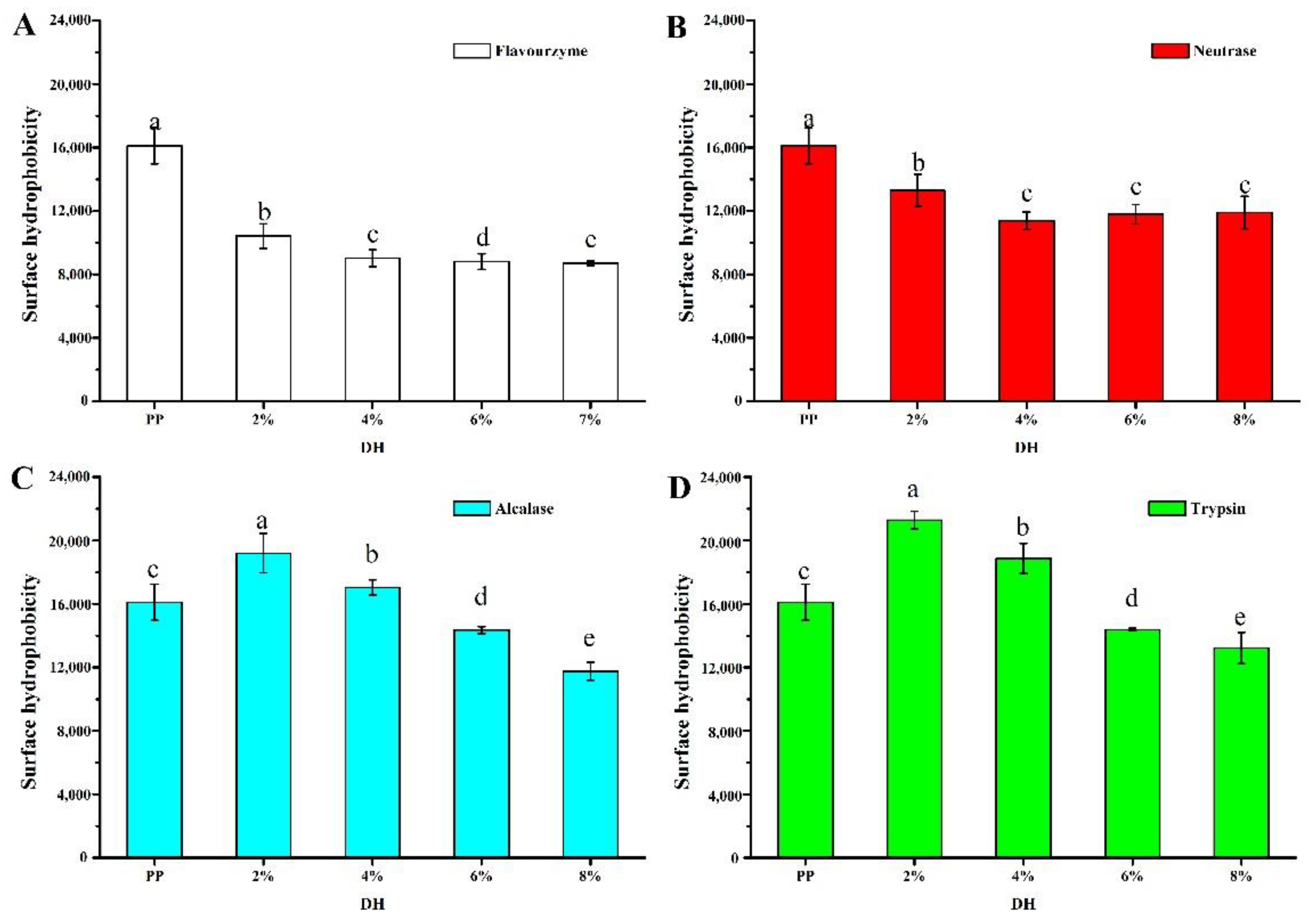
2.3.3. Surface Hydrophobicity Measurements
2.3.4. Circular Dichroism (CD) Spectroscopy Studies
2.3.5. Morphology Observation
2.3.6. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.3.7. Determination of Sulfhydryl Groups (SH)
2.3.8. Amino Acid Composition Analysis
2.3.9. Hydrophobicity Analysis Based on Amino Acid Composition
2.3.10. Sensory Evaluation
2.4. Functional Properties
2.4.1. Solubility
2.4.2. Foaming Performance
2.4.3. Emulsifying Performance
2.5. Statistical Analyses
3. Results and Discussion
3.1. Structure Properties
3.1.1. Degree of Enzymolysis (DH)
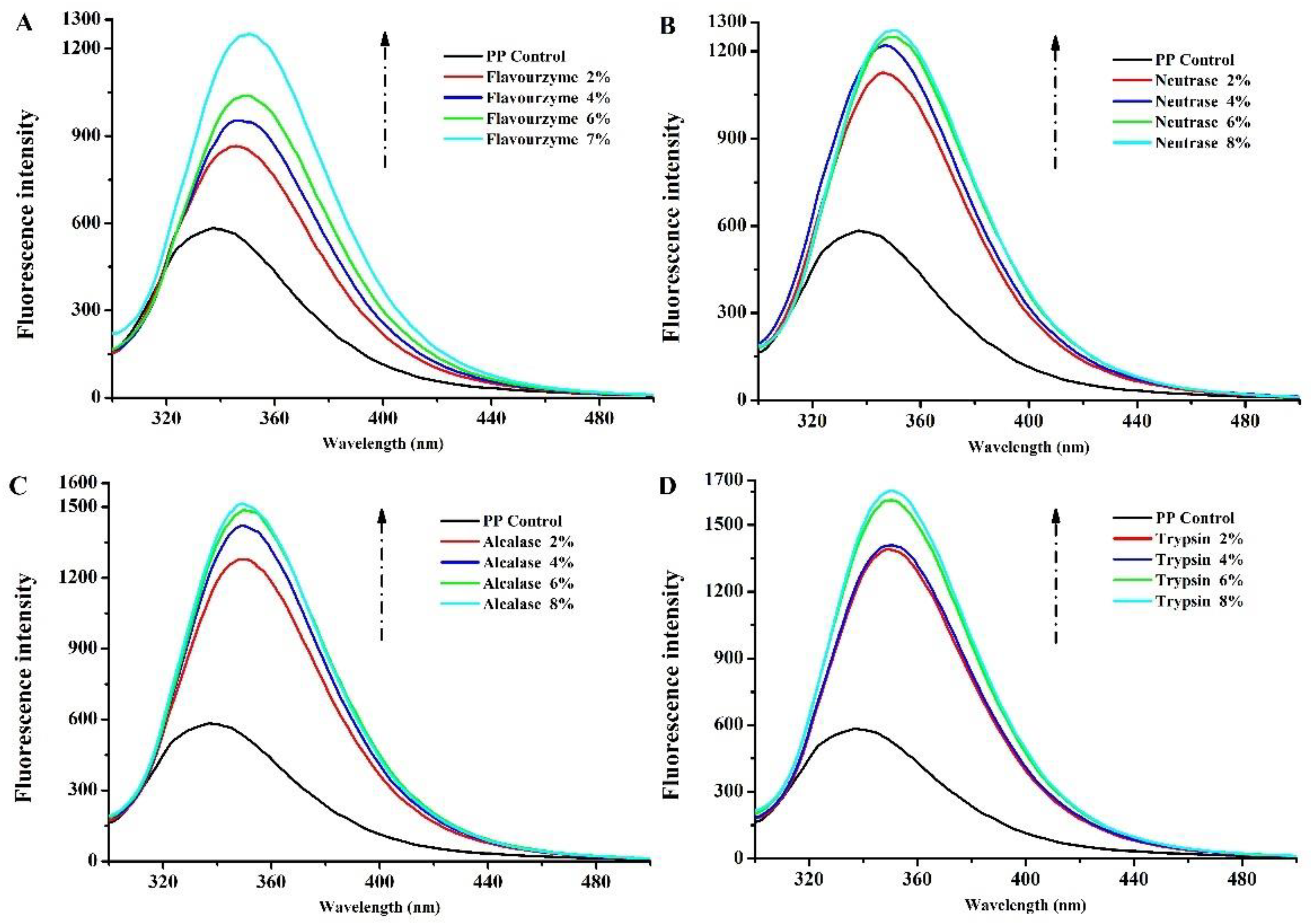
3.1.2. Intrinsic Fluorescence Spectroscopy
3.1.3. Surface Hydrophobicity
3.1.4. Circular Dichroism (CD) Spectroscopy Studies
3.1.5. Morphology Observation
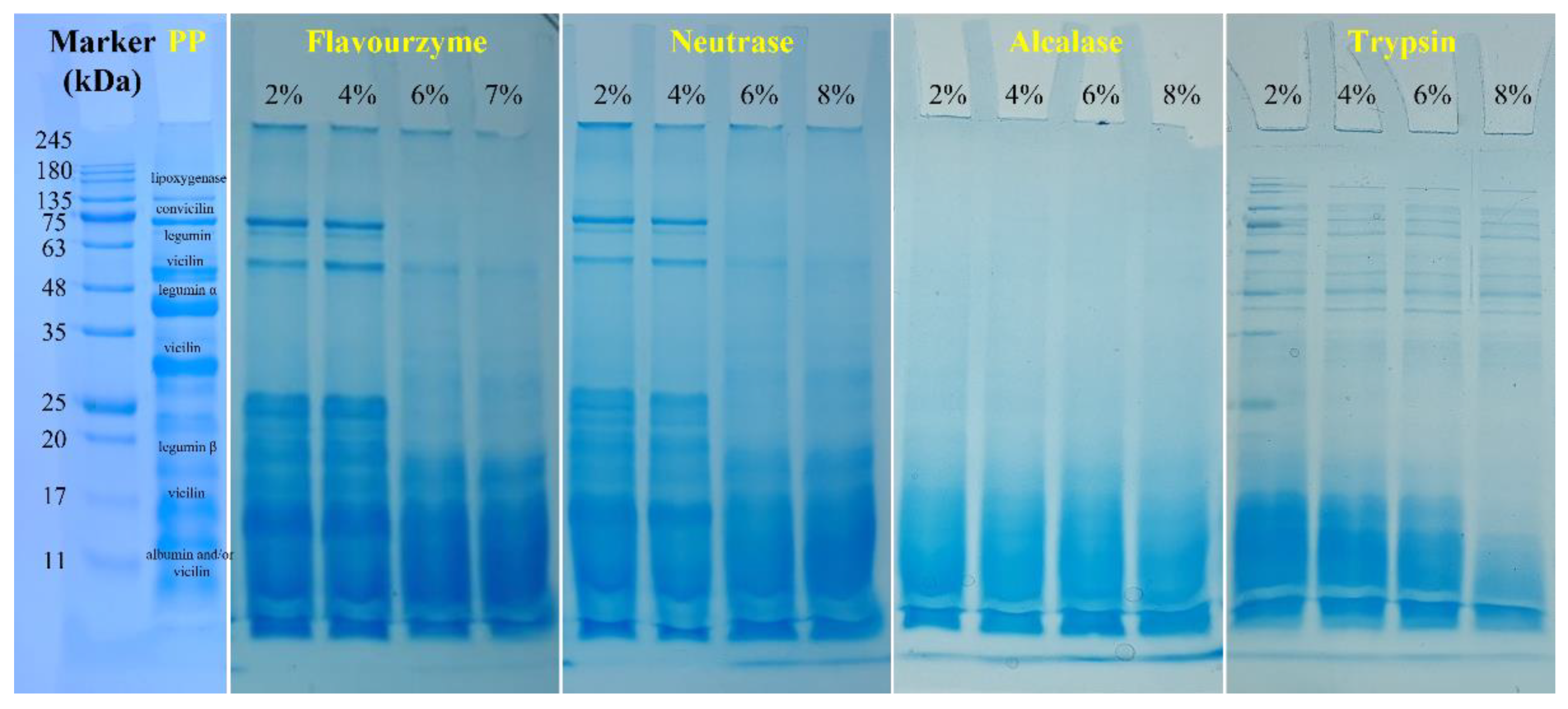
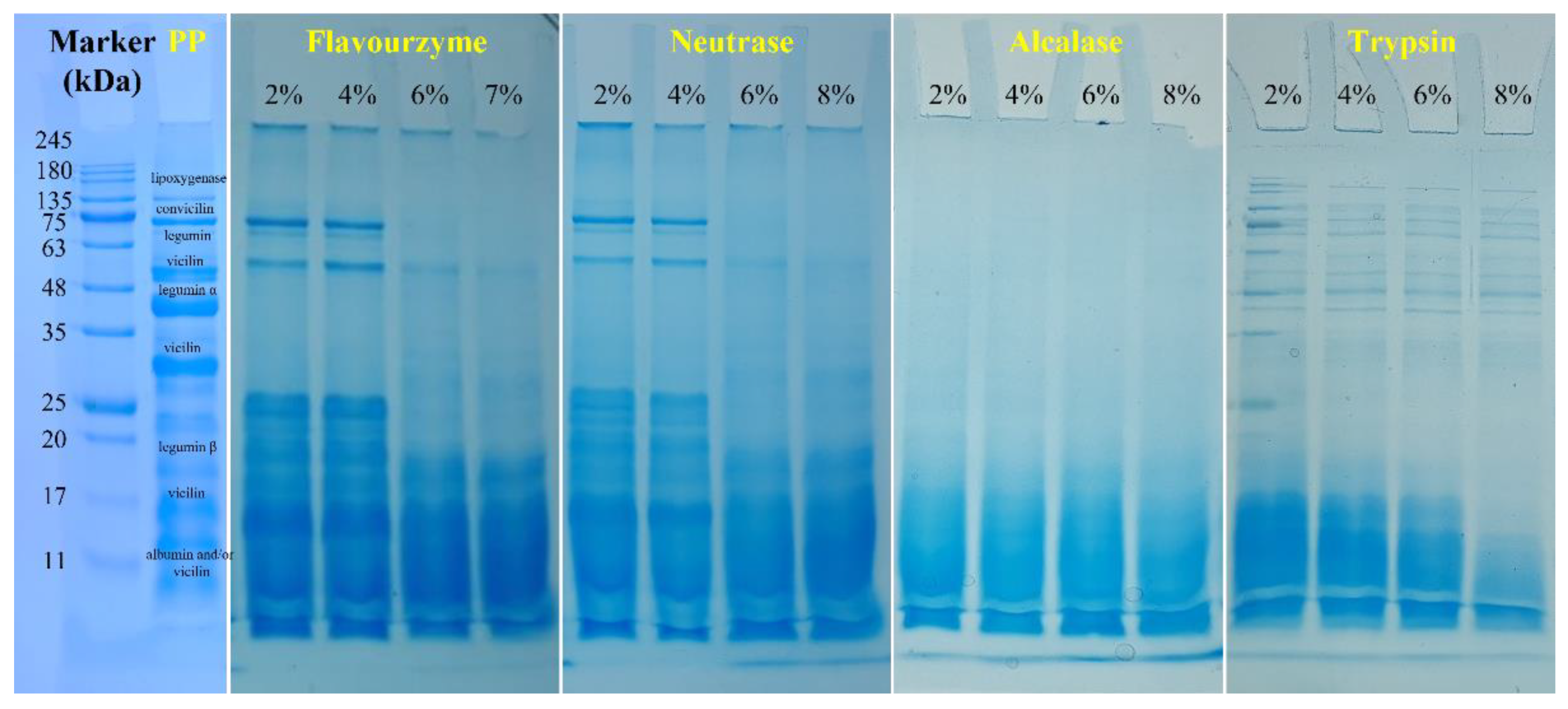
3.1.6. SDS-PAGE
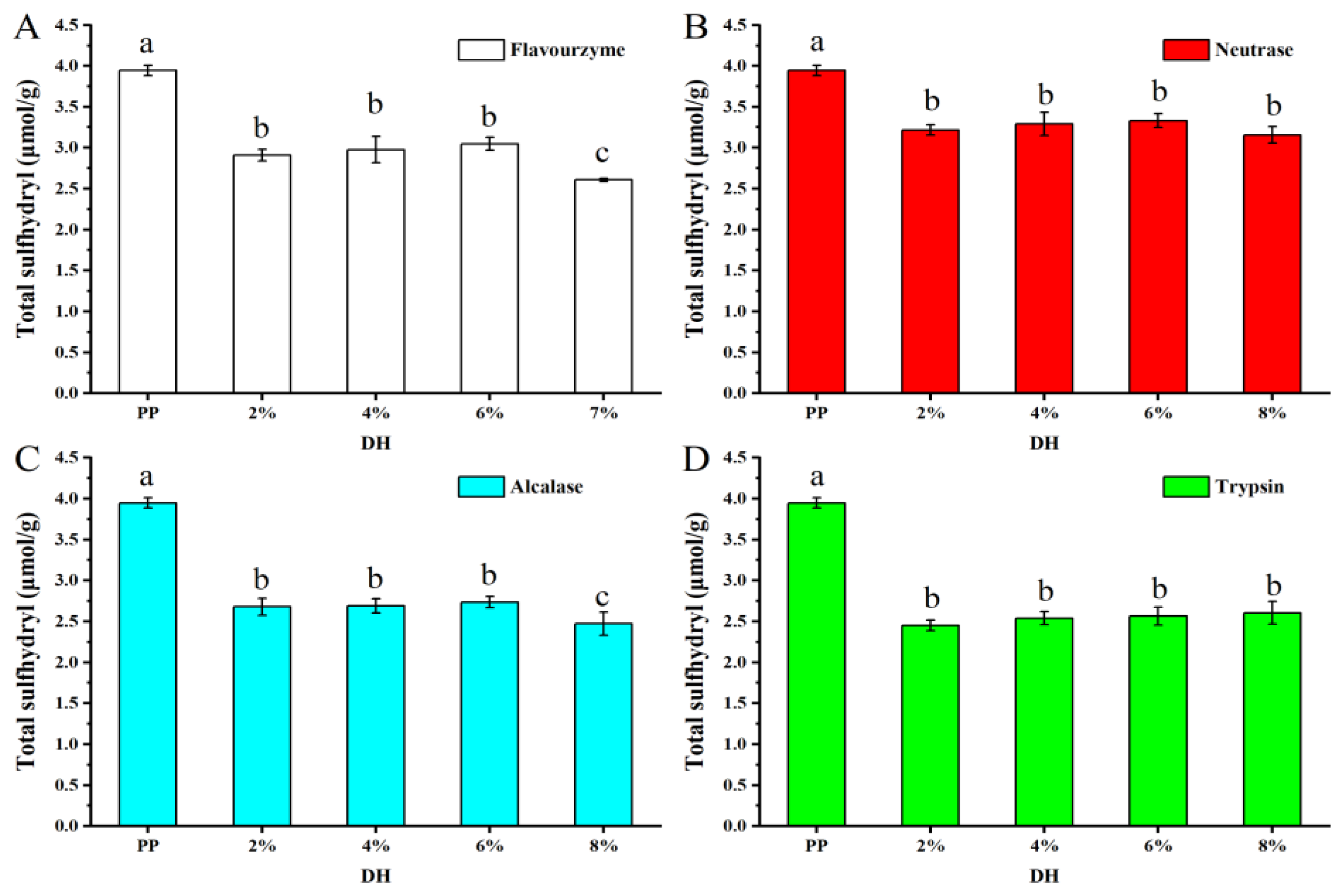
3.1.7. Determination of Total Sulfhydryl Groups
3.1.8. Amino Acid Composition and Average Hydrophobicity Analysis
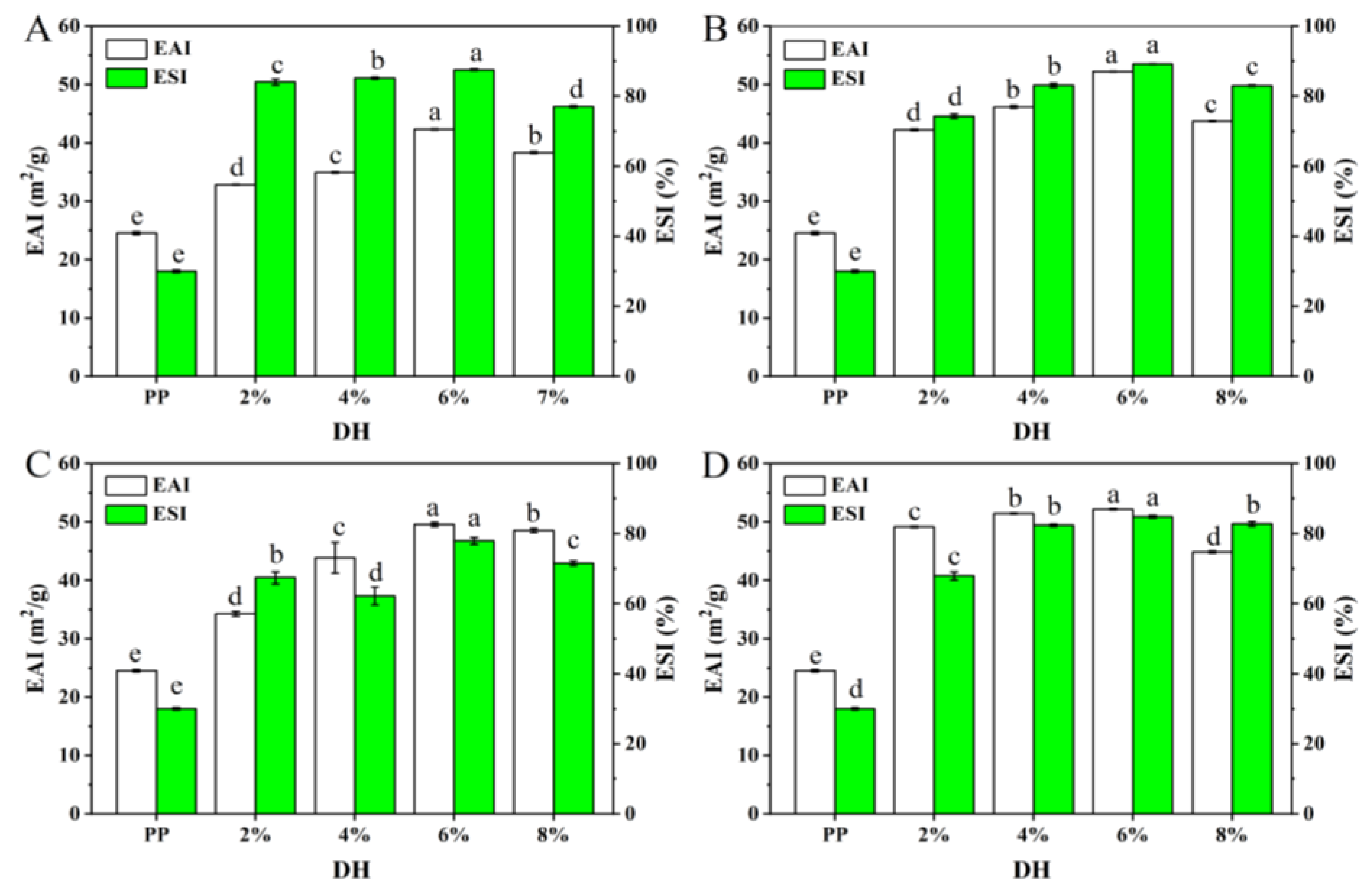
3.2. Functional Properties
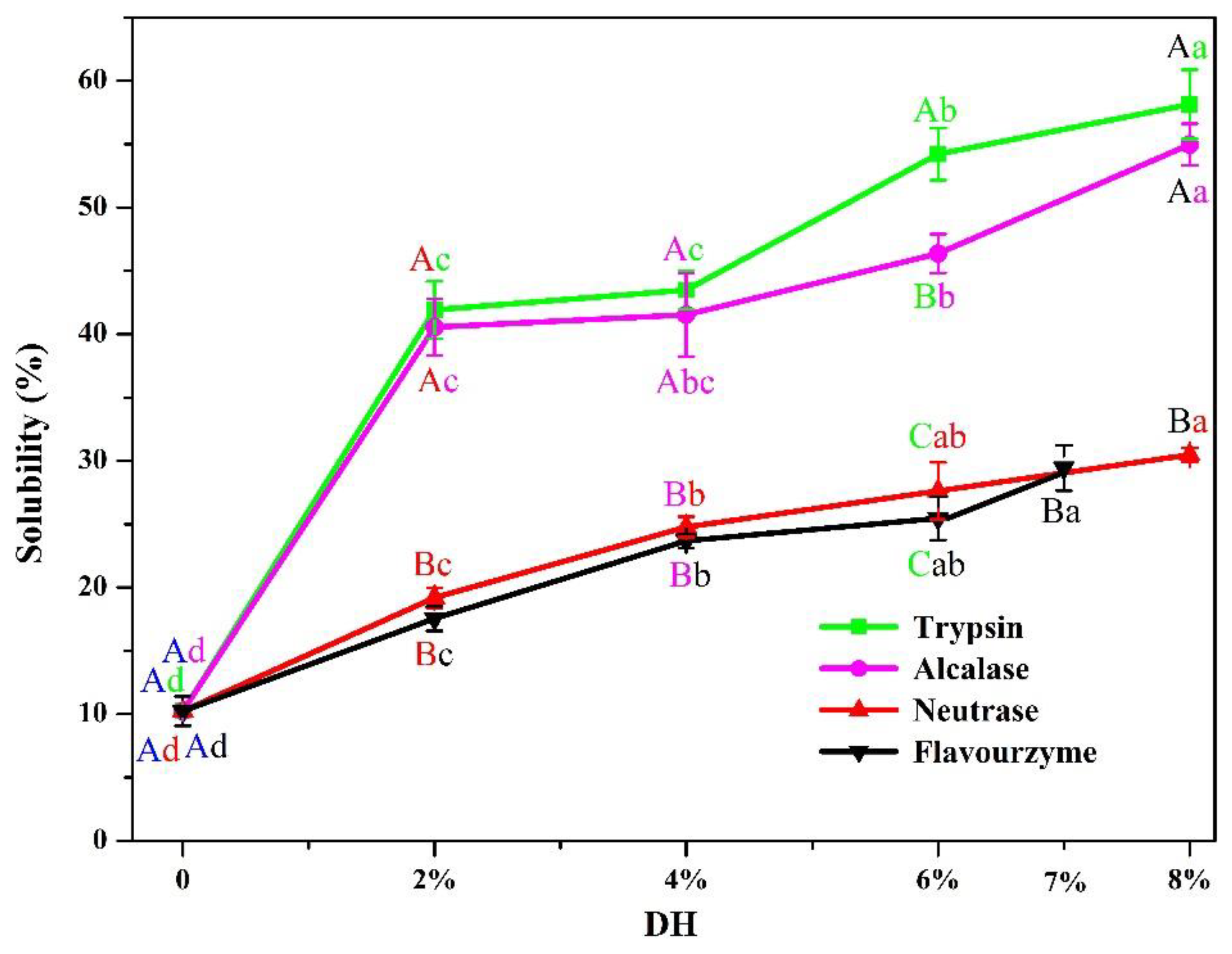
3.2.1. Solubility
3.2.2. Foaming Performance
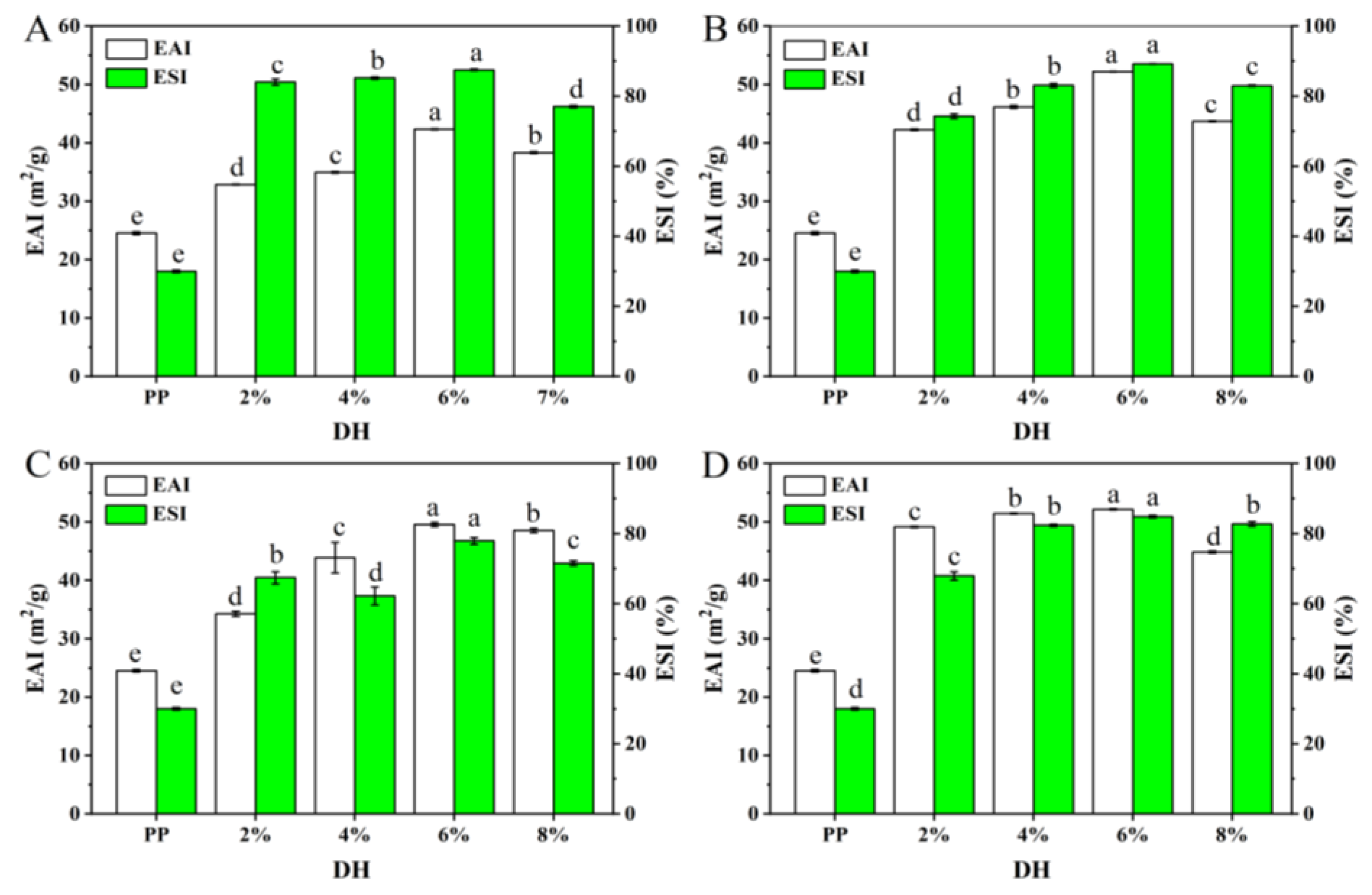
3.2.3. Emulsifying Performance
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burger, T.G.; Zhang, Y. Recent progress in the utilization of pea protein as an emulsifier for food applications. Trends Food Sci. Technol. 2019, 86, 25–33. [Google Scholar] [CrossRef]
- Dai, T.; Li, T.; Li, R.; Zhou, H.; Liu, C.; Chen, J.; McClements, D.J. Utilization of plant-based protein-polyphenol complexes to form and stabilize emulsions: Pea proteins and grape seed proanthocyanidins. Food Chem. 2020, 329, 127219. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Sun, C.X.; Corke, H.; Gul, K.; Gan, R.Y.; Fang, Y. The health benefits, functional properties, modifications, and applications of pea (Pisum sativum L.) protein: Current status, challenges, and perspectives. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1835–1876. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Xu, M.; Ohm, J.B.; Chen, B.; Rao, J. Solid dispersion-based spray-drying improves solubility and mitigates beany flavour of pea protein isolate. Food Chem. 2019, 278, 665–673. [Google Scholar] [CrossRef]
- He, X.; Chen, J.; He, X.; Feng, Z.; Li, C.; Liu, W.; Dai, T.; Liu, C. Industry-scale microfluidization as a potential technique to improve solubility and modify structure of pea protein. Innov. Food Sci. Emerg. Technol. 2020, 67, 102582. [Google Scholar] [CrossRef]
