
Are Ischemic Stroke and Alzheimer’s Disease Genetically Consecutive Pathologies?
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Alzheimer’s Disease (AD)
3.2. Ischemic Stroke (IS) and AD
3.3. Genome-Wide Association Studies of IS
3.4. GWAS of AD
3.5. Crosstalk between GWAS of IS and AD
3.6. Transcriptomics of IS
3.7. Transcriptomics of AD
3.8. Crosstalk between Transcriptomic Data of IS and AD
3.9. MicroRNAs in IS and AD
3.10. Circular RNAs in IS and AD
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Igarashi, M.; Okuyama, K.; Ueda, N.; Sano, H.; Takahashi, K.; P Qureshi, Z.; Tokita, S.; Ogawa, A.; Okumura, Y.; Okuda, S. Incremental Medical Cost of Delirium in Elderly Patients with Cognitive Impairment: Analysis of a Nationwide Administrative Database in Japan. BMJ Open 2022, 12, e062141. [Google Scholar] [CrossRef] [PubMed]
- Smid, J.; Studart-Neto, A.; César-Freitas, K.G.; Dourado, M.C.N.; Kochhann, R.; Barbosa, B.J.A.P.; Schilling, L.P.; Balthazar, M.L.F.; Frota, N.A.F.; de Souza, L.C.; et al. Declínio Cognitivo Subjetivo, Comprometimento Cognitivo Leve e Demência—Diagnóstico Sindrômico: Recomendações Do Departamento Científico de Neurologia Cognitiva e Do Envelhecimento Da Academia Brasileira de Neurologia. Dement. Neuropsychol. 2022, 16, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Patil, C.G. Epidemiology and the Global Burden of Stroke. World Neurosurg. 2011, 76, S85–S90. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Stroke Collaborators. Global, Regional, and National Burden of Stroke and Its Risk Factors, 1990–2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet Neurol 2021, 12, 941–948. [Google Scholar] [CrossRef]
- Tahami Monfared, A.A.; Byrnes, M.J.; White, L.A.; Zhang, Q. Alzheimer’s Disease: Epidemiology and Clinical Progression. Neurol. Ther. 2022, 11, 553–569. [Google Scholar] [CrossRef]
- Kalaria, R.N.; Ballard, C. Stroke and Cognition. Curr. Atheroscler. Rep. 2001, 3, 334–339. [Google Scholar]
- Seshadri, S.; Beiser, A.; Kelly-Hayes, M.; Kase, C.S.; Au, R.; Kannel, W.B.; Wolf, P.A. The Lifetime Risk of Stroke: Estimates from the Framingham Study. Stroke 2006, 37, 345–350. [Google Scholar] [CrossRef]
- Campbell, B.C.V.; De Silva, D.A.; Macleod, M.R.; Coutts, S.B.; Schwamm, L.H.; Davis, S.M.; Donnan, G.A. Ischaemic Stroke. Nat. Rev. Dis. Primers 2019, 5, 70. [Google Scholar] [CrossRef]
- Gatz, M.; Reynolds, C.A.; Fratiglioni, L.; Johansson, B.; Mortimer, J.A.; Berg, S.; Fiske, A.; Pedersen, N.L. Role of Genes and Environments for Explaining Alzheimer Disease. Arch. Gen. Psychiatry 2006, 63, 168–174. [Google Scholar] [CrossRef]
- Bekris, L.M.; Yu, C.E.; Bird, T.D.; Tsuang, D.W. Review Article: Genetics of Alzheimer Disease. J. Geriatr. Psychiatry Neurol. 2010, 23, 213–227. [Google Scholar]
- Gorelick, P.B.; Scuteri, A.; Black, S.E.; Decarli, C.; Greenberg, S.M.; Iadecola, C.; Launer, L.J.; Laurent, S.; Lopez, O.L.; Nyenhuis, D.; et al. Vascular Contributions to Cognitive Impairment and Dementia: A Statement for Healthcare Professionals from the American Heart Association/American Stroke Association. Stroke 2011, 42, 2672–2713. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yu, J.T.; Wang, H.F.; Meng, X.F.; Tan, C.C.; Wang, J.; Wang, C.; Tana, L. Association between Stroke and Alzheimer’s Disease: Systematic Review and Meta-Analysis. J. Alzheimers Dis. 2015, 43, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Waziry, R.; Chibnik, L.B.; Bos, D.; Ikram, M.K.; Hofman, A. Risk of Hemorrhagic and Ischemic Stroke in Patients with Alzheimer Disease: A Synthesis of the Literature. Neurology 2020, 94, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Pinho, J.; Quintas-Neves, M.; Dogan, I.; Reetz, K.; Reich, A.; Costa, A.S. Incident Stroke in Patients with Alzheimer’s Disease: Systematic Review and Meta-Analysis. Sci. Rep. 2021, 11, 16385. [Google Scholar] [CrossRef]
- Boehme, A.K.; Esenwa, C.; Elkind, M.S.V. Stroke Risk Factors, Genetics, and Prevention. Circ. Res. 2017, 120, 472–495. [Google Scholar] [CrossRef]
- Silva, M.V.F.; Loures, C.D.M.G.; Alves, L.C.V.; De Souza, L.C.; Borges, K.B.G.; Carvalho, M.D.G. Alzheimer’s Disease: Risk Factors and Potentially Protective Measures. J. Biomed. Sci. 2019, 26, 33. [Google Scholar] [CrossRef]
- Nordestgaard, L.T.; Christoffersen, M.; Frikke-Schmidt, R. Shared Risk Factors between Dementia and Atherosclerotic Cardiovascular Disease. Int. J. Mol. Sci. 2022, 23, 9777. [Google Scholar] [CrossRef]
- Pluta, R.; Kida, E.; Lossinsky, A.S.; Golabek, A.A.; Mossakowski, M.J.; Wisniewski, H.M. Complete Cerebral Ischemia with Short-Term Survival in Rats Induced by Cardiac Arrest. I. Extracellular Accumulation of Alzheimer’s Beta-Amyloid Protein Precursor in the Brain. Brain Res. 1994, 649, 323–328. [Google Scholar] [CrossRef]
- Stephenson, D.T.; Rash, K.; Clemens, J.A. Amyloid Precursor Protein Accumulates in Regions of Neurodegeneration Following Focal Cerebral Ischemia in the Rat. Brain Res. 1992, 593, 128–135. [Google Scholar] [CrossRef]
- Qi, J.P.; Wu, H.; Yang, Y.; Wang, D.D.; Chen, Y.X.; Gu, Y.H.; Liu, T. Cerebral Ischemia and Alzheimer’s Disease: The Expression of Amyloid-Beta and Apolipoprotein E in Human Hippocampus. J. Alzheimers Dis. 2007, 12, 335–341. [Google Scholar] [CrossRef]
- Ouyang, F.; Jiang, Z.; Chen, X.; Chen, Y.; Wei, J.; Xing, S.; Zhang, J.; Fan, Y.; Zeng, J. Is Cerebral Amyloid-β Deposition Related to Post-Stroke Cognitive Impairment? Transl. Stroke Res. 2021, 12, 946–957. [Google Scholar] [CrossRef] [PubMed]
- Bloom, G.S. Amyloid-β and Tau: The Trigger and Bullet in Alzheimer Disease Pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Michalicova, A.; Majerova, P.; Kovac, A. Tau Protein and Its Role in Blood-Brain Barrier Dysfunction. Front. Mol. Neurosci. 2020, 13, 570045. [Google Scholar] [CrossRef]
- Banks, W.A.; Kovac, A.; Majerova, P.; Bullock, K.M.; Shi, M.; Zhang, J. Tau Proteins Cross the Blood-Brain Barrier. J. Alzheimers Dis. 2017, 55, 411–419. [Google Scholar] [CrossRef]
- Yu, S.P.; Jiang, M.Q.; Shim, S.S.; Pourkhodadad, S.; Wei, L. Extrasynaptic NMDA Receptors in Acute and Chronic Excitotoxicity: Implications for Preventive Treatments of Ischemic Stroke and Late-Onset Alzheimer’s Disease. Mol. Neurodegener. 2023, 18, 43. [Google Scholar] [CrossRef] [PubMed]
- Attems, J.; Jellinger, K.A. The Overlap between Vascular Disease and Alzheimer’s Disease—Lessons from Pathology. BMC Med. 2014, 12, 206. [Google Scholar] [CrossRef]
- Pluta, R.; Ułamek-Kozioł, M.; Januszewski, S.; Czuczwar, S.J. Shared Genomic and Proteomic Contribution of Amyloid and Tau Protein Characteristic of Alzheimer’s Disease to Brain Ischemia. Int. J. Mol. Sci. 2020, 21, 3186. [Google Scholar]
- Wang, T.; Ni, Q.B.; Wang, K.; Han, Z.; Sun, B.L. Stroke and Alzheimer’s Disease: A Mendelian Randomization Study. Front. Genet. 2020, 11, 581. [Google Scholar] [CrossRef]
