Abstract
Children carrying the minor allele ‘A’ at the fat mass and obesity-associated protein (FTO) gene have higher obesity prevalence. We examined the link between FTO rs9939609 polymorphism and plasma adiponectin and the mediating role of body adiposity, in a cross-sectional study comprising 323 children aged 6–11 years. Adiponectin and FTO genotypes were assessed using a commercial kit and a real-time polymerase chain reaction with high-resolution melting analysis, respectively. Body adiposity included body mass index z-score, body fat percentage and waist-to-hip ratio. To investigate adiponectin (outcome) associations with FTO and adiposity, linear regressions were implemented in additive models and across genotype categories, adjusting for sex, age and Tanner’s stage. Using mediation analysis, we determined the proportion of the association adiponectin-FTO mediated by body adiposity. Lower adiponectin concentrations were associated with one additional risk allele (βadditive = −0.075 log-μg/mL [−0.124; −0.025]), a homozygous risk genotype (βAA/TT = −0.150 [−0.253; −0.048]) and a higher body mass index z-score (β = −0.130 [−0.176; −0.085]). Similar results were obtained for body fat percentage and waist-to-hip ratio. Body adiposity may mediate up to 29.8% of the FTO-adiponectin association. In conclusion, FTO rs9939609-related differences in body adiposity may partially explain lower adiponectin concentrations. Further studies need to disentangle the biological pathways independent from body adiposity.
1. Introduction
In the last decades, overweight and obesity prevalence has increased worldwide, affecting adults, children and adolescents in developed and developing countries [1]. In Chile, childhood obesity has steadily risen since the late 1980’s [2,3]. By 2020, prevalence of childhood overweight and obesity reached up to 64%, across different school grades [4]. At all ages the life course of obesity is associated with various detrimental health effects, such as metabolic syndrome (MetS) and type 2 diabetes (T2D). It is estimated that around 60% of those suffering from obesity in their early life will show at least one metabolic alteration related to non-communicable chronic diseases (NCDs) in their adulthood, such as hypertension, dyslipidemia, insulin resistance or metabolic syndrome [5,6,7].
Obesity, a condition of excessive body fat, has been linked to lower concentrations of plasma adiponectin, a type of adipokine secreted by the adipose tissue [8]. Adiponectin has received special attention due to its pleiotropic role and beneficial effects on tissues such as skeletal muscle, liver, heart, and kidney tissues [9]. Several studies have provided evidence of adiponectin’s anti-inflammatory, anti-atherogenic [10,11] and insulin-sensitizing effects [12,13,14]. Moreover, lower plasma adiponectin has been observed among obese and T2D individuals [15,16], which has drawn more attention as a promising therapeutic target against T2D and cardio-metabolic traits.
Although there are several behavioral risk factors that contribute to obesity, genetic load plays a relevant role in the etiology of obesity. Since 2007, more than 1100 independent loci from almost 60 genome-wide association studies (GWAS) have been identified in association with several obesity traits [17]. Among them, the strongest susceptibility gene identified is the Fat Mass and Obesity (FTO)-associated gene [18]. The relationship between obesity and the minor allele ‘A’ of the FTO single-nucleotide polymorphism (SNP) rs9939609 is well established [18,19] among adults and children from diverse ethnic backgrounds, including Chileans [20,21,22,23,24,25]. For example, a study in Caucasian adults found that carriers of the A-allele had higher odds of overweight and obesity by 19% (95% CI 1.06; 1.20) and 27% (1.20; 1.34), respectively [23]. Moreover, the FTO rs9939609 risk variant has been suggested to predispose carriers to obesity-related diseases, such as T2D, and this association may be explained by body adiposity [23,26,27].
The link between the FTO rs9939609 risk variant and plasma levels of adiponectin is less clear. One study reported that adults carrying the rs9939609 SNP risk variant had no differences in adiponectin circulating levels [28], while others showed that A allele carriers had significantly lower adiponectin concentration [21,29], which was attenuated after BMI adjustment, suggesting that such association might occur through changes in body adiposity [29].
The evidence from studies in populations of children is limited. Therefore, we sought to research the association of plasma adiponectin concentration with the FTO rs9939609 genotypes (AA, TA, and AA) and measurements of general and central adiposity in Chilean children. In addition, we investigated the potential role of adiposity markers as mediators in the association between FTO rs9939609 and adiponectin.
2. Materials and Methods
2.1. Study Design and Population
This cross-sectional study included a sample of children living in the Biobío Region of Chile. Participants were 6 to 11 years old and free of any chronic disease. Participants with incomplete data on anthropometric and adiposity measures, FTO genotype and circulating adiponectin levels were excluded from the study. Out of the 361 recruited individuals, 37 participants with missing data on body fat percentage (BF%) and one individual with adiponectin blood concentration beyond five standard deviations were removed from the analyses. The final analytical sample size was n = 323.
2.2. Plasma Determinations
A sample of 4 mL of fasting peripheral blood was collected. Plasma adiponectin was determined using a commercial ELISA kit (Linco Research, St. Charles, MO, USA) and a multi-reader (Synergy 2, Biotek, Winooski, VT, USA).