- Mahdavian Mehr, H.; Koocheki, A. Effect of atmospheric cold plasma on structure, interfacial and emulsifying properties of Grass pea (Lathyrus sativus L.) protein isolate. Food Hydrocoll. 2020, 106, 105899. [Google Scholar] [CrossRef]
- Wu, D.; Wu, C.; Ma, W.; Wang, Z.; Yu, C.; Du, M. Effects of ultrasound treatment on the physicochemical and emulsifying properties of proteins from scallops (Chlamys farreri). Food Hydrocoll. 2019, 89, 707–714. [Google Scholar] [CrossRef]
- Jiang, S.; Ding, J.; Andrade, J.; Rababah, T.M.; Almajwal, A.; Abulmeaty, M.M.; Feng, H. Modifying the physicochemical properties of pea protein by pH-shifting and ultrasound combined treatments. Ultrason.-Sonochem. 2017, 38, 835–842. [Google Scholar] [CrossRef]
- Wouters, A.G.B.; Rombouts, I.; Fierens, E.; Brijs, K.; Delcour, J.A. Relevance of the Functional Properties of Enzymatic Plant Protein Hydrolysates in Food Systems. Compr. Rev. Food Sci. Food Saf. 2016, 15, 786–800. [Google Scholar] [CrossRef] [Green Version]
- Meinlschmidt, P.; Sussmann, D.; Schweiggert-Weisz, U.; Eisner, P. Enzymatic treatment of soy protein isolates: Effects on the potential allergenicity, technofunctionality, and sensory properties. Food Sci. Nutr. 2016, 4, 11–23. [Google Scholar] [CrossRef]
- Schlegel, K.; Sontheimer, K.; Hickisch, A.; Wani, A.A.; Eisner, P.; Schweiggert-Weisz, U. Enzymatic hydrolysis of lupin protein isolates-Changes in the molecular weight distribution, technofunctional characteristics, and sensory attributes. Food Sci. Nutr. 2019, 7, 2747–2759. [Google Scholar] [CrossRef] [PubMed]
- García Arteaga, V.; Apéstegui Guardia, M.; Muranyi, I.; Eisner, P.; Schweiggert-Weisz, U. Effect of enzymatic hydrolysis on molecular weight distribution, techno-functional properties and sensory perception of pea protein isolates. Innov. Food Sci. Emerg. Technol. 2020, 65, 102449. [Google Scholar] [CrossRef]
- Klost, M.; Drusch, S. Functionalisation of pea protein by tryptic hydrolysis–Characterisation of interfacial and functional properties. Food Hydrocoll. 2019, 86, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Tamm, F.; Herbst, S.; Brodkorb, A.; Drusch, S. Functional properties of pea protein hydrolysates in emulsions and spray-dried microcapsules. Food Hydrocoll. 2016, 58, 204–214. [Google Scholar] [CrossRef]