- Traylor, M.; Adib-Samii, P.; Harold, D.; Dichgans, M.; Williams, J.; Lewis, C.M.; Markus, H.S.; Fornage, M.; Holliday, E.G.; Sharma, P.; et al. Shared Genetic Contribution to Ischemic Stroke and Alzheimer’s Disease. Ann. Neurol. 2016, 79, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Rost, N.S.; Meschia, J.F.; Gottesman, R.; Wruck, L.; Helmer, K.; Greenberg, S.M.; Barrett, K.; Biffi, A.; Boden-Albala, B.; Fornage, M.; et al. Cognitive Impairment and Dementia After Stroke: Design and Rationale for the DISCOVERY Study. Stroke 2021, 52, e499–e516. [Google Scholar] [CrossRef]
- Brabec, J.L.; Lara, M.K.; Tyler, A.L.; Mahoney, J.M. System-Level Analysis of Alzheimer’s Disease Prioritizes Candidate Genes for Neurodegeneration. Front. Genet. 2021, 12, 625246. [Google Scholar] [CrossRef] [PubMed]
- Droś, J.; Klimkowicz-Mrowiec, A. Current View on Post-Stroke Dementia. Psychogeriatrics 2021, 21, 407–417. [Google Scholar] [PubMed]
- Leys, D.; Hénon, H.; Mackowiak-Cordoliani, M.A.; Pasquier, F. Poststroke Dementia. Lancet Neurol. 2005, 4, 752–759. [Google Scholar] [PubMed]
- Bi, M.; Gladbach, A.; Van Eersel, J.; Ittner, A.; Przybyla, M.; Van Hummel, A.; Chua, S.W.; Van Der Hoven, J.; Lee, W.S.; Müller, J.; et al. Tau Exacerbates Excitotoxic Brain Damage in an Animal Model of Stroke. Nat. Commun. 2017, 8, 473. [Google Scholar] [CrossRef] [PubMed]
- Giannakopoulos, P.; Gold, G.; Kövari, E.; von Gunten, A.; Imhof, A.; Bouras, C.; Hof, P.R. Assessing the Cognitive Impact of Alzheimer Disease Pathology and Vascular Burden in the Aging Brain: The Geneva Experience. Acta Neuropathol. 2007, 113, 1–12. [Google Scholar] [CrossRef]
- Gallardo, G.; Holtzman, D.M. Amyloid-β and Tau at the Crossroads of Alzheimer’s Disease. Adv. Exp. Med. Biol. 2019, 1184, 187–203. [Google Scholar] [CrossRef]
- Krajcovicova, L.; Klobusiakova, P.; Rektorova, I. Gray Matter Changes in Parkinson’s and Alzheimer’s Disease and Relation to Cognition. Curr. Neurol. Neurosci. Rep. 2019, 19, 85. [Google Scholar] [CrossRef]
- Nikolac Perkovic, M.; Pivac, N. Genetic Markers of Alzheimer’s Disease. Adv. Exp. Med. Biol. 2019, 1192, 27–52. [Google Scholar] [CrossRef]
- Lanoiselée, H.M.; Nicolas, G.; Wallon, D.; Rovelet-Lecrux, A.; Lacour, M.; Rousseau, S.; Richard, A.C.; Pasquier, F.; Rollin-Sillaire, A.; Martinaud, O.; et al. APP, PSEN1, and PSEN2 Mutations in Early-Onset Alzheimer Disease: A Genetic Screening Study of Familial and Sporadic Cases. PLoS Med. 2017, 14, e1002270. [Google Scholar] [CrossRef]
- van Heusden, F.C.; van Nifterick, A.M.; Souza, B.C.; França, A.S.C.; Nauta, I.M.; Stam, C.J.; Scheltens, P.; Smit, A.B.; Gouw, A.A.; van Kesteren, R.E. Neurophysiological Alterations in Mice and Humans Carrying Mutations in APP and PSEN1 Genes. Alzheimers Res. Ther. 2023, 15, 142. [Google Scholar] [CrossRef]
- Quan, M.; Cao, S.; Wang, Q.; Wang, S.; Jia, J. Genetic Phenotypes of Alzheimer’s Disease: Mechanisms and Potential Therapy. Phenomics 2023, 3, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jin, F.; Yang, Z.; Lu, Z.; Kan, R.; Li, S.; Zheng, C.; Wang, L. The Insertion Polymorphism in Angiotensin-Converting Enzyme Gene Associated with the APOE Epsilon 4 Allele Increases the Risk of Late-Onset Alzheimer Disease. J. Mol. Neurosci. 2006, 30, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Padilla, V.; Lozano, S.; Gamez, D.; Su, B.B.; Wang, X.; Maestre, G.; Wang, K. APOE Gene Associated with Dementia-Related Traits, Depression, and Anxiety in the Hispanic Population. Genes 2023, 14, 1405. [Google Scholar] [CrossRef]
- Jackson, R.J.; Keiser, M.S.; Meltzer, J.C.; Fykstra, D.P.; Dierksmeier, S.E.; Melloni, A.; Nakajima, T.; Tecedor, L.; Ranum, P.T.; Carrell, E.; et al. APOE2 Gene Therapy Reduces Amyloid Deposition, and Improves Markers of Neuroinflammation and Neurodegeneration in a Mouse Model of Alzheimer Disease. bioRxiv 2023. [Google Scholar] [CrossRef]
- Elman-Shina, K.; Efrati, S. Ischemia as a Common Trigger for Alzheimer’s Disease. Front. Aging Neurosci. 2022, 14, 1012779. [Google Scholar] [CrossRef] [PubMed]
- Pluta, R.; Januszewski, S.; Czuczwar, S.J. Neuroinflammation in Post-Ischemic Neurodegeneration of the Brain: Friend, Foe, or Both? Int. J. Mol. Sci. 2021, 22, 4405. [Google Scholar] [CrossRef]
- Rost, N.S.; Brodtmann, A.; Pase, M.P.; van Veluw, S.J.; Biffi, A.; Duering, M.; Hinman, J.D.; Dichgans, M. Post-Stroke Cognitive Impairment and Dementia. Circ. Res. 2022, 130, 1252–1271. [Google Scholar] [CrossRef]
- Ułamek-Kozioł, M.; Czuczwar, S.J.; Januszewski, S.; Pluta, R. Proteomic and Genomic Changes in Tau Protein, Which Are Associated with Alzheimer’s Disease after Ischemia-Reperfusion Brain Injury. Int. J. Mol. Sci. 2020, 21, 892. [Google Scholar] [CrossRef]
- Kumari, S.; Dhapola, R.; Reddy, D.H.K. Apoptosis in Alzheimer’s Disease: Insight into the Signaling Pathways and Therapeutic Avenues. Apoptosis 2023, 28, 943–957. [Google Scholar] [CrossRef]
- Koenig, L.N.; McCue, L.M.; Grant, E.; Massoumzadeh, P.; Roe, C.M.; Xiong, C.; Moulder, K.L.; Wang, L.; Zazulia, A.R.; Kelly, P.; et al. Lack of Association between Acute Stroke, Post-Stroke Dementia, Race, and β-Amyloid Status. Neuroimage Clin. 2021, 29, 102553. [Google Scholar] [CrossRef]
- Montaner, J.; Ramiro, L.; Simats, A.; Tiedt, S.; Makris, K.; Jickling, G.C.; Debette, S.; Sanchez, J.C.; Bustamante, A. Multilevel Omics for the Discovery of Biomarkers and Therapeutic Targets for Stroke. Nat. Rev. Neurol. 2020, 16, 247–264. [Google Scholar] [PubMed]
- Malik, R.; Chauhan, G.; Traylor, M.; Sargurupremraj, M.; Okada, Y.; Mishra, A.; Rutten-Jacobs, L.; Giese, A.K.; Van Der Laan, S.W.; Gretarsdottir, S.; et al. Multiancestry Genome-Wide Association Study of 520,000 Subjects Identifies 32 Loci Associated with Stroke and Stroke Subtypes. Nat. Genet. 2018, 50, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, G.; Debette, S. Genetic Risk Factors for Ischemic and Hemorrhagic Stroke. Curr. Cardiol. Rep. 2016, 18, 124. [Google Scholar] [CrossRef] [PubMed]
- Titov, B.V.; Matveeva, N.A.; Martynov, M.Y.; Favorova, O.O. Ischemic Stroke as a Complex Polygenic Disease. Mol. Biol. 2015, 49, 224–248. [Google Scholar]
- Gudbjartsson, D.F.; Holm, H.; Gretarsdottir, S.; Thorleifsson, G.; Walters, G.B.; Thorgeirsson, G.; Gulcher, J.; Mathiesen, E.B.; Njølstad, I.; Nyrnes, A.; et al. A Sequence Variant in ZFHX3 on 16q22 Associates with Atrial Fibrillation and Ischemic Stroke. Nat. Genet. 2009, 41, 876–878. [Google Scholar] [CrossRef]
- Gudbjartsson, D.F.; Arnar, D.O.; Helgadottir, A.; Gretarsdottir, S.; Holm, H.; Sigurdsson, A.; Jonasdottir, A.; Baker, A.; Thorleifsson, G.; Kristjansson, K.; et al. Variants Conferring Risk of Atrial Fibrillation on Chromosome 4q25. Nature 2007, 448, 353–357. [Google Scholar] [CrossRef]
- International Stroke Genetics Consortium (ISGC); Wellcome Trust Case Control Consortium 2 (WTCCC2); Bellenguez, C.; Bevan, S.; Gschwendtner, A.; Spencer, C.C.A.; Burgess, A.I.; Pirinen, M.; Jackson, C.A.; Traylor, M.; et al. Genome-Wide Association Study Identifies a Variant in HDAC9 Associated with Large Vessel Ischemic Stroke. Nat. Genet. 2012, 44, 328–333. [Google Scholar] [CrossRef]
