Convergent Variations in the Leaf Traits of Desert Plants

 ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. Species Composition, Leaf Traits and Soil Parameters
2.2. Patterns of Leaf Traits among Different Functional Groups
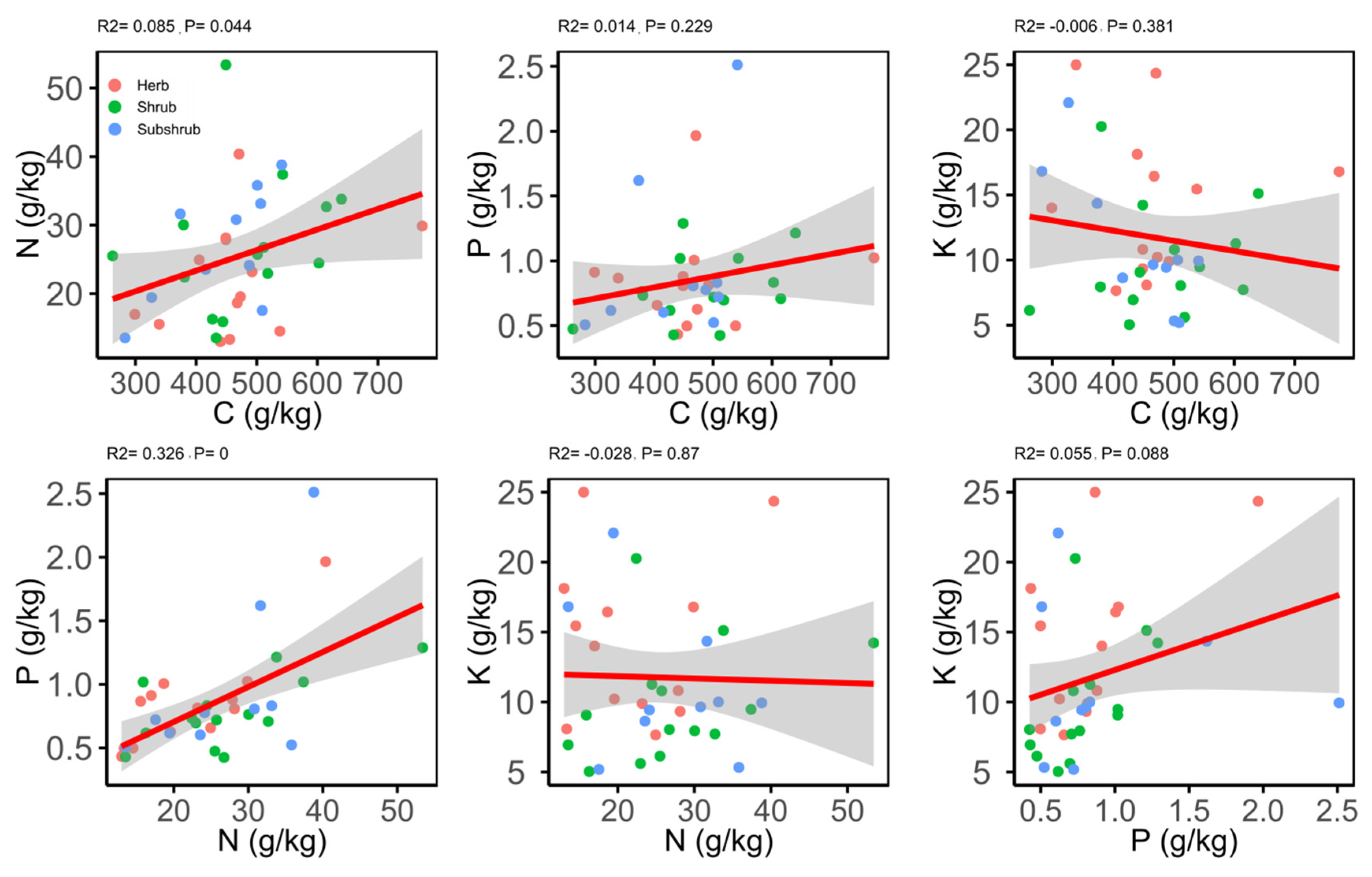
2.3. Correlations between Different Leaf Traits
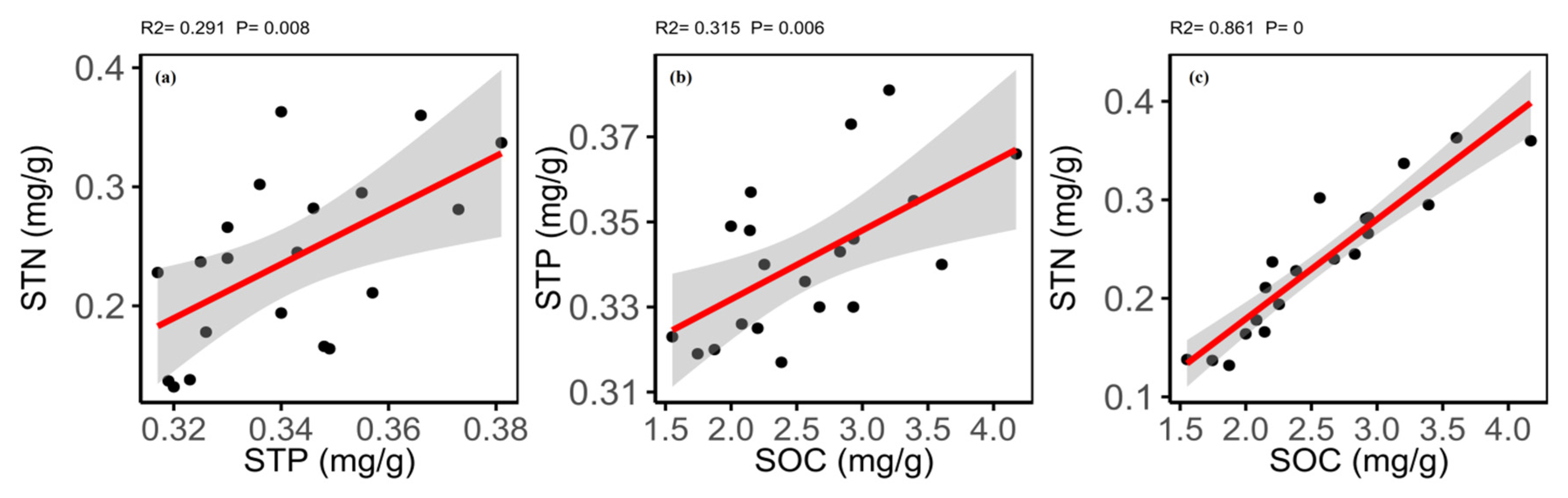
2.4. Relationships between Traits of Plant and Soil
2.5. Taxonomic Effects on Leaf Traits at Species and Family Level
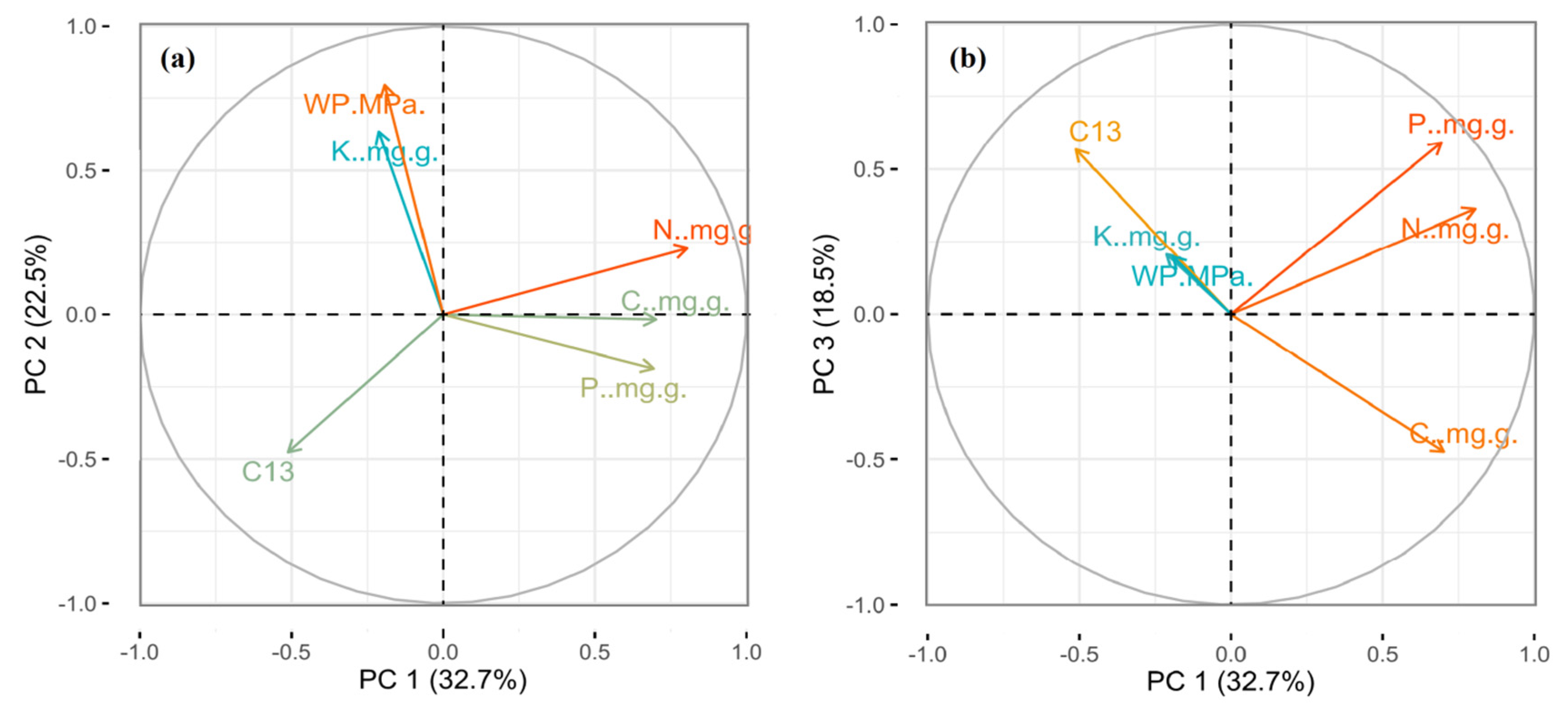
2.6. Partitioning of Variance in Leaf Traits
3. Discussion
3.1. Patterns of Leaf Traits in All Species
3.2. Patterns of Leaf Traits among Different Functional Groups
3.3. Correlations between Different Leaf Traits
3.4. Relationships between the Different Traits of Plant and Soil
3.5. Taxonomic Effect on Leaf Traits
3.6. Partitioning of Variance in Different Leaf Traits
4. Materials and Methods
4.1. Site Description
4.2. Data Collection and Methods
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Losos, J.B. Convergence, adaptation, and constraint. Evolution 2011, 65, 1827–1840. [Google Scholar] [CrossRef] [PubMed]
- Givnish, T. Convergent Evolution, Adaptive Radiation, and Species Diversification in Plants; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Stubbs, W.J.; Wilson, J.B. Evidence for limiting similarity in a sand dune community. J. Ecol. 2004, 92, 557–567. [Google Scholar] [CrossRef]
- Griffiths, H.; Males, J. Succulent plants. Curr. Biol. 2017, 27, R890–R896. [Google Scholar] [CrossRef] [PubMed]
- Kraft, N.J.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Ács, É.; Földi, A.; Vad, C.F.; Trábert, Z.; Kiss, K.T.; Duleba, M.; Borics, G.; Grigorszky, I.; Botta-Dukát, Z. Trait-based community assembly of epiphytic diatoms in saline astatic ponds: A test of the stress-dominance hypothesis. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Botta-Dukát, Z.; Czúcz, B. Testing the ability of functional diversity indices to detect trait convergence and divergence using individual-based simulation. Methods Ecol. Evol. 2016, 7, 114–126. [Google Scholar] [CrossRef]
- Coyle, J.R.; Halliday, F.W.; Lopez, B.E.; Palmquist, K.A.; Wilfahrt, P.A.; Hurlbert, A.H. Using trait and phylogenetic diversity to evaluate the generality of the stress-dominance hypothesis in eastern North American tree communities. Ecography 2014, 37, 814–826. [Google Scholar] [CrossRef]
- Givnish, T.J. On the Economy of Plant Form and Function: Proceedings of the Sixth Maria Moors Cabot Symposium; Cambridge University Press: Cambridge, UK, 1986; Volume 6. [Google Scholar]
- Burns, J.H.; Strauss, S.Y. More closely related species are more ecologically similar in an experimental test. Proc. Natl. Acad. Sci. USA 2011, 108, 5302–5307. [Google Scholar] [CrossRef]
- Reich, P.; Wright, I.; Cavender-Bares, J.; Craine, J.; Oleksyn, J.; Westoby, M.; Walters, M. The evolution of plant functional variation: Traits, spectra, and strategies. Int. J. Plant Sci. 2003, 164, S143–S164. [Google Scholar] [CrossRef]
- Shipley, B.; De Bello, F.; Cornelissen, J.H.C.; Laliberté, E.; Laughlin, D.C.; Reich, P.B. Reinforcing loose foundation stones in trait-based plant ecology. Oecologia 2016, 180, 923–931. [Google Scholar] [CrossRef]
- Merritt, D.M.; Scott, M.L.; LeRoy Poff, N.; Auble, G.T.; Lytle, D.A. Theory, methods and tools for determining environmental flows for riparian vegetation: Riparian vegetation-flow response guilds. Freshw. Biol. 2010, 55, 206–225. [Google Scholar] [CrossRef]