- Klost, M.; Giménez-Ribes, G.; Drusch, S. Enzymatic hydrolysis of pea protein: Interactions and protein fractions involved in fermentation induced gels and their influence on rheological properties. Food Hydrocoll. 2020, 105, 105793. [Google Scholar] [CrossRef]
- Phoon, P.Y.; San Martin-Gonzalez, M.F.; Narsimhan, G. Effect of hydrolysis of soy β-conglycinin on the oxidative stability of O/W emulsions. Food Hydrocoll. 2014, 35, 429–443. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Enzymic Hydrolysis of Food Proteins; Elsevier Applied Science Publishers: New York, NY, USA, 1986; pp. 2210–2230. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Liu, W.; Liu, C.; Luo, L.; Chen, J.; Luo, S.; McClements, D.J.; Wu, L. Effect of limited enzymatic hydrolysis on structure and emulsifying properties of rice glutelin. Food Hydrocoll. 2016, 61, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Hu, P.; Dai, T.; Li, P.; Ye, X.; Chen, J.; Liu, C. Comparing the binding interaction between beta-lactoglobulin and flavonoids with different structure by multi-spectroscopy analysis and molecular docking. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 201, 197–206. [Google Scholar] [CrossRef]
- Li, X.; Dai, T.; Hu, P.; Zhang, C.; Chen, J.; Liu, C.; Li, T. Characterization the non-covalent interactions between beta lactoglobulin and selected phenolic acids. Food Hydrocoll. 2020, 105, 105761. [Google Scholar] [CrossRef]
- Yang, X.; Xu, J.; Fu, C.; Jia, Z.; Yao, K.; Chen, X. The cataract-related S39C variant increases gammaS-crystallin sensitivity to environmental stress by destroying the intermolecular disulfide cross-links. Biochem. Biophys. Res. Commun. 2020, 526, 459–465. [Google Scholar] [CrossRef]
- Xie, H.; Huang, J.; Woo, M.W.; Hu, J.; Xiong, H.; Zhao, Q. Effect of cold and hot enzyme deactivation on the structural and functional properties of rice dreg protein hydrolysates. Food Chem. 2021, 345, 128784. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.; Rosebrough, N.; Farr, A.L.; Randall, R. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Fathollahy, I.; Farmani, J.; Kasaai, M.R.; Hamishehkar, H. Characteristics and functional properties of Persian lime (Citrus latifolia) seed protein isolate and enzymatic Hydrolysates. LWT-Food Sci. Technol. 2021, 140, 110765. [Google Scholar] [CrossRef]
- Hou, F.; Ding, W.; Qu, W.; Oladejo, A.O.; Xiong, F.; Zhang, W.; He, R.; Ma, H. Alkali solution extraction of rice residue protein isolates: Influence of alkali concentration on protein functional, structural properties and lysinoalanine formation. Food Chem. 2017, 218, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Avramenko, N.A.; Low, N.H.; Nickerson, M.T. The effects of limited enzymatic hydrolysis on the physicochemical and emulsifying properties of a lentil protein isolate. Food Res. Int. 2013, 51, 162–169. [Google Scholar] [CrossRef]
