Upstream Regulator Analysis of Wooden Breast Myopathy Proteomics in Commercial Broilers and Comparison to Feed Efficiency Proteomics in Pedigree Male Broilers

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Wooden Breast Myopathy Samples in Commercial Broilers
2.3. Breast Muscle Samples in Pedigree Male Broilers
2.4. Protein Extraction
2.5. Shotgun Proteomics
2.6. Upstream Regulator Analysis
3. Results and Discussion
3.1. Phenotypic Data for Birds in Wooden Breast and Feed Efficiency Studies
3.2. Proteomic Dataset Comparison
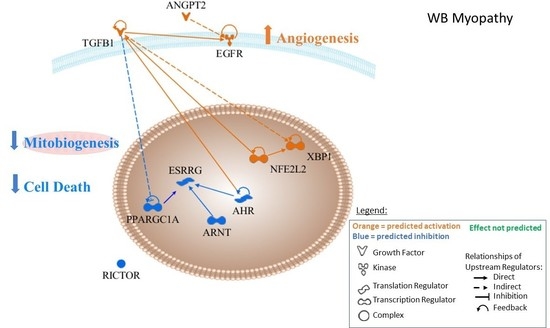
3.3. Upstream Regulators and Functional Analysis
3.4. Upstream Regulators
3.4.1. Angiopoeitin 2
3.4.2. Epidermal Growth Factor Receptor
3.4.3. Transforming Growth Factor Beta 1 (TGFB1)
3.4.4. Nuclear Factor (Erythroid-Derived 2)-Like 2 (NFE2L2)
3.4.5. X-Box Binding Protein 1
3.4.6. Rapamycin-Insensitive Companion of Mammalian Target of Rapamycin (RICTOR)
3.4.7. Peroxisome Proliferator-Activating Gamma Coactivator 1 Alpha (PPARGC1α or PGC1α)
3.4.8. Aryl Hydrocarbon Receptor Nuclear Translocator (ARNT) and Aryl Hydrocarbon Receptor Nuclear (AHR)
3.4.9. Estrogen Regulator Receptor Gamma (ERSSγ)
3.5. Summary and Synopsis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Velleman, S.G. Relationship of Skeletal Muscle Development and Growth to Breast Muscle Myopathies: A Review. Avian Dis. 2015, 59, 525–531. [Google Scholar] [CrossRef]
- Kuttappan, V.A.; Bottje, W.; Ramnathan, R.; Hartson, S.D.; Coon, C.N.; Kong, B.-W.; Owens, C.M.; Vazquez-Añon, M.; Hargis, B.M. Proteomic analysis reveals changes in carbohydrate and protein metabolism associated with broiler breast myopathy. Poult. Sci. 2017, 96, 2992–2999. [Google Scholar] [CrossRef] [PubMed]
- Lake, J.A.; Abasht, B. Glucolipotoxicity: A Proposed Etiology for Wooden Breast and Related Myopathies in Commercial Broiler Chickens. Front. Physiol. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Velleman, S.G.; Clark, K.L. Histopatholoic and myogenic gene expression changes associated with wooden breast in broiler breast muscle. Avian Dis. 2015, 59, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Abasht, B.; Mutryn, M.F.; Mickalek, R.D.; Lee, W.R. Oxidative stress and metabolic perturbations in wooden breast disorder in chickens. PLoS ONE 2016, 11, e0153750. [Google Scholar] [CrossRef]
- Papah, M.B.; Brannick, E.M.; Schmidt, C.J.; Abasht, B. Evidence and role of phlebitis and lipid infiltration in the onset and pathogenesis of Wooden Breast Disease in modern broiler chickens. Avian Pathol. 2017, 46, 623–643. [Google Scholar] [CrossRef]
- Sihvo, H.-K.; Lindén, J.; Airas, N.; Immonen, K.; Valaja, J.; Puolanne, E. Wooden Breast Myodegeneration of Pectoralis Major Muscle over the Growth Period in Broilers. Vet. Pathol. 2017, 54, 119–128. [Google Scholar] [CrossRef]