2.3. Anthropometric Measurements
Data on weight (kg), height (cm), body mass index (BMI, kg/m2), waist–height index (WHtR) and BF% were collected using an anthropometrics manual [30]. Briefly, weight was assessed on light clothing and without shoes on a Tanita scale (TANITA TBF-300, TANITA, Tokyo, Japan; 1 g accuracy). Height was measured using wall-mounted stadiometers (Seca, model 208, 0.1 cm precision). The waist circumference was measured at the midpoint between the last rib and the upper border of the iliac crest with a non-elastic flexible tape (Seca, model 201, accuracy 0.1 cm). Body composition (body lean mass, body fat mass and BF%) was determined using a bioelectrical impedance analysis (TANITA TBF-300, Tokyo, Japan). The pubertal stage was established by a pediatrician according to the Tanner criteria [31]. BMI z-score was computed following the WHO definitions. Body weight was divided by height in meters squared and then normalized based on age and sex. Nutritional status was classified, as normal weight (z-score BMI > −2SD and <+1SD); overweight (z-score BMI > +1SD or <+2SD); obesity (z-score BMI > +2SD), as proposed by WHO [32].
2.4. Identification of Allelic Variants of FTO rs9939609 Polymorphism
We used the Mini Kit QIAamp DNA Blood (Qiagen GmbH, Hilden, Germany) to extract genomic DNA from leukocytes following the manufacturer’s instructions. The DNA amplification by real-time polymerase chain reaction (PCR) amplifications and the high-resolution melting analysis (HRM) were performed with the thermocycler Rotor-Gene 6500 (Corbett Research, Sydney, Australia). Briefly, 100 ng/µL of genomic DNA were incubated at 95 degrees Celsius (C) for 10 min with 3.0 mM magnesium, 12.5 µL SensiMix HRM (Quantace) reagent, 1 µL EvaGreen dye and 600 nM primers. It was followed by 40 cycles at 95 C for 15 s, 59 C for 10 s and 72 C for 10 s following a standardized protocol [33]. We based our primer selection on the work by López-Bermejo and colleagues. We used forward primer: 5′-AACTG GCTCTTGAATGAAATAGGATTCAGA-3′ and reverse primer: 5′-GTGATGCACTTGGATAGTCTCTGTTACTCT-3′ [34]. A graphic representation of PCR product sequence of 182 base pairs can be found in Supplementary Figure S1. Wild type (TT), heterozygous (TA) and homozygous for the mutation (AA) genotypes were identified using the HRM method, which allows variations in single nucleic acids to be identified by detecting small differences in the DNA melting temperature [35]. Our HRM analysis investigated melting temperatures from 70 to 85 C, with increments of 0.1 C in each PCR cycle [33]. PCR melting curves were obtained using the Rotor-Gene 6500-incorporated software and compared with the melting curves from samples with known sequence (controls), using 95% confidence interval. The melting curves of TT, TA and AA FTO genotypes are shown in Supplementary Figure S2. During HRM technique standardization, 20 random samples underwent sequencing (Faculty of Biological Sciences, Pontificia Universidad Católica de Chile) in order to confirm genotypes and be used as controls. As an example, Supplementary Figure S3 displays the sequencing data of one control sample. We carried out a 3% agarose gel electrophoresis in order to confirm the presence of a single PCR product. All samples were analysed in duplicates, with a 98% genotyping success rate.
2.5. Ethics
We adhered to the Declaration of Helsinki (1964), the Convention of the Council of Europe regarding human rights and biomedicine (1997), and the Universal Declaration on the human genome and human rights (UNESCO, 1997). Moreover, we met the requirements of the Chilean legislation in the field of biomedical research, data privacy and bioethics, according to Decree No. 114 of 2010, Law No. 20,120, and Decree update on 14 January 2013. In addition, this study protocol was approved by the Bioethics Committee of the Vice-Rectory of Research of Universidad de Concepción with number 352-2019 and date January 2019.
2.6. Statistical Analyses
The study population characteristics were presented as mean and standard deviation (SD), or as median and interquartile range (IQR) for continuous variables with normal or non-normal distribution, respectively. Absolute and relative frequency was used to describe categorical variables.
Linear regressions were used to investigate associations between circulating concentrations of adiponectin (dependent variable) and various measurements of general and central adiposity: BMI z-score, BF % and waist-to-height ratio (WHtR), as exposures (independent variables). To account for potential confounders, three statistical models were used. Model 1 was unadjusted, Model 2 was adjusted for sex, age and Tanner’s stage, and Model 3 added an adjustment for the FTO SNP rs9939609 genotype. The FTO SNP rs9939609 genotype was coded according to an additive model where the FTO genotype was coded as 0 = TT (homozygous for the wild type allele); 1 = TA (heterozygous for risk allele); 2 = AA (homozygous for the risk allele). The Chi-square test was used to estimate the Hardy–Weinberg equilibrium of the FTO alleles. Blood adiponectin concentrations were transformed to their natural logarithm to approximate normal distribution. The results were expressed in beta estimates and 95% lower and upper confidence intervals (β, 95% CI), for the variation in log-transformed adiponectin concentrations in µg/mL, per one unit increase in adiposity measures.
Next, we studied the relationship between adiponectin concentrations and the variants of the FTO SNP rs9939609 genotype in linear associations using statistical Models 1 and 2. For the independent variable, we followed two approaches: (1) as a numerical variable according to the additive model (0 = TT; 1 = AT; and 2 = AA); and (2) as genotype categories, comparing carriers of the AT or AA genotype with the TT genotype, respectively, thus using the wild type as the reference group. Further, a third model was performed with additional adjustment for a marker of general adiposity, BMI z-score. The results were expressed in beta estimates and 95% lower and upper confidence intervals (β, 95% CI), for the variation in log-transformed adiponectin, per one additional risk allele (approach 1), or among carriers of AT or AA, compared with TT carriers, respectively (approach 2).