- Roger, P.H. Determining the fluorescence spectrum of a protein. Curr. Protoc. Protein Sci. 2004, 7, 7.7.1. [Google Scholar] [CrossRef]
- Vera, A.; Valenzuela, M.A.; Yazdani-Pedram, M.; Tapia, C.; Abugoch, L. Conformational and physicochemical properties of quinoa proteins affected by different conditions of high-intensity ultrasound treatments. Ultrason. Sonochem. 2019, 51, 186–196. [Google Scholar] [CrossRef]
- Ling, B.; Ouyang, S.; Wang, S. Effect of radio frequency treatment on functional, structural and thermal behaviors of protein isolates in rice bran. Food Chem. 2019, 289, 537–544. [Google Scholar] [CrossRef]
- Burstein, E.A.; Vedenkina, N.S.; Ivkova, M.N. Fluorescence and the location of tryptophan residues in protein molecules. Photochem. Photobiol. 1973, 18, 263–279. [Google Scholar] [CrossRef]
- Sponton, O.E.; Perez, A.A.; Carrara, C.; Santiago, L.G. Effect of limited enzymatic hydrolysis on linoleic acid binding properties of beta-lactoglobulin. Food Chem. 2014, 146, 577–582. [Google Scholar] [CrossRef]
- Kang, D.C.; Zou, Y.H.; Cheng, Y.P.; Xing, L.J.; Zhou, G.H.; Zhang, W.G. Effects of power ultrasound on oxidation and structure of beef proteins during curing processing. Ultrason. Sonochem. 2016, 33, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Selomulya, C.; Wang, S.; Xiong, H.; Chen, X.D.; Li, W.; Peng, H.; Xie, J.; Sun, W.; Zhou, Q. Enhancing the oxidative stability of food emulsions with rice dreg protein hydrolysate. Food Res. Int. 2012, 48, 876–884. [Google Scholar] [CrossRef]
- Jung, S.; Murphy, P.A.; Johnson, L.A. Physicochemical and functional properties of soy protein substrates modified by low levels of protease hydrolysis. J. Food Sci. 2005, 70, 180–187. [Google Scholar] [CrossRef]
- Guan, X.; Yao, H.; Chen, Z.; Shan, L.; Zhang, M. Some functional properties of oat bran protein concentrate modified by trypsin. Food Chem. 2007, 101, 163–170. [Google Scholar] [CrossRef]
- Yong, Y.H.; Yamaguchi, S.; Matsumura, Y. Effects of enzymatic deamidation by protein-glutaminase on structure and functional properties of wheat gluten. J. Agric. Food Chem. 2006, 54, 6034–6040. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Shi, H.; Chen, X.; Xu, P.; Jiang, D.; Xu, W.; Wang, D. Modifying the structure, emulsifying and rheological properties of water-soluble protein from chicken liver by low-frequency ultrasound treatment. Int. J. Biol Macromol. 2019, 139, 810–817. [Google Scholar] [CrossRef]