- Brothers, B.; Zhuo, Z.; Papah, M.B.; Abasht, B. RNA-Seq Analysis Reveals Spatial and Sex Differences in Pectoralis Major Muscle of Broiler Chickens Contributing to Difference in Susceptibility to Wooden Breast Disease. Front. Physiol. 2019, 10, 764. [Google Scholar] [CrossRef]
- Petracci, M.; Soglia, F.; Madruga, M.; Carvalho, L.; Ida, E.; Estévez, M. Wooden-Breast, White Striping, and Spaghetti Meat: Causes, Consequences and Consumer Perception of Emerging Broiler Meat Abnormalities. Compr. Rev. Food Sci. Food Saf. 2019, 18, 565–583. [Google Scholar] [CrossRef]
- Gratta, F.; Fasolato, L.; Birolo, M.; Zomeño, C.; Novelli, E.; Massimiliano, P.; Pascual, A.; Xiccato, G.; Trocino, A. Effect of breast myopathies on quality and microbial shelf life of broiler meat. Poult. Sci. 2019, 98, 2641–2651. [Google Scholar] [CrossRef]
- Kong, B.; Lassiter, K.; Piekarski-Welsher, A.; Dridi, S.; Reverter-Gomez, A.; Hudson, N.J.; Bottje, W.G. Proteomics of breast muscle tissue associated with the phenotypic expression of feed efficiency within a pedigree male broiler line: I. Highlight on mitochondria. PLoS ONE 2016, 11, e0155679. [Google Scholar] [CrossRef]
- Tijare, V.; Yang, V.; Kuttappan, V.; Alvarado, C.; Coon, C.; Owens, C. Meat quality of broiler breast fillets with white string and woody breast muscle myopathies. Poult. Sci. 2016, 95, 2167–2173. [Google Scholar] [CrossRef] [PubMed]
- Bottje, W.; Brand, M.D.; Ojano-Dirain, C.; Lassiter, K.; Toyomizu, M.; Wing, T. Mitochondrial proton leak kinetics and relationship with feed efficiency within a single genetic line of male broilers. Poult. Sci. 2009, 88, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Voruganti, S.; Lacroix, J.C.; Rogers, C.N.; Rogers, J.; Matts, R.L.; Hartson, S. The Anticancer Drug AUY922 Generates a Proteomics Fingerprint That Is Highly Conserved among Structurally Diverse Hsp90 Inhibitors. J. Proteome Res. 2013, 12, 3697–3706. [Google Scholar] [CrossRef] [PubMed]
- Mutryn, M.F.; Brannick, E.M.; Fu, W.; Lee, W.R.; Abasht, B. Characterization of a novel chicken muscle disorder through differential gene expression and pathway analysis using RNA-sequencing. BMC Genom. 2015, 16, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Mofarrahi, M.; Husain, S.N.A. Expression and Functional Roles of Angiopoietin-2 in Skeletal Muscles. PLoS ONE 2011, 6, e22882. [Google Scholar] [CrossRef]
- Reverter, A.; Okimoto, R.; Sapp, R.; Bottje, W.G.; Hawken, R.; Hudson, N.J. Chicken muscle mitochondrial content appears co-ordinately regulated and is associated with performance phenotypes. Biol. Open 2017, 6, 50–58. [Google Scholar] [CrossRef]
- Olwin, B.B.; Haushka, S.D. Cell surface fibroblast growth factor and epidermal growth factor receptors are permanently lost during skeletal muscle terminal differentiation in culture. J. Cell Biol. 1988, 107, 761–769. [Google Scholar] [CrossRef]