- McCoy-Sulentic, M.E.; Kolb, T.E.; Merritt, D.M.; Palmquist, E.C.; Ralston, B.E.; Sarr, D.A. Variation in species-level plant functional traits over wetland indicator status categories. Ecol. Evol. 2017, 7, 3732–3744. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M. The worldwide leaf economics spectrum. Nature 2004, 428, 821. [Google Scholar] [CrossRef] [PubMed]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Bönisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J. TRY–a global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, É. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Westoby, M.; Wright, I.J. Land-plant ecology on the basis of functional traits. Trends Ecol. Evol. 2006, 21, 261–268. [Google Scholar] [CrossRef]
- Zhang, S.B.; Zhang, J.L.; Slik, J.F.; Cao, K.F. Leaf element concentrations of terrestrial plants across China are influenced by taxonomy and the environment. Glob. Ecol. Biogeogr. 2012, 21, 809–818. [Google Scholar] [CrossRef]
- Edwards, E.J.; Chatelet, D.S.; Sack, L.; Donoghue, M.J. Leaf life span and the leaf economic spectrum in the context of whole plant architecture. J. Ecol. 2014, 102, 328–336. [Google Scholar] [CrossRef]
- Enrique, G.; Olmo, M.; Poorter, H.; Ubera, J.L.; Villar, R. Leaf mass per area (LMA) and its relationship with leaf structure and anatomy in 34 Mediterranean woody species along a water availability gradient. PLoS ONE 2016, 11, e0148788. [Google Scholar]
- Schellenberger Costa, D.; Gerschlauer, F.; Kiese, R.; Fischer, M.; Kleyer, M.; Hemp, A. Plant niche breadths along environmental gradients and their relationship to plant functional traits. Divers. Distrib. 2018, 24, 1869–1882. [Google Scholar] [CrossRef]
- Nielsen, R.L.; James, J.J.; Drenovsky, R.E. Functional traits explain variation in chaparral shrub sensitivity to altered water and nutrient availability. Front. Plant Sci. 2019, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Onoda, Y.; Westoby, M.; Adler, P.B.; Choong, A.M.; Clissold, F.J.; Cornelissen, J.H.; Díaz, S.; Dominy, N.J.; Elgart, A.; Enrico, L. Global patterns of leaf mechanical properties. Ecol. Lett. 2011, 14, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Fridley, J.D.; Lynn, J.S.; Grime, J.; Askew, A. Longer growing seasons shift grassland vegetation towards more-productive species. Nat. Clim. Chang. 2016, 6, 865. [Google Scholar] [CrossRef]
- Bruelheide, H.; Dengler, J.; Purschke, O.; Lenoir, J.; Jiménez-Alfaro, B.; Hennekens, S.M.; Botta-Dukát, Z.; Chytrý, M.; Field, R.; Jansen, F. Global trait–environment relationships of plant communities. Nat. Ecol. Evol. 2018, 2, 1906. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; He, W.; An, L.; Xu, S. Nutrient resorption or accumulation of desert plants with contrasting sodium regulation strategies. Sci. Rep. 2017, 7, 17035. [Google Scholar] [CrossRef]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B.; Vose, J.M.; Gresham, C.; Volin, J.C.; Bowman, W.D. Generality of leaf trait relationships: A test across six biomes. Ecology 1999, 80, 1955–1969. [Google Scholar] [CrossRef]
- Nagy, L.G. Many roads to convergence. Science 2018, 361, 125–126. [Google Scholar]
- Liu, H.-y.; Xu, L.-h.; Chen, C.-d.; Cui, H.-t.; Xu, X.-y. Vegetation patterns and nature reserve construction in an extremely-arid desert in Anxi, NW China’s Gansu Province. J. Environ. Sci. 2002, 14, 380–387. [Google Scholar]
- Liu, F. Vegetation Growth and Conservation Efficacy Assessment in South Part of Gansu Anxi National Nature Reserve in Hyper-Arid Desert. Acta Eco. Sin. 2016, 36. [Google Scholar]
- Tang, Z.; Xu, W.; Zhou, G.; Bai, Y.; Li, J.; Tang, X.; Chen, D.; Liu, Q.; Ma, W.; Xiong, G. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2018, 115, 4033–4038. [Google Scholar] [CrossRef]
