Shading Affects Yield, Elemental Composition and Antioxidants of Perennial Wall Rocket Crops Grown from Spring to Summer in Southern Italy



 ,
,  ,
,  ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
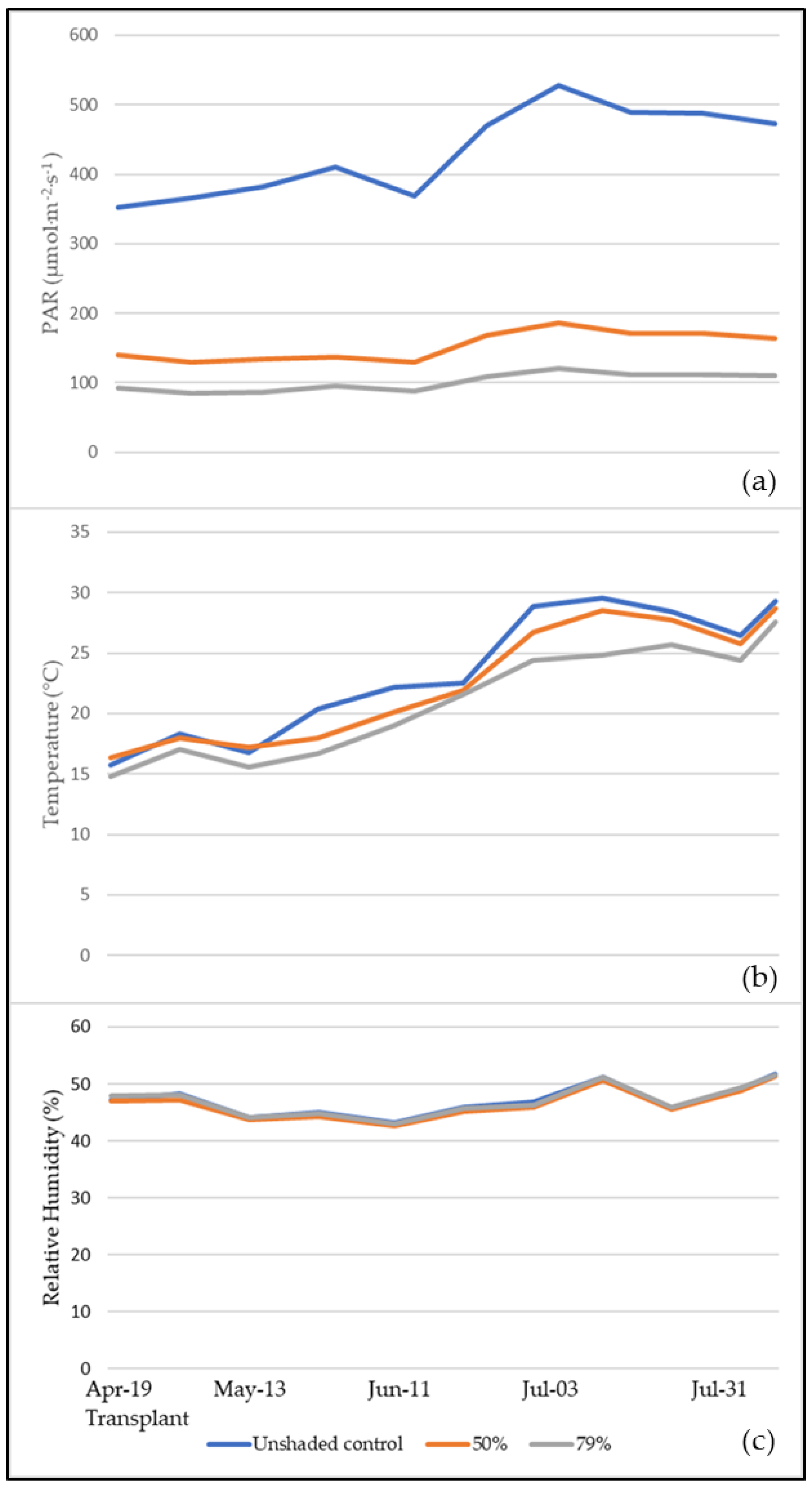
2.1. Meteorological Parameters
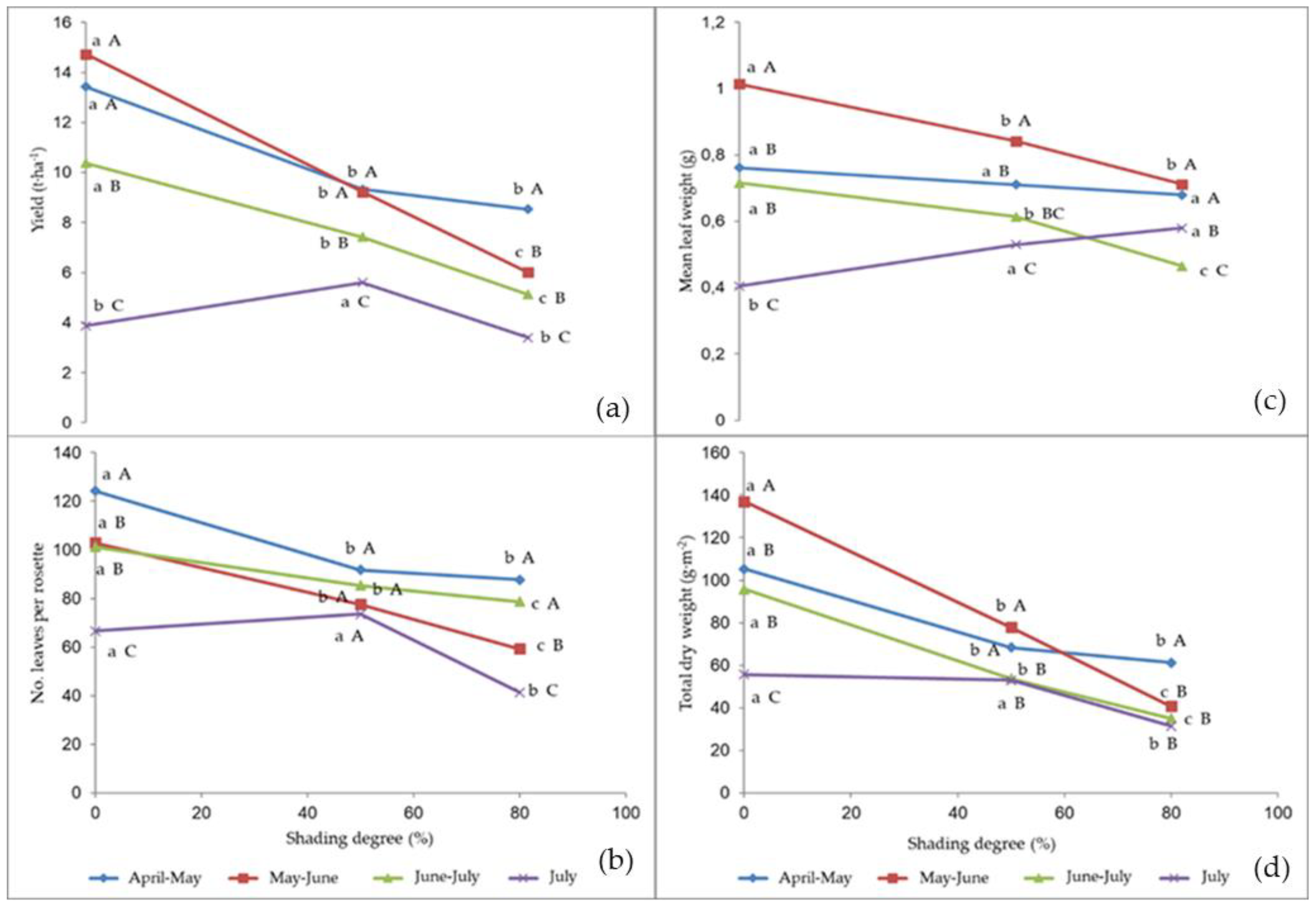
2.2. Plant Growth and Yield
2.3. Leaf Color Parameters and Chemical Composition
2.4. Antioxidant Compounds and Activity
3. Materials and Methods
3.1. Growing Conditions and Experimental Protocol
3.2. Dry Weight
3.3. Leaf Colorimetric Parameters
3.4. Mineral Elements
3.5. Antioxidant Compounds and Activity
3.6. Statistical Processing
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martínez-Sánchez, A.; Llorach, R.; Gil, M.I.; Ferreres, F. Identification of new flavonoid glycosides and flavonoid profiles to characterize rocket leafy salads (Eruca vesicaria and Diplotaxis tenuifolia). J. Agric. Food Chem. 2007, 55, 1356–1363. [Google Scholar] [CrossRef]
- Bennett, R.N.; Rosa, E.A.; Mellon, F.A.; Kroon, P.A. Ontogenic profiling of glucosinolates, flavonoids, and other secondary metabolites in Eruca sativa (salad rocket), Diplotaxis erucoides (wall rocket), Diplotaxis tenuifolia (wild rocket), and Bunias orientalis (Turkish rocket). J. Agrci. Food Chem. 2006, 54, 4005–4015. [Google Scholar] [CrossRef]
- Winkler, S.; Faragher, J.; Franz, P.; Imsic, M.; Jones, R. Glucoraphanin and flavonoid levels remain stable during simulated transport and marketing of broccoli (Brassica oleracea var. italica) heads. Postharvest Biol. Technol. 2007, 43, 89–94. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lelandais, M.; Kunert, K.J. Photooxidative stress in plants. Phys. Plant. 1994, 92, 696–717. [Google Scholar] [CrossRef]
- Lefsrud, M.G.; Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Air temperature affects biomass and carotenoid pigment accumulation in kale and spinach grown in a controlled environment. HortScience 2005, 40, 2026–2030. [Google Scholar] [CrossRef]
- Gruda, N. Impact of environmental factors on product quality of greenhouse vegetables for fresh consumption. Crit. Rev. Plant. Sci. 2005, 24, 227–247. [Google Scholar] [CrossRef]
- Ayala-Tafoya, F.; Zatarain-López, D.M.; Valenzuela-López, M.; Partida-Ruvalcaba, L.; Velázquez-Alcaraz, T.D.J.; Díaz-Valdés, T.; Osuna-Sánchez, J.A. Growth and yield of tomato in response to sun radiation transmitted by shade nets. Terra Latinoam. 2011, 29, 403–410. [Google Scholar]
- Castellano, S.; Scarascia-Mugnozza, G.; Russo, G.; Briassoulis, D.; Mistriotis, A.; Hemming, S.; Waaijenberg, D. Plastic nets in agriculture: A general review of types and applications. Appl. Eng. Agric. 2008, 24, 799–808. [Google Scholar] [CrossRef]
