Photochemical Efficiency and Oxidative Metabolism of Tree Species during Acclimation to High and Low Irradiance



Abstract
1. Introduction
2. Material and Methods
2.1. Plant Material and Growth Conditions
2.2. Chloroplastidic Pigments Content
2.3. Chlorophyll Fluorescence Parameters
2.4. Extraction of Soluble Protein and Measurement of Enzyme Activities
2.5. Lipid Peroxidation (Determination of Malonaldehyde Content)
2.6. Leaf Phenols Compounds
2.7. Leaf-Nutrient Content
2.8. Plasticity Index
2.9. Experimental Design and Statistical Analysis
3. Results
3.1. Chloroplastidic Pigments Content
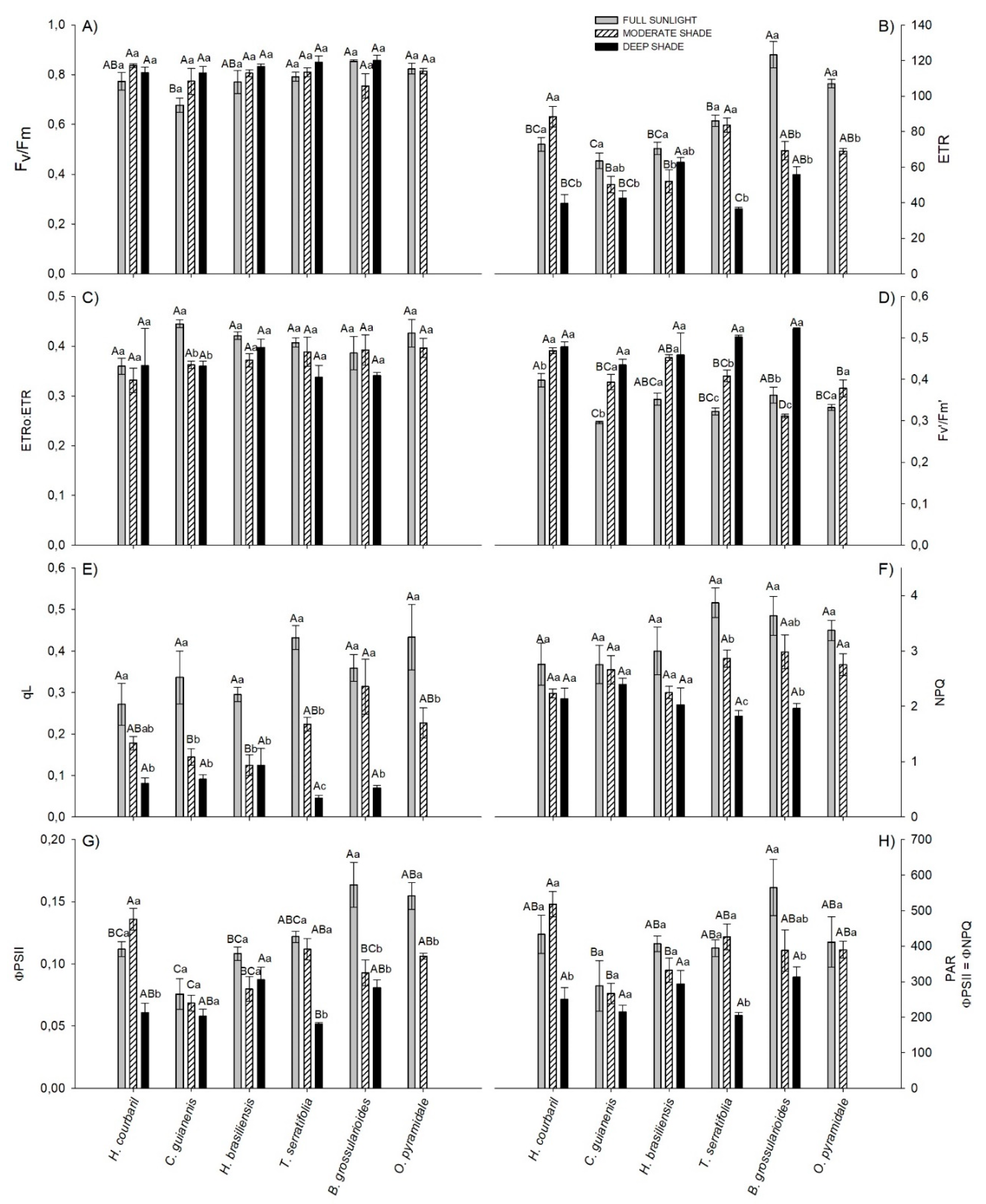
3.2. Chlorophyll Fluorescence Parameters
3.3. Leaf-Nutrient Content
3.4. Enzymatic Activities, Leaf Phenolic Compounds, and Lipid Peroxidation
3.5. Plasticity Index
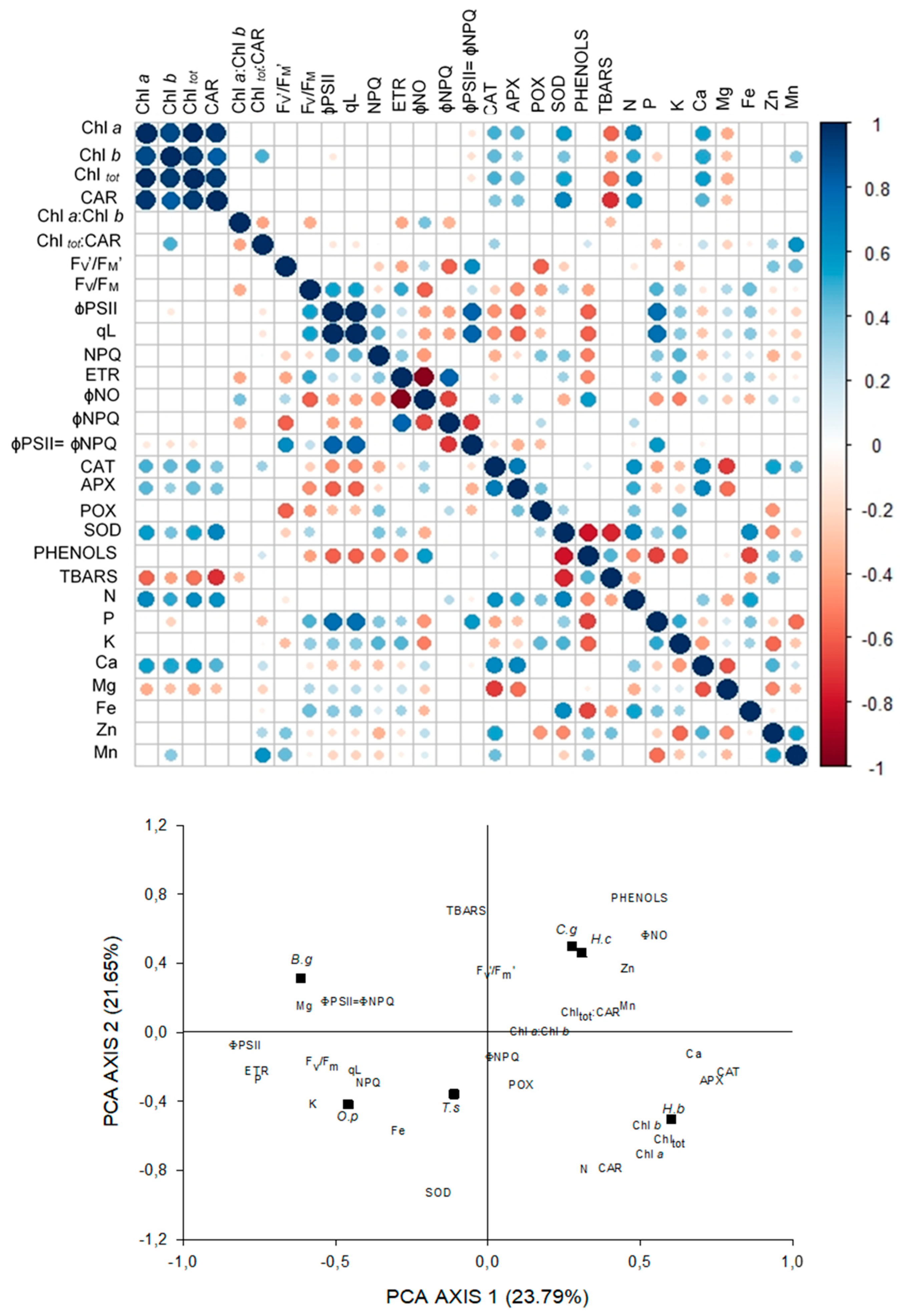
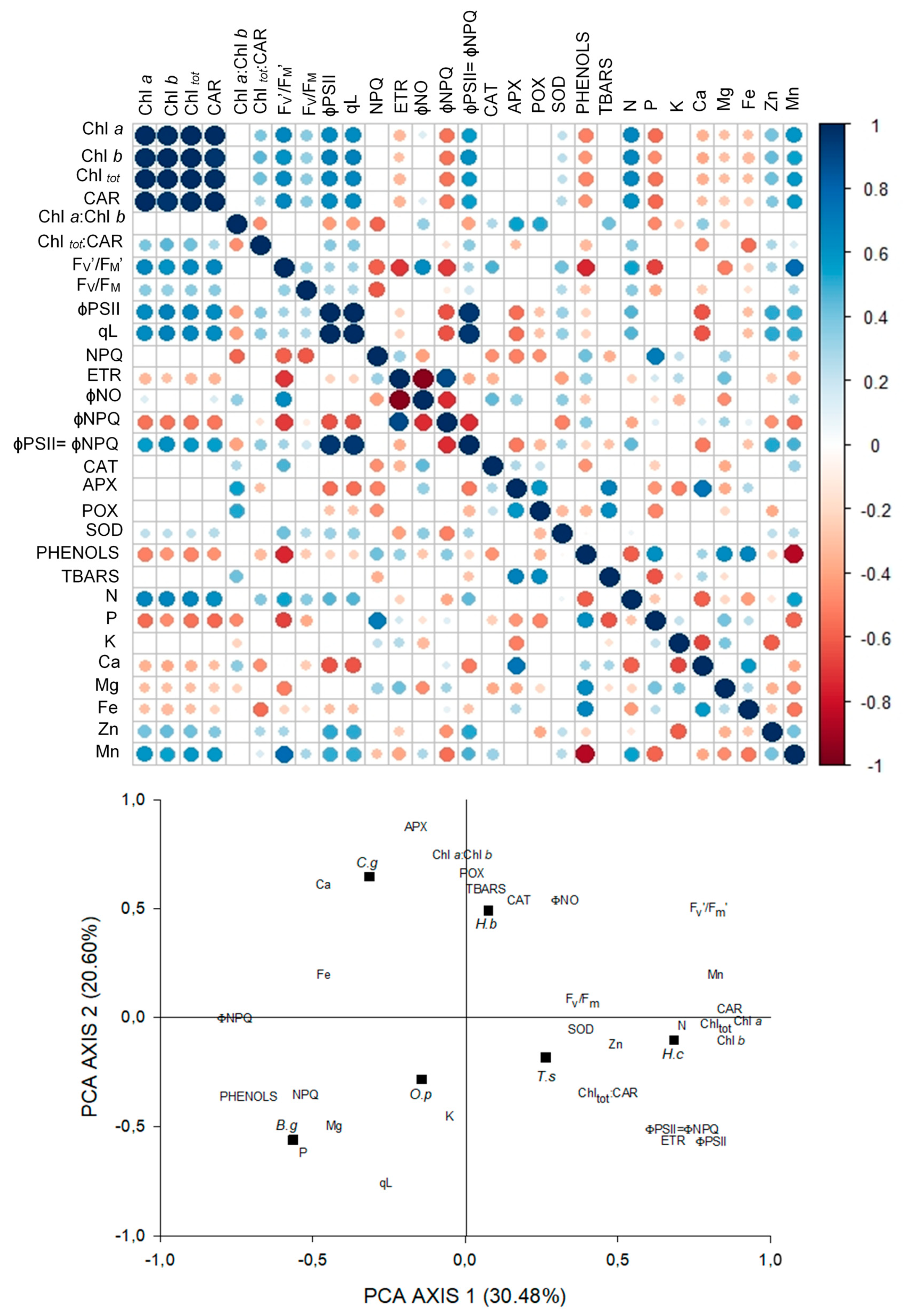
3.6. Relationships between Light Capture and Use Parameters, Antioxidant Activity, and Leaf-Nutrient Content
4. Discussion
4.1. Chloroplastidic Pigment Content
4.2. Chlorophyll Fluorescence Parameters
4.3. Leaf-Nutrient Content
4.4. Enzymatic Activities, Leaf Phenolic Compounds, and Lipid Peroxidation
4.5. Plasticity Index
4.6. Relationships between Light Capture and USE Parameters, Antioxidant Activity, and Leaf-Nutrient Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baird, A.S.; Anderegg, L.D.L.; Lacey, M.E.; Hillerislambers, J.; Van Volkenburgh, E. Comparative leaf growth strategies in response to low-water and low-light availability: Variation in leaf physiology underlies variation in leaf mass per area in Populus tremuloides. Tree Physiol. 2017, 37, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Costa, K.C.; Jaquetti, R.; de Carvalho Gonçalves, J.F. Special issue in honour of Prof. Reto J. StrasserChlorophyll a fluorescence of Bertholletia excelsa Bonpl. plantations under thinning, liming, and phosphorus fertilization. Photosynthetica 2020, 58. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, S.-B.; Liu, T.; Huang, W. Decreased photosystem II activity facilitates acclimation to fluctuating light in the understory plant Paris polyphylla. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148135. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Laanisto, L.; Niinemets, Ü.; Zavala, M.A. Shedding light on shade: Ecological perspectives of understorey plant life. Plant Ecol. Divers. 2016, 9, 237–251. [Google Scholar] [CrossRef]
- Favaretto, V.F.; Martinez, C.A.; Soriani, H.H.; Furriel, R.P.M. Differential responses of antioxidant enzymes in pioneer and late-successional tropical tree species grown under sun and shade conditions. Environ. Exp. Bot. 2011, 70, 20–28. [Google Scholar] [CrossRef]