- Malik, R.; Traylor, M.; Pulit, S.L.; Bevan, S.; Hopewell, J.C.; Holliday, E.G.; Zhao, W.; Abrantes, P.; Amouyel, P.; Attia, J.R.; et al. Low-Frequency and Common Genetic Variation in Ischemic Stroke. Neurology 2016, 86, 1217–1226. [Google Scholar] [CrossRef]
- Georgakis, M.K.; Parodi, L.; Frerich, S.; Mayerhofer, E.; Tsivgoulis, G.; Pirruccello, J.P.; Slowik, A.; Rundek, T.; Malik, R.; Dichgans, M.; et al. Genetic Architecture of Stroke of Undetermined Source: Overlap with Known Stroke Etiologies and Associations with Modifiable Risk Factors. Ann. Neurol. 2022, 91, 640–651. [Google Scholar] [CrossRef]
- Mishra, A.; Malik, R.; Hachiya, T.; Jürgenson, T.; Namba, S.; Posner, D.C.; Kamanu, F.K.; Koido, M.; Le Grand, Q.; Shi, M.; et al. Stroke Genetics Informs Drug Discovery and Risk Prediction across Ancestries. Nature 2022, 611, 115–123. [Google Scholar] [CrossRef]
- Sun, S.; Dong, B.; Zou, Q. Revisiting Genome-Wide Association Studies from Statistical Modelling to Machine Learning. Brief. Bioinform. 2021, 22, bbaa263. [Google Scholar] [CrossRef]
- Nicholls, H.L.; John, C.R.; Watson, D.S.; Munroe, P.B.; Barnes, M.R.; Cabrera, C.P. Reaching the End-Game for GWAS: Machine Learning Approaches for the Prioritization of Complex Disease Loci. Front. Genet. 2020, 11, 350. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hou, L.; Xu, S.; Li, H.; Chen, X.; Gao, J.; Wang, Z.; Han, B.; Liu, X.; Wan, S. Discovering Cerebral Ischemic Stroke Associated Genes Based on Network Representation Learning. Front. Genet. 2021, 12, 728333. [Google Scholar] [CrossRef] [PubMed]
- Kamboh, M.I. Genomics and Functional Genomics of Alzheimer’s Disease. Neurotherapeutics 2022, 19, 152–172. [Google Scholar] [CrossRef]
- Harold, D.; Abraham, R.; Hollingworth, P.; Sims, R.; Gerrish, A.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; Dowzell, K.; Williams, A.; et al. Genome-Wide Association Study Identifies Variants at CLU and PICALM Associated with Alzheimer’s Disease. Nat. Genet. 2009, 41, 1088–1093. [Google Scholar] [CrossRef]
- Lambert, J.C.; Heath, S.; Even, G.; Campion, D.; Sleegers, K.; Hiltunen, M.; Combarros, O.; Zelenika, D.; Bullido, M.J.; Tavernier, B.; et al. Genome-Wide Association Study Identifies Variants at CLU and CR1 Associated with Alzheimer’s Disease. Nat. Genet. 2009, 41, 1094–1099. [Google Scholar] [CrossRef]
- Hollingworth, P.; Harold, D.; Sims, R.; Gerrish, A.; Lambert, J.C.; Carrasquillo, M.M.; Abraham, R.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; et al. Common Variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP Are Associated with Alzheimer’s Disease. Nat. Genet. 2011, 43, 429–436. [Google Scholar] [CrossRef]
- Naj, A.C.; Jun, G.; Beecham, G.W.; Wang, L.S.; Vardarajan, B.N.; Buros, J.; Gallins, P.J.; Buxbaum, J.D.; Jarvik, G.P.; Crane, P.K.; et al. Common Variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 Are Associated with Late-Onset Alzheimer’s Disease. Nat. Genet. 2011, 43, 436–443. [Google Scholar] [CrossRef]
- Benitez, B.A.; Jin, S.C.; Guerreiro, R.; Graham, R.; Lord, J.; Harold, D.; Sims, R.; Lambert, J.C.; Gibbs, J.R.; Bras, J.; et al. Missense Variant in TREML2 Protects against Alzheimer’s Disease. Neurobiol. Aging 2014, 35, 1510.e19–1510.e26. [Google Scholar] [CrossRef]
- Ruiz, A.; Dols-Icardo, O.; Bullido, M.J.; Pastor, P.; Rodríguez-Rodríguez, E.; López de Munain, A.; de Pancorbo, M.M.; Pérez-Tur, J.; Álvarez, V.; Antonell, A.; et al. Assessing the Role of the TREM2 p.R47H Variant as a Risk Factor for Alzheimer’s Disease and Frontotemporal Dementia. Neurobiol. Aging 2014, 35, 444.e1–444.e4. [Google Scholar] [CrossRef]
- Lambert, J.C.; Ibrahim-Verbaas, C.A.; Harold, D.; Naj, A.C.; Sims, R.; Bellenguez, C.; Jun, G.; DeStefano, A.L.; Bis, J.C.; Beecham, G.W.; et al. Meta-Analysis of 74,046 Individuals Identifies 11 New Susceptibility Loci for Alzheimer’s Disease. Nat. Genet. 2013, 45, 1452–1458. [Google Scholar] [CrossRef]
- Rajabli, F.; Tosto, G.; Hamilton-Nelson, K.L.; Kunkle, B.W.; Vardarajan, B.N.; Naj, A.; Whitehead, P.G.; Gardner, O.K.; Bush, W.S.; Sariya, S.; et al. Admixture Mapping Identifies Novel Alzheimer’s Disease Risk Regions in African Americans. Alzheimers Dement. 2022, 19, 2538–2548. [Google Scholar] [CrossRef]
- Jansen, I.E.; Savage, J.E.; Watanabe, K.; Bryois, J.; Williams, D.M.; Steinberg, S.; Sealock, J.; Karlsson, I.K.; Hägg, S.; Athanasiu, L.; et al. Genome-Wide Meta-Analysis Identifies New Loci and Functional Pathways Influencing Alzheimer’s Disease Risk. Nat. Genet. 2019, 51, 404–413. [Google Scholar] [CrossRef]
- Marioni, R.E.; Harris, S.E.; Zhang, Q.; McRae, A.F.; Hagenaars, S.P.; Hill, W.D.; Davies, G.; Ritchie, C.W.; Gale, C.R.; Starr, J.M.; et al. GWAS on Family History of Alzheimer’s Disease. Transl. Psychiatry 2018, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- Schwartzentruber, J.; Cooper, S.; Liu, J.Z.; Barrio-Hernandez, I.; Bello, E.; Kumasaka, N.; Young, A.M.H.; Franklin, R.J.M.; Johnson, T.; Estrada, K.; et al. Genome-Wide Meta-Analysis, Fine-Mapping and Integrative Prioritization Implicate New Alzheimer’s Disease Risk Genes. Nat. Genet. 2021, 53, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Wightman, D.P.; Jansen, I.E.; Savage, J.E.; Shadrin, A.A.; Bahrami, S.; Holland, D.; Rongve, A.; Børte, S.; Winsvold, B.S.; Drange, O.K.; et al. A Genome-Wide Association Study with 1,126,563 Individuals Identifies New Risk Loci for Alzheimer’s Disease. Nat. Genet. 2021, 53, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.; Leonenko, G.; Schmidt, K.M.; Hill, M.; Myers, A.J.; Shoai, M.; de Rojas, I.; Tesi, N.; Holstege, H.; van der Flier, W.M.; et al. What Does Heritability of Alzheimer’s Disease Represent? PLoS ONE 2023, 18, e0281440. [Google Scholar] [CrossRef]
- Wei, C.J.; Cui, P.; Li, H.; Lang, W.J.; Liu, G.Y.; Ma, X.F. Shared Genes between Alzheimer’s Disease and Ischemic Stroke. CNS Neurosci. Ther. 2019, 25, 855–864. [Google Scholar] [CrossRef]
- Ford, G.; Xu, Z.; Gates, A.; Jiang, J.; Ford, B.D. Expression Analysis Systematic Explorer (EASE) Analysis Reveals Differential Gene Expression in Permanent and Transient Focal Stroke Rat Models. Brain Res. 2006, 1071, 226–236. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.; Zhang, L.; Wang, G.; Zhang, M.; Yu, Y. Neuroprotection of Dexmedetomidine against Cerebral Ischemia-Reperfusion Injury in Rats: Involved in Inhibition of NF-ΚB and Inflammation Response. Biomol. Ther. 2017, 25, 383–389. [Google Scholar] [CrossRef]
- Marcheselli, V.L.; Hong, S.; Lukiw, W.J.; Tian, X.H.; Gronert, K.; Musto, A.; Hardy, M.; Gimenez, J.M.; Chiang, N.; Serhan, C.N.; et al. Novel Docosanoids Inhibit Brain Ischemia-Reperfusion-Mediated Leukocyte Infiltration and Pro-Inflammatory Gene Expression. J. Biol. Chem. 2003, 278, 43807–43817. [Google Scholar] [CrossRef] [PubMed]