- Ben-Yakir, D.; Hadar, M.D.; Offir, Y.; Chen, M.; Tregerman, M. Protecting crops from pests using OptiNet (R) screens and ChromatiNet (R) shading nets. Acta Hortic. 2008, 770, 205–212. [Google Scholar] [CrossRef]
- Elad, Y.; Messika, Y.; Brand, M.; David, D.R.; Sztejnberg, A. Effect of colored shade nets on pepper powdery mildew (Leveillula taurica). Phytoparasit. 2007, 35, 285–299. [Google Scholar] [CrossRef]
- Shahak, Y. Photo-selective netting for improved performance of horticultural crops. A review of ornamental and vegetable studies carried out in israel. Acta Hortic. 2008, 770, 161–168. [Google Scholar] [CrossRef]
- Stamps, R.H. Use of colored shade netting in horticulture. HortScience 2009, 44, 239–241. [Google Scholar] [CrossRef]
- Ferreira, T.; Valadares, K.; Souza, M.; Santana, J.; Balbino, M.; Ferreira, R. Yellow and red sweet pepper quality under photoselective screens and field crop conditions. Acta Hortic. 2012, 956, 473–479. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Milenković, L.; Šunić, L.; Manojlović, M. Color shade nets improve vegetables quality at harvest and maintain quality during storage. Contemp. Agric. 2018, 67, 9–19. [Google Scholar] [CrossRef]
- Mashabela, M.N.; Selahle, K.M.; Soundy, P.; Crosby, K.M.; Sivakumar, D. Bioactive compounds and fruit quality of green sweet pepper grown under different coloured shade netting during postharvest storage. J. Food Sci. 2015, 80, 2612–2618. [Google Scholar] [CrossRef] [PubMed]
- Selahle, K.M.; Sivakumar, D.; Jifon, J.; Soundy, P. Postharvest responses of red and yellow sweet peppers grown under photo-selective nets. Food Chem. 2015, 173, 951–956. [Google Scholar] [CrossRef]
- Sivakumar, D.; Jifon, J. Influence of photoselective shade nettings on postharvest quality of vegetables. In Preharvest Modulation of Postharvest Fruit and Vegetable Quality; Elsevier BV: Duivendrecht, The Netherlands, 2018; pp. 121–138. [Google Scholar]
- Jin, J.; Koroleva, O.A.; Gibson, T.; Swanston, J.; Magan, J.; Zhang, Y.; Rowland, I.R.; Wagstaff, C. Analysis of phytochemical composition and chemoprotective capacity of rocket (Eruca sativa and Diplotaxis tenuifolia) leafy salad following cultivation in different environments. J. Agric. Food Chem. 2009, 57, 5227–5234. [Google Scholar] [CrossRef]
- Francke, A. Effect of flat covers on macronutrient concentrations in arugula leaves. J. Elementology 2014, 19, 351–360. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Fallik, E. Light quality manipulation improves vegetable quality at harvest and postharvest: A review. Environ. Exp. Bot. 2017, 139, 79–90. [Google Scholar] [CrossRef]
- Justen, V.L.; Fritz, V.A.; Cohen, J.D. Seasonal variation in glucosinolate accumulation in turnips grown under photoselective nettings. Hortic. Environ. Biotechnol. 2012, 53, 108–115. [Google Scholar] [CrossRef]