- Zhang, B.; Gao, X.; Li, L.; Lu, Y.; Shareef, M.; Huang, C.; Liu, G.; Gui, D.; Zeng, F. Groundwater Depth Affects Phosphorus But Not Carbon and Nitrogen Concentrations of a Desert Phreatophyte in Northwest China. Front. Plant Sci. 2018, 9, 338. [Google Scholar] [CrossRef] [PubMed]
- Abdala-Roberts, L.; Covelo, F.; Parra-Tabla, V.; y Terán, J.C.B.M.; Mooney, K.A.; Moreira, X. Intra-Specific Latitudinal Clines in Leaf Carbon, Nitrogen, and Phosphorus and their Underlying Abiotic Correlates in Ruellia Nudiflora. Sci. Rep. 2018, 8, 596. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wu, G.; Zhang, Y.; Zhou, X. Leaf N and P stoichiometry of 57 plant species in the Karamori Mountain Ungulate Nature Reserve, Xinjiang, China. J. Arid Land 2016, 8, 935–947. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A.; Zhang, K.; Tan, H.; Zhao, Y.; Li, X. Influence of life form, taxonomy, climate, and soil properties on shoot and root concentrations of 11 elements in herbaceous plants in a temperate desert. Plant Soil 2016, 398, 339–350. [Google Scholar] [CrossRef]
- Elser, J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.; Markow, T.; Cotner, J.B.; Harrison, J.; Hobbie, S.E.; Odell, G.; Weider, L. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef]
- Crain, G.; McLaren, J.; Brunner, B.; Darrouzet-Nardi, A. Biologically Available Phosphorus in Biocrust-Dominated Soils of the Chihuahuan Desert. Soil Syst. 2018, 2, 56. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood. Glob. Ecol. Biogeogr. 2013, 22, 494–507. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.; Pearse, I.; Sardans, J.; Sayol, F.; Koenig, W.; LaMontagne, J.; Bogdziewicz, M.; Collalti, A.; Hacket-Pain, A.; Vacchiano, G. Nutrient scarcity as a selective pressure for mast seeding. Nat. Plants 2019, 1–7. [Google Scholar] [CrossRef]
- Ågren, G.I. Stoichiometry and nutrition of plant growth in natural communities. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 153–170. [Google Scholar] [CrossRef]
- Ågren, G.I.; Wetterstedt, J.M.; Billberger, M.F. Nutrient limitation on terrestrial plant growth–modeling the interaction between nitrogen and phosphorus. New Phytol. 2012, 194, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Fernández-Martínez, M.; Ciais, P.; Jou, D.; Piao, S.; Obersteiner, M.; Vicca, S.; Janssens, I.A.; Sardans, J. The bioelements, the elementome, and the biogeochemical niche. Ecology 2019, 100, e02652. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J. Drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct. Ecol. 2007, 21, 191–201. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Martínez, M.; Llusià, J.; Filella, I.; Niinemets, Ü.; Arneth, A.; Wright, I.J.; Loreto, F.; Peñuelas, J. Nutrient-rich plants emit a less intense blend of volatile isoprenoids. New Phytol. 2018, 220, 773–784. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Wright, I.J.; Niklas, K.J.; Hedin, L.; Elser, J.J. Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes. Proc. R. Soc. B 2009, 277, 877–883. [Google Scholar] [CrossRef]
- Yuan, S.; Tang, H. Research advances in the eco-physiological characteristics of ephemerals adaptation to habitats. Acta Prataculturae Sin. 2010, 19, 240–247. [Google Scholar]