- Peng, M.; Palin, M.-F.; Véronneau, S.; Lebel, D.; Pelletier, G. Ontogeny of epidermal growth factor (EGF), EGF receptor (EGFR) and basic fibroblast growth factor (bFGF) mRNA levels in pancreas, liver, kidney, and skeletal muscle of pig. Domest. Anim. Endocrinol. 1997, 14, 286–294. [Google Scholar] [CrossRef]
- Leroy, M.C.; Perroud, J.; Darbellay, B.; Bernheim, L.; Konig, S. Epidermal Growth Factor Receptor Down-Regulation Triggers Human Myoblast Differentiation. PLoS ONE 2013, 8, e71770. [Google Scholar] [CrossRef]
- Van Obberghen-Schilling, E.; Roche, N.S.; Flanders, K.C.; Sporn, M.B.; Roberts, A.B. Transforming growth factor B1 positively regulates its own expression in normal and transformed cells. J. Biol. Chem. 1988, 263, 7741–7746. [Google Scholar] [CrossRef]
- Hubert, S.M.; Williams, T.J.; Athrey, G. Insights into the molecular basis of wooden breast based on comparative analysis of fast-and slow-growth broilers. bioRxiv 2018, 356683. [Google Scholar] [CrossRef]
- Casalena, G.; Daehn, I.; Bottinger, E.P. Transforming Growth Factor-β, Bioenergetics, and Mitochondria in Renal Disease. Semin. Nephrol. 2012, 32, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Sakairi, T.; Beeson, C.; Kopp, J.B. TGF-beta1 stimulates mitochondrial oxidative phosphorylation and generation of reactive oxygen species in cultured mouse podocytes, mediated in part by the mTOR pathway. Am. J. Physiol. Ren. Physiol. 2013, 305, F1477–F1490. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Rivera, S.; Monclus, E.A.; Synenki, L.; Zirk, A.; Eisenbart, J.; Feghali-Bostwick, C.; Mutlu, G.M.; Budinger, G.R.; Chandel, N.S. Mitochondrial reactive oxygen species regulate transforming growth factor-beta signaling. J. Biol. Chem. 2013, 288, 770–777. [Google Scholar] [CrossRef]
- Liu, D.; Black, B.L.; Derynck, R. TGF-β inhibits muscle differentiation through functional repression of myogenic transcription factors by Smad3. Genes Dev. 2001, 15, 2950–2966. [Google Scholar] [CrossRef]
- Negmadjanov, U.; Godic, Z.; Rizvi, F.; Emelyanova, L.; Ross, G.; Richards, J.; Holmuhamedov, E.; Jahangir, A. TGF-β1-Mediated Differentiation of Fibroblasts Is Associated with Increased Mitochondrial Content and Cellular Respiration. PLoS ONE 2015, 10, e0123046. [Google Scholar] [CrossRef]
- Widmann, P.; Reverter, A.; Weikard, R.; Suhre, K.; Hammon, H.M.; Albrecht, E.; Kühn, C. Systems Biology Analysis Merging Phenotype, Metabolomic and Genomic Data Identifies Non-SMC Condensin I Complex, Subunit G (NCAPG) and Cellular Maintenance Processes as Major Contributors to Genetic Variability in Bovine Feed Efficiency. PLoS ONE 2015, 10, e0124574. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, P.A.; Naval-Sanchez, M.; Porto-Neto, L.R.; Ferraz, J.B.S.; Reverter, A.; Fukumasu, H. Systems Biology Reveals NR2F6 and TGFB1 as Key Regulators of Feed Efficiency in Beef Cattle. Front. Genet. 2019, 10, 230. [Google Scholar] [CrossRef]