- Ombodi, A.; Zoltán, P.É.K.; Szuvandzsiev, P.; Taskovics, Z.T.; Kohazi-Kis, A.; Kovacs, A.; Helyes, L. Effects of external coloured shade nets on sweet peppers cultivated in walk-in plastic tunnels. Not. Bot. Hort. Agrobot. Cluj-Nap. 2015, 43, 398–403. [Google Scholar] [CrossRef]
- Caruso, G.; Cozzolino, E.; Cuciniello, A.; Maiello, R.; Cenvinzo, V.; Giordano, M.; De Pascale, S.; Rouphael, Y. Yield and quality of greenhouse organic pepper as affected by shading net in Mediterranean area. Acta Hortic. 2020, 335–340. [Google Scholar] [CrossRef]
- Ueno, O.; Bang, S.W.; Wada, Y.; Kondo, A.; Ishihara, K.; Kaneko, Y.; Matsuzawa, Y. Structural and biochemical dissection of photorespiration in hybrids differing in genome constitution between Diplotaxis tenuifolia (C3-C4) and radish (C3). Plant. Physiol. 2003, 132, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Flaishman, M.; Peles, Y.; Dahan, Y.; Milo-Cochavi, S.; Frieman, A.; Naor, A. Differential response of cell-cycle and cell-expansion regulators to heat stress in apple (Malus domestica) fruitlets. Plant. Sci. 2015, 233, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Bonasia, A.; Lazzizera, C.; Elia, A.; Conversa, G. Nutritional, biophysical and physiological characteristics of wild rocket genotypes as affected by soilless cultivation system, salinity level of nutrient solution and growing period. Front. Plant. Sci. 2017, 8, 300. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Stoleru, V.; Giordano, M.; Cozzolino, E.; Pannico, A.; Giordano, M.; Teliban, G.; Cuciniello, A.; Rouphael, Y. Production, leaf quality and antioxidants of perennial wall rocket as affected by crop cycle and mulching type. Agronomy 2019, 9, 194. [Google Scholar] [CrossRef]
- Jaimez, R.E.; Rada, F. Flowering and fruit production dynamics of sweet pepper (Capsicum chinense Jacq) under different shade conditions in a humid tropical region. J. Sustain. Agric. 2006, 27, 97–108. [Google Scholar] [CrossRef]
- Arthurs, S.P.; Stamps, R.H.; Giglia, F.F. Environmental modification inside photoselective shadehouses. HortScience 2013, 48, 975–979. [Google Scholar] [CrossRef]
- Ilić, S.; Milenković, L.; Dimitrijević, A.; Stanojević, L.; Cvetković, D.; Kevrešan, Ž.; Fallik, E.; Mastilović, J. Light modification by color nets improve quality of lettuce from summer production. Sci. Hortic. 2017, 226, 389–397. [Google Scholar] [CrossRef]
- Greer, D.H.; Berry, J.A.; Björkman, O. Photoinhibition of photosynthesis in intact bean leaves: Role of light and temperature, and requirement for chloroplast-protein synthesis during recovery. Planta 1986, 168, 253–260. [Google Scholar]
