Nutritional Value of Savory Herb (Satureja hortensis L.) and Plant Response to Variable Mineral Nutrition Conditions in Various Phases of Development

 , , and
, , and
Abstract
1. Introduction
2. Results and Discussion
2.1. Nitrogen in Savory Raw Material
2.2. Other Macroelements Present in the Herb
2.3. Nutrients Available in Soil
3. Material and Methods
3.1. Plant Material and Experimental Conditions
3.2. Plant Analysis
3.3. Soil Analysis
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grusak, M.A. Enhancing mineral content in plant food products. J. Am. Coll. Nutr. 2002, 21, 178S–183S. [Google Scholar] [CrossRef]
- Özcan, M.M. Mineral contents of some plants used as condiments in Turkey. Food Chem. 2004, 84, 437–440. [Google Scholar] [CrossRef]
- Jafari, F.; Ghavidel, F.; Zarshenas, M.M. A Critical Overview on the Pharmacological and Clinical Aspects of Popular Satureja Species. J. Acupunct. Meridian Stud. 2016, 9, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Kudełka, W.; Kosowska, A. Components of spices and herbs determining their functional properties and their role in human nutrition and prevention of diseases. Zesz. Nauk. Univ. Ekon. Cracow. 2008, 781, 83–111. [Google Scholar]
- Seidler-Łożykowska, K.; Kozik, E.; Golcz, A.; Mieloszyk, E. Macroelements and essential oil content in the raw material of the selected medicinal plants species from organic cultivation. J. Res. Appl. Agric. Eng. 2006, 51, 161–163. [Google Scholar]
- Seidler-Łożykowska, K.; Golcz, A.; Kozik, E.; Kucharski, W.; Mordalski, R.; Wójcik, J. Evaluation of quality of savory (Satureja hortensis L.) herb from organic cultivation. J. Res. Appl. Agric. Eng. 2007, 52, 48–51. [Google Scholar]
- Mumivand, H.; Babalar, M.; Hadian, J.; Tabatabaei, S.M.F. Influence of nitrogen and calcium carbonate application rates on the minerals content of summer savory (Satureja hortensis L.) leaves. Hort. Env. Biot. 2010, 51, 173–177. [Google Scholar]
- Karimi, N.; Yari, M.; Ghasmpour, H.R. Identification and comparison of essential oil composition and mineral changes in different phenological stages of Satureja hortensis L. Iran J. Plant Phys. 2012, 3, 577–582. [Google Scholar]
- Soltani, F.; Heidari, S.; Azizi, M.; Hadian, J. Influence of Ca(NO3)2 and KNO3 Application on Biomass, Yield, Oil and Mineral Contents of Tarragon in “Ray” Region. J. Agric. Sci. 2014, 7, 19–29. [Google Scholar] [CrossRef]
- Kłosowska, K. Plant Responses to salinity. Kosmos 2010, 59, 539–549. [Google Scholar]
- Siyal, A.A.; Siyal, A.G.; Abro, Z.A. Salt affected soils their identification and reclamation. Pak. J. Appl. Sci. 2002, 2, 537–540. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Zhu, J. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Boil. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Sankar, B.; Sridharan, R.; Panneerselvam, R. Soil salinity alters growth, chlorophyll content, and secondary metabolite accumulation in Catharanthus roseus. Turk. J. Bot. 2008, 32, 79–83. [Google Scholar]
- Baranauskienė, R.; Venskutonis, P.R.; Viškelis, P.; Dambrauskienė, E. Influence of Nitrogen Fertilizers on the Yield and Composition of Thyme (Thymus vulgaris). J. Agric. Food Chem. 2003, 51, 7751–7758. [Google Scholar] [CrossRef]
- Nowacki, E. Nitrogen Management of Crops; PWRiL: Warsaw, Polish, 1980; pp. 47–50, 64–71. [Google Scholar]
- Fageria, V.D. Nutrient Interactions in Crop Plants. J. Plant Nutr. 2001, 24, 1269–1290. [Google Scholar] [CrossRef]