- Dos Anjos, L.; Oliva, M.A.; Kuki, K.N.; Mielke, M.S.; Ventrella, M.C.; Galvão, M.F.; Pinto, L.R.M. Key leaf traits indicative of photosynthetic plasticity in tropical tree species. Trees Struct. Funct. 2015, 29, 247–258. [Google Scholar] [CrossRef]
- Walters, R.G. Towards an understanding of photosynthetic acclimation. J. Exp. Bot. 2005, 56, 435–447. [Google Scholar] [CrossRef]
- Dietz, K.J. Efficient high light acclimation involves rapid processes at multiple mechanistic levels. J. Exp. Bot. 2015, 66, 2401–2414. [Google Scholar] [CrossRef]
- Lestari, D.P.; Nichols, J.D. Seedlings of subtropical rainforest species from similar successional guild show different photosynthetic and morphological responses to varying light levels. Tree Physiol. 2017, 37, 186–198. [Google Scholar] [CrossRef][Green Version]
- Sukhova, E.; Khlopkov, A.; Vodeneev, V.; Sukhov, V. Simulation of a nonphotochemical quenching in plant leaf under different light intensities. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148138. [Google Scholar] [CrossRef]
- Tikkanen, M.; Mekala, N.R.; Aro, E. Biochimica et Biophysica Acta Photosystem II photoinhibition-repair cycle protects Photosystem I from irreversible damage. BBA Bioenerg. 2014, 1837, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Aro, E.; Millar, A.H. Mechanisms of Photodamage and Protein Turnover in Photoinhibition. Trends Plant Sci. 2018, 23, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Derks, A.; Schaven, K.; Bruce, D. Diverse mechanisms for photoprotection in photosynthesis. Dynamic regulation of photosystem II excitation in response to rapid environmental change. Biochim. Biophys. Acta Bioenerg. 2015, 1847, 468–485. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.R.; de A. Silva, M.M.; Willadino, L.; Ulisses, C.; Camara, T.R. Geração e desintoxicação enzimática de espécies reativas de oxigênio em plantas. Ciência Rural 2014, 44, 453–460. [Google Scholar] [CrossRef]
- Wu, H.; Jiang, H.; Liu, C.; Deng, Y. South African Journal of Botany Growth, pigment composition, chlorophyll fl uorescence and antioxidant defenses in the red alga Gracilaria lemaneiformis (Gracilariales, Rhodophyta) under light stress. S. Afr. J. Bot. 2015, 100, 27–32. [Google Scholar] [CrossRef]
- Filstrup, C.; Downing, J. Relationship of chlorophyll to phosphorus and nitrogen in nutrient-rich lakes. Inl. Waters 2017, 7. [Google Scholar] [CrossRef]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- De Oliveira dos Santos, O.; Mendes, K.R.; Martins, S.V.C.; Batista-Silva, W.; Dos Santos, M.A.; de Figueirôa, J.M.; de Souza, E.R.; Fernandes, D.; Araújo, W.L.; Pompelli, M.F. Physiological parameters and plasticity as key factors to understand pioneer and late successional species in the Atlantic Rainforest. Acta Physiol. Plant. 2019, 41, 1–18. [Google Scholar] [CrossRef]