- Yamori, W.; Masumoto, C.; Fukayama, H.; Makino, A. Rubisco activase is a key regulator of non-steady-state photosynthesis at any leaf temperature and, to a lesser extent, of steady-state photosynthesis at high temperature. Plant. J. 2012, 71, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Noguchi, K.; Kashino, Y.; Terashima, I. The role of electron transport in determining the temperature dependence of the photosynthetic rate in spinach leaves grown at contrasting temperatures. Plant. Cell Physiol. 2008, 49, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Von Caemmerer, S.; Evans, J.R. Temperature responses of mesophyll conductance differ greatly between species. Plant., Cell Environ. 2014, 38, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, P.; Gonnella, M.; Elia, A.; Parente, A.; Serio, F. Ways of reducing rocket salad nitrate content. Acta Hortic. 2001, 529–536. [Google Scholar] [CrossRef]
- Padulosi, S.; Pignone, D. Rocket: A Mediterranean Crop for the World. In Report of A Workshop 13–14 December 1996; Bioversity International: Rome, Italy, 1997; p. 97. [Google Scholar]
- Wolff, X.Y.; Coltman, R.R. Productivity of eight leafy vegetable crops grown under shade in Hawaii. J. Am. Soc. Hortic. Sci. 1990, 115, 182–188. [Google Scholar] [CrossRef]
- Kavga, A.; Trypanagnostopoulos, G.; Zervoudakis, G.; Tripanagnostopoulos, Y. Growth and physiological characteristics of lettuce (Lactuca sativa L.) and rocket (Eruca sativa Mill.) plants cultivated under photovoltaic panels. Not. Bot. Horti Agrobot. Cluj-Napoc. 2018, 46, 206–212. [Google Scholar] [CrossRef]
- Tindall, J.A.; Mills, H.; Radcliffe, D. The effect of root zone temperature on nutrient uptake of tomato. J. Plant. Nutr. 1990, 13, 939–956. [Google Scholar] [CrossRef]
- Gregory, P.J. Growth and functioning of plant roots. In Russell’s Soil Conditions and Plant Growth; Wild, A., Russell, J., Eds.; Longman Scientific and Technical: Essex, UK, 1988; pp. 113–167. [Google Scholar]
- Stagnari, F.; Galieni, A.; Pisante, M. Shading and nitrogen management affect quality, safety and yield of greenhouse-grown leaf lettuce. Sci. Hortic. 2015, 192, 70–79. [Google Scholar] [CrossRef]
- Díaz-Pérez, J.C. Bell pepper (Capsicum annum L.) crop as affected by shade level: Microenvironment, plant growth, leaf gas exchange, and leaf mineral nutrient concentration. HortScience 2013, 48, 175–182. [Google Scholar] [CrossRef]
- Zhao, D.; Oosterhuis, D.M. Influence of shade on mineral nutrient status of field-grown cotton. J. Plant. Nutr. 1998, 21, 1681–1695. [Google Scholar] [CrossRef]
- Chen, K.; Hu, G.; Lenz, F. Apple yield and quality as affected by training and shading. Acta Hortic. 1998, 466, 53–58. [Google Scholar] [CrossRef]
