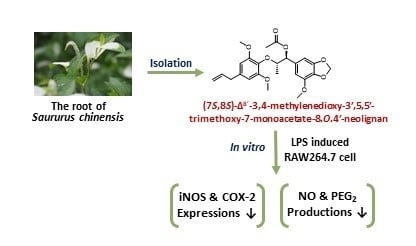
Anti-Inflamatory Activity of Neolignan Compound Isolated from the Roots of Saururus chinensis


Abstract
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
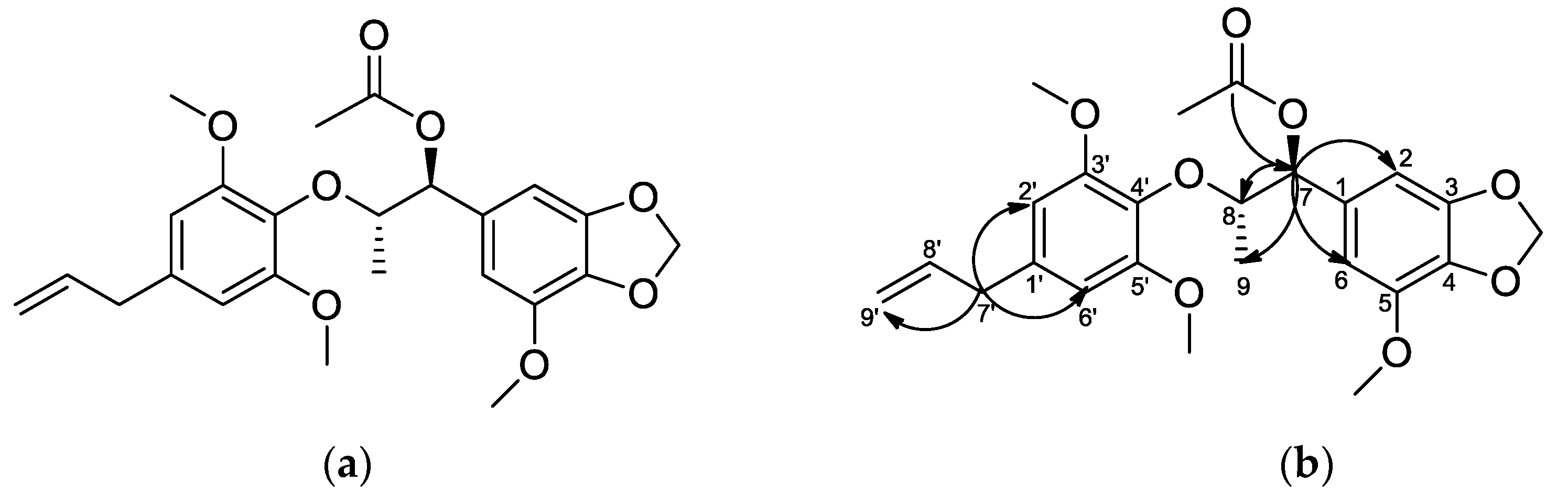
2.1. Structure Determination of MTMN
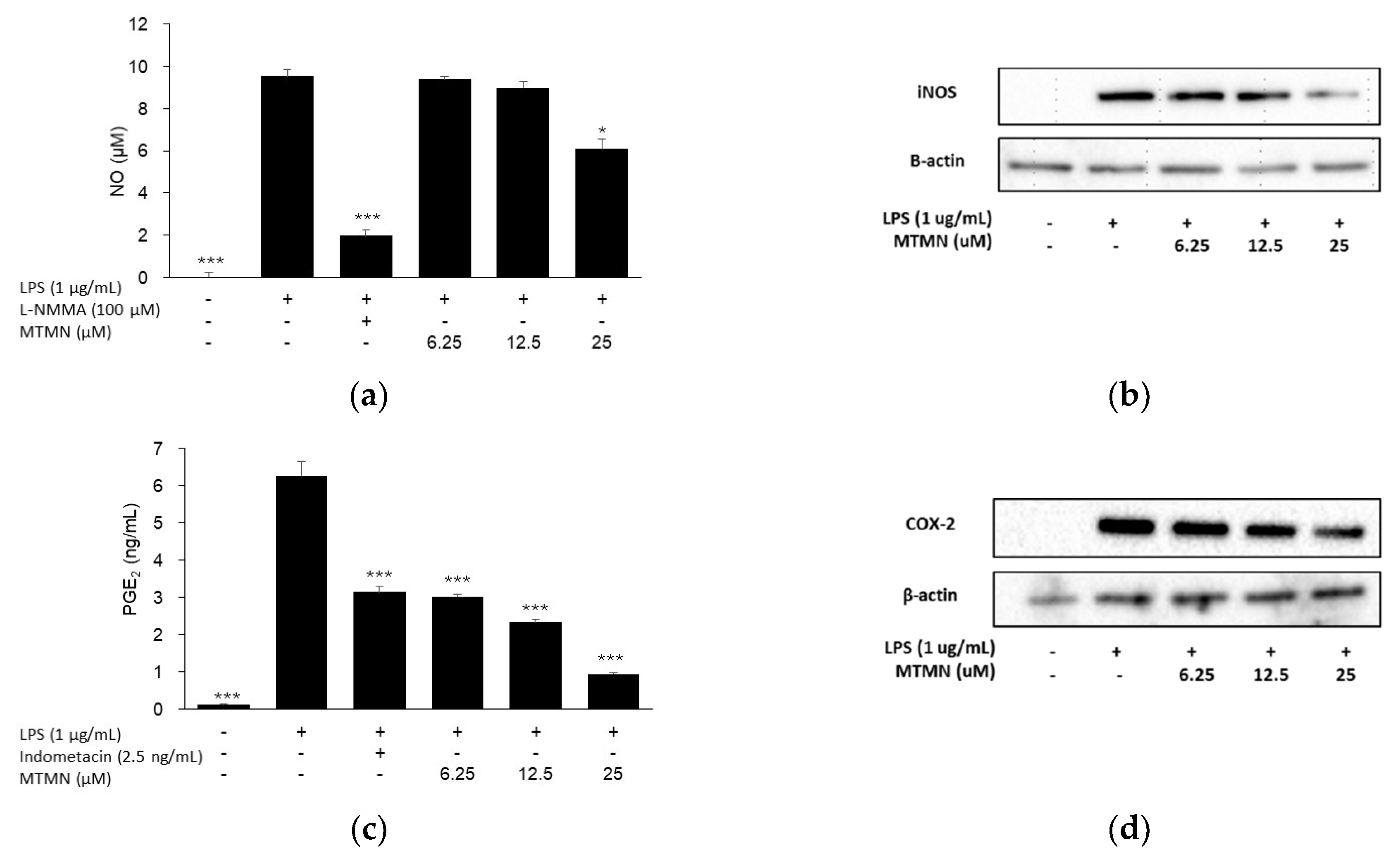
2.2. Inhibitory Effect of Inflammation Related Mediator of MTMN
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Cytotoxicity Assay
3.3. Determination of Nitric Oxide (NO) and Prostaglandin E2 (PGE2)
3.4. Western Blotting
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pirillo, A.; Bonacina, F.; Norata, G.D.; Catapano, A.L. The Interplay of Lipids, Lipoproteins, and Immunity in Atherosclerosis. Curr. Atheroscler. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Herron, J.W.; Nerurkar, L.; Cavanagh, J. Neuroimmune Biomarkers in Mental Illness. Curr. Top. Behav. Neurosci. 2018, 40, 45–78. [Google Scholar] [PubMed]
- Miller, A.H.; Raison, C.L. The role of inflammation in depression: From evolutionary imperative to modern treatment target. Nat. Rev. Immunol. 2016, 16, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Leung, K.S.; Chow, S.K.; Cheung, W.H. Inflammation and age-associated skeletal muscle deterioration (sarcopaenia). J. Orthop. Translat. 2017, 10, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Avalos, Y.; Kerr, B.; Maliqueo, M.; Dorfman, M. Cell and molecular mechanisms behind diet-induced hypothalamic inflammation and obesity. J. Neuroendocrinol. 2018, 30, e12598. [Google Scholar] [CrossRef] [PubMed]
- Bally, M.; Dendukuri, N.; Rich, B.; Nadeau, L.; Helin-Salmivaara, A.; Garbe, E.; Brophy, J.M. Risk of acute myocardial infarction with NSAIDs in real world use: Bayesian meta-analysis of individual patient data. BMJ Br. Med. J. 2017, 357. [Google Scholar] [CrossRef] [PubMed]