- De Souza, F.M.; Batista, J.L.F. Restoration of seasonal semideciduous forests in Brazil: Influence of age and restoration design on forest structure. For. Ecol. Manag. 2004, 191, 185–200. [Google Scholar] [CrossRef]
- Vinson, C.C.; Azevedo, V.C.R.; Sampaio, I.; Ciampi, A.Y. Development of microsatellite markers for Carapa guianensis (Aublet), a tree species from the Amazon forest. Mol. Ecol. Notes 2005, 5, 33–34. [Google Scholar] [CrossRef]
- Amaral, D.D.; Viera, I.C.G.; de Almeida, S.S.; Salomão, R.P.; da Silva, A.S.L.; Jardim, M.A.G. Checklist da flora arbórea de remanescentes florestais da região metropolitana de Belém e valor histórico dos fragmentos, Pará, Brasil. Bol. do Mus. Para. Emilio Goeldi, Ciências Nat. 2009, 4, 231–289. [Google Scholar]
- Gualberto, M.L.C.; da Silva Ribeiro, R.B.; Gama, J.R.V.; dos Santos Vieira, D. Fitossociologia E Potencial De Espécies Arbóreas Em Ecossistema Sucessional Na Floresta Nacional Do Tapajós, Pará. Agrossistemas 2014, 6, 42–57. [Google Scholar] [CrossRef][Green Version]
- Bentos, T.V.; Nascimento, H.E.M.; Dos Anjos Vizcarra, M.; Williamson, G.B. Effects of lightgaps and topography on Amazon secondary forest: Changes in species richness and community composition. For. Ecol. Manag. 2017, 396, 124–131. [Google Scholar] [CrossRef]
- Slot, M.; Winter, K. High tolerance of tropical sapling growth and gas exchange to moderate warming. Funct. Ecol. 2018, 32, 599–611. [Google Scholar] [CrossRef]
- Lichtenthaler, H.; Wellburn, A. Determinations of total carotenoids and chlorophylls b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Hendry, G.A.F.; Price, A.H. Stress indicators: Chlorophylls and carotenoids. Methods Comp. Plant Ecol. 1993, 148–152. [Google Scholar]
- Krause, G.H.; Weiss, E. Chlorophyll fluorescence and photosynthesis: The basis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.N.; Olavi, K.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxe. Microsc. Microanal. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Genty, B.; Brientais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron-transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Epron, D.; Godard, D.; Cornic, G.; Genty, B.; Sud, U.F. Limitation of net CO2 assimilation rate by internal resistances to CO2 transfer in the leaves of two tree species (Fagus sylvatica L and Castanea sativa Mill.). Plant Cell Environ. 1995, 18, 43–51. [Google Scholar] [CrossRef]