- Stagnari, F.; Galieni, A.; Cafiero, G.; Pisante, M. Application of photo-selective films to manipulate wavelength of transmitted radiation and photosynthate composition in red beet (Beta vulgaris var. conditiva Alef.). J. Sci. Food Agric. 2013, 94, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Conti, S.; La Rocca, G. Influence of crop cycle and nitrogen fertilizer form on yield and nitrate content in different species of vegetables. Adv. Hortic. Sci. 2011, 25, 81–89. [Google Scholar]
- Potter, J.D. Vegetables, fruit, and cancer. Lancet 2005, 366, 527–530. [Google Scholar] [CrossRef]
- Steingröver, E.; Ratering, P.; Siesling, J. Daily changes in uptake, reduction and storage of nitrate in spinach grown at low light intensity. Physiol. Plant. 1986, 66, 550–556. [Google Scholar] [CrossRef]
- Behr, U.; Wiebe, H.J. Relation between photosynthesis and nitrate content of lettuce cultivars. Sci. Hortic. 1992, 49, 175–179. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
- Tamura, Y. Environmental changes and genetic variation of accumulation of bioactive compounds in plantain (Plantago lanceolata L.). Bull. Nat. Agric. Res. Cent. Reg. 2002, 100, 75–92. [Google Scholar]
- Luthria, D.L. A simplified UV spectral scan method for the estimation of phenolic acids and antioxidant capacity in eggplant pulp extracts. J. Funct. Foods 2012, 4, 238–242. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2014, 95, 869–877. [Google Scholar] [CrossRef]
- Cano, A.; Arnao, M.B. Hydrophilic and lipophilic antioxidant activity in different leaves of three lettuce varieties. Int. J. Food Prop. 2005, 8, 521–528. [Google Scholar] [CrossRef]
- Zdravković, J.; Pavlović, N.; Girek, Z.; Zdravkovic, M.; Cvikic, D. Characteristics important for organic breeding of vegetable crops. Org. Agric. Pract. 2014, 211–222. [Google Scholar] [CrossRef]
- Crowe, F.; Roddam, A.W.; Key, T.J.; Appleby, P.N.; Overvad, K.; Jakobsen, M.U.; Tjønneland, A.; Hansen, L.; Boeing, H.; Weikert, C.; et al. Fruit and vegetable intake and mortality from ischaemic heart disease: Results from the European Prospective Investigation into Cancer and Nutrition (EPIC)-Heart study. Eur. Heart J. 2011, 32, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Yen, G.C. Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: Phenolic acids, monophenol, polyphenol, and their derivatives. Cancer Treat. Rev. 2012, 38, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Commenges, D.; Scotet, V.; Renaud, S.; Jacqmin-Gadda, H.; Barberger-Gateau, P.; Dartigues, J.F. Intake of flavonoids and risk of dementia. Eur. J. Epidemiol. 2000, 16, 357–363. [Google Scholar] [CrossRef]
- Dai, Q.; Borenstein, A.R.; Wu, Y.; Jackson, J.C.; Larson, E.B. Fruit and vegetable juices and Alzheimer’s Disease: The Kame project. Am. J. Med. 2006, 119, 751–759. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and human health: Prevention of disease and mechanisms of action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef]