- Lepedda, A.J.; Zinellu, A.; Nieddu, G.; Zinellu, E.; Carru, C.; Spirito, R.; Guarino, A.; De Muro, P.; Formato, M. Protein sulfhydryl group oxidation and mixed-disulfide modifications in stable and unstable human carotid plaques. Oxidative Med. Cell. Longev. 2013, 2013, 66–67. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Crombag, J.J.R.; Senden, J.M.G.; Waterval, W.A.H.; Bierau, J.; Verdijk, L.B.; van Loon, L.J.C. Protein content and amino acid composition of commercially available plant-based protein isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.G.; Zhao, M.; Wang, S.; Cui, C. Proteolytic degradation and amino acid liberation during extensive hydrolysis of porcine blood hemoglobin by protease admixture. J. Food Process Eng. 2007, 30, 640–659. [Google Scholar] [CrossRef]
- Cho, M.J.; Unklesbay, N.; Hsieh, F.H.; Clarke, A.D. Hydrophobicity of bitter peptides from soy protein hydrolysates. J. Agric. Chem. 2004, 52, 5895–5901. [Google Scholar] [CrossRef]
- Ney, K.H. Bitterness of peptides: Amino acid composition and chain length. Acs Symp. Ser. 1979, 115, 149–173. [Google Scholar] [CrossRef]
- Noman, A.; Xu, Y.; Al-Bukhaiti, W.Q.; Abed, S.M.; Ali, A.H.; Ramadhan, A.H.; Xia, W. Influence of enzymatic hydrolysis conditions on the degree of hydrolysis and functional properties of protein hydrolysate obtained from Chinese sturgeon (Acipenser sinensis) by using papain enzyme. Process. Biochem. 2018, 67, 19–28. [Google Scholar] [CrossRef]
- Malik, M.A.; Saini, C.S. Polyphenol removal from sunflower seed and kernel: Effect on functional and rheological properties of protein isolates. Food Hydrocoll. 2017, 63, 705–715. [Google Scholar] [CrossRef]
- Tang, S.; Hettiarachchy, N.S.; Horax, R.; Eswaranandam, S. Physicochemical properties and functionality of rice bran protein hydrolyzate prepared from heat-stabilized defatted rice bran with the aid of enzymes. J. Food Sci. 2003, 68, 152–157. [Google Scholar] [CrossRef]
- Hettiarachchy, N.S.; Ziegler, G.R. Ift Basic Symposium Series. In Protein Functionality in Food Systems; Marcel Dekker, Inc.: New York, NY, USA, 1994. [Google Scholar]
- Mokni Ghribi, A.; Maklouf Gafsi, I.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of enzymatic hydrolysis on conformational and functional properties of chickpea protein isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef]