- Grzebisz, W. Fertilization of Crops; PWRiL: Warsaw, Polish, 2015. [Google Scholar]
- Szewczuk, C.; Mazur, M. Influence of different doses of nitrogen fertilizers on the chemical composition of nettle (Urtica dioica L.) harvested in three development stages, part II mineral content. Acta Scien. Pol. Agricul. 2004, 3, 239–248. [Google Scholar]
- Fabek, S.; Toth, N.; Redovniković, I.R.; Ćustić, M.H.; Benko, B.; Žutić, I. The effect of nitrogen fertilization on nitrate accumulation, and the content of minerals and glucosinolates in broccoli cultivars. Food Technol. Biotechnol. 2012, 50, 183–191. [Google Scholar]
- Kozera, W.; Majcherczak, E.; Barczak, B. Effect of varied NPK fertilisation on the yield size, content of essential oil and mineral composition of caraway fruit (Carum carvi L.). J. Elem. 2012, 8, 255–267. [Google Scholar] [CrossRef]
- Jalili, F. The effect of N, P, and micronutrients on yield and essential oil of Satureja hortensis L. Advan. Env. Biol. 2015, 9, 860–865. [Google Scholar]
- Ghaly, F.; Baddour, G.; El-Azazy, H. Nitrate Accumulation and Oxalate Formation in Spinach Plants (Spinacia oleracea, L.) as Affected by Nitrogen Fertilization levels and Iron Foliar Application. J. Soil Sci. Agric. Eng. 2017, 8, 571–576. [Google Scholar] [CrossRef]
- Baligar, V.C.; Fageria, N.K.; He, Z. Nutrient Use Efficiency in Plants. Commun. Soil Sci. Plant Anal. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Wang, Y.; Wu, W.-H. Membrane Transporters for Nitrogen, Phosphate and Potassium Uptake in Plants. J. Integr. Plant Boil. 2008, 50, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Rietra, R.; Heinen, M.; Dimkpa, C.O.; Bindraban, P.S. Effects of Nutrient Antagonism and Synergism on Yield and Fertilizer Use Efficiency. Commun. Soil Sci. Plant Anal. 2017, 48, 1895–1920. [Google Scholar] [CrossRef]
- Geilfus, C.-M. Chloride in soil: From nutrient to soil pollutant. Environ. Exp. Bot. 2019, 157, 299–309. [Google Scholar] [CrossRef]
- Jakobsen, S.T. Interaction between Plant Nutrients: III. Antagonism between Potassium, Magnesium and Calcium. Acta Agric. Scand. Sect. B Plant Soil Sci. 1993, 43, 1–5. [Google Scholar] [CrossRef]
- Singh, M.; Khan, M.M.A.; Naeem, M. Effect of nitrogen on growth, nutrient assimilation, essential oil content, yield and quality attributes in Zingiber officinale Rosc. J. Saudi Soc. Agric. Sci. 2016, 15, 171–178. [Google Scholar] [CrossRef]
- Dzida, K.; Jarosz, Z. Impact of nitrogen-potassium fertilization on the yield and chemical composition of summer savory (Satureja hortensis L.). Acta Agroph. 2006, 7, 879–884. [Google Scholar]
- Nurzyńska-Wierdak, R.; Rożek, E.; Borowski, B. Response of different basil cultivars to nitrogen and potassium fertilization: Total and mineral nitrogen content in herb. Acta Sci. Pol. Hort. Cult. 2011, 10, 217–232. [Google Scholar]
- Effect of nitrogen fertilization on morphological and biochemical traits of some Apiaceae crops under arid regions in Egypt. Nusant. Biosci. 2013, 5, 15–21. [CrossRef]
- Hendawy, S.F. Comparative study of organic and mineral fertilization on Plantago arenaria plant. J. Appl. Sci. Res. 2008, 4, 500–506. [Google Scholar]
- Singh, M.; Rao, R.S.G.; Ramesh, S. Effects of N and K on growth, herbage, oil yield and nutrient uptake patterns in rosemary (Rosmarinus officinalis L.) under semi-arid tropical conditions. J. Hortic. Sci. Biotechnol. 2007, 82, 414–419. [Google Scholar] [CrossRef]