- Oh, M.M.; Carey, E.; Rajashekar, C.B. Antioxidant phytochemicals in lettuce grown in high tunnels and open field. Hortic. Environ. Biotechnol. 2011, 52, 133–139. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Zhang, Y.Y.; Zhao, X.; Yu, H.J.; Shi, K.; Yu, J.Q. Impact of light variation on development of photoprotection, antioxidants, and nutritional value in Lactuca sativa L. J. Agric. Food Chem. 2009, 57, 5494–5500. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.M.; Carey, E.; Rajashekar, C. Environmental stresses induce health-promoting phytochemicals in lettuce. Plant. Physiol. Biochem. 2009, 47, 578–583. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, S.; He, X.; Li, Y.; Zhang, Y.; Chen, W. Response of total phenols, flavonoids, minerals, and amino acids of four edible fern species to four shading treatments. Plant Biol. 2020, 8, e8354. [Google Scholar] [CrossRef]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Boil. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef]
- Kosma, C.; Triantafyllidis, V.; Papasavvas, A.; Salahas, G.; Patakas, A. Yield and nutritional quality of greenhouse lettuce as affected by shading and cultivation season. Emir. J. Food Agric. 2013, 25, 974–979. [Google Scholar] [CrossRef]
- Hamner, K.C.; Bernstein, L.; Maynard, L.A. Effects of light intensity, day length, temperature, and other environmental factors on the ascorbic acid content of tomatoes. J. Nutr. 1945, 29, 85–97. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Yu, J.; Gómez, F.; Fernández, L.; McIntosh, L.; Foyer, C.H. Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J. Exp. Bot. 2006, 57, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Requirements. In FAO Irrigation and Drainage Paper No. 56.; FAO: Rome, Italy, 1998. [Google Scholar]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis; Black, C.A., Evans, D.D., White, I.L., Ensminger, L.E., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1149–1178. [Google Scholar]
- Kampfenkel, K.; Vanmontagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Experimental Treatment | Cycle Length (Days from Transplant) | Yield (t·ha−1) | Number of Leaves per Rosette | Mean Weight (g) | Total Dry Matter (g·m−2) |
|---|---|---|---|---|---|
| Cropping season | |||||
| April–May | 27.0 ± 3.6 b | 10.43 ± 1.61 a | 101.2 ± 16.2 a | 0.72 ± 0.06 b | 78.3 ± 13.3 a |
| May–June | 31.7 ± 3.1 a | 9.99 ± 3.54 a | 79.9 ± 17.8 b | 0.86 ± 0.15 a | 85.3 ± 34.2 a |
| June–July | 25.0 ± 2.6 b,c | 7.65 ± 1.65 b | 88.5 ± 16.8 b | 0.60 ± 0.10 c | 61.6 ± 16.6 b |
| July | 24.0 ± 3.6 c | 4.29 ± 1.19 c | 60.6 ± 15.4 c | 0.50 ± 0.08 d | 46.7 ± 7.8 c |
| Shading degree (%) | |||||