- Yin, X.; Struik, P.C.; Romero, P.; Harbinson, J.; Evers, J.B.; Van Der Putten, P.E.L.; Vos, J. Using combined measurements of gas exchange and chlorophyll fluorescence to estimate parameters of a biochemical C3photosynthesis model: A critical appraisal and a new integrated approach applied to leaves in a wheat (Triticum aestivum) canopy. Plant Cell Environ. 2009, 32, 448–464. [Google Scholar] [CrossRef] [PubMed]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Walters, R.; Horton, G.P. Resolution of components of non- photochemical chlorophyll fluorescence quenching in barley leaves. Photosynth. Res. 1991, 27, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Calcerrada, J.; Reich, P.B.; Rosenqvist, E.; Pardos, J.A.; Cano, F.J.; Aranda, I. Leaf physiological versus morphological acclimation to high-light exposure at different stages of foliar development in oak. Tree Physiol. 2008, 28, 761–771. [Google Scholar] [CrossRef]
- Azevedo, R.A.; Alas, R.M.; Smith, R.J.; Lea, P.J. Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiol. Plant. 1998, 104, 280–292. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 880. [Google Scholar]
- Kar, M.; Mishraa, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef]
- Beauchamp, C.O.; Fridovich, I. Superoxide dismutase. Improved assays and an assay applicable to acrylamide gel. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases in higher-plants. Plant Physiol. 1977, 309–314. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase and perroxidase activies in root tips of soyabean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochmestry Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Jennings, A.C. The determination of dihydroxy phenolic compounds in extracts of plant tissues. Anal. Biochem. 1981, 118, 396–398. [Google Scholar] [CrossRef]
- Miyazawa, M.; Pavan, M.A.; Muraoka, T.; Carmo, C.A.F.S.; Mello, W.J. Análise química de tecidos vegetais. In Manual de Análise Química de Solos, Plantas e Fertilizantes; Silva, F.S., Ed.; EMBRAPA: Brasilia, Brazil, 1999; pp. 172–223. [Google Scholar]
- Vitti, G.C.; Ferreira, A.C. Síntese de Análises Químicas em Tecido Vegetal; Escola Superior de Agricultura Luiz de Queiroz: Piracicaba, Brazil, 1997. [Google Scholar]
- Valladares, F.; Niinemets, Ü. Shade Tolerance, a Key Plant Feature of Complex Nature and Consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Dos Santos Magalhães, N.; Marenco, R.A.; Mendes, K.R. Aclimatação de mudas de acariquara à alta irradiância. Pesqui. Agropecu. Bras. 2009, 44, 687–694. [Google Scholar] [CrossRef]
- Gonçalves, J.F.D.C.; da Silva, C.E.M.; Justino, G.C.; Nina Junior, A.D.R. Efeito do ambiente de luz no crescimento de plantas jovens de mogno (Swietenia macrophylla King). Sci. For. Sci. 2012, 40, 337–344. [Google Scholar]
- Quevedo-Rojas, A.; García-Núñez, C.; Jerez-Rico, M.; Jaimez, R.; Schwarzkopf, T. Leaf acclimation strategies to contrasting light conditions in saplings of different shade tolerance in a tropical cloud forest. Funct. Plant Biol. 2018, 45, 968–982. [Google Scholar] [CrossRef]