- Shelton, L.M.; Park, B.K.; Copple, I.M. Role of Nrf2 in protection against acute kidney injury. Kidney Int. 2013, 84, 1090–1095. [Google Scholar] [CrossRef]
- Chen, Q.M.; Maltagliati, A.J. Nrf2 at the heart of oxidative stress and cardiac protection. Physiol. Genom. 2018, 50, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kang, M.-I.; Watai, Y.; Tong, K.I.; Shibata, T.; Uchida, K.; Yamamoto, M. Oxidative and Electrophilic Stresses Activate Nrf2 through Inhibition of Ubiquitination Activity of Keap1. Mol. Cell. Biol. 2006, 26, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Lee, W.R.; Abasht, B. Messenger RNA sequencing and pathway analysis provide novel insights into the biological basis of chickens’ feed efficiency. BMC Genom. 2015, 16, 195. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, M.; Xiong, Y. BTB Protein Keap1 Targets Antioxidant Transcription Factor Nrf2 for Ubiquitination by the Cullin 3-Roc1 Ligase. Mol. Cell. Biol. 2005, 25, 162–171. [Google Scholar] [CrossRef]
- Maher, J.; Yamamoto, M. The rise of antioxidant signaling—The evolution and hormetic actions of Nrf2. Toxicol. Appl. Pharmacol. 2010, 244, 4–15. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Abramov, A.Y. The emerging role of Nrf2 in mitochondrial function. Free. Radic. Biol. Med. 2015, 88, 179–188. [Google Scholar] [CrossRef]
- Acosta-Alvear, D.; Zhou, Y.; Blais, A.; Tsikitis, M.; Lents, N.H.; Arias, C.; Lennon, C.J.; Kluger, Y.; Dynlacht, B.D. XBP1 Controls Diverse Cell Type- and Condition-Specific Transcriptional Regulatory Networks. Mol. Cell 2007, 27, 53–66. [Google Scholar] [CrossRef]
- Greene, E.; Flees, J.; Dadgar, S.; Mallman, B.; Orlowski, S.; Dhamad, A.; Rochell, S.; Kidd, M.; Laurendon, C.; Whitfield, H.; et al. Quantum blue reduces the severity of muscle myopathy via modulation of oxygen homeostasis-related genes in broiler chickens. Front. Physiol. 2019, 10, 1251. [Google Scholar] [CrossRef]
- Jacinto, E.; Loewith, R.; Schmidt, A.; Lin, S.; Rüegg, M.A.; Hall, A.; Hall, M.N. Mammalian TOR complex 2 controls the actin cytoskeleton and is rapamycin insensitive. Nat. Cell Biol. 2004, 6, 1122–1128. [Google Scholar] [CrossRef]
- Kim, D.-H.; Sarbassov, D.D.; Ali, S.M.; King, J.E.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. mTOR Interacts with Raptor to Form a Nutrient-Sensitive Complex that Signals to the Cell Growth Machinery. Cell 2002, 110, 163–175. [Google Scholar] [CrossRef]