- Nogawa, S.; Zhang, F.; Ross, M.E.; Iadecola, C. Cyclo-Oxygenase-2 Gene Expression in Neurons Contributes to Ischemic Brain Damage. J. Neurosci. 1997, 17, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, M.; Pan, Y.; Bai, B.; Chen, J. Global Gene Expression Profile of Cerebral Ischemia-Reperfusion Injury in Rat MCAO Model. Oncotarget 2017, 8, 74607–74622. [Google Scholar] [CrossRef]
- DeGracia, D.J. Regulation of MRNA Following Brain Ischemia and Reperfusion. Wiley Interdiscip. Rev. RNA 2017, 8, e1415. [Google Scholar] [CrossRef]
- Dergunova, L.V.; Filippenkov, I.B.; Stavchansky, V.V.; Denisova, A.E.; Yuzhakov, V.V.; Mozerov, S.A.; Gubsky, L.V.; Limborska, S.A. Genome-Wide Transcriptome Analysis Using RNA-Seq Reveals a Large Number of Differentially Expressed Genes in a Transient MCAO Rat Model. BMC Genom. 2018, 19, 655. [Google Scholar] [CrossRef]
- Bub, A.; Brenna, S.; Alawi, M.; Kügler, P.; Gui, Y.; Kretz, O.; Altmeppen, H.; Magnus, T.; Puig, B. Multiplexed MRNA Analysis of Brain-Derived Extracellular Vesicles upon Experimental Stroke in Mice Reveals Increased MRNA Content with Potential Relevance to Inflammation and Recovery Processes. Cell Mol. Life Sci. 2022, 79, 329. [Google Scholar] [CrossRef] [PubMed]
- Torres-Querol, C.; Torres, P.; Vidal, N.; Portero-Otín, M.; Arque, G.; Purroy, F. Acute Ischemic Stroke Triggers a Cellular Senescence-Associated Secretory Phenotype. Sci. Rep. 2021, 11, 15752. [Google Scholar] [CrossRef]
- Behrouzifar, S.; Vakili, A.; Barati, M. The Effects of Mouse Recombinant Resistin on MRNA Expression of Proinflammatory and Anti-Inflammatory Cytokines and Heat Shock Protein-70 in Experimental Stroke Model. J. Stroke Cerebrovasc. Dis. 2018, 27, 3272–3279. [Google Scholar] [CrossRef]
- Dagonnier, M.; Wilson, W.J.; Favaloro, J.M.; Rewell, S.S.J.; Lockett, L.J.; Sastra, S.A.; Jeffreys, A.L.; Dewey, H.M.; Donnan, G.A.; Howells, D.W. Hyperacute Changes in Blood MRNA Expression Profiles of Rats after Middle Cerebral Artery Occlusion: Towards a Stroke Time Signature. PLoS ONE 2018, 13, e0206321. [Google Scholar] [CrossRef]
- Yeh, C.F.; Chuang, T.Y.; Lan, M.Y.; Chin, Y.C.; Wang, W.H.; Lin, Y.Y. Excessive Expression of Microglia/Macrophage and Proinflammatory Mediators in Olfactory Bulb and Olfactory Dysfunction After Stroke. In Vivo 2019, 33, 1893–1899. [Google Scholar] [CrossRef]
- Zheng, K.; Lin, L.; Jiang, W.; Chen, L.; Zhang, X.; Zhang, Q.; Ren, Y.; Hao, J. Single-Cell RNA-Seq Reveals the Transcriptional Landscape in Ischemic Stroke. J. Cereb. Blood Flow. Metab. 2022, 42, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lyu, J.; Li, R.; Jain, V.; Shen, Y.; del Águila, Á.; Hoffmann, U.; Sheng, H.; Yang, W. Single-Cell Transcriptomic Analysis of the Immune Cell Landscape in the Aged Mouse Brain after Ischemic Stroke. J. Neuroinflamm. 2022, 19, 83. [Google Scholar] [CrossRef] [PubMed]
- Dickey, C.A.; Loring, J.F.; Montgomery, J.; Gordon, M.N.; Eastman, P.S.; Morgan, D. Selectively Reduced Expression of Synaptic Plasticity-Related Genes in Amyloid Precursor Protein + Presenilin-1 Transgenic Mice. J. Neurosci. 2003, 23, 5219–5226. [Google Scholar] [CrossRef] [PubMed]
- Frautschy, S.A.; Yang, F.; Irrizarry, M.; Hyman, B.; Saido, T.C.; Hsiao, K.; Cole, G.M. Microglial Response to Amyloid Plaques in APPsw Transgenic Mice. Am. J. Pathol. 1998, 152, 307–317. [Google Scholar] [PubMed]
- Howlett, D.R.; Bowler, K.; Soden, P.E.; Riddell, D.; Davis, J.B.; Richardson, J.C.; Burbidge, S.A.; Gonzalez, M.I.; Irving, E.A.; Lawman, A.; et al. Aß Deposition and Related Pathology in an APP x PS1 Transgenic Mouse Model of Alzheimer’s Disease. Histol. Histopathol. 2008, 23, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; McWeeney, S.; Park, B.S.; Manczak, M.; Gutala, R.V.; Partovi, D.; Jung, Y.; Yau, V.; Searles, R.; Mori, M.; et al. Gene Expression Profiles of Transcripts in Amyloid Precursor Protein Transgenic Mice: Up-Regulation of Mitochondrial Metabolism and Apoptotic Genes Is an Early Cellular Change in Alzheimer’s Disease. Hum. Mol. Genet. 2004, 13, 1225–1240. [Google Scholar] [CrossRef]
- Wirz, K.T.S.; Bossers, K.; Stargardt, A.; Kamphuis, W.; Swaab, D.F.; Hol, E.M.; Verhaagen, J. Cortical Beta Amyloid Protein Triggers an Immune Response, but No Synaptic Changes in the APPswe/PS1dE9 Alzheimer’s Disease Mouse Model. Neurobiol. Aging 2013, 34, 1328–1342. [Google Scholar] [CrossRef][Green Version]
- Matarin, M.; Salih, D.A.; Yasvoina, M.; Cummings, D.M.; Guelfi, S.; Liu, W.; NahabooSolim, M.A.; Moens, T.G.; Paublete, R.M.; Ali, S.S.; et al. A Genome-Wide Gene-Expression Analysis and Database in Transgenic Mice during Development of Amyloid or Tau Pathology. Cell Rep. 2015, 10, 633–644. [Google Scholar] [CrossRef]
- Sierksma, A.; Lu, A.; Mancuso, R.; Fattorelli, N.; Thrupp, N.; Salta, E.; Zoco, J.; Blum, D.; Buée, L.; De Strooper, B.; et al. Novel Alzheimer Risk Genes Determine the Microglia Response to Amyloid-β but Not to TAU Pathology. EMBO Mol. Med. 2020, 12, e10606. [Google Scholar] [CrossRef]
- De Strooper, B.; Karran, E. The Cellular Phase of Alzheimer’s Disease. Cell 2016, 164, 603–615. [Google Scholar]
- Chen, W.T.; Lu, A.; Craessaerts, K.; Pavie, B.; Sala Frigerio, C.; Corthout, N.; Qian, X.; Laláková, J.; Kühnemund, M.; Voytyuk, I.; et al. Spatial Transcriptomics and In Situ Sequencing to Study Alzheimer’s Disease. Cell 2020, 182, 976–991.e19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Song, W.M.; Andhey, P.S.; Swain, A.; Levy, T.; Miller, K.R.; Poliani, P.L.; Cominelli, M.; Grover, S.; Gilfillan, S.; et al. Human and Mouse Single-Nucleus Transcriptomics Reveal TREM2-Dependent and TREM2-Independent Cellular Responses in Alzheimer’s Disease. Nat. Med. 2020, 26, 131–142. [Google Scholar] [CrossRef]
- Hammond, T.R.; Dufort, C.; Dissing-Olesen, L.; Giera, S.; Young, A.; Wysoker, A.; Walker, A.J.; Gergits, F.; Segel, M.; Nemesh, J.; et al. Single-Cell RNA Sequencing of Microglia throughout the Mouse Lifespan and in the Injured Brain Reveals Complex Cell-State Changes. Immunity 2019, 50, 253–271.e6. [Google Scholar] [CrossRef] [PubMed]
- Majerníková, N.; den Dunnen, W.F.A.; Dolga, A.M. The Potential of Ferroptosis-Targeting Therapies for Alzheimer’s Disease: From Mechanism to Transcriptomic Analysis. Front. Aging Neurosci. 2021, 13, 745046. [Google Scholar]
- Liu, W.; Wan, M.; Shi, Y.; Yang, X.Z. Transcriptomic Analysis Identifies Shared Biological Foundations between Ischemic Stroke and Alzheimer’s Disease. Front. Neurosci. 2022, 16, 1008752. [Google Scholar] [CrossRef]
- Song, J. Comparison of Cerebral Cortex Transcriptome Profiles in Ischemic Stroke and Alzheimer’s Disease Models. Clin. Nutr. Res. 2022, 11, 159. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Wang, Q.; Ma, J.; Gao, X.; Wang, M. TREM2, Microglial and Ischemic Stroke. J. Neuroimmunol. 2023, 381, 578108. [Google Scholar] [CrossRef]
- Ruffo, P.; De Amicis, F.; Giardina, E.; Conforti, F. Long-Noncoding RNAs as Epigenetic Regulators in Neurodegenerative Diseases. Neural Regen. Res. 2023, 18, 1243. [Google Scholar] [CrossRef]