Finally, we used model 3 to interrogate the role of body adiposity as a potential mediator in the association between adiponectin concentrations and the FTO SNP rs9939609 genotype, using mediation analysis from the mediation package in R [36]. This analysis dissects the total effect of the exposure (presence of one additional FTO rs9939609 risk allele) on the outcome (adiponectin concentration) into the direct and indirect effects. The direct effect is the part of the effect that goes directly or through mediators other than those currently studied. The indirect effect represents the portion of the effect that goes via (is mediated by) the variable under study (adiposity markers); then, the proportion mediated is quantified and expressed in percentage mediated. Further, this analysis works under the sequential ignorability assumption, which assumes no unmeasured confounding. Note that, despite that the notation in the mediation analysis uses the word ‘effect’, their results must not be interpreted as causal, given the observational and cross-sectional design of this study. Quasi-Bayesian confidence intervals were constructed for the estimated effects with 5000 simulations. Results are expressed as proportion mediated in percentage and 95% Cis. A schematic representation of the mediation analysis concept is displayed in Figure 1. All analyses were performed using R statistical software v.4.0.1 (R Foundation, Vienna, Austria).
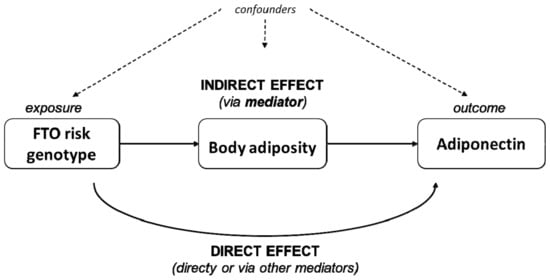
Figure 1.
Schematic representation of the causal mediation analysis. The total effect of an exposure (FTO genotype) over an outcome (concentration of circulating adiponectin) is dissected into direct and indirect effects. The indirect effect is the one being exerted via a potential mediator of interest (body adiposity). The remaining part of the effect goes directly or via mediators other than the ones under current study.
3. Results
3.1. Sample Description
The study population’s characteristics are shown in Table 1. Briefly, participants were on average 8.8 (SD ± 2.2) years old, were more often at pre-pubertal stage (77.4%) and female (50.8%), and they lived in the Biobío Region, Chile.

Table 1.
Characteristics of the study population.
3.2. Associations between Circulating Adiponectin and FTO rs9939609
Table 2 shows the inverse association between the presence of one additional risk allele at SNP rs9939609 and adiponectin concentration (βadditive= −0.075 [−0.124; −0.025], p = 0.003) controlling for age, sex and Tanner’s stage. For the FTO genotype, only the homozygous for the risk allele (AA) showed an association with adiponectin concentrations (βAA vs. TT = −0.150 [−0.253; −0.048], p = 0.004). This association remained after adjusting the analysis by BMI (β = −0.104 [−0.101; −0.004], p = 0.041) (Table 2 and Figure 2).
Table 2.
Associations between adiponectin (log-transformed, µg/mL) and several markers of general and central adiposity and the FTO rs9939609 genotype.
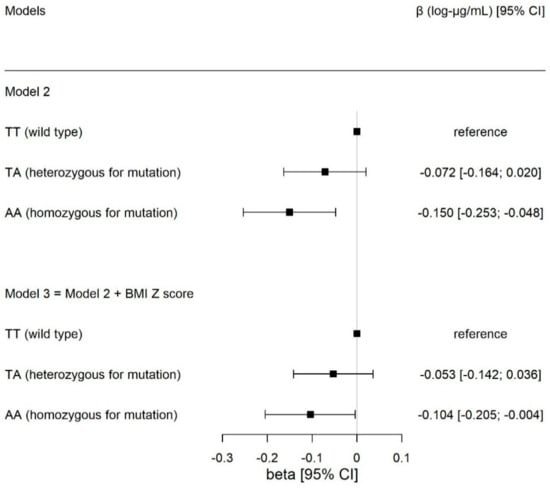
Figure 2.
Associations between circulating adiponectin concentrations in log−transformed μg/mL, with FTO genotype for SNP rs9939609, comparing carriers of the heterozygous (TA) and homozygous (AA) genotype for the mutation, with the wild-type individuals (TT, reference group), respectively. The results are expressed in beta effect estimates of and their respective 95% CIs. Model 2 was adjusted for age, sex, and Tanner’s stage. Model 3 included the adjustments in model 2 plus BMI z-score.
3.3. Link between Circulating Adiponectin and Body Adiposity
Using Model 2, we found that natural log-transformed adiponectin levels in blood were inversely associated with various measures of overall (BMI z-score and total BF %) and central obesity (WHtR), independent of sex, age and Tanner’s stage. A one-unit increase in BMI z-score and one-percent increase in total body fat were associated with lower adiponectin concentration (β = −0.130 log μg/mL [95% CI −0.176; −0.085], p = 4.63 × 10−8 and β= −0.012 [−0.016; −0.007], p = 2.38 × 10−7, respectively) (Table 2 and Figure 3). Similarly, the increase in one unit of WHtR was related to lower adiponectin (β = −1.386 [−1.959; −0.814], p = 2.91 × 10−6) (Table 2). After adjusting for the FTO SNP rs9939609, the associations remained (Table 2).