- Xia, Y.; Bamdad, F.; Ganzle, M.; Chen, L. Fractionation and characterization of antioxidant peptides derived from barley glutelin by enzymatic hydrolysis. Food Chem. 2012, 134, 1509–1518. [Google Scholar] [CrossRef]
- Severin, S.; Xia, W.S. Enzymatic hydrolysis of whey proteins by two different proteases and their effect on the functional properties of resulting protein hydrolysates. J. Food Biochem. 2006, 30, 77–97. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Flavourzyme | Protein | Neutrase | ||||||
| α-Helix (%) | β-Sheet (%) | β-Turn (%) | Random (%) | α-Helix (%) | β-Sheet (%) | β-Turn (%) | Random (%) | ||
| PP | 15.0 | 28.8 | 22.1 | 34.2 | PP | 15.0 | 28.8 | 22.1 | 34.2 |
| 2% | 20.1 | 24.4 | 19.6 | 35.9 | 2% | 21.2 | 23.5 | 20.4 | 34.9 |
| 4% | 20.5 | 23.8 | 19.8 | 35.9 | 4% | 22.5 | 22.8 | 19.0 | 35.7 |
| 6% | 21.6 | 22.4 | 19.8 | 36.2 | 6% | 23.0 | 22.2 | 18.2 | 36.6 |
| 7% | 22.2 | 21.7 | 19.7 | 36.4 | 8% | 21.2 | 23.5 | 20.4 | 34.9 |
| Protein | Alcalase | Protein | Trypsin | ||||||
| α-Helix (%) | β-Sheet (%) | β-Turn (%) | Random (%) | α-Helix (%) | β-Sheet (%) | β-Turn (%) | Random (%) | ||
| PP | 15.0 | 28.8 | 22.1 | 34.2 | PP | 15.0 | 28.8 | 22.1 | 34.2 |
| 2% | 24.5 | 21.7 | 19.5 | 34.3 | 2% | 25.9 | 20.8 | 19.1 | 34.2 |
| 4% | 24.7 | 21.4 | 19.5 | 34.4 | 4% | 27.9 | 19.3 | 19.0 | 33.8 |
| 6% | 25.6 | 20.3 | 19.0 | 35.1 | 6% | 29.2 | 16.9 | 19.9 | 34.0 |
| 8% | 26.9 | 18.5 | 18.3 | 36.3 | 8% | 32.1 | 14.1 | 19.0 | 34.8 |
| Amino Acid | Flavourzyme | Neutrase | Alcalase | Trypsin | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2% | 4% | 6% | 7% | 2% | 4% | 6% | 8% | 2% | 4% | 6% | 8% | 2% | 4% | 6% | 8% | |
| ASP | 7.79 ± 0.01 a | 8.66 ± 0.02 c | 8.09 ± 0.00 b | 8.61 ± 0.02 c | 9.09 ± 0.02 a | 9.17 ± 0.01 b | 9.58 ± 0.03 c | 9.70 ± 0.01 d | 10.24 ± 0.03 c | 10.02 ± 0.01 a | 10.13 ± 0.02 b | 10.18 ± 0.03 b | 9.58 ± 0.01 a | 9.70 ± 0.02 b | 9.58 ± 0.04 a | 9.73 ± 0.02 b |
| THR | 2.05 ± 0.01 a | 2.25 ± 0.00 d | 2.18 ± 0.01 c | 2.07 ± 0.02 b | 2.76 ± 0.02 a | 2.89 ± 0.00 b | 3.06 ± 0.01 c | 3.10 ± 0.01 d | 3.05 ± 0.01 a | 3.05 ± 0.03 a | 3.04 ± 0.01 a | 3.24 ± 0.02 b | 3.11 ± 0.00 b | 3.06 ± 0.02 a | 3.13 ± 0.01 b | 3.18 ± 0.03 c |
| SER | 3.36 ± 0.00 c | 3.38 ± 0.01 c | 2.98 ± 0.02 b | 2.67 ± 0.00 a | 3.85 ± 0.01 a | 3.83 ± 0.02 a | 3.93 ± 0.01 b | 4.04 ± 0.02 c | 4.01 ± 0.01 c | 3.86 ± 0.02 a | 3.82 ± 0.01 a | 3.95 ± 0.02 b | 4.05 ± 0.03 c | 3.92 ± 0.01 a | 3.93 ± 0.01 a | 3.98 ± 0.01 b |