| Unshaded control | 24.3 ± 3.9 b | 12.16 ± 3.41 a | 106.1 ± 14.3 a | 0.80 ± 0.24 a | 104.1 ± 29.0 a |
| 50 | 26.0 ± 3.6 b | 8.42 ± 1.21 b | 84.0 ± 7.5 b | 0.71 ± 0.11 b | 61.6 ± 9.7 b |
| 79 | 30.5 ± 3.3 a | 6.31 ± 1.49 c | 72.7 ± 17.5 c | 0.62 ± 0.11 c | 42.2 ± 11.4 c |
| Treatment | L* | a* | b* |
|---|---|---|---|
| Cropping Season | |||
| April–May | 41.1 ± 1.0 b | –14.5 ± 0.9 | 23.1 ± 1.3 |
| May–June | 41.1 ± 1.5 b | –15.3 ± 1.2 | 21.7 ± 1.4 |
| June–July | 42.9 ± 0.5 a | –14.6 ± 0.5 | 22.6 ± 2.1 |
| July | 43.4 ± 0.5 a | –15.1 ± 0.9 | 21.8 ± 1.1 |
| n.s. | n.s. | ||
| Shading degree (%) | |||
| Unshaded control | 42.6 ± 1.6 | –14.8 ± 1.1 | 22.0 ± 2.4 |
| 50 | 42.2 ± 2.3 | –14.6 ± 1.0 | 22.6 ± 1.3 |
| 79 | 41.6 ± 1.7 | –15.2 ± 1.0 | 22.7 ± 1.5 |
| n.s. | n.s. | n.s. |
| Treatment | NO3 | N | K | P | S | Ca | Mg | Na |
|---|---|---|---|---|---|---|---|---|
| mg·kg−1 f.w. | g·kg−1 d.w. | g·kg−1 d.w. | g·kg−1 d.w. | g·kg−1 d.w. | g·kg−1 d.w. | g·kg−1 d.w. | g·kg−1 d.w. | |
| Cropping season | ||||||||
| April–May | 6863 ± 488 a | 4.50 ± 0.36 | 47.6 ± 6.3 b | 3.02 ± 0.31 a | 7.37 ± 0.36 | 29.3 ± 1.4 a | 3.56 ± 0.15 a | 3.15 ± 0.27 |
| June–July | 6615 ± 303 a,b | 4.56 ± 0.41 | 50.3 ± 4.8 a,b | 2.84 ± 0.25 a,b | 7.25 ± 0.33 | 27.8 ± 1.5 a,b | 3.37 ± 0.12 a,b | 3.24 ± 0.11 |
| July | 6404 ± 305 b | 4.65 ± 0.43 | 53.4 ± 3.1 a | 2.71 ± 0.23 b | 7.18 ± 0.28 | 25.5 ± 3.0 b | 3.14 ± 0.32 b | 3.30 ± 0.18 |
| n.s. | n.s. | n.s. | ||||||
| Shading degree (%) | ||||||||
| Unshaded control | 6218 ± 182 b | 4.70 ± 0.35 | 55.5 ± 1.6 a | 3.08 ± 0.14 a | 7.34 ± 0.33 | 29.2 ± 1.9 a | 3.50 ± 0.20 a | 3.30 ± 0.19 |
| 50 | 6594 ± 158 b | 4.58 ± 0.44 | 51.6 ± 2.0 b | 2.85 ± 0.11 a,b | 7.28 ± 0.23 | 27.7 ± 1.0 a,b | 3.39 ± 0.10 a,b | 3.22 ± 0.11 |
| 79 | 7070 ± 336 a | 4.44 ± 0.20 | 44.0 ± 4.7 c | 2.65 ± 0.07 b | 7.20 ± 0.36 | 25.5 ± 3.9 b | 3.20 ± 0.42 b | 3.18 ± 0.42 |
| n.s. | n.s. | n.s. |
| Treatment | Cu | Fe | Mn | Se | Zn |
|---|---|---|---|---|---|
| mg·kg−1 d.w. | mg·kg−1 d.w. | mg·kg−1 d.w. | µg·kg−1 d.w. | mg·kg−1 d.w. | |
| Cropping season | |||||
| April–May | 18 ± 3 b | 486 ± 49 b | 53 ± 4 | 237 ± 50 c | 45 ± 2 a |
| June–July | 20 ± 2 a,b | 515 ± 23 a,b | 56 ± 2 | 272 ± 51 b | 42 ± 2 a,b |
| July | 21 ± 3 a | 532 ± 58 a | 58 ± 7 | 326 ± 81 a | 38 ± 4 b |
| n.s. | |||||
| Shading degree (%) | |||||
| Unshaded control | 22 ± 2 a | 544 ± 47 a | 60 ± 5 a | 218 ± 24 c | 44 ± 3 a |
| 50 | 20 ± 1 a | 517 ± 22 a,b | 56 ± 3 a,b | 350 ± 58 a | 42 ± 1 a,b |
| 79 | 17 ± 1 b | 472 ± 44 b | 51 ± 4 b | 267 ± 57 b | 39 ± 5 b |
| Treatment | LAA | HAA | Total Phenols | TAA |
|---|---|---|---|---|
| (mmol Trolox 100 g−1 d.w.) | (mmol AA 100 g−1 d.w.) | (mg gallic acid 100 g−1 d.w.) | (mg 100 g−1 f.w.) | |
| Cropping season (CS) | ||||