- Sufianova, G.; Shumadalova, A.; Wenhao, Y.; Gareev, I. Long Non-Coding RNAs as Biomarkers and Therapeutic Targets for Ischemic Stroke. Noncoding RNA Res. 2022, 7, 226–232. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Wen, Z.-J.; Xu, H.-M.; Zhang, Y.; Zhang, Y.-F. Exosomal Noncoding RNAs in Central Nervous System Diseases: Biological Functions and Potential Clinical Applications. Front. Mol. Neurosci. 2022, 15, 1004221. [Google Scholar] [CrossRef]
- He, L.; Zhang, F.; Zhu, Y.; Lu, M. A Crosstalk between Circular RNA, MicroRNA, and Messenger RNA in the Development of Various Brain Cognitive Disorders. Front. Mol. Neurosci. 2022, 15, 960657. [Google Scholar] [CrossRef] [PubMed]
- Sherazi, S.; Abbasi, A.; Jamil, A.; Uzair, M.; Ikram, A.; Qamar, S.; Olamide, A.; Arshad, M.; Fried, P.; Ljubisavljevic, M.; et al. Molecular Hallmarks of Long Non-Coding RNAs in Aging and Its Significant Effect on Aging-Associated Diseases. Neural Regen. Res. 2023, 18, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Fullerton, J.L.; Thomas, J.M.; Gonzalez-Trueba, L.; Trivett, C.; van Kralingen, J.C.; Allan, S.M.; Quinn, T.J.; Work, L.M. Systematic Review: Association between Circulating MicroRNA Expression & Stroke. J. Cereb. Blood Flow. Metab. 2022, 42, 935–951. [Google Scholar] [CrossRef] [PubMed]
- Toor, S.M.; Aldous, E.K.; Parray, A.; Akhtar, N.; Al-Sarraj, Y.; Abdelalim, E.M.; Arredouani, A.; El-Agnaf, O.; Thornalley, P.J.; Pananchikkal, S.V.; et al. Circulating MicroRNA Profiling Identifies Distinct MicroRNA Signatures in Acute Ischemic Stroke and Transient Ischemic Attack Patients. Int. J. Mol. Sci. 2022, 24, 108. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.P.; McKay, W.; Edwards, J.S.; Swaminathan, R.; Santacruz, K.S.; Mims, R.L.; Yonas, H.; Roitbak, T. MicroRNA Analysis of Human Stroke Brain Tissue Resected during Decompressive Craniectomy/Stroke-Ectomy Surgery. Genes 2021, 12, 1860. [Google Scholar] [CrossRef]
- Voelz, C.; Ebrahimy, N.; Zhao, W.; Habib, P.; Zendedel, A.; Pufe, T.; Beyer, C.; Slowik, A. Transient Focal Cerebral Ischemia Leads to MiRNA Alterations in Different Brain Regions, Blood Serum, Liver, and Spleen. Int. J. Mol. Sci. 2021, 23, 161. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, F.; Zhao, X.; Zhang, Y.; Wang, X.; Zhang, Z.; Yang, L.Z.; Badshah, J.S.; Xu, B.; Xie, R.; et al. Expression Analysis and Targets Prediction of MicroRNAs in OGD/R Treated Astrocyte-Derived Exosomes by SmallRNA Sequencing. Genomics 2023, 115, 110594. [Google Scholar] [CrossRef]
- Denk, J.; Oberhauser, F.; Kornhuber, J.; Wiltfang, J.; Fassbender, K.; Schroeter, M.L.; Volk, A.E.; Diehl-Schmid, J.; Prudlo, J.; Danek, A.; et al. Specific Serum and CSF MicroRNA Profiles Distinguish Sporadic Behavioural Variant of Frontotemporal Dementia Compared with Alzheimer Patients and Cognitively Healthy Controls. PLoS ONE 2018, 13, e0197329. [Google Scholar] [CrossRef]
- Müller, M.; Kuiperij, H.B.; Claassen, J.A.; Küsters, B.; Verbeek, M.M. MicroRNAs in Alzheimer’s Disease: Differential Expression in Hippocampus and Cell-Free Cerebrospinal Fluid. Neurobiol. Aging 2014, 35, 152–158. [Google Scholar] [CrossRef]
- Takousis, P.; Sadlon, A.; Schulz, J.; Wohlers, I.; Dobricic, V.; Middleton, L.; Lill, C.M.; Perneczky, R.; Bertram, L. Differential Expression of MicroRNAs in Alzheimer’s Disease Brain, Blood, and Cerebrospinal Fluid. Alzheimers Dement. 2019, 15, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Kim, S.E.; Ko, Y.; Jeong, G.H.; Lee, K.H.; Lee, J.; Solmi, M.; Jacob, L.; Smith, L.; Stickley, A.; et al. Differential Expression of MicroRNAs in Alzheimer’s Disease: A Systematic Review and Meta-Analysis. Mol. Psychiatry 2022, 27, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Jiang, H.L.; Ashraf, G.; Li, Z.R.; Liu, R. MicroRNA and MRNA Profiling of Cerebral Cortex in a Transgenic Mouse Model of Alzheimer’s Disease by RNA Sequencing. Neural Regen. Res. 2021, 16, 2099. [Google Scholar] [CrossRef]
- Mirzaei, H.; Momeni, F.; Saadatpour, L.; Sahebkar, A.; Goodarzi, M.; Masoudifar, A.; Kouhpayeh, S.; Salehi, H.; Mirzaei, H.R.; Jaafari, M.R. MicroRNA: Relevance to Stroke Diagnosis, Prognosis, and Therapy. J. Cell Physiol. 2018, 233, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Wang, Z.Y.; Ma, L.N.; Zhang, T.T.; Cao, Y.; Li, H. MicroRNAs in Alzheimer’s Disease: Function and Potential Applications as Diagnostic Biomarkers. Front. Mol. Neurosci. 2020, 13, 160. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.; Chen, D.; Chen, N. The Regulation of MicroRNAs in Alzheimer’s Disease. Front. Neurol. 2020, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.L.; Chokkalla, A.K.; Bathula, S.; Vemuganti, R. MicroRNA MiR-7 Is Essential for Post-Stroke Functional Recovery. Transl. Stroke Res. 2023, 14, 111–115. [Google Scholar] [CrossRef]
- La Rosa, F.; Mancuso, R.; Agostini, S.; Piancone, F.; Marventano, I.; Saresella, M.; Hernis, A.; Fenoglio, C.; Galimberti, D.; Scarpini, E.; et al. Pharmacological and Epigenetic Regulators of NLRP3 Inflammasome Activation in Alzheimer’s Disease. Pharmaceuticals 2021, 14, 1187. [Google Scholar] [CrossRef]
- Puthiyedth, N.; Riveros, C.; Berretta, R.; Moscato, P. Identification of Differentially Expressed Genes through Integrated Study of Alzheimer’s Disease Affected Brain Regions. PLoS ONE 2016, 11, e0152342. [Google Scholar] [CrossRef]
- Deng, Y.; Ding, Y.; Hou, D. Research Status of the Regulation of MiRNA on BACE1. Int. J. Neurosci. 2014, 124, 474–477. [Google Scholar] [CrossRef]
- Shi, Z.; Chen, T.; Yao, Q.; Zheng, L.; Zhang, Z.; Wang, J.; Hu, Z.; Cui, H.; Han, Y.; Han, X.; et al. The Circular RNA CiRS-7 Promotes APP and BACE1 Degradation in an NF-ΚB-Dependent Manner. FEBS J. 2017, 284, 1096–1109. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Xu, J.; Zhong, P.; Fang, L. MiR-125a-5p Silencing Inhibits Cerebral Ischemia-Induced Injury through Targeting IGFBP3. Folia Neuropathol. 2021, 59, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, H.; Feng, J.; Chen, L. Overexpression of CircRNA CircUCK2 Attenuates Cell Apoptosis in Cerebral Ischemia-Reperfusion Injury via MiR-125b-5p/GDF11 Signaling. Mol. Ther. Nucleic Acids 2020, 22, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Chen, Z.; Cai, P.; Wang, R.; Yang, Q.; Li, L.; Yang, H.; Zhu, R. Targeting MicroRNA-125b Promotes Neurite Outgrowth but Represses Cell Apoptosis and Inflammation via Blocking PTGS2 and CDK5 in a FOXQ1-Dependent Way in Alzheimer Disease. Front. Cell Neurosci. 2020, 14, 587747. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.P.; Wang, X.H.; Zhang, H.X.; Shang, M.M.; Liu, X.X.; Sun, H.M.; Song, Y.P. MiR-103 Regulates the Angiogenesis of Ischemic Stroke Rats by Targeting Vascular Endothelial Growth Factor (VEGF). Iran. J. Basic. Med. Sci. 2018, 21, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Vinciguerra, A.; Formisano, L.; Cerullo, P.; Guida, N.; Cuomo, O.; Esposito, A.; Di Renzo, G.; Annunziato, L.; Pignataro, G. MicroRNA-103-1 Selectively Downregulates Brain NCX1 and Its Inhibition by Anti-MiRNA Ameliorates Stroke Damage and Neurological Deficits. Mol. Ther. 2014, 22, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Liang, Y.; Yu, S. Downregulation of MicroRNA-103a Reduces Microvascular Endothelial Cell Injury in a Rat Model of Cerebral Ischemia by Targeting AXIN2. J. Cell Physiol. 2020, 235, 4720–4733. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Shang, H.; Chen, X.; Yang, S.; Qu, Y.; Ding, J.; Li, X. Circular RNA Circ_0000950 Promotes Neuron Apoptosis, Suppresses Neurite Outgrowth and Elevates Inflammatory Cytokines Levels via Directly Sponging MiR-103 in Alzheimer’s Disease. Cell Cycle 2019, 18, 2197–2214. [Google Scholar] [CrossRef]