- Arboretum, K.N. English Names for Korean Native Plants; Korea National Arboretum: Pocheon, Korea, 2015; p. 778. [Google Scholar]
- Park, J.H.; Lee, J.G. The Encycolpedia of Medicinal Plants; Shin Il Publishing Co.: Seoul, Korea, 2000; pp. 202–203. [Google Scholar]
- Li, C.; Li, N.; Yue, J.; Song, Q.; Fan, Q. Two new lignans from Saururus chinensis. Nat. Prod. Res. 2017, 31, 1598–1603. [Google Scholar] [CrossRef] [PubMed]
- Seo, C.S.; Lee, Y.K.; Kim, Y.J.; Jung, J.S.; Jahng, Y.; Chang, H.W.; Song, D.K.; Son, J.K. Protective effect of lignans against sepsis from the roots of Saururus chinensis. Biol. Pharm. Bull. 2008, 31, 523–526. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Seo, C.S.; Zheng, M.S.; Woo, M.H.; Lee, C.S.; Lee, S.H.; Jeong, B.S.; Chang, H.W.; Jahng, Y.; Lee, E.S.; Son, J.K. Lignans from the roots of Saururus chinensis. J. Nat. Prod. 2008, 71, 1771–1774. [Google Scholar] [CrossRef] [PubMed]
- Sung, S.H. A new dineolignan from Saururus chinensis root. Fitoterapia 2006, 77, 487–488. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lee, D.S.; Choi, H.G.; Kim, K.S.; Kang, D.G.; Lee, H.S.; Jeong, G.S.; Kim, Y.C. Sauchinone Suppresses Pro-inflammatory Mediators by Inducing Hesla me Oxygenase-1 in RAW264.7 Macrophages. Biol. Pharm. Bull. 2011, 34, 1566–1571. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Kim, H.G.; Sim, Y.; Sung, S.H.; Oh, M.S. Sauchinone, a Lignan from Saururus chinensis, Protects Human Skin Keratinocytes against Ultraviolet B-Induced Photoaging by Regulating the Oxidative Defense System. Biol. Pharm. Bull. 2013, 36, 1134–1139. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Silva, M.S.; Barbosa-Filho, J.M.; Yoshida, M.; Gottlieb, O.R. Benzodioxane and β–aryloxy-arylpropane type neolignans from Licaria chrysophylla. Phytochemistry 1989, 28, 3477–3482. [Google Scholar] [CrossRef]