- Krause, G.H.; Koroleva, O.Y.; Dalling, J.W.; Winter, K. Acclimation of tropical tree seedlings to excessive light in simulated tree-fall gaps. Plant Cell Environ. 2001, 24, 1345–1352. [Google Scholar] [CrossRef]
- Gonçalves, J.F.D.C.; Marenco, R.A.; Vieira, G. Concentration of photosynthetic pigments and chlorophyll fluorescence of mahogany and tonka bean under two light environments. Rev. Bras. Fisiol. Veg. 2001, 13, 149–157. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Bussotti, F.; Gerosa, G.; Digrado, A.; Pollastrini, M. Selection of chlorophyll fluorescence parameters as indicators of photosynthetic efficiency in large scale plant ecological studies. Ecol. Indic. 2020, 108, 105686. [Google Scholar] [CrossRef]
- Morais, R.R.; Gonçalves, J.F.D.C.; Dos Santos, U.M.; Dünisch, O.; Dos Santos, A.L.W. Chloroplastid pigment contents and chlorophyll a fluorescence in Amazonian tropical three species. Rev. Arvore 2007, 31, 959–966. [Google Scholar] [CrossRef]
- de Carvalho Gonçalves, J.F.; Silva, C.E.; Guimarães, D.G.; Bernardes, R.S. Análise dos transientes da fluorescência da clorofila a de plantas jovens de Carapa guianensis e de Dipteryx odorata submetidas a dois ambientes de luz. Acta Amaz. 2010, 40, 89–98. [Google Scholar] [CrossRef]
- Azevedo, G.F.C.; Marenco, R.A. Growth and physiological changes in saplings of Minquartia guianensis and Swietenia macrophylla during acclimation to full sunlight. Photosynthetica 2012, 50, 86–94. [Google Scholar] [CrossRef]
- Voss, I.; Sunil, B.; Scheibe, R.; Raghavendra, A.S. Emerging concept for the role of photorespiration as an important part of abiotic stress response. Plant Biol. 2013, 15, 713–722. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef]
- Guidi, L.; Calatayud, A. Non-invasive tools to estimate stress-induced changes in photosynthetic performance in plants inhabiting Mediterranean areas. Environ. Exp. Bot. 2014, 103, 42–52. [Google Scholar] [CrossRef]
- Schimpl, F.C.; Ribeiro, R.V.; Pereira, L.; Rodrigues, H.S.; Mazzafera, P. Photochemical responses to abrupt and gradual chilling treatments in eucalyptus species. Theor. Exp. Plant Physiol. 2018, 30, 9–17. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102–112. [Google Scholar] [CrossRef]
- Portsmuth, A.; Niinemets, Ü. Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Funct. Ecol. 2007, 21, 61–77. [Google Scholar] [CrossRef]
- Cunha, H.F.V.; de Carvalho Gonçalves, J.F.; Dos Santos, U.M.; Ferreira, M.J.; Peixoto, P.H.P. Biomassa, trocas gasosas e aspectos nutricionais de plantas jovens de pau de balsa (Ochroma pyramidale (Cav. Ex Lamb.) Urb.) submetidas à fertilização fosfatada em ambientes contrastantes de irradiância. Sci. For. Sci. 2016, 44, 215–230. [Google Scholar] [CrossRef]