- Han, B.; Zhang, Y.; Zhang, Y.; Bai, Y.; Chen, X.; Huang, R.; Wu, F.; Leng, S.; Chao, J.; Zhang, J.H.; et al. Novel Insight into Circular RNA HECTD1 in Astrocyte Activation via Autophagy by Targeting MIR142-TIPARP: Implications for Cerebral Ischemic Stroke. Autophagy 2018, 14, 1164–1184. [Google Scholar] [CrossRef]
- Liang, W.; Xie, Z.; Liao, D.; Li, Y.; Li, Z.; Zhao, Y.; Li, X.; Dong, M. Inhibiting MicroRNA-142-5p Improves Learning and Memory in Alzheimer’s Disease Rats via Targeted Regulation of the PTPN1-Mediated Akt Pathway. Brain Res. Bull. 2023, 192, 107–114. [Google Scholar] [CrossRef]
- Liu, W.; Miao, Y.; Zhang, L.; Xu, X.; Luan, Q. MiR-211 Protects Cerebral Ischemia/Reperfusion Injury by Inhibiting Cell Apoptosis. Bioengineered 2020, 11, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Qi, X.; Liu, C.; Wang, D.; Li, L.; Liu, X.; Hou, Y.; Su, X.; Lin, H. The Silent Information Regulator 1 Pathway Attenuates ROS-Induced Oxidative Stress in Alzheimer’s Disease. J. Integr. Neurosci. 2020, 19, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wu, Q.; Ye, X.; Luo, H.; Yan, D.; Xiong, Y.; Zhu, H.; Diao, Y.; Zhang, W.; Wan, J. Role of MiR-211 in Neuronal Differentiation and Viability: Implications to Pathogenesis of Alzheimer’s Disease. Front. Aging Neurosci. 2016, 8, 166. [Google Scholar] [CrossRef]
- Huo, H.; Hu, C.; Lu, Y.; Zhou, J.; Mai, Z. Silencing of CircCDC14A Prevents Cerebral Ischemia-Reperfusion Injury via MiR-23a-3p/CXCL12 Axis. J. Biochem. Mol. Toxicol. 2022, 36, e22982. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Liu, J.; Guo, S.; Zeng, L.; Cai, Z.; Zhang, J.; Wang, L.; Li, Z.; Liu, R. MiR-23b-3p Rescues Cognition in Alzheimer’s Disease by Reducing Tau Phosphorylation and Apoptosis via GSK-3β Signaling Pathways. Mol. Ther. Nucleic Acids 2022, 28, 539–557. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Chen, D.; Gao, F.; Lv, H.; Zhang, G.; Sun, X.; Liu, L.; Mo, D.; Ma, N.; Song, L.; et al. Exosomes Derived from MicroRNA-138-5p-Overexpressing Bone Marrow-Derived Mesenchymal Stem Cells Confer Neuroprotection to Astrocytes Following Ischemic Stroke via Inhibition of LCN2. J. Biol. Eng. 2019, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Tang, C.; Wang, D. LncRNA H19 Promotes Inflammatory Response Induced by Cerebral Ischemia-Reperfusion Injury through Regulating the MiR-138-5p-P65 Axis. Biochem. Cell Biol. 2020, 98, 525–536. [Google Scholar] [CrossRef]
- Lu, Y.; Tan, L.; Wang, X. Circular HDAC9/MicroRNA-138/Sirtuin-1 Pathway Mediates Synaptic and Amyloid Precursor Protein Processing Deficits in Alzheimer’s Disease. Neurosci. Bull. 2019, 35, 877–888. [Google Scholar] [CrossRef]
- Wang, X.; Tan, L.; Lu, Y.; Peng, J.; Zhu, Y.; Zhang, Y.; Sun, Z. MicroRNA-138 Promotes Tau Phosphorylation by Targeting Retinoic Acid Receptor Alpha. FEBS Lett. 2015, 589, 726–729. [Google Scholar] [CrossRef]
- Wu, F.; Han, B.; Wu, S.; Yang, L.; Leng, S.; Li, M.; Liao, J.; Wang, G.; Ye, Q.; Zhang, Y.; et al. Circular RNA TLK1 Aggravates Neuronal Injury and Neurological Deficits after Ischemic Stroke via MiR-335-3p/TIPARP. J. Neurosci. 2019, 39, 7369–7393. [Google Scholar] [CrossRef]
- Si, W.; Ye, S.; Ren, Z.; Liu, X.; Wu, Z.; Li, Y.; Zhou, J.; Zhang, S.; Li, Y.; Deng, R.; et al. MiR-335 Promotes Stress Granule Formation to Inhibit Apoptosis by Targeting ROCK2 in Acute Ischemic Stroke. Int. J. Mol. Med. 2019, 43, 1452–1466. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Fei, Z.; Luo, S.; Wang, H. MiR-335-5p Inhibits β-Amyloid (Aβ) Accumulation to Attenuate Cognitive Deficits Through Targeting c-Jun-N-Terminal Kinase 3 in Alzheimer’s Disease. Curr. Neurovasc. Res. 2020, 17, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Jun, J.; Cong, L.; Du, L.; Wang, C. MiR-328-3p, a Predictor of Stroke, Aggravates the Cerebral Ischemia-Reperfusion Injury. Int. J. Gen. Med. 2021, 14, 2367–2376. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, X.; Fan, H.; Sun, J.; Ni, M.; Zhang, L.; Fang, F.; Zhang, W.; Ma, P. Circular RNA AXL Increases Neuron Injury and Inflammation through Targeting MicroRNA-328 Mediated BACE1 in Alzheimer’s Disease. Neurosci. Lett. 2022, 776, 136531. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Tao, Z.; Wang, R.; Liu, P.; Yan, F.; Li, J.; Zhang, C.; Ji, X.; Luo, Y. MicroRNA-23a-3p Attenuates Oxidative Stress Injury in a Mouse Model of Focal Cerebral Ischemia-Reperfusion. Brain Res. 2014, 1592, 65–72. [Google Scholar] [CrossRef]
- Filippenkov, I.B.; Stavchansky, V.V.; Denisova, A.E.; Valieva, L.V.; Remizova, J.A.; Mozgovoy, I.V.; Zaytceva, E.I.; Gubsky, L.V.; Limborska, S.A.; Dergunova, L. V Genome-Wide RNA-Sequencing Reveals Massive Circular RNA Expression Changes of the Neurotransmission Genes in the Rat Brain after Ischemia-Reperfusion. Genes 2021, 12, 1870. [Google Scholar] [CrossRef]
- Suzuki, H.; Tsukahara, T. A View of Pre-MRNA Splicing from RNase R Resistant RNAs. Int. J. Mol. Sci. 2014, 15, 9331–9342. [Google Scholar] [CrossRef]
- Suzuki, H.; Zuo, Y.; Wang, J.; Zhang, M.Q.; Malhotra, A.; Mayeda, A. Characterization of RNase R-Digested Cellular RNA Source That Consists of Lariat and Circular RNAs from Pre-MRNA Splicing. Nucleic Acids Res. 2006, 34, e63. [Google Scholar] [CrossRef]
- Liang, D.; Tatomer, D.C.; Luo, Z.; Wu, H.; Yang, L.; Chen, L.L.; Cherry, S.; Wilusz, J.E. The Output of Protein-Coding Genes Shifts to Circular RNAs When the Pre-MRNA Processing Machinery Is Limiting. Mol. Cell 2017, 68, 940–954.e3. [Google Scholar] [CrossRef]
- Liang, D.; Wilusz, J.E. Short Intronic Repeat Sequences Facilitate Circular RNA Production. Genes. Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef]
- Chen, B.J.; Yang, B.; Janitz, M. Region-Specific Expression of Circular RNAs in the Mouse Brain. Neurosci. Lett. 2018, 666, 44–47. [Google Scholar] [CrossRef]
- Filippenkov, I.B.; Sudarkina, O.Y.; Limborska, S.A.; Dergunova, L. V Circular RNA of the Human Sphingomyelin Synthase 1 Gene: Multiple Splice Variants, Evolutionary Conservatism and Expression in Different Tissues. RNA Biol. 2015, 12, 1030–1042. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA Circles Function as Efficient MicroRNA Sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Holdt, L.M.; Stahringer, A.; Sass, K.; Pichler, G.; Kulak, N.A.; Wilfert, W.; Kohlmaier, A.; Herbst, A.; Northoff, B.H.; Nicolaou, A.; et al. Circular Non-Coding RNA ANRIL Modulates Ribosomal RNA Maturation and Atherosclerosis in Humans. Nat. Commun. 2016, 7, 12429. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, C.; Yang, J.; Geng, X.; Du, H.; Ji, X.; Zhao, H. Screening Circular RNA Expression Patterns Following Focal Cerebral Ischemia in Mice. Oncotarget 2017, 8, 86535–86547. [Google Scholar] [CrossRef]