| GLU | 18.59 ± 0.05 c | 18.99 ± 0.03 d | 18.05 ± 0.06 b | 17.39 ± 0.03 a | 16.73 ± 0.03 d | 15.89 ± 0.02 a | 16.25 ± 0.02 b | 16.62 ± 0.01 c | 18.52 ± 0.02 d | 17.52 ± 0.03 c | 16.49 ± 0.01 a | 16.83 ± 0.04 b | 16.99 ± 0.01 d | 15.90 ± 0.02 c | 15.70 ± 0.01 a | 15.78 ± 0.02 b |
| PRO | 2.68 ± 0.02 a | 3.00 ± 0.01 b | 3.13 ± 0.01 c | 3.18 ± 0.01 d | 3.19 ± 0.02 a | 3.25 ± 0.01 b | 3.36 ± 0.01 c | 3.40 ± 0.00 d | 3.44 ± 0.01 a | 3.45 ± 0.02 a | 3.50 ± 0.00 b | 3.63 ± 0.01 c | 3.42 ± 0.02 a | 3.42 ± 0.01 a | 3.43 ± 0.00 a | 3.44 ± 0.01 a |
| GLY | 2.75 ± 0.01 a | 2.90 ± 0.00 b | 2.89 ± 0.02 b | 2.92 ± 0.01 b | 2.95 ± 0.02 a | 2.99 ± 0.00 b | 3.08 ± 0.01 c | 3.12 ± 0.01 d | 3.22 ± 0.00 c | 3.17 ± 0.02 b | 3.09 ± 0.01 a | 3.31 ± 0.01 d | 3.15 ± 0.00 b | 3.09 ± 0.01 a | 3.13 ± 0.02 b | 3.16 ± 0.01 b |
| ALA | 2.55 ± 0.00 a | 2.82 ± 0.02 b | 2.89 ± 0.00 c | 2.98 ± 0.01 d | 3.08 ± 0.01 a | 3.15 ± 0.01 b | 3.29 ± 0.01 c | 3.31 ± 0.01 c | 3.30 ± 0.01 a | 3.33 ± 0.01 a | 3.39 ± 0.01 b | 3.45 ± 0.01 c | 3.28 ± 0.01 a | 3.35 ± 0.01 b | 3.36 ± 0.01 b | 3.39 ± 0.02 b |
| VAL | 2.94 ± 0.01 a | 3.24 ± 0.01 b | 3.33 ± 0.03 c | 3.54 ± 0.02 d | 3.45 ± 0.01 a | 3.56 ± 0.01 b | 3.67 ± 0.02 c | 3.81 ± 0.02 d | 3.68 ± 0.02 a | 3.71 ± 0.01 a | 3.72 ± 0.01 a | 3.84 ± 0.02 b | 3.64 ± 0.01 a | 3.70 ± 0.02 b | 3.74 ± 0.00 c | 3.77 ± 0.01 d |
| MET | 0.05 ± 0.00 a | 0.19 ± 0.01 b | 0.22 ± 0.01 b | 0.25 ± 0.00 c | 0.33 ± 0.01 a | 0.40 ± 0.01 b | 0.42 ± 0.00 b | 0.45 ± 0.01 c | 0.34 ± 0.01 a | 0.36 ± 0.00 a | 0.43 ± 0.00 b | 0.49 ± 0.01 c | 0.42 ± 0.01 a | 0.45 ± 0.01 a | 0.46 ± 0.01 a | 0.54 ± 0.00 b |
| ILE | 2.76 ± 0.02 a | 3.03 ± 0.03 b | 3.16 ± 0.01 c | 3.33 ± 0.01 d | 3.19 ± 0.01 a | 3.32 ± 0.02 b | 3.42 ± 0.01 c | 3.46 ± 0.02 c | 3.40 ± 0.01 a | 3.45 ± 0.02 b | 3.46 ± 0.01 b | 3.61 ± 0.02 c | 3.31 ± 0.02 a | 3.33 ± 0.01 a | 3.36 ± 0.01 b | 3.40 ± 0.01 c |
| LEU | 4.72 ± 0.01 a | 5.43 ± 0.01 b | 5.63 ± 0.03 c | 5.81 ± 0.01 d | 5.75 ± 0.02 a | 5.89 ± 0.01 b | 6.09 ± 0.02 c | 6.17 ± 0.01 d | 6.15 ± 0.01 a | 6.19 ± 0.02 a | 6.25 ± 0.02 b | 6.34 ± 0.02 c | 6.14 ± 0.01 a | 6.15 ± 0.03 a | 6.31 ± 0.02 b | 6.44 ± 0.01 c |
| TYR | 1.32 ± 0.01 a | 1.56 ± 0.00 b | 1.69 ± 0.01 c | 1.87 ± 0.00 d | 1.83 ± 0.00 a | 2.03 ± 0.01 b | 2.17 ± 0.01 c | 2.19 ± 0.01 c | 2.17 ± 0.00 a | 2.34 ± 0.01 b | 2.36 ± 0.00 b | 2.54 ± 0.01 c | 2.37 ± 0.01 a | 2.41 ± 0.01 b | 2.57 ± 0.01 c | 2.61 ± 0.02 d |
| PHE | 2.91 ± 0.01 a | 3.80 ± 0.02 b | 3.90 ± 0.00 c | 4.03 ± 0.03 d | 3.92 ± 0.01 a | 4.03 ± 0.01 b | 4.18 ± 0.01 c | 4.20 ± 0.01 c | 4.08 ± 0.01 a | 4.15 ± 0.01 b | 4.16 ± 0.01 b | 4.30 ± 0.01 c | 4.10 ± 0.00 a | 4.13 ± 0.01 b | 4.20 ± 0.01 c | 4.25 ± 0.01 d |