- Dos Santos Junior, U.M.; de Carvalho Gonçalves, J.F.; Feldpausch, T.R. Growth, leaf nutrient concentration and photosynthetic nutrient use efficiency in tropical tree species planted in degraded areas in central Amazonia. For. Ecol. Manag. 2006, 226, 299–309. [Google Scholar] [CrossRef]
- Mendes, K.R.; Marenco, R.A. Photosynthetic traits of tree species in response to leaf nutrient content in the central Amazon. Theor. Exp. Plant Physiol. 2015, 27, 51–59. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Khorobrykh, S.; Havurinne, V.; Mattila, H.; Tyystjärvi, E. Oxygen and ROS in Photosynthesis. Plants 2020, 9, 91. [Google Scholar] [CrossRef]
- Noctor, G.; Lelarge-trouverie, C.; Mhamdi, A. Phytochemistry The metabolomics of oxidative stress. Phytochemistry 2015, 112, 33–53. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Inès, J.; Al-Juburi, H.J.; Chang-Xing, Z.; Hong-Bo, S.; Panneerselvam, R. Antioxidant defense responses: Physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant. 2009, 31, 427–436. [Google Scholar] [CrossRef]
- Yanik, T.; Donaldson, R.P. A protective association between catalase and isocitrate lyase in peroxisomes. Arch. Biochem. Biophys. 2005, 435, 243–252. [Google Scholar] [CrossRef]
- Sousa, R.H.V.; Carvalho, F.E.L.; Lima-Melo, Y.; Alencar, V.T.C.B.; Daloso, D.M.; Margis-Pinheiro, M.; Komatsu, S.; Silveira, J.A.G. Impairment of peroxisomal APX and CAT activities increases protection of photosynthesis under oxidative stress. J. Exp. Bot. 2019, 70, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Bela, K.; Riyazuddin, R.; Horváth, E.; Hurton, Á.; Gallé, Á. Comprehensive analysis of antioxidant mechanisms in Arabidopsis glutathione peroxidase-like mutants under salt- and osmotic stress reveals organ-speci fi c signi fi cance of the AtGPXL’s activities. Environ. Exp. Bot. 2018, 150, 127–140. [Google Scholar] [CrossRef]
- Hansen, U.; Fiedler, B.; Rank, B. Variation of pigment composition and antioxidative systems along the canopy light gradient in a mixed beech/oak forest: A comparative study on deciduous tree species differing in shade tolerance. Trees 2002, 16, 354–364. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. Oxidative stress and antioxidative systems: Recipes for successful data collection and interpretation. Plant Cell Environ. 2016, 39, 1140–1160. [Google Scholar] [CrossRef]
- Lima, C.S.; Ferreira-Silva, S.L.; Carvalho, F.E.L.; Lima Neto, M.C.; Aragão, R.M.; Silva, E.N.; Sousa, R.M.J.; Silveira, J.A.G. Antioxidant protection and PSII regulation mitigate photo-oxidative stress induced by drought followed by high light in cashew plants. Environ. Exp. Bot. 2018, 149, 59–69. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Picket, S.T.A. Physiological ecology of tropical succession: A comparative review. Annu. Rev. Ecol. Syst. 1980, 11, 297–310. [Google Scholar] [CrossRef]