- Ameen, F.; AlYahya, S.A.; AlNadhari, S.; Alasmari, H.; Alhoshani, F.; Wainwright, M. Phosphate solubilizing bacteria and fungi in desert soils: Species, limitations and mechanisms. Arch. Agron. Soil Sci. 2019, 65, 1446–1459. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Nahar, K.; Hossain, M.; Mahmud, J.A.; Hossen, M.; Masud, A.A.C.; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 135–189. [Google Scholar]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research progress and perspective on drought stress in legumes: A review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Fang, J.; Reich, P.B.; Ian Woodward, F.; Wang, Z. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. Lett. 2011, 14, 788–796. [Google Scholar] [CrossRef]
- Li, M.; Huang, C.; Yang, T.; Drosos, M.; Wang, J.; Kang, X.; Liu, F.; Xi, B.; Hu, Z. Role of plant species and soil phosphorus concentrations in determining phosphorus: Nutrient stoichiometry in leaves and fine roots. Plant Soil 2019, 445, 231–242. [Google Scholar] [CrossRef]
- Fuentes-Ramirez, A.; Schafer, J.L.; Mudrak, E.L.; Schat, M.; Parag, H.A.; Holzapfel, C.; Moloney, K.A. Spatio-temporal impacts of fire on soil nutrient availability in Larrea tridentata shrublands of the Mojave Desert, USA. Geoderma 2015, 259, 126–133. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, G.; Jia, Y. Foliar δ13C showed no altitudinal trend in an arid region and atmospheric pressure exerted a negative effect on plant δ13C. Front. Plant Sci. 2017, 8, 1070. [Google Scholar] [CrossRef]
- Čada, V.; Šantrůčková, H.; Šantrůček, J.; Kubištová, L.; Seedre, M.; Svoboda, M. Complex physiological response of Norway Spruce to atmospheric pollution–Decreased carbon isotope discrimination and unchanged tree biomass increment. Front. Plant Sci. 2016, 7, 805. [Google Scholar] [CrossRef]
- Kohn, M.J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo) ecology and (paleo) climate. Proc. Natl. Acad. Sci. USA 2010, 107, 19691–19695. [Google Scholar] [CrossRef]
- Ratzmann, G.; Zakharova, L.; Tietjen, B. Optimal leaf water status regulation of plants in drylands. Sci. Rep. 2019, 9, 3768. [Google Scholar] [CrossRef]
- Donovan, L.; Linton, M.; Richards, J. Predawn plant water potential does not necessarily equilibrate with soil water potential under well-watered conditions. Oecologia 2001, 129, 328–335. [Google Scholar] [CrossRef]
- Vesala, T.; Sevanto, S.; Grönholm, T.; Salmon, Y.; Nikinmaa, E.; Hari, P.; Hölttä, T. Effect of leaf water potential on internal humidity and CO2 dissolution: Reverse transpiration and improved water use efficiency under negative pressure. Front. Plant Sci. 2017, 8, 54. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: London, UK; Waltham, MA, USA, 2011. [Google Scholar]
- Reddy, P.S. Breeding for Abiotic Stress Resistance in Sorghum. In Breeding Sorghum for Diverse End Uses; Elsevier: Amsterdam, The Netherlands, 2019; pp. 325–340. [Google Scholar]
- Anderegg, W.R.; Wolf, A.; Arango-Velez, A.; Choat, B.; Chmura, D.J.; Jansen, S.; Kolb, T.; Li, S.; Meinzer, F.; Pita, P. Plant water potential improves prediction of empirical stomatal models. PLoS ONE 2017, 12, e0185481. [Google Scholar] [CrossRef] [PubMed]
- Sandquist, D.R. Plants in deserts. Ecol. Environ. 2014, 297–326. [Google Scholar] [CrossRef]
- Thompson, K.; Parkinson, J.A.; Band, S.R.; Spencer, R.E. A comparative study of leaf nutrient concentrations in a regional herbaceous flora. New Phytol. 1997, 136, 679–689. [Google Scholar] [CrossRef]
- Zhang, Y.; Nan, W.; Zhang, B.; Zhang, J. Species composition, distribution patterns and ecological functions of biological soil crusts in the Gurbantunggut Desert. J. Arid Land 2010, 2, 180–189. [Google Scholar]
- Zhang, H.; Su, P.; Li, S.; Zhou, Z.; Xie, T.; Zhao, Q. Indicative effect of the anatomical structure of plant photosynthetic organ on WUE in desert region. Acta Ecol. Sin. 2013, 33, 4909–4918. [Google Scholar] [CrossRef]