- Sarbassov, D.D.; Ali, S.M.; Kim, D.-H.; Guertin, D.A.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. Rictor, a Novel Binding Partner of mTOR, Defines a Rapamycin-Insensitive and Raptor-Independent Pathway that Regulates the Cytoskeleton. Curr. Biol. 2004, 14, 1296–1302. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Romanino, K.; Cloëtta, D.; Lin, S.; Mascarenhas, J.B.; Oliveri, F.; Xia, J.; Casanova, E.; Costa, C.F.; Brink, M.; et al. Skeletal muscle-specific ablation of raptor, but not of rictor, causes metabolic changes and results in muscle dystrophy. Cell Metab. 2008, 8, 411–424. [Google Scholar] [CrossRef]
- Nisoli, E.; Clementi, E.; Moncada, S.; Carruba, M.O. Mitochondrial biogenesis as a cellular signaling framework. Biochem. Pharmacol. 2004, 67, 1–15. [Google Scholar] [CrossRef]
- Nisoli, E.; Clementi, E.; Paolucci, C.; Cozzi, V.; Tonello, C.; Sciorati, C.; Bracale, R.; Valerio, A.; Francolini, M.; Moncada, S.; et al. Mitochondrial Biogenesis in Mammals: The Role of Endogenous Nitric Oxide. Science 2003, 299, 896–899. [Google Scholar] [CrossRef]
- Hosotani, M.; Kawasaki, T.; Hasegawa, Y.; Wakasa, Y.; Hoshino, M.; Takahashi, N.; Ueda, H.; Takaya, T.; Iwasaki, T.; Watanabe, T. Physiological and Pathological Mitochondrial Clearance Is Related to Pectoralis Major Muscle Pathogenesis in Broilers with Wooden Breast Syndrome. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef]
- Bottje, W.; Carstens, G.E. Association of mitochondrial function and feed efficiency in poultry and livestock species1. J. Anim. Sci. 2009, 87, E48–E63. [Google Scholar] [CrossRef]
- Papah, M.B.; Brannick, E.M.; Schmidt, C.J.; Abasht, B. Gene expression profiling of the early pathogenesis of wooden breast disease in commercial broiler chickens using RNA-sequencing. PLoS ONE 2018, 13, e0207346. [Google Scholar] [CrossRef]
- Lake, J.A.; Papah, M.B.; Abasht, B. Increased Expression of Lipid Metabolism Genes in Early Stages of Wooden Breast Links Myopathy of Broilers to Metabolic Syndrome in Humans. Genes 2019, 10, 746. [Google Scholar] [CrossRef]
- Guo, J.; Sartor, M.; Karyala, S.; Medvedovic, M.; Kann, S.; Puga, A.; Ryan, P.; Tomlinson, C.R. Expression of genes in the TGF-B signaling pathway is significantly deregulated in smooth muscle cells from aorta of aryl hydrocarbon receptor knockout mice. Toxicol. Appl. Pharmacol. 2004, 194, 76–89. [Google Scholar] [CrossRef]
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR signaling pathways and regulatory functions. Biochim. Open 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Gutierrez-Vasquez, C.; Quintana, F. Regulation of the immune response by the aryl hydrocarbon receptor. Immun. Cell Biol. 2018, 48, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; He, N.; Lin, C.S.; Wei, Z.; Hah, N.; Waizenegger, W.; Ming-Xiao, J.; Liddle, C.; Yu, R.T.; Atkins, A.R.; et al. Errg promotes angiogenesis, mitochondrial biogenesis, and oxidative remodeling in PGC1a/b-deficient mice. Cell Rep. 2018, 22, 2521–2529. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Commercial Broilers 2 | Pedigree Male Broilers 3 | ||||||