- Chadzon, R.; Pearcy, R.W.; Lee, D.W.; Fetcher, N. Photosynthetic responses of tropical forest plants to contrasting light environments. In Tropical Forest Plant Ecophysiology; Mulkey, S.S., Chazdon, R.L., Smith, A.P., Eds.; Chapman and Hall: New York, NY, USA, 1996; pp. 5–55. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specie | Family | Successional Groups |
|---|---|---|
| Hymenea courbaril | Fabaceae | Late successional [19] |
| Carapa guianensis | Meliaceae | Late successional [20] |
| Hevea brasiliensis | Euphorbiaceae | Mid-successional [21] |
| Tabebuia serratifolia | Bignoniaceae | Mid-successional [22] |
| Bellucia grossularioides | Melastomataceae | Pioneer [23] |
| Ochroma pyramidale | Malvaceae | Pioneer [24] |
| Full Sunlight (FS) | Moderate Shade (MS) | Deep Shade (DS) | |
|---|---|---|---|
| PPFD average (µmol photons m−2 s−1) | 1027.51 ± 10.49 | 362.50 ± 9.37 | 47.95 ± 7.98 |
| Maximum PPFD (µmol photons m−2 s−1) | 1866.17 (13:00 h) | 795.61 (12:00 h) | 74.50 (14:00 h) |
| % PPFD | 100 | 35.23 | 4.66 |
| Fluorescence Parameter | Name and Physiological Interpretation |
|---|---|
| Fv/Fm = (Fm − F0)/Fm | Estimated maximum quantum efficiency (yield) of PSII photochemistry [27] |
| Fv′/Fm’ = (Fm′ − F0′)/Fm′ | Maximum efficiency of PSII photochemistry in the light, if all centers were open [28] |
| ɸPSII = (Fm′ − F′)/Fm′ | Estimated effective quantum yield (efficiency) of PSII photochemistry at incident PAR [29] |
| ETR = αβ x PAR x ɸPSII | Electron transport rate in PSII at incident PAR. (αβ was determined from curves obtained under nonphotorespiratory conditions in an atmosphere containing less than 1% O2) [30,31] |
| NPQ = (Fm − Fm′/Fm′) | Non-photochemical quenching [32,33] |
| qP = (Fm′ − Fs)/(Fm′ − F0′) | Coefficient of photochemical quenching based on the “puddle” model (i.e., unconnected PSII units) [34] |
| qL = qP x (F0′/Fs) | Coefficient of photochemical quenching based on the “lake” model (i.e., fully connected PSII units) [28] |
| ɸNO=1/[NPQ+1+qL(Fm/F0 − 1)] | Quantum yield of nonregulated energy dissipation in PSII [28] |
| ɸNPQ = 1 − ɸPSII − ɸNO | Quantum yield for dissipation by down-regulation [28] |
| H. courbaril | C. guianensis | H. brasiliensis | T. serratifolia | B. grossularioides | O. pyramidale | Mean Values | |
|---|---|---|---|---|---|---|---|
| Fluorescence Parameters | |||||||
| FV/FM | 0.22 | 0.28 | 0.23 | 0.17 | 0.27 | 0.11 | 0.21 ± 0.07 D |
| ETR | 0.74 | 0.50 | 0.48 | 0.63 | 0.68 | 0.42 | 0.57 ± 0.13 B |