- Taylor, S.H.; Ripley, B.S.; Martin, T.; De-Wet, L.A.; Woodward, F.I.; Osborne, C.P. Physiological advantages of C4 grasses in the field: A comparative experiment demonstrating the importance of drought. Glob. Chang. Biol. 2014, 20, 1992–2003. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, R.; Porder, S.; Balvanera, P.; Edwards, E.J. Ecological and evolutionary variation in community nitrogen use traits during tropical dry forest secondary succession. Ecology 2016, 97, 1194–1206. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.; Ainsworth, E.A.; Leakey, A.D. Will elevated carbon dioxide concentration amplify the benefits of nitrogen fixation in legumes? Plant Physiol. 2009, 151, 1009–1016. [Google Scholar] [CrossRef]
- Liu, A.; Contador, C.A.; Fan, K.; Lam, H.-M. Interaction and regulation of carbon, nitrogen, and phosphorus metabolisms in root nodules of legumes. Front. Plant Sci. 2018, 9, 1860. [Google Scholar] [CrossRef]
- Guo, Y.; Yang, X.; Schöb, C.; Jiang, Y.; Tang, Z. Legume shrubs are more nitrogen-homeostatic than non-legume shrubs. Front. Plant Sci. 2017, 8, 1662. [Google Scholar] [CrossRef]
- Pérez-Fernández, M.; Míguez-Montero, Á.; Valentine, A. Phosphorus and Nitrogen Modulate Plant Performance in Shrubby Legumes from the Iberian Peninsula. Plants 2019, 8, 334. [Google Scholar] [CrossRef] [PubMed]
- Divito, G.A.; Sadras, V.O. How do phosphorus, potassium and sulphur affect plant growth and biological nitrogen fixation in crop and pasture legumes? A meta-analysis. Field Crops Res. 2014, 156, 161–171. [Google Scholar] [CrossRef]
- Garten, C.T., Jr. Correlations between concentrations of elements in plants. Nature 1976, 261, 686. [Google Scholar] [CrossRef]
- Kamilar, J.M.; Cooper, N. Phylogenetic signal in primate behaviour, ecology and life history. Philos. Trans. R. Soc. B 2013, 368, 20120341. [Google Scholar] [CrossRef]
- Jiménez, J.J.; Lorenz, K.; Lal, R. Organic carbon and nitrogen in soil particle-size aggregates under dry tropical forests from Guanacaste, Costa Rica—Implications for within-site soil organic carbon stabilization. Catena 2011, 86, 178–191. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Gou, M.; Lei, P.; Chen, L.; Deng, X. Variations in soil carbon, nitrogen, phosphorus and stoichiometry along forest succession in southern China. Biogeosci. Discuss. 2018. [Google Scholar] [CrossRef]
- Liu, Z.-P.; Shao, M.-A.; Wang, Y.-Q. Spatial patterns of soil total nitrogen and soil total phosphorus across the entire Loess Plateau region of China. Geoderma 2013, 197, 67–78. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.-R.; An, S.-S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Fang, Z.; Li, D.-D.; Jiao, F.; Yao, J.; Du, H.-T. The latitudinal patterns of leaf and soil C: N: P stoichiometry in the Loess Plateau of China. Front. Plant Sci. 2019, 10, 85. [Google Scholar] [CrossRef]
- Hume, A.; Chen, H.Y.; Taylor, A.R.; Kayahara, G.J.; Man, R. Soil C: N: P dynamics during secondary succession following fire in the boreal forest of central Canada. For. Ecol. Manag. 2016, 369, 1–9. [Google Scholar] [CrossRef]
- Wang, Z.; Lu, J.; Yang, M.; Yang, H.; Zhang, Q. Stoichiometric characteristics of carbon, nitrogen, and phosphorus in leaves of differently aged Lucerne (Medicago sativa) Stands. Front. Plant Sci. 2015, 6, 1062. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Wu, X.; Xiang, W.; Fang, X.; Zeng, Y.; Ouyang, S.; Lei, P.; Deng, X.; Peng, C. Spatial variations in soil organic carbon, nitrogen and phosphorus concentrations related to stand characteristics in subtropical areas. Plant Soil 2017, 413, 289–301. [Google Scholar] [CrossRef]