- Filippenkov, I.B.; Stavchansky, V.V.; Denisova, A.E.; Ivanova, K.A.; Limborska, S.A.; Dergunova, L. V Experimental Cerebral Ischemia Affects the Expression of Circular RNA Genes of Metabotropic Glutamate Receptors MGluR3 and MGluR5 in Rat Brain. Russ. J. Bioorg. Chem. 2018, 44, 302–309. [Google Scholar] [CrossRef]
- Piscopo, P.; Manzini, V.; Rivabene, R.; Crestini, A.; Le Pera, L.; Pizzi, E.; Veroni, C.; Talarico, G.; Peconi, M.; Castellano, A.E.; et al. A Plasma Circular RNA Profile Differentiates Subjects with Alzheimer’s Disease and Mild Cognitive Impairment from Healthy Controls. Int. J. Mol. Sci. 2022, 23, 13232. [Google Scholar] [CrossRef]
- Lukiw, W.J. Circular RNA (CircRNA) in Alzheimer’s Disease (AD). Front. Genet. 2013, 4, 307. [Google Scholar] [CrossRef]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded Identification and Characterization of Mammalian Circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef]
- Denzler, R.; Agarwal, V.; Stefano, J.; Bartel, D.P.; Stoffel, M. Assessing the CeRNA Hypothesis with Quantitative Measurements of MiRNA and Target Abundance. Mol. Cell 2014, 54, 766–776. [Google Scholar] [CrossRef] [PubMed]
- He, A.T.; Liu, J.; Li, F.; Yang, B.B. Targeting Circular RNAs as a Therapeutic Approach: Current Strategies and Challenges. Signal Transduct. Target. Ther. 2021, 6, 185. [Google Scholar] [CrossRef] [PubMed]
- Vromman, M.; Vandesompele, J.; Volders, P.J. Closing the Circle: Current State and Perspectives of Circular RNA Databases. Brief. Bioinform. 2021, 22, 288–297. [Google Scholar] [CrossRef]
- Ostolaza, A.; Blanco-Luquin, I.; Urdánoz-Casado, A.; Rubio, I.; Labarga, A.; Zandio, B.; Roldán, M.; Martínez-Cascales, J.; Mayor, S.; Herrera, M.; et al. Circular RNA Expression Profile in Blood According to Ischemic Stroke Etiology. Cell Biosci. 2020, 10, 34. [Google Scholar] [CrossRef]
- Siracusa, C.; Sabatino, J.; Leo, I.; Eyileten, C.; Postuła, M.; De Rosa, S. Circular RNAs in Ischemic Stroke: Biological Role and Experimental Models. Biomolecules 2023, 13, 214. [Google Scholar] [CrossRef]
- Liu, M.; Liu, X.; Zhou, M.; Guo, S.; Sun, K. Impact of CircRNAs on Ischemic Stroke. Aging Dis. 2022, 13, 329–339. [Google Scholar] [CrossRef]
- Mehta, S.L.; Chokkalla, A.K.; Bathula, S.; Arruri, V.; Chelluboina, B.; Vemuganti, R. CDR1as Regulates α-Synuclein-Mediated Ischemic Brain Damage by Controlling MiR-7 Availability. Mol. Ther. Nucleic Acids 2022, 31, 57–67. [Google Scholar] [CrossRef]
- Cervera-Carles, L.; Dols-Icardo, O.; Molina-Porcel, L.; Alcolea, D.; Cervantes-Gonzalez, A.; Muñoz-Llahuna, L.; Clarimon, J. Assessing Circular RNAs in Alzheimer’s Disease and Frontotemporal Lobar Degeneration. Neurobiol. Aging 2020, 92, 7–11. [Google Scholar] [CrossRef]
- Dube, U.; Del-Aguila, J.L.; Li, Z.; Budde, J.P.; Jiang, S.; Hsu, S.; Ibanez, L.; Fernandez, M.V.; Farias, F.; Norton, J.; et al. An Atlas of Cortical Circular RNA Expression in Alzheimer Disease Brains Demonstrates Clinical and Pathological Associations. Nat. Neurosci. 2019, 22, 1903–1912. [Google Scholar] [CrossRef]
- Huang, J.L.; Su, M.; Wu, D.P. Functional Roles of Circular RNAs in Alzheimer’s Disease. Ageing Res. Rev. 2020, 60, 101058. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Alexandrov, P.N.; Jaber, V.; Lukiw, W.J. Deficiency in the Ubiquitin Conjugating Enzyme UBE2A in Alzheimer’s Disease (AD) Is Linked to Deficits in a Natural Circular MiRNA-7 Sponge (CircRNA; CiRS-7). Genes 2016, 7, 116. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liang, R.; Lian, Y.; Zhou, Y. Circ_0002945 Functions as a Competing Endogenous RNA to Promote Aβ25-35-Induced Endoplasmic Reticulum Stress and Apoptosis in SK-N-SH Cells and Human Primary Neurons. Brain Res. 2022, 1785, 147878. [Google Scholar] [CrossRef]
- Chen, J.; Yang, J.; Fei, X.; Wang, X.; Wang, K. CircRNA CiRS-7: A Novel Oncogene in Multiple Cancers. Int. J. Biol. Sci. 2021, 17, 379–389. [Google Scholar] [CrossRef]
- Pluta, R.; Ułamek-Kozioł, M.; Kocki, J.; Bogucki, J.; Januszewski, S.; Bogucka-Kocka, A.; Czuczwar, S.J. Expression of the Tau Protein and Amyloid Protein Precursor Processing Genes in the CA3 Area of the Hippocampus in the Ischemic Model of Alzheimer’s Disease in the Rat. Mol. Neurobiol. 2020, 57, 1281–1290. [Google Scholar] [CrossRef]
- Pluta, R.; Ouyang, L.; Januszewski, S.; Li, Y.; Czuczwar, S.J. Participation of Amyloid and Tau Protein in Post-Ischemic Neurodegeneration of the Hippocampus of a Nature Identical to Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 2460. [Google Scholar]
- Zhang, Y.; Zhao, Y.; Ao, X.; Yu, W.; Zhang, L.; Wang, Y.; Chang, W. The Role of Non-Coding RNAs in Alzheimer’s Disease: From Regulated Mechanism to Therapeutic Targets and Diagnostic Biomarkers. Front. Aging Neurosci. 2021, 13, 654978. [Google Scholar] [CrossRef]
- Bao, M.H.; Szeto, V.; Yang, B.B.; Zhu, S.Z.; Sun, H.S.; Feng, Z.P. Long Non-Coding RNAs in Ischemic Stroke. Cell Death Dis. 2018, 9, 281. [Google Scholar] [CrossRef]
- Vital, S.A.; Gavins, F.N.E. Surgical Approach for Middle Cerebral Artery Occlusion and Reperfusion Induced Stroke in Mice. J. Vis. Exp. 2016, 2016, e54302. [Google Scholar] [CrossRef]
- Shahjouei, S.; Cai, P.Y.; Ansari, S.; Sharififar, S.; Azari, H.; Ganji, S.; Zand, R. Middle Cerebral Artery Occlusion Model of Stroke in Rodents: A Step-by-Step Approach. J. Vasc. Interv. Neurol. 2016, 8, 1–8. [Google Scholar]
- Sasaguri, H.; Nilsson, P.; Hashimoto, S.; Nagata, K.; Saito, T.; De Strooper, B.; Hardy, J.; Vassar, R.; Winblad, B.; Saido, T.C. APP Mouse Models for Alzheimer’s Disease Preclinical Studies. EMBO J. 2017, 36, 2473–2487. [Google Scholar] [CrossRef]
- Amado, B.; Melo, L.; Pinto, R.; Lobo, A.; Barros, P.; Gomes, J.R. Ischemic Stroke, Lessons from the Past towards Effective Preclinical Models. Biomedicines 2022, 10, 2561. [Google Scholar] [CrossRef]
- Mullane, K.; Williams, M. Preclinical Models of Alzheimer’s Disease: Relevance and Translational Validity. Curr. Protoc. Pharmacol. 2019, 84, e57. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, Y.; Xia, J. CircRNAs: Key Molecules in the Prevention and Treatment of Ischemic Stroke. Biomed. Pharmacother. 2022, 156, 113845. [Google Scholar] [CrossRef]
- Vakili, O.; Asili, P.; Babaei, Z.; Mirahmad, M.; Keshavarzmotamed, A.; Asemi, Z.; Mafi, A. Circular RNAs in Alzheimer’s Disease: A New Perspective of Diagnostic and Therapeutic Targets. CNS Neurol. Disord. Drug Targets 2022, 21, 1335–1354. [Google Scholar] [CrossRef]
- Moretti, A.; Ferrari, F.; Villa, R.F. Neuroprotection for Ischaemic Stroke: Current Status and Challenges. Pharmacol. Ther. 2015, 146, 23–34. [Google Scholar] [CrossRef]