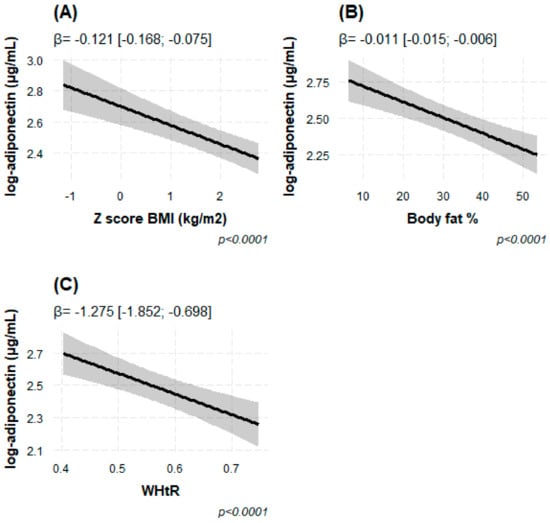
Figure 3.
(A–C) Associations of circulating adiponectin concentrations in log−transformed μg/mL, with several measures of overall and central adiposity. (A) BMI z-score, (B) total body fat % and (C) Waist to height ratio (WHtR). The statistical model was adjusted for age, sex, Tanner’s stage and FTO rs9939609 genotype. The results are expressed in beta effect estimates of and their respective 95% CIs.
3.4. Body Adiposity as Partial Mediator of the Adiponectin–FTO rs9939609 Association
Further, we investigated the potential role of general and central adiposity markers as mediators in the FTO genotype–adiponectin associations. In the mediation analysis, we observed that higher adiposity measures of BMI, WHtR and BF%, respectively, mediate a proportion from 23.9% (6.8; 68.0, Pmediation = 0.006, for WHtR) to 29.8% (10.4; 79.0, Pmediation = 0.003, for Z-score BMI) of the total effect of one additional FTO rs9939609 risk allele on plasma adiponectin levels, called the indirect effect (Table 3).
Table 3.
Mediation analysis for the association between adiponectin and FTO rs9939609 genotype, with several measures of adiposity modelled as potential mediators.
4. Discussion
In this population of Chilean children, we found associations between lower plasma adiponectin and the presence of one additional A allele at the FTO rs9939609 polymorphism, and across FTO genotypes. Moreover, inverse associations between adiponectin and measures of general and central adiposity markers (BMI Z-score, WHtR and BF%) were observed. Furthermore, we quantified the proportion of the adiponectin–FTO association explained by body adiposity differences, finding that up to 30% of the association is mediated by FTO-related levels of BMI Z-score, WHtR or BF%.
Adiponectin is an endocrine factor secreted by adipocytes [37] and displays beneficial effects on various tissues, such those related to glucose uptake and fatty acid metabolism [38,39]. Additionally, lower adiponectin concentrations are related to intermediate risk factors for T2D, such as higher blood glucose, insulin, and triglycerides [33,40].
Experimental studies show that plasmatic adiponectin and other hormones regulating food intake and satiety regulation, such as leptin and ghrelin, are associated with FTO polymorphisms [40]. Several observational studies in populations of diverse ethnic backgrounds (Caucasian, Mexican, Turkish, Indian and Chinese) have investigated the relationship between FTO rs9939609 and plasma adiponectin, reporting significant associations [20,21,22,23,24,25]. In agreement, our study in a pediatric Chilean population found a significant relationship between lower concentrations of adiponectin and the presence of one additional A allele at rs9939609, in multivariate regressions adjusted for age, sex and Tanner’s stage. Similar results were found among AA carriers, as compared to wild-type (TT) carriers, while no significant results were found when comparing TA with TT carriers. Nevertheless, our results contradict studies carried out in Tunisian [41] and Iranian [28] adults and in Romanian children [42], which failed to find significant differences in adiponectin levels across FTO rs9939609 genotypes. Moreover, a randomized controlled trial on a two-year calorie restriction revealed that the presence of the A allele did not influence adiponectin levels in response to the intervention, although they found significant associations at baseline [43] similar to other studies [44,45].
Mechanisms underlying the potential relationship FTO-adiponectin have been elucidated to a limited extent. On the one hand, lower plasma adiponectin concentration has been observed among obese or higher BMI adults [46,47] and among obese children [48,49], in line with our results. On the other hand, adiposity excess and metabolic syndrome is well known to be prevalent among carriers of the A allele at rs9939609 [18,19]. Moreover, we previously reported significant associations of rs9939609 with higher body adiposity [20], obesity [33] and MetS prevalence in Chilean children, finding the strongest effect among AA carriers [50].