- Fang, J.M.; Lee, C.K.; Cheng, Y.S. Lignans from leaves of Juniperus chinensis. Phytochemistry 1992, 31, 3659–3661. [Google Scholar]
- Lee, I.S.; Kim, Y.S.; Jung, S.H.; Yu, S.Y.; Kim, J.H.; Sun, H.; Kim, J.S. Lignans from the stems and leaves of Brandisia hancei and their effects on VEGF-induced vascular permeability and migration of HRECs and DLAV formation in zebrafish. Biosci. Biotechnol. Biochem. 2015, 79, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Barata, L.E.S.; Baker, P.M.; Gottlieb, O.R.; Rùveda, E.A. Neolignans of Virola surinamensis. Phytochemistry 1978, 17, 783–786. [Google Scholar] [CrossRef]
- Conserva, L.M.; da Silva, M.S.; Filho, R.B. Lignans from Aristolochia birostris. Phytochemistry 1990, 29, 257–260. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar] [PubMed]
- Williams, C.S.; Mann, M.; DuBois, R.N. The role of cyclooxygenases in inflammation, cancer, and development. Oncogene 1999, 18, 7908–7916. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, S.-R.; Ha, H.; Shin, H.-K.; Seo, C.-S. Anti-Inflamatory Activity of Neolignan Compound Isolated from the Roots of Saururus chinensis. Plants 2020, 9, 932. https://doi.org/10.3390/plants9080932
Yoo S-R, Ha H, Shin H-K, Seo C-S. Anti-Inflamatory Activity of Neolignan Compound Isolated from the Roots of Saururus chinensis. Plants. 2020; 9(8):932. https://doi.org/10.3390/plants9080932
Chicago/Turabian StyleYoo, Sae-Rom, Hyekyung Ha, Hyeun-Kyoo Shin, and Chang-Seob Seo. 2020. "Anti-Inflamatory Activity of Neolignan Compound Isolated from the Roots of Saururus chinensis" Plants 9, no. 8: 932. https://doi.org/10.3390/plants9080932
APA StyleYoo, S.-R., Ha, H., Shin, H.-K., & Seo, C.-S. (2020). Anti-Inflamatory Activity of Neolignan Compound Isolated from the Roots of Saururus chinensis. Plants, 9(8), 932. https://doi.org/10.3390/plants9080932