|---|---|---|---|---|---|---|---|
| Normal | Severe WB | High FE | Low FE | ||||
| Variable | (n = 5) | (n = 5) | p | Variable | (n = 4) | (n = 4) | p |
| BW Kg (52 days) | 3.2 + 0.2 | 3.8 + 0.2 | 0.031 | BW (49 days) | 3.13 + 0.08 | 3.18 + 0.03 | 0.573 |
| WB Score | 0.2 + 0.2 | 1.4 + 0.2 | 0.003 | Gain (kg/7 days) | 0.64 + 0.04 | 0.47 + 0.04 | 0.022 |
| FI (kg/7 days) | 0.99 + 0.06 | 1.03 + 0.09 | 0.689 | ||||
| FE (G:F) | 0.65 + 0.01 | 0.46 + 0.01 | <0.0001 | ||||
| Functions | Activation z-Score | p-Value of Overlap | Differentially Expressed Proteins 1 |
|---|---|---|---|
| Development of vasculature | 2.45 | 1.21 × 10−3 | ACP1, ANXA2, C3, CALR, CAPNS1, CDH13, CRYAB, CTSB, FGA, FLNA, HSPA5, IGHM, LDHA, PGK1, PKM, PPIA, RHOA, VIM, YWHAZ |
| Vasculogenesis | 2.43 | 4.46 × 10−4 | ACP1, ANXA2, C3, CALR, CAPNS1, CDH13, CRYAB, CTSB, FLNA, HSPA5, LDHA, PKM, PPIA, RHOA, VIM, YWHAZ |
| angiogenesis | 2.30 | 1.67 ×10−3 | ACP1, ANXA2, C3, CALR, CAPNS1, CDH13, CRYAB, CTSB, FLNA, HSPA5, LDHA, PGK1, PKM, PPIA, RHOA, VIM, YWHAZ |
| Necrosis | −3.33 | 2.68 × 10−11 | ACP1, ANXA2, C3, CALR, CAPNS1, CBR1, CRYAB, CS, CTSB, DDX3X, DYNC1H1, EEF1A1, EIF3L, FDPS, FGA, FLNA, GAPDH, GLO1, HINT1, HK1, HNRNPH1, HSP90AA1, HSP90B1, HSPA5, IGHM,KLHL40, LDHA, P4HB, PARK7, PDCD6IP, PDIA3, PEBP1, PKM, POSTN, PPIA, PRDX6, PSMC2, RACK1, RAN, RHOA, RPL11, RPL6, RPLP0, RPS11, RPS14, RPS24, RPS3, Rps3a1, RPS6, RPS7, TAGLN2, UBE2V2, VIM, YWHAE, YWHAZ |
| Cell death | −3.05 | 2.01 × 10−9 | ACP1, AK1, ALDOC, ANXA2, C3, CALR, CAPNS1, CBR1, CRYAB, CS, CTSB, DDX3X, DES, DYNC1H1, EEF1A1, EIF3L, FDPS, FGA, FLNA, GAPDH, GLO1, HINT1, HK1, HNRNPH1, HSP90AA1, HSP90B1, HSPA2, HSPA5, IGHM, KLHL40, LDHA, P4HB, PARK7, PDCD6IP, PDIA3, PEBP1, PKM, POSTN, PPIA, PRDX6, PSMC2, RAB1A, RACK1, RAN, RHOA, RPL11, RPL6, RPLP0, RPS11, RPS14, RPS24, RPS3, Rps3a1, RPS6, RPS7, TAGLN2, UBE2V2, VIM, YWHAE, YWHAZ |
| Apoptosis | −1.98 | 4.61 × 10−6 | ALDOC, ANXA2, C3, CALR, CAPNS1, CRYAB, CS, CTSB, DDX3X, DES, DYNC1H1, EEF1A1, FLNA, GAPDH, GLO1, HINT1, HK1, HNRNPH1, HSP90AA1, HSP90B1, HSPA2, HSPA5, IGHM, KLHL40, LDHA, P4HB, PARK7, PDCD6IP, PDIA3, PEBP1, PKM, PPIA, PRDX6, RACK1, RHOA, RPLP0, RPS24, RPS3, RPS6, TAGLN2, UBE2V2, VIM, YWHAE, YWHAZ |
| Upstream Regulator | Protein Name | Activation z-Score | p-Value of Overlap | Differentially Expressed Downstream Target Proteins 1 |
|---|---|---|---|---|
| ANGPT2 | Angiopoeitin 2 | 3.08 | 7.41 × 10−9 | CALR, CRYAB, HSP90AA1, HSPA2, HSPA5, P4HB, PDIA3, PDIA6, POSTN, RPS6 |
| EGFR | Epidermal growth factor | 2.62 | 1.38 × 10−3 | CRYAB, HNRNPH1, HSP90B1, HSPA5, POSTN, PPIA, VIM |
| XBP1 | X box binding protein 1 | 2.41 | 8.79 × 10−3 | CALR, HSP90B1, HSPA5, PDIA3, PDIA4, PDIA6 |