The current study provides additional evidence on the role of body adiposity in the observed associations using two approaches. Firstly, additional adjustment of the association adiponectin-FTO for BMI z-score resulted in an attenuated beta estimate size, although the associations remained significant. This observation is in line with previous studies [21,22,23,24,25,26,27,28,29] which performed multivariate analysis and found attenuation of the association after accounting for body adiposity measures. Further, we implemented a more advanced method, mediation analyses, in order to compute FTO’s indirect effect on adiponectin levels, defined as the effect that goes through the mediator (body adiposity). We found that, of the total effect of FTO on adiponectin concentrations, from 23.9% (WHtR) to 29.8% (BMI z-score) is mediated by FTO-related differences in general or central body adiposity, with the largest effect attributed to the BMI z-score. Nevertheless, there is still a major part of the association that is not explained by FTO-related adiposity, but by other biological pathways beyond the scope of this study. Given the possible role of adiponectin as a promising therapeutic target against highly prevalent metabolic diseases such as T2D [51], future studies aiming to unravel mechanisms related to FTO rs9939609 regulating plasma adiponectin concentrations different from body adiposity are warranted.
Based on the available evidence, part of the unexplained effect might be determined by the gene-environment interactions. Some studies suggest that FTO polymorphism’s effect may be modulated by lifestyle factors [52], such as diet quality and physical exercise [21,53]. For example, macronutrient–gene interactions might affect obesity phenotypes, potentially by regulation of FTO and IRX3 gene expression [54]. Recent research shows that adherence to a Mediterranean dietary pattern, higher intake of unsaturated fatty acids, whole grains, polyphenols and probiotics improves the inflammatory biomarker profile, including the elevation of plasma adiponectin [55,56]. Moreover, it is well known that physical exercise triggers a number of signaling pathways stimulating the production of the bioactive molecules, adiponectin among them, that exert beneficial health effects [57]. A study reported that among individuals with the lowest physical activity levels, those carrying the A allele at rs9939609 had lower plasma adiponectin concentrations as compared to carriers of the TT genotype [21]. More studies are needed to dissect the extent to which environmental factors may contribute to attenuating the genetic predisposition to adverse metabolic health outcomes conferred by FTO risk genotypes.
Our study has some strengths. We used a well-characterized children population, including details on their pubertal stage as a biochemical marker of obesity. In addition, we used a formal mediation analysis test, an advanced statistical method to compute the proportion of the effect. Among the weaknesses of our study, we count a study population of a restricted size, although we were able to detect significant associations. Further, we used classic anthropometric and bio-impedance measures of body fat, while more accurate methods for body composition are preferable, such as dual-energy X-ray absorptiometry and computed tomography [58]. Moreover, we had limited data in lifestyle factors and other genetic polymorphisms regulating adiponectin production, such as ADIPOQ or genetic ancestry. About the latter, a previous work from our research group performed the ethnic characterization of the same study sample based on Amerindian haplogroups assessed in mitochondrial DNA [33]. The authors observed that 85% had Amerindian lineages, and that among normal-weight and obese children, the proportion of non-Amerindian individuals did not differ per group. Thus, we do not expect that genetic ancestry affects our results.
Further, the cross-sectional design of our study restrains the mediation analysis interpretation, given that the studied mediators were collected at the same time point as blood collection for adiponectin measurement. In addition, it contains some strong assumptions, such as the ignorability assumption, wherein no potential unmeasured confounding is present [36]. Although we addressed the relevant confounders, unmeasured confounding should not be ruled out. For this reason, the results from our observational study must not be interpreted as causal; however, they do provide evidence to inspire future well-powered studies and the use of causal inference methods. Finally, these results are not generalizable, and replication of the findings to other populations merits further research.
5. Conclusions
To the extent of our knowledge, this is the first study to formally suggest that the association between adiponectin and the FTO risk variant might be partly mediated by changes in body adiposity induced by FTO rs9939609 in a Chilean children population. Further research is needed to unravel the biological pathways linking SNP rs9939609 and adiponectin independently from body adiposity. More studies using accurate methods to assess body fat and a causal design are needed confirm these findings. This study enlarges the body of evidence and confirms previous findings embedded in populations with diverse ethnic backgrounds and age groups.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/children10030426/s1, Supplementary Figure S1: sequence of the polymerase chain reaction (PCR) product; Supplementary Figure S2: High Resolution Melting (HRM) curves for genetic variants of FTO rs9939609; Supplementary Figure S3: sequencing data of one of the control samples.
Author Contributions
Conceptualization, C.O.-R. and N.U.; Methodology and statistical analysis, C.O.-R.; Funding acquisition, N.U. and C.A.; Laboratory analysis, N.U. and C.A.; Writing–original draft, C.O.-R. and N.U.; Writing—review and editing, L.M., C.C.-M., M.V. and M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was conducted at Universidad de Concepción, and was funded by the grant INNOVA from the Corporación de Fomento de la Producción CORFO (Production Development Corporation), Chilean Ministry of Economy, Development and Tourism, grant number 07CN131SM-196, years 2010–2011.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Universidad de Concepción for studies involving humans.
Informed Consent Statement
Informed consent was obtained from the parent or legal guardian of all the subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We greatly appreciate the collaboration of Benilde Riffo, for providing us with valuable information on the laboratory protocols for genotyping through high-resolution melting analysis. We also thank the study participants and technical personnel for their key contribution to this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kelly, T.; Yang, W.; Chen, C.-S.; Reynolds, K.; He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. 2008, 32, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Kain, J.; Uauy, R.; Lera, L.; Taibo, M.; Espejo, F.; Albala, C. Evolution of the nutritional status of six years old Chilean children (1987–2003). Rev. Med. Chil. 2005, 133, 1013–1020. [Google Scholar] [PubMed]
- Herrera, J.C.; Lira, M.; Kain, J. Socioeconomic vulnerability and obesity in Chilean schoolchildren attending first grade: Comparison between 2009 and 2013. Rev. Chil. Pediatr. 2017, 88, 736–743. [Google Scholar] [CrossRef] [PubMed]
- JUNAEB. Mapa Nutricional 2019; Ministerio de Educación, Gobierno de Chile: Santiago, Chile, 2020.