- Ghazali, M.F.; Wikantika, K.; Harto, A.B.; Kondoh, A. Generating soil salinity, soil moisture, soil pH from satellite imagery and its analysis. Inf. Process. Agric. 2019. [Google Scholar] [CrossRef]
- Mohd-Aizat, A.; Mohamad-Roslan, M.; Sulaiman, W.N.A.; Karam, D.S. The relationship between soil pH and selected soil properties in 48 years logged-over forest. Int. J. Environ. Sci. 2014, 4, 1129. [Google Scholar]
- Harmon, M.E.; Fasth, B.; Woodall, C.W.; Sexton, J. Carbon concentration of standing and downed woody detritus: Effects of tree taxa, decay class, position, and tissue type. For. Ecol. Manag. 2013, 291, 259–267. [Google Scholar] [CrossRef]
- Wu, H.; Xiang, W.; Fang, X.; Lei, P.; Ouyang, S.; Deng, X. Tree functional types simplify forest carbon stock estimates induced by carbon concentration variations among species in a subtropical area. Sci. Rep. 2017, 7, 4992. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, L.; He, W.; Zhang, X.; An, L.; Xu, S. Patterns of leaf N: P stoichiometry along climatic gradients in sandy region, north of China. J. Plant Ecol. 2016, 11, 218–225. [Google Scholar] [CrossRef]
- Gutterman, Y. Survival Strategies of Annual Desert Plants; Springer Science & Business Media, Springer GmbH & Co. KG: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Gray, E.F.; Wright, I.J.; Falster, D.S.; Eller, A.S.; Lehmann, C.; Bradford, M.G.; Cernusak, L.A. Leaf: Wood allometry and functional traits together explain substantial growth rate variation in rainforest trees. AoB Plants 2019, 11, plz024. [Google Scholar] [CrossRef]
- Boczulak, S.; Hawkins, B.; Roy, R. Temperature effects on nitrogen form uptake by seedling roots of three contrasting conifers. Tree Physiol. 2013, 34, 513–523. [Google Scholar] [CrossRef]
- Barbosa, E.R.; Tomlinson, K.W.; Carvalheiro, L.G.; Kirkman, K.; de Bie, S.; Prins, H.H.; van Langevelde, F. Short-term effect of nutrient availability and rainfall distribution on biomass production and leaf nutrient content of savanna tree species. PLoS ONE 2014, 9, e92619. [Google Scholar] [CrossRef]
- Gong, Y.; Lv, G.; Guo, Z.; Chen, Y.; Cao, J. Influence of aridity and salinity on plant nutrients scales up from species to community level in a desert ecosystem. Sci. Rep. 2017, 7, 6811. [Google Scholar] [CrossRef] [PubMed]
- Balemi, T.; Negisho, K. Management of soil phosphorus and plant adaptation mechanisms to phosphorus stress for sustainable crop production: A review. J. Soil Sci. Plant Nutr. 2012, 12, 547–562. [Google Scholar] [CrossRef]
- Singh, B.; Schulze, D. Soil minerals and plant nutrition. Nat. Educ. Knowl. 2015, 6, 1. [Google Scholar]
- Zhao, K.; Li, F. The halophytes in China. Chin. Bull. Bot. 1999, 16, 201–207. [Google Scholar]
- Sparks, D.L.; Helmke, P.; Page, A. Methods of Soil Analysis: Chemical Methods; Soil Science Society of America (SSSA) Inc., American Society of Agronomy, Inc.: Madison, WI, USA, 1996. [Google Scholar]
- Lhotsky, B.; Kovács, B.; Ónodi, G.; Csecserits, A.; Rédei, T.; Lengyel, A.; Kertész, M.; Botta-Dukát, Z. Changes in assembly rules along a stress gradient from open dry grasslands to wetlands. J. Ecol. 2016, 104, 507–517.s. [Google Scholar] [CrossRef]
- Twohey, R.J., III; Roberts, L.M.; Studer, A.J. Leaf stable carbon isotope composition reflects transpiration efficiency in Zea mays. Plant J. 2019, 97, 475–484. [Google Scholar] [CrossRef]
- Group, A.P. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar]