| TGFB1 | Transforming growth factor beta 1 | 2.21 | 1.95 × 10−6 | AK1, ANXA2, CAPNS1, COL7A1, CTSB, DES, EEF1A1, FLNA, FSCN1, GATD3A/GATD3B, HNT1, HNRNPH1, HSP90A1, HAPA5, IARS, IGHM, LDHA, PGK1, PLS3, POSTN, RAB1A, RACK1, RHOA, VIM, YWHAE |
| NFE2L2 | Nuclear factor (erythroid-derived 2)-like 2 | 2.18 | 4.3 × 10−9 | ARF1, CBR1, DSTN, HSP90AA1, HSP90B1, ME1, PDIA3, PDIA4, PDIA6, PGD, RACK1, RAN, RPL18, RPLPO, TPI1 |
| RICTOR | Regulatory-associated protein Independent of mTOR complex 2 | −3.61 | 2.06 × 10−9 | PSMC2, RPL11, RPL18, RPL4, RPL6, RPLP0, RPS11, RPS2, RPS24, RPS3 |
| PPARGC1A | Peroxisome proliferator activator receptor gamma-coactivator 1 α | −2.35 | 1.92 × 10−4 | AK1, C3, CS, GOT2, LDHA, ME1, PGAM1 |
| AHR | Aryl hydrocarbon receptor | −2.22 | 2.02 × 10−3 | ABCB6, CBR1, GOT1, HSP90AA1, HSPA5, IGHM, VIM |
| ARNT | Aryl hydrocarbon receptor nuclear translocator | −2.20 | 3.35 × 10−6 | ALDOC, GAPDH, IGHM, LDHA, PGK1, TPI1, VIM |
| ESRRG (NR3B3) | Estrogen related receptor gamma | −2.17 | 1.89 × 10−6 | ALDOC, GAPDH, LDHA, PKM, TPI1 |
| Upstream Regulator | WB | FE Overlay | Upstream Regulator | WB | FE Overlay |
|---|---|---|---|---|---|
| ANGPT2 | + | no | RICTOR | -- | no |
| EGFR | + | no | PPARGC1α | -- | no |
| XBP1 | + | no | AHR | -- | no |
| TGFB1 | + | + | ARNT | -- | no |
| NFE2L2 | + | + | ESRRG | -- | no |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bottje, W.G.; Lassiter, K.R.; Kuttappan, V.A.; Hudson, N.J.; Owens, C.M.; Abasht, B.; Dridi, S.; Kong, B.C. Upstream Regulator Analysis of Wooden Breast Myopathy Proteomics in Commercial Broilers and Comparison to Feed Efficiency Proteomics in Pedigree Male Broilers. Foods 2021, 10, 104. https://doi.org/10.3390/foods10010104
Bottje WG, Lassiter KR, Kuttappan VA, Hudson NJ, Owens CM, Abasht B, Dridi S, Kong BC. Upstream Regulator Analysis of Wooden Breast Myopathy Proteomics in Commercial Broilers and Comparison to Feed Efficiency Proteomics in Pedigree Male Broilers. Foods. 2021; 10(1):104. https://doi.org/10.3390/foods10010104
Chicago/Turabian StyleBottje, Walter G., Kentu R. Lassiter, Vivek A. Kuttappan, Nicholas J. Hudson, Casey M. Owens, Behnam Abasht, Sami Dridi, and Byungwhi C. Kong. 2021. "Upstream Regulator Analysis of Wooden Breast Myopathy Proteomics in Commercial Broilers and Comparison to Feed Efficiency Proteomics in Pedigree Male Broilers" Foods 10, no. 1: 104. https://doi.org/10.3390/foods10010104
APA StyleBottje, W. G., Lassiter, K. R., Kuttappan, V. A., Hudson, N. J., Owens, C. M., Abasht, B., Dridi, S., & Kong, B. C. (2021). Upstream Regulator Analysis of Wooden Breast Myopathy Proteomics in Commercial Broilers and Comparison to Feed Efficiency Proteomics in Pedigree Male Broilers. Foods, 10(1), 104. https://doi.org/10.3390/foods10010104