- Allman-Farinelli, M.A.; King, L.; Bauman, A.E. Overweight and obesity from childhood to adulthood: A follow-up of participants in the 1985 Australian Schools Health and Fitness Survey. Med. J. Aust. 2007, 187, 314–315. [Google Scholar] [CrossRef]
- Eckel, R.H. Obesity and heart disease: A statement for healthcare professionals from the Nutrition Committee, American Heart Association. Circulation 1997, 96, 3248–3250. [Google Scholar] [CrossRef]
- Freedman, D.S.; Dietz, W.H.; Tang, R.; Mensah, G.; Bond, M.G.; Urbina, E.M.; Srinivasan, S.; Berenson, G.S. The relation of obesity throughout life to carotid intima-media thickness in adulthood: The Bogalusa Heart Study. Int. J. Obes. Relat. Metab. Disord 2004, 28, 159–166. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Roy, B.; Palaniyandi, S.S. Tissue-specific role and associated downstream signaling pathways of adiponectin. Cell Biosci. 2021, 11, 77. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef]
- Ohashi, K.; Ouchi, N.; Matsuzawa, Y. Anti-inflammatory and anti-atherogenic properties of adiponectin. Biochimie 2012, 94, 2137–2142. [Google Scholar] [CrossRef]
- Berner, H.S.; Lyngstadaas, S.P.; Spahr, A.; Monjo, M.; Thommesen, L.; Drevon, C.A.; Syversen, U.; Reseland, J.E. Adiponectin and its receptors are expressed in bone-forming cells. Bone 2004, 35, 842–849. [Google Scholar] [CrossRef]
- Chen, J.; Du, B. Novel positioning from obesity to cancer: FTO, an m(6)A RNA demethylase, regulates tumour progression. J. Cancer Res. Clin. Oncol. 2019, 145, 19–29. [Google Scholar] [CrossRef]
- Pajvani, U.B.; Hawkins, M.; Combs, T.P.; Rajala, M.W.; Doebber, T.; Berger, J.P.; Wagner, J.A.; Wu, M.; Knopps, A.; Xiang, A.H.; et al. Complex distribution, not absolute amount of adiponectin, correlates with thiazolidinedione-mediated improvement in insulin sensitivity. J. Biol. Chem. 2004, 279, 12152–12162. [Google Scholar] [CrossRef]
- Hakim, O.; Bello, O.; Ladwa, M.; Peacock, J.L.; Umpleby, A.M.; Charles-Edwards, G.; Amiel, S.A.; Goff, L.M. The Link between Obesity and Inflammatory Markers in the Development of Type 2 Diabetes in Men of Black African and White European Ethnicity. Nutrients 2020, 12, 3796. [Google Scholar] [CrossRef]
- Yasuda, Y.; Miyake, N.; Matsuoka, H.; Sugihara, S. Adiponectin, ALT and family history as critical markers for the development of type 2 diabetes in obese Japanese children. Endocrinol. Diabetes Metab. 2021, 4, e00178. [Google Scholar] [CrossRef]
- Loos, R.J.F.; Yeo, G.S.H. The genetics of obesity: From discovery to biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [CrossRef]
- Loos, R.J.F.; Yeo, G.S.H. The bigger picture of FTO: The first GWAS-identified obesity gene. Nat. Rev. Endocrinol. 2014, 10, 51–61. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, Y.; Sun, B.-F.; Zhao, Y.-L.; Yang, Y.-G. FTO and obesity: Mechanisms of association. Curr. Diab. Rep. 2014, 14, 486. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, N.; Villagrán, M.; Riffo, B.; Gleisner, A.; Petermann-Rocha, F.; Mardones, L.; Leiva, A.M.; Martínez-Sanguinetti, M.A.; Celis-Morales, C. Association between FTO gene rs9939609 and adiposity markers in Chilean children. Rev. Chil. Pediatr. 2020, 91, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Isgin-Atici, K.; Alsulami, S.; Turan-Demirci, B.; Surendran, S.; Sendur, S.N.; Lay, I.; Karabulut, E.; Ellahi, B.; Lovegrove, J.A.; Alikasifoglu, M.; et al. FTO gene-lifestyle interactions on serum adiponectin concentrations and central obesity in a Turkish population. Int. J. Food Sci. Nutr. 2021, 72, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Al-Serri, A.; Alroughani, R.; Al-Temaimi, R.A. The FTO gene polymorphism rs9939609 is associated with obesity and disability in multiple sclerosis patients. Sci. Rep. 2019, 9, 19071. [Google Scholar] [CrossRef]
- Andreasen, C.H.; Stender-Petersen, K.L.; Mogensen, M.S.; Torekov, S.S.; Wegner, L.; Andersen, G.; Nielsen, A.L.; Albrechtsen, A.; Borch-Johnsen, K.; Rasmussen, S.S.; et al. Low physical activity accentuates the effect of the FTO rs9939609 polymorphism on body fat accumulation. Diabetes 2008, 57, 95–101. [Google Scholar] [CrossRef]