- Blomberg, S.P.; Garland, T., Jr.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Code | Family | Life Form | Phylogeny | Photosynthetic Pathway |
|---|---|---|---|---|---|
| Phragmites australis | Pa. | Poaceae | Herb | Monocots | C3 |
| Allium mongolicum | Am. | Amaryllidaceae | Herb | Monocots | C3 |
| Karelinia caspia | Kc. | Asteraceae | Herb | Dicots | C3 |
| Scorzonera pseudodivaricata | Sp. | Asteraceae | Herb | Dicots | C3 |
| Halogeton glomeratus | Hg. | Amaranthaceae | Herb | Dicots | C4 |
| Asterothamnus centraliasiaticus | Ac. | Asteraceae | Herb | Dicots | C3 |
| Artemisia desertorum | Ad. | Asteraceae | Herb | Dicots | C3 |
| Hedysarum scoparium | Hs. | Fabaceae | Herb | Dicots | C3 |
| Sonchus oleraceus | So. | Asteraceae | Herb | Dicots | C3 |
| Salsola ikoikovii | Si. | Amaranthaceae | Herb | Dicots | C4 |
| Hedysarum multijugum | Hm. | Fabaceae | Herb | Dicots | C3 |
| Zygophyllum gobicum | Zg. | Zygophyllaceae | Herb | Dicots | C3 |
| Limonium aureum | La. | Plumbaginaceae | Herb | Dicots | C3 |
| Lycium ruthenicum | Lr. | Solanaceae | Shrub | Dicots | C3 |
| Tamarix chinensis | Tc. | Tamaricaceae | Shrub | Dicots | C3 |
| Elaeagnus angustifolia | Ea. | Elaeagnaceae | Shrub | Dicots | C3 |
| Calligonum mongolicum | Cm. | Polygonaceae | Shrub | Dicots | C4 |
| Artemisia frigida | Af. | Asteraceae | Shrub | Dicots | C3 |
| Gymnocarpos przewalskii | Gp. | Caryophyllaceae | Shrub | Dicots | C3 |
| Sarcozygium xanthoxylon | Sx. | Zygophyllaceae | Shrub | Dicots | C3 |
| Xanthoceras sorbifolia | Xs. | Sapindaceae | Shrub | Dicots | C3 |
| Salsola arbuscula | Sa. | Amaranthaceae | Shrub | Dicots | C4 |
| Caragana roborovskyi | Cr. | Fabaceae | Shrub | Dicots | C3 |
| Caragana korshinskii | Ck. | Fabaceae | Shrub | Dicots | C3 |
| Reaumuria songarica | Rs. | Tamaricaceae | Shrub | Dicots | C3 |
| Ammopiptanthus mongolicus | Amo. | Fabaceae | Shrub | Dicots | C3 |
| Nitraria sphaerocarpa | Ns. | Nitrariaceae | Shrub | Dicots | C3 |
| Caryopteris mongholia | Cam. | Lamiaceae | Subshrub | Dicots | C3 |
| Alhagi sparsifolia | As. | Fabaceae | Subshrub | Dicots | C3 |
| Clematis tangutica | Ct. | Ranunculaceae | Subshrub | Dicots | C3 |
| Bassia dasyphylla | Bd. | Amaranthaceae | Subshrub | Dicots | C3 |
| Cynanchum chinense | Cc. | Apocynaceae | Subshrub | Dicots | C3 |
| Sympegma regelii | Sr. | Amaranthaceae | Subshrub | Dicots | C3 |
| Ephedra przewalskii | Ep. | Ephedraceae | Subshrub | Gymnosperm | C3 |
| Sophora alopecuroides | Sa. | Fabaceae | Subshrub | Dicots | C3 |
| Ephedra sinica | Es. | Ephedraceae | Subshrub | Gymnosperm | C3 |
| Haloxylon ammodendron | Ha. | Amaranthaceae | Subshrub | Dicots | C4 |
| Life Form | Statistic | Leaf Traits | |||||
|---|---|---|---|---|---|---|---|
| C (mg g−1) | N (mg g−1) | P (mg g−1) | K (mg g−1) | δ13C | LWP (MPa) | ||
| Herb | Mean | 465.47 | 22.00 | 0.85 | 8.38 | −24.88 | −8.66 |
| (n = 13) | G. Mean | 454.53 | 20.75 | 0.78 | 7.05 | - | - |
| Max | 772.99 | 40.38 | 1.97 | 12.12 | −13.63 | −3.01 | |
| Min | 298.86 | 12.99 | 0.43 | 4.32 | −28.98 | −19.60 | |
| SE | 30.94 | 2.23 | 0.11 | 0.65 | 1.40 | 1.24 | |
| CV | 0.24 | 0.37 | 0.46 | 0.28 | −0.20 | −0.52 | |
| Shrub | Mean | 478.97 | 27.20 | 0.78 | 14.91 | −24.47 | −7.29 |
| (n = 14) | G. Mean | 467.52 | 25.57 | 0.74 | 14.06 | - | - |
| Max | 639.65 | 53.40 | 1.29 | 26.35 | −14.76 | −4.24 | |
| Min | 262.25 | 13.52 | 0.42 | 8.08 | −27.59 | −16.33 | |
| SE | 27.75 | 2.73 | 0.07 | 1.47 | 1.05 | 1.00 | |
| CV | 0.22 | 0.38 | 0.35 | 0.37 | −0.16 | −0.51 | |
| Subshrub | Mean | 441.01 | 26.84 | 0.95 | 11.16 | −24.63 | −6.96 |