| LYS | 5.61 ± 0.03 b | 5.79 ± 0.01 c | 5.63 ± 0.03 b | 5.06 ± 0.02 a | 5.99 ± 0.02 a | 5.95 ± 0.01 a | 6.21 ± 0.02 b | 6.21 ± 0.01 b | 6.42 ± 0.02 c | 6.29 ± 0.01 b | 5.99 ± 0.02 a | 6.26 ± 0.03 b | 6.08 ± 0.02 c | 5.83 ± 0.03 a | 5.83 ± 0.01 a | 5.94 ± 0.00 b |
| HIS | 1.01 ± 0.00 b | 1.16 ± 0.01 c | 0.92 ± 0.00 a | 0.90 ± 0.02 a | 1.13 ± 0.00 b | 1.12 ± 0.01 b | 1.13 ± 0.00 b | 1.07 ± 0.00 a | 1.07 ± 0.00 b | 1.09 ± 0.01 b | 1.04 ± 0.01 a | 1.12 ± 0.01 b | 1.07 ± 0.00 a | 1.09 ± 0.01 b | 1.06 ± 0.00 a | 1.10 ± 0.01 b |
| ARG | 6.13 ± 0.03 c | 6.45 ± 0.03 d | 5.44 ± 0.01 b | 4.98 ± 0.02 a | 7.26 ± 0.02 b | 7.16 ± 0.02 a | 7.39 ± 0.01 c | 7.49 ± 0.01 d | 7.65 ± 0.02 c | 7.42 ± 0.03 b | 7.09 ± 0.01 a | 6.41 ± 0.02 b | 7.47 ± 0.01 c | 7.11 ± 0.00 b | 7.07 ± 0.01 a | 7.11 ± 0.03 ab |
| HAA | 19.92 ± 0.03 a | 23.08 ± 0.02 b | 23.94 ± 0.06 c | 24.99 ± 0.02 d | 24.73 ± 0.05 a | 25.62 ± 0.03 b | 26.59 ± 0.01 c | 26.99 ± 0.02 d | 26.58 ± 0.05 a | 26.99 ± 0.05 b | 27.61 ± 0.01 c | 28.18 ± 0.03 d | 26.50 ± 0.03 a | 27.13 ± 0.02 b | 27.43 ± 0.03 c | 27.83 ± 0.05 d |
| Q value | 1039.94 ± 1.21 a | 1074.08 ± 0.86 b | 1108.16 ± 1.11 c | 1126.82 ± 2.01 d | 1089.13 ± 1.01 a | 1101.133 ± 2.11 b | 1104.35 ± 1.33 b | 1108.58 ± 0.56 c | 1083.81 ± 1.51 a | 1098.21 ± 2.11 b | 1104.91 ± 0.96 c | 1111.09 ± 1.71 d | 1094.78 ± 1.44 a | 1105.858 ± 1.66 b | 1110.83 ± 1.13 c | 1112.40 ± 0.56 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shuai, X.; Gao, L.; Geng, Q.; Li, T.; He, X.; Chen, J.; Liu, C.; Dai, T. Effects of Moderate Enzymatic Hydrolysis on Structure and Functional Properties of Pea Protein. Foods 2022, 11, 2368. https://doi.org/10.3390/foods11152368
Shuai X, Gao L, Geng Q, Li T, He X, Chen J, Liu C, Dai T. Effects of Moderate Enzymatic Hydrolysis on Structure and Functional Properties of Pea Protein. Foods. 2022; 11(15):2368. https://doi.org/10.3390/foods11152368
Chicago/Turabian StyleShuai, Xixiang, Lizhi Gao, Qin Geng, Ti Li, Xuemei He, Jun Chen, Chengmei Liu, and Taotao Dai. 2022. "Effects of Moderate Enzymatic Hydrolysis on Structure and Functional Properties of Pea Protein" Foods 11, no. 15: 2368. https://doi.org/10.3390/foods11152368
APA StyleShuai, X., Gao, L., Geng, Q., Li, T., He, X., Chen, J., Liu, C., & Dai, T. (2022). Effects of Moderate Enzymatic Hydrolysis on Structure and Functional Properties of Pea Protein. Foods, 11(15), 2368. https://doi.org/10.3390/foods11152368