- Fang, H.; Li, Y.; Du, S.; Hu, X.; Zhang, Q.; Liu, A.; Ma, G. Variant rs9939609 in the FTO gene is associated with body mass index among Chinese children. BMC Med. Genet. 2010, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Prakash, J.; Mittal, B.; Srivastava, A.; Awasthi, S.; Srivastava, N. Association of FTO rs9939609 SNP with Obesity and Obesity- Associated Phenotypes in a North Indian Population. Oman Med. J. 2016, 31, 99–106. [Google Scholar] [PubMed]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.B.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Rees, S.D.; Islam, M.; Hydrie, M.Z.I.; Chaudhary, B.; Bellary, S.; Hashmi, S.; O’Hare, J.P.; Kumar, S.; Sanghera, D.K.; Chaturvedi, N.; et al. An FTO variant is associated with Type 2 diabetes in South Asian populations after accounting for body mass index and waist circumference. Diabet. Med. 2011, 28, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Mehrdad, M.; Doaei, S.; Gholamalizadeh, M.; Fardaei, M.; Fararouei, M.; Eftekhari, M.H. Association of FTO rs9939609 polymorphism with serum leptin, insulin, adiponectin, and lipid profile in overweight adults. Adipocyte 2020, 9, 51–56. [Google Scholar] [CrossRef]
- Qi, L.; Kang, K.; Zhang, C.; van Dam, R.M.; Kraft, P.; Hunter, D.; Lee, C.H.; Hu, F.B. Fat mass-and obesity-associated (FTO) gene variant is associated with obesity: Longitudinal analyses in two cohort studies and functional test. Diabetes 2008, 57, 3145–3151. [Google Scholar] [CrossRef]
- Callaway, C.; Chumlea, W.; Bouchard, C.; Himes, J.; Lohman, T.; Martin, A.; Mitchell, C.D.; Mueller, W.H. Circumferences. In Anthropometric Standardization Reference Manual; Lohman, T.G., Roche, A., Martorell, R., Eds.; Human Kinetics Books: Champaign, IL, USA, 1991; pp. 44–45. [Google Scholar]
- Tanner, J.M.; Whitehouse, R.H. Clinical longitudinal standards for height, weight, height velocity, weight velocity, and stages of puberty. Arch. Dis. Child. 1976, 51, 170–179. [Google Scholar] [CrossRef]
- WHO. Growth Reference Data for 5–19 Years. 2006. Available online: https://www.who.int/tools/growth-reference-data-for-5to19-years (accessed on 22 April 2021).
- Riffo, B.; Asenjo, S.; Sáez, K.; Aguayo, C.; Muñoz, I.; Bustos, P.; Celis-Morales, C.; Lagos, J.; Sapunar, J.; Ulloa, N. FTO gene is related to obesity in Chilean Amerindian children and impairs HOMA-IR in prepubertal girls. Pediatr. Diabetes 2012, 13, 384–391. [Google Scholar] [CrossRef]
- López-Bermejo, A.; Petry, C.J.; Díaz, M.; Sebastiani, G.; de Zegher, F.; Dunger, D.B.; Ibáñez, L. The association between the FTO gene and fat mass in humans develops by the postnatal age of two weeks. J. Clin. Endocrinol. Metab. 2008, 93, 1501–1505. [Google Scholar] [CrossRef] [PubMed]
- Erali, M.; Voelkerding, K.V.; Wittwer, C.T. High resolution melting applications for clinical laboratory medicine. Exp. Mol. Pathol. 2008, 85, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Tingley, D.; Yamamoto, T.; Hirose, K.; Keele, L.; Imai, K. Mediation: R Package for Causal Mediation Analysis. J. Stat. Softw. 2014, 59, 1–38. [Google Scholar] [CrossRef]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. Compr. Physiol. 2018, 8, 1031–1063. [Google Scholar]
- Elghazy, A.M.; Elsaeid, A.M.; Refaat, M.; Youssef, M.M. Biochemical studies of adiponectin gene polymorphism in patients with obesity in Egyptians. Arch. Physiol. Biochem. 2022, 128, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, Y.; Kim, J.W.; Son, Y.-J.; Ma, M.J.; Um, J.-H.; Kim, N.D.; Min, S.H.; Kim, D.I.; Kim, B.B. Discovery of a novel potent peptide agonist to adiponectin receptor 1. PLoS ONE 2018, 13, e0199256. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.R. The ‘Fat Mass and Obesity Related’ (FTO) gene: Mechanisms of Impact on Obesity and Energy Balance. Curr. Obes. Rep. 2015, 4, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Ben Halima, M.; Kallel, A.; Baara, A.; Ben Wafi, S.; Sanhagi, H.; Slimane, H.; Jemaa, R.; Feki, M. The rs9939609 polymorphism in the fat mass and obesity associated (FTO) gene is associated with obesity in Tunisian population. Biomarkers 2018, 23, 787–792. [Google Scholar] [CrossRef]
- Duicu, C.; Mărginean, C.O.; Voidăzan, S.; Tripon, F.; Bănescu, C. FTO rs 9939609 SNP Is Associated With Adiponectin and Leptin Levels and the Risk of Obesity in a Cohort of Romanian Children Population. Medicine 2016, 95, e3709. [Google Scholar] [CrossRef]