| (n = 10) | G. Mean | 432.29 | 25.53 | 0.83 | 10.03 | - | - |
| Max | 541.11 | 38.79 | 2.51 | 22.08 | −13.62 | −3.05 | |
| Min | 282.72 | 13.54 | 0.51 | 4.13 | −28.13 | −11.00 | |
| SE | 27.61 | 2.66 | 0.20 | 1.70 | 1.31 | 0.95 | |
| CV | 0.20 | 0.31 | 0.67 | 0.48 | −0.17 | −0.43 | |
| All | Mean | 463.97 | 25.27 | 0.85 | 11.60 | −24.66 | −7.68 |
| (n = 37) | SE | 9.83 | 0.87 | 0.04 | 0.50 | 0.41 | 0.37 |
| ANOVA Results | |||||||
| Life form | F | 0.40 | 1.32 | 0.45 | 6.87 | 0.03 | 0.67 |
| P | 0.67 | 0.28 | 0.64 | 0.00 | 0.97 | 0.52 | |
| Statistic | Soil Parameters | ||||||
|---|---|---|---|---|---|---|---|
| SOC (mg g−1) | STN (mg g−1) | STP (mg g−1) | SAP (mg g−1) | SWC | pH | SEC (µS/cm) | |
| Mean | 2.58 | 0.239 | 0.343 | 5.24 | 0.456 | 8.97 | 485.29 |
| SE | 1.49 | 0.138 | 0.198 | 3.02 | 0.263 | 5.18 | 280.19 |
| CV | 0.26 | 0.305 | 0.053 | 0.58 | 0.580 | 0.03 | 0.57 |
| Plant Category | Statistic | Leaf Traits | |||||
|---|---|---|---|---|---|---|---|
| C (mg g−1) | N (mg g−1) | P (mg g−1) | K (mg g−1) | δ13C | LWP (MPa) | ||
| C3 | Mean | 481.92 | 26.59 | 0.86 | 11.63 | −26.25 | −7.45 |
| (n = 32) | SE | 16.60 | 1.60 | 0.08 | 0.96 | 0.24 | 0.60 |
| CV | 0.19 | 0.34 | 0.53 | 0.47 | −0.05 | −0.46 | |
| C4 | Mean | 349.08 | 16.87 | 0.81 | 11.45 | −14.48 | −9.14 |
| (n = 5) | SE | 29.21 | 1.49 | 0.09 | 2.06 | 0.52 | 2.70 |
| CV | 0.19 | 0.20 | 0.24 | 0.40 | −0.08 | −0.66 | |
| ANOVA Results | |||||||
| F | 9.20 | 5.53 | 0.05 | 0.00 | 336.07 | 0.85 | |
| P | 0.00 | 0.02 | 0.82 | 0.95 | 0.00 | 0.36 | |
| Legumes | Mean | 488.89 | 27.00 | 0.61 | 12.61 | −25.83 | −6.79 |
| (n = 7) | SE | 26.40 | 2.09 | 0.04 | 2.55 | 0.57 | 1.42 |
| CV | 14.29 | 20.45 | 17.66 | 53.56 | −5.88 | −55.13 | |
| Nonlegumes | Mean | 458.15 | 24.87 | 0.91 | 11.37 | −24.38 | −7.88 |
| (n = 30) | SE | 19.53 | 1.79 | 0.08 | 0.91 | 0.85 | 0.70 |
| CV | 23.35 | 39.40 | 50.12 | 43.96 | −19.20 | −48.84 | |
| ANOVA Results | |||||||
| F | 0.52 | 0.30 | 2.91 | 0.31 | 0.64 | 0.46 | |
| P | 0.48 | 0.58 | 0.10 | 0.58 | 0.43 | 0.50 | |
| C | N | P | K | δ13C | LWP | |
|---|---|---|---|---|---|---|
| C | 0.350 * | 0.341 * | 0.033 | −0.099 | −0.180 | |
| N | 0.333 * | 0.709 ** | −0.048 | −0.417 * | 0.059 | |
| P | 0.201 | 0.586 ** | −0.494 ** | −0.332 * | −0.409 * | |
| K | −0.069 | 0.023 | −0.154 | −0.024 | 0.610 ** | |
| δ13C | −0.404 * | −0.294 | 0.004 | 0.038 | −0.135 | |
| LWP | −0.243 | 0.052 | −0.122 | 0.258 | –0.162 |
| Leaf Traits | Total Effects (r2, %) | ||
|---|---|---|---|
| Full | Taxonomy | Soil | |
| C | 65.61 | 57.76 | 7.84 |
| N | 82.02 | 73.65 | 8.37 |
| P | 90.38 | 82.18 | 8.19 |
| K | 39.32 | 35.44 | 3.88 |
| δ13C | 94.87 | 76.45 | 18.42 |
| LWP | 31.14 | 30.84 | 0.30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akram, M.A.; Wang, X.; Hu, W.; Xiong, J.; Zhang, Y.; Deng, Y.; Ran, J.; Deng, J. Convergent Variations in the Leaf Traits of Desert Plants. Plants 2020, 9, 990. https://doi.org/10.3390/plants9080990
Akram MA, Wang X, Hu W, Xiong J, Zhang Y, Deng Y, Ran J, Deng J. Convergent Variations in the Leaf Traits of Desert Plants. Plants. 2020; 9(8):990. https://doi.org/10.3390/plants9080990
Chicago/Turabian StyleAkram, Muhammad Adnan, Xiaoting Wang, Weigang Hu, Junlan Xiong, Yahui Zhang, Yan Deng, Jinzhi Ran, and Jianming Deng. 2020. "Convergent Variations in the Leaf Traits of Desert Plants" Plants 9, no. 8: 990. https://doi.org/10.3390/plants9080990
APA StyleAkram, M. A., Wang, X., Hu, W., Xiong, J., Zhang, Y., Deng, Y., Ran, J., & Deng, J. (2020). Convergent Variations in the Leaf Traits of Desert Plants. Plants, 9(8), 990. https://doi.org/10.3390/plants9080990