| ETRO/ETR | 0.64 | 0.27 | 0.25 | 0.39 | 0.32 | 0.25 | 0.35 ± 0.15 CD |
| FV′/FM′ | 0.30 | 0.39 | 0.43 | 0.41 | 0.43 | 0.26 | 0.37 ± 0.07 CD |
| qL | 0.86 | 0.84 | 0.87 | 0.95 | 0.89 | 0.76 | 0.86 ± 0.06 A |
| NPQ | 0.54 | 0.44 | 0.65 | 0.64 | 0.60 | 0.33 | 0.53 ± 0.12 BC |
| ΦPSII | 0.74 | 0.54 | 0.48 | 0.63 | 0.68 | 0.42 | 0.58 ± 0.12 B |
| Mean Values | 0.50 ± 0.08 (2) | ||||||
| Nutritional Parameters | |||||||
| N | 0.60 | 0.49 | 0.61 | 0.61 | 0.63 | 0.41 | 0.56 ± 0.09 A |
| P | 0.55 | 0.46 | 0.68 | 0.50 | 0.82 | 0.62 | 0.61 ± 0.13 A |
| K | 0.83 | 0.75 | 0.84 | 0.72 | 0.73 | 0.73 | 0.77 ± 0.06 A |
| Ca | 0.65 | 0.54 | 0.74 | 0.84 | 0.54 | 0.35 | 0.61 ± 0.17 A |
| Mg | 0.78 | 0.77 | 0.83 | 0.69 | 0.43 | 0.53 | 0.67 ± 0.16 A |
| Fe | 0.59 | 0.76 | 0.38 | 0.79 | 0.77 | 0.35 | 0.61 ± 0.20 A |
| Mn | 0.84 | 0.78 | 0.49 | 0.74 | 0.74 | 0.75 | 0.72 ± 0.12 A |
| Zn | 0.65 | 0.49 | 0.88 | 0.61 | 0.53 | 0.46 | 0.60 ± 0.15 A |
| N:P | 0.48 | 0.61 | 0.76 | 0.66 | 0.91 | 0.59 | 0.67 ± 0.15 A |
| Mean Values | 0.65 ± 0.06 (1,2) | ||||||
| Antioxidant Parameters | |||||||
| CAT | 0.83 | 0.75 | 0.90 | 0.71 | 0.88 | 0.20 | 0.71 ± 0.26 A |
| APX | 0.99 | 0.90 | 0.79 | 0.33 | 0.93 | 0.57 | 0.75 ± 0.25 A |
| POX | 0.99 | 0.75 | 0.83 | 0.49 | 0.70 | 0.72 | 0.75 ± 0.17 A |
| SOD | 0.87 | 0.75 | 0.95 | 0.64 | 0.71 | 0.43 | 0.73 ± 0.18 A |
| PHENOLS | 0.62 | 0.57 | 0.71 | 0.63 | 0.57 | 0.67 | 0.63 ± 0.06 A |
| Mean Values | 0.71 ± 0.14 (1) | ||||||
| Mean Values | 0.68± 0.20 a | 0.60±0.18 ab | 0.66 ± 0.22 a | 0.61 ± 0.18 ab | 0.66 ± 0.19 a | 0.47 ± 0.19 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Rocha Nina Junior, A.; Furtunato Maia, J.M.; Vitor Martins, S.C.; Gonçalves, J.F.d.C. Photochemical Efficiency and Oxidative Metabolism of Tree Species during Acclimation to High and Low Irradiance. Plants 2020, 9, 1047. https://doi.org/10.3390/plants9081047
da Rocha Nina Junior A, Furtunato Maia JM, Vitor Martins SC, Gonçalves JFdC. Photochemical Efficiency and Oxidative Metabolism of Tree Species during Acclimation to High and Low Irradiance. Plants. 2020; 9(8):1047. https://doi.org/10.3390/plants9081047
Chicago/Turabian Styleda Rocha Nina Junior, Adamir, Jair Max Furtunato Maia, Samuel Cordeiro Vitor Martins, and José Francisco de Carvalho Gonçalves. 2020. "Photochemical Efficiency and Oxidative Metabolism of Tree Species during Acclimation to High and Low Irradiance" Plants 9, no. 8: 1047. https://doi.org/10.3390/plants9081047
APA Styleda Rocha Nina Junior, A., Furtunato Maia, J. M., Vitor Martins, S. C., & Gonçalves, J. F. d. C. (2020). Photochemical Efficiency and Oxidative Metabolism of Tree Species during Acclimation to High and Low Irradiance. Plants, 9(8), 1047. https://doi.org/10.3390/plants9081047