- Dorling, J.L.; Belsky, D.W.; Racette, S.B.; Das, S.K.; Ravussin, E.; Redman, L.M.; Höchsmann, C.; Huffman, K.M.; Kraus, W.E.; Kobor, M.S.; et al. Association between the FTO rs9939609 single nucleotide polymorphism and dietary adherence during a 2-year caloric restriction intervention: Exploratory analyses from CALERIE phase 2. Exp. Gerontol. 2021, 155, 111555. [Google Scholar] [CrossRef]
- de Luis, D.A.; Aller, R.; Izaola, O.; de la Fuente, B.; Conde, R.; Sagrado, M.G.; Primo, D. Evaluation of weight loss and adipocytokines levels after two hypocaloric diets with different macronutrient distribution in obese subjects with rs9939609 gene variant. Diabetes/Metab. Res. Rev. 2012, 28, 663–668. [Google Scholar] [CrossRef]
- Zheng, Y.; Huang, T.; Zhang, X.; Rood, J.; Bray, G.A.; Sacks, F.M.; Qi, L. Dietary Fat Modifies the Effects of FTO Genotype on Changes in Insulin Sensitivity. J. Nutr. 2015, 145, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Bergmark, B.A.; Cannon, C.P.; White, W.B.; Jarolim, P.; Liu, Y.; Bonaca, M.P.; Zannad, F.; Morrow, D.A. Baseline adiponectin concentration and clinical outcomes among patients with diabetes and recent acute coronary syndrome in the EXAMINE trial. Diabetes Obes. Metab. 2017, 19, 962–969. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.W.; Lee, W.Y.; Rhee, E.J.; Baek, K.H.; Yoon, K.H.; Kang, M.I.; Yun, E.J.; Park, C.Y.; Ihm, S.H.; Choi, M.G.; et al. The relationship between serum resistin, leptin, adiponectin, ghrelin levels and bone mineral density in middle-aged men. Clin. Endocrinol. 2005, 63, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Ramon-Krauel, M.; Leal-Witt, M.J.; Osorio-Conles, Ó.; Amat-Bou, M.; Lerin, C.; Selva, D.M. Relationship between adiponectin, TNFα, and SHBG in prepubertal children with obesity. Mol. Cell Pediatr. 2021, 8, 3. [Google Scholar] [CrossRef]
- Weiss, R.; Dufour, S.; Groszmann, A.; Petersen, K.; Dziura, J.; Taksali, S.E.; Shulman, G.; Caprio, S. Low adiponectin levels in adolescent obesity: A marker of increased intramyocellular lipid accumulation. J. Clin. Endocrinol. Metab. 2003, 88, 2014–2018. [Google Scholar] [CrossRef]
- Molina-Luque, R.; Ulloa, N.; Romero-Saldaña, M.; Zilic, M.; Gleisner, A.; Lanuza, F.; Molina-Recio, G. Association between the FTO SNP rs9939609 and Metabolic Syndrome in Chilean Children. Nutrients 2021, 13, 2014. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef]
- Evans, C.; Curtis, J.; Antonio, J. FTO and Anthropometrics: The Role of Modifiable Factors. J. Funct. Morphol. Kinesiol. 2022, 7, 90. [Google Scholar] [CrossRef]
- Goodarzi, G.; Hosseini-Esfahani, F.; Ataie-Jafari, A.; Haji-Hosseini-Gazestani, N.; Daneshpour, M.S.; Keshavarz, S.-A.; Mirmiran, P. Dietary diversity modifies the association between FTO polymorphisms and obesity phenotypes. Int. J. Food Sci. Nutr. 2021, 72, 997–1007. [Google Scholar] [CrossRef]
- Doaei, S.; Kalantari, N.; Izadi, P.; Salonurmi, T.; Jarrahi, A.M.; Rafieifar, S.; Tabesh, G.A.; Rahimzadeh, G.; Gholamalizadeh, M.; Goodarzi, M.O. Interactions between macro-nutrients’ intake, FTO and IRX3 gene expression, and FTO genotype in obese and overweight male adolescents. Adipocyte 2019, 8, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Noormohammadi, M.; Ghorbani, Z.; Löber, U.; Mahdavi-Roshan, M.; Bartolomaeus, T.U.; Kazemi, A.; Shoaibinobarian, N.; Forslund, S.K. The effect of probiotic and synbiotic supplementation on appetite-regulating hormones and desire to eat: A systematic review and meta-analysis of clinical trials. Pharmacol. Res. 2023, 187, 106614. [Google Scholar] [CrossRef]
- Sánchez-Rosales, A.I.; Guadarrama-López, A.L.; Gaona-Valle, L.S.; Martínez-Carrillo, B.E.; Valdés-Ramos, R. The Effect of Dietary Patterns on Inflammatory Biomarkers in Adults with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2022, 14, 4577. [Google Scholar] [CrossRef] [PubMed]
- García-Hermoso, A.; Ramírez-Vélez, R.; Díez, J.; González, A.; Izquierdo, M. Exercise training-induced changes in exerkine concentrations may be relevant to the metabolic control of type 2 diabetes mellitus patients: A systematic review and meta-analysis of randomized controlled trials. J. Sport Health Sci. 2022; in press. [Google Scholar]
- Drolet, R.; Bélanger, C.; Fortier, M.; Huot, C.; Mailloux, J.; Légaré, D.; Tchernof, A. Fat depot-specific impact of visceral obesity on adipocyte adiponectin release in women. Obesity 2009, 17, 424–430. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).