- Passeri, E.; Elkhoury, K.; Morsink, M.; Broersen, K.; Linder, M.; Tamayol, A.; Malaplate, C.; Yen, F.T.; Arab-Tehrany, E. Alzheimer’s Disease: Treatment Strategies and Their Limitations. Int. J. Mol. Sci. 2022, 23, 13954. [Google Scholar] [CrossRef]
- Yang, L.; Han, B.; Zhang, Z.; Wang, S.; Bai, Y.; Zhang, Y.; Tang, Y.; Du, L.; Xu, L.; Wu, F.; et al. Extracellular Vesicle-Mediated Delivery of Circular RNA SCMH1 Promotes Functional Recovery in Rodent and Nonhuman Primate Ischemic Stroke Models. Circulation 2020, 142, 556–574. [Google Scholar] [CrossRef]
- Yu, X.; Bai, Y.; Han, B.; Ju, M.; Tang, T.; Shen, L.; Li, M.; Yang, L.; Zhang, Z.; Hu, G.; et al. Extracellular Vesicle-Mediated Delivery of CircDYM Alleviates CUS-Induced Depressive-like Behaviours. J. Extracell. Vesicles 2022, 11, e12185. [Google Scholar] [CrossRef]


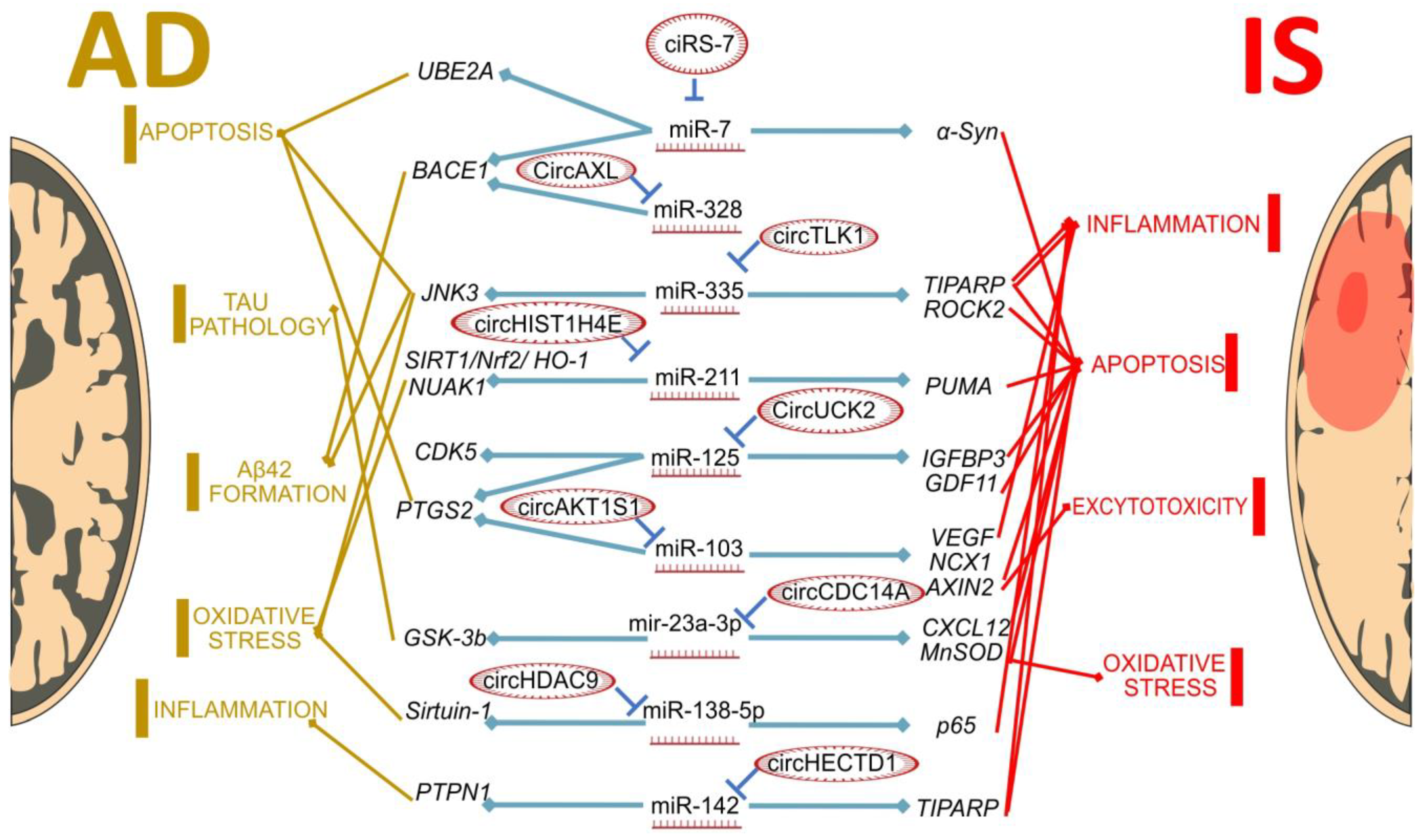{kind=link}
| MiRNA | Disease | Differential Expression Data * | Validated Targets | Target Function | References |
|---|---|---|---|---|---|
| MiR-7 | IS | tMCAO rats ↓ | α-Syn | Promotes neuronal death | [127] |
| AD | PBMCs of AD patients ↑ | BACE1 | Required for the generation of all forms of Aβ, including Aβ42 | [128,129,130,131] | |
| MiR-125 | IS | OGD/R triggered BV2 cells ↓ | IGFBP3 | Promotes apoptosis | [132] |
| GDF11 | Promotes apoptosis via the TGF pathway | [133] | |||
| AD | Neuro2a APPSwe/Δ9 cells; AD patients ↑ | PTGS2, CDK5 | Represses neurite outgrowth but promotes cell apoptosis | [134] | |
| MiR-103 | IS | tMCAO rats ↑ | VEGF | Regulates the angiogenesis | [135] |
| NCX1 | Regulates Ca2+ and Na+ homeostasis | [136] | |||
| AXIN2 | Reduces apoptosis through regulating the mitochondria-associated apoptosis signaling pathway | [137] | |||
| AD | Mouse model of AD, CSF of AD patients ↓ | PTGS2 | Promotes neurite outgrowth and suppresses cells apoptosis | [138] | |
| MiR-142 | IS | AIS patients, primary mouse astrocytes and A172 cells ↓ | TIPARP | Inhibits astrocyte activation via macroautophagy/autophagy | [139] |
| AD | Aβ42-treated SH-SY5Y cells and AD patients ↓ | PTPN1 | Regulates VEGFR2 and Akt signaling | [140] | |
| MiR-211 | IS | OGD/R triggered C12 cells, penumbra of tMCAO mice ↓ | PUMA | Promotes apoptosis | [141] |
| AD | APPswe/PS1ΔE9 mice ↑ | SIRT1/Nrf2/HO-1 | SIRT1 pathway attenuates ROS-induced oxidative stress | [142] | |
| NUAK1 | Regulates cortical neuron differentiation and survival | [143] | |||
| MiR-23-3p | IS | Cortex of C57/BL6 tMCAO mice ↑ | CXCL12 | Suppresses apoptosis | [144] |
| AD | Cortex of APP/PS1 mice ↓ | GSK-3β | Implicated in tau pathology | [145] | |
| MiR-138 | IS | OGD/R induced astrocytes ↓ | p65 | Essential for NF-κB dimerization | [146,147] |
| AD | N2a/APP and HEK293/tau cell lines ↑ | SIRT-1 | Mediates synaptic function and APP processing via macroautophagy/autophagy | [148,149] | |
| MiR-335 | IS | tMCAO mice ↓ | TIPARP | Astrocyte activation | [150] |
| ROCK2 | Promotes apoptosis | [151] | |||
| AD | tissues of AD patients ↓ | JNK3 | Has key role in AD, mediated by many mechanisms | [152] | |
| MiR-328 | IS | Serum of IS patients ↓ | No validated targets in IS conditions | Role in IS development is yet unknown, but connection between serum levels and short-term prognosis of stroke had been reported | [153] |
| AD | SK-N-SH and SK-SY5Y cell lines ↓ | BACE1 | Required for the generation of all forms of Aβ, including Aβ42 | [154] |
| circRNA | Differential Expression Data | Validated Targets | Reported Axis Function | References |
|---|---|---|---|---|
| ciRS-7 (CDR1as) | tMCAO mice ↓ | miR-7/α-Syn (SNCA) | CDR1as overexpression suppressed α-Syn protein induction, promoted motor function recovery, decreased infarct size, curtailed the markers of apoptosis, autophagy and inflammation in the post-stroke brain | [178] |
| CircUCK2 (Circ_001128) | tMCAO mice ↓, OGD HT-22 cells ↓ | miR-125b-5p/ GDF11 | Upregulated circUCK2 levels decreased infarct volumes, attenuated neuronal injury and improved neurological deficits | [133] |
| CircHECTD1 (Circ_0000375) | tMCAO mice ↑, plasma of acute IS patients ↑, OGD-R A172 cells ↑ | miR-142/ TIPARP | circHECTD1 leads to the inhibition of expression with subsequent inhibition of astrocyte activation via macroautophagy/autophagy | [139] |
| CircTLK1 (circ_0004442) | tMCAO mice ↓ | miR-335/ TIPARP | Knockdown of circTLK1 decreased infarct volumes, attenuated neuronal injury, and improved neurological deficits | [150] |
| CircCDC14A | tMCAO mice ↓ | miR-23a-3p/ CXCL12 | Knockdown of circCDC14A suppressed MCAO-induced cerebral infarction and neurological damage, as well as the brain tissue damage and neuronal apoptosis in vivo | [144] |
| circRNA | Differential Expression Data | Validated Targets | Reported Axis Function | References |
|---|---|---|---|---|
| ciRS-7 (CDR1as) | Hippocampal region of AD patients ↓ | miR-7/ UBE2A | Drives amyloid accumulation and the formation of senile plaque deposits | [131,170,182] |
| Circ_0000950 (circAKT1S1) | NGF-stimulated PC 12 cells, primary cerebral cortex neurons from rat embryo cells—no expression changes | miR-103/ PTGS2 | Regulates neuron apoptosis, neurite outgrowth and IL-1β, IL-6 and TNF-α levels | [138] |
| CircHDAC9 | Serum of AD patients and individuals with mild cognitive impairment ↓ | miR-138/ Sirtuin-1 | Mediates synaptic function and APP processing in AD | [148] |
| CircHIST1H4E (circ_0001588) | Streptozotocin-induced rat model of AD ↓ | miR-211-5p/ SIRT1/Nrf2/HO-1 | Control of reactive oxygen species production and malonaldehyde levels, regulates superoxide dismutase and glutathione levels | [142] |
| CircAXL (Circ_0002945) | AD serum and amyloid beta Aβ25-35-stimulated SK-N-SH cells and human primary neurons (HPNs) ↑ | miR-328/ BACE1 | Regulates apoptosis, neurite outgrowth and inflammatory cytokines in cellular AD models | [154] |
| miR-431-5p/ TNFAIP1 | Controls Aβ25-35-induced cell apoptosis and endoplasmic reticulum stress. | [183] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippenkov, I.B.; Khrunin, A.V.; Mozgovoy, I.V.; Dergunova, L.V.; Limborska, S.A. Are Ischemic Stroke and Alzheimer’s Disease Genetically Consecutive Pathologies? Biomedicines 2023, 11, 2727. https://doi.org/10.3390/biomedicines11102727
Filippenkov IB, Khrunin AV, Mozgovoy IV, Dergunova LV, Limborska SA. Are Ischemic Stroke and Alzheimer’s Disease Genetically Consecutive Pathologies? Biomedicines. 2023; 11(10):2727. https://doi.org/10.3390/biomedicines11102727
Chicago/Turabian StyleFilippenkov, Ivan B., Andrey V. Khrunin, Ivan V. Mozgovoy, Lyudmila V. Dergunova, and Svetlana A. Limborska. 2023. "Are Ischemic Stroke and Alzheimer’s Disease Genetically Consecutive Pathologies?" Biomedicines 11, no. 10: 2727. https://doi.org/10.3390/biomedicines11102727
APA StyleFilippenkov, I. B., Khrunin, A. V., Mozgovoy, I. V., Dergunova, L. V., & Limborska, S. A. (2023). Are Ischemic Stroke and Alzheimer’s Disease Genetically Consecutive Pathologies? Biomedicines, 11(10), 2727. https://doi.org/10.3390/biomedicines11102727