| April–May | 14.76 ± 0.72 c | 7.28 ± 0.89 a | 2.45 ± 0.14 | 79.60 ± 12.0 a |
| June–July | 18.42 ± 1.38 b | 6.32 ± 0.75 b | 2.52 ± 0.13 | 65.97 ± 6.72 b |
| July | 20.14 ± 0.75 a | 6.71 ± 0.60 b | 2.30 ± 0.11 | 82.08 ± 16.0 a |
| * | * | n.s. | * | |
| Shading degree (SD, %) | ||||
| Unshaded control | 14.84 ± 0.76 c | 8.83 ± 0.19 a | 2.42 ± 0.15 a,b | 119.7 ± 8.30 a |
| 50 | 17.87 ± 1.19 b | 7.41 ± 0.48 b | 2.23 ± 0.07 b | 57.12 ± 4.55 b |
| 79 | 20.61 ± 0.88 a | 4.08 ± 0.13 c | 2.62 ± 0.12 a | 50.81 ± 3.37 b |
| SD × CS | ||||
| Control × April–May | 13.32 ± 0.33 c | 8.98 ± 0.54 a | 2.85 ± 0.11 a,b | 123.9 ± 2.77 b |
| Control × June–July | 13.71 ± 1.06 c | 9.02 ± 0.21 a | 2.53 ± 0.14 a,b,c | 90.12 ± 4.23 c |
| Control × July | 17.51 ± 0.53 b | 8.47 ± 0.11 a | 1.88 ± 0.02 d | 145.1 ± 5.56 a |
| 50 × April–May | 13.61 ± 0.72 c | 9.06 ± 0.22 a | 2.12 ± 0.14 c,d | 72.13 ± 5.72 d |
| 50 × June–July | 18.66 ± 0.57 b | 6.03 ± 0.41 c | 2.12 ± 0.10 c,d | 47.21 ± 3.97 e,f |
| 50 × July | 21.33 ± 0.96 a | 7.13 ± 0.40 b | 2.44 ± 0.02 b,c | 52.03 ± 4.99 e,f |
| 79 × April–May | 17.36 ± 0.65 b | 3.81 ± 0.02 d | 2.38 ± 0.26 c | 42.77 ± 3.61 f |
| 79 × June–July | 22.90 ± 0.48 a | 3.93 ± 0.16 d | 2.90 ± 0.16 a | 60.60 ± 5.11 d,e |
| 79 × July | 21.57 ± 0.60 a | 4.51 ± 0.22 d | 2.58 ± 0.13 a,b,c | 49.06 ± 3.92 e,f |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, G.; Formisano, L.; Cozzolino, E.; Pannico, A.; El-Nakhel, C.; Rouphael, Y.; Tallarita, A.; Cenvinzo, V.; De Pascale, S. Shading Affects Yield, Elemental Composition and Antioxidants of Perennial Wall Rocket Crops Grown from Spring to Summer in Southern Italy. Plants 2020, 9, 933. https://doi.org/10.3390/plants9080933
Caruso G, Formisano L, Cozzolino E, Pannico A, El-Nakhel C, Rouphael Y, Tallarita A, Cenvinzo V, De Pascale S. Shading Affects Yield, Elemental Composition and Antioxidants of Perennial Wall Rocket Crops Grown from Spring to Summer in Southern Italy. Plants. 2020; 9(8):933. https://doi.org/10.3390/plants9080933
Chicago/Turabian StyleCaruso, Gianluca, Luigi Formisano, Eugenio Cozzolino, Antonio Pannico, Christophe El-Nakhel, Youssef Rouphael, Alessio Tallarita, Vincenzo Cenvinzo, and Stefania De Pascale. 2020. "Shading Affects Yield, Elemental Composition and Antioxidants of Perennial Wall Rocket Crops Grown from Spring to Summer in Southern Italy" Plants 9, no. 8: 933. https://doi.org/10.3390/plants9080933
APA StyleCaruso, G., Formisano, L., Cozzolino, E., Pannico, A., El-Nakhel, C., Rouphael, Y., Tallarita, A., Cenvinzo, V., & De Pascale, S. (2020). Shading Affects Yield, Elemental Composition and Antioxidants of Perennial Wall Rocket Crops Grown from Spring to Summer in Southern Italy. Plants, 9(8), 933. https://doi.org/10.3390/plants9080933