- Golcz, A.; Politycka, B.; Seidler-Łożykowska, K. The effect of nitrogen fertilization and stage of plant development on the mass and quality of sweet basil leaves (Ocimum basilicum L.). Herba Pol. 2006, 52, 22–30. [Google Scholar]
- Dzida, K.; Zawiślak, G.; Karczmarz, K. Yields and quality of three herbal species from Lamiaceae familly. J. Elem. 2015, 20, 273–283. [Google Scholar] [CrossRef]
- Santamaria, P.; Elia, A.; Serio, F. Effect of Solution Nitrogen Concentration on Yield, Leaf Element Content, and Water and Nitrogen Use Efficiency of Three Hydroponically-Grown Rocket Salad Genotypes. J. Plant Nutr. 2002, 25, 245–258. [Google Scholar] [CrossRef]
- Nurzyńska-Wierdak, R. Growth and yield of garden rocket (Eruca sativa Mill.) affected by nitrogen and potassium fertilization. Acta Sci. Pol. Hort. Cult. 2009, 8, 23–33. [Google Scholar]
- Baričevič, D.; Zupančič, A. The Impact of Drought Stress and/or Nitrogen Fertilization in Some Medicinal Plants. J. Herbs Spices Med. Plants 2002, 9, 53–64. [Google Scholar] [CrossRef]
- Jakobsen, S.T. Interaction between plant nutrients: I. theory and analytical procedures. Acta Agric. Scand. Sect. B Soil Plant Scien. 1992, 42, 208–212. [Google Scholar] [CrossRef]
- Kafkafi, U.; Xu, G.; Imas, P.; Magen, H.; Tarchitzky, J.; Johnston, A.E. Potassium and chloride in crops and soils: The role of potassium chloride fertilizer in crop nutrition. In IPI Reserch Topics 22; Johnston, A., Ed.; International Potash Institute: Berne, Switzerland, 2002. [Google Scholar]
- Johnston, A.E.; Polulton, P.R.; Syers, J.K. Phosphorus, Potassium and Sulfur Cycles in Agricultural Soils; International Fertiliser Society: York, UK, 2001. [Google Scholar]
- Nurzyńska-Wierdak, R.; Rożek, E.; Borowski, B.; Dzida, K.; Jarosz, Z. Changes in the content of some macronutrients in basil herbage induced by different nitrogen and potassium fertilization rates. J. Elem. 2012, 13, 465–477. [Google Scholar] [CrossRef]
- Kafkafi, U.; Valoras, N.; Letey, J. Chloride interaction with nitrate and phosphate nutrition in tomato (Lycopersicon esculentum L.). J. Plant Nutr. 1982, 5, 1369–1385. [Google Scholar] [CrossRef]
- David, E.; Rothstein, D. Effects of nitrogen from on nutrient uptake and physiology of Abies fraseri. J. For. Ecol. Manag. 2005, 219, 69–80. [Google Scholar]
- Grzebisz, W.; Potarzycki, J. Factors affecting phosphorus uptake by plants. J. Elem. 2003, 8, 47–60. [Google Scholar]
- Sapek, B. Accumulation and release of phosphorus in soil—Sources, processes, causes. Wat. Env. Rur. Are. 2014, 14, 77–100. [Google Scholar]
- Abbas, M.K. Effect of calcium nitrate, potassium nitrate and anfaton on growth and storability of plastic houses cucumber (Cucumis sativus L. cv. Al-Hytham). Am. J. Plant Physiol. 2010, 5, 278–290. [Google Scholar] [CrossRef]
- Haneklaus, S.; Bloem, E.; Schnug, E. Sulphur in agroecosystems. Fol. Univ. Tetin. 2000, 204, 17–32. [Google Scholar]
- Jamal, A.; Moon, Y.S.; Abdin, M.Z. Sulphur-a general overview and interaction with nitrogen. Aust. J. Crop Sci. 2010, 4, 523–529. [Google Scholar]
- Fazli, I.S.; Jamal, A.; Ahmad, S.; Masoodi, M.; Khan, J.S.; Abdin, M.Z. Interactive effect of sulphur and nitrogen on nitrogen accumulation and harvest in oilseed crops differing in nitrogen assimilation potential. J. Plant Nutr. 2008, 31, 1203–1220. [Google Scholar] [CrossRef]
- Zhao, F.-J.; Bilsborrow, P.E.; Evans, E.J.; McGrath, S.P. Nitrogen to sulphur ratio in rapeseed and in rapeseed protein and its use in diagnosing sulphur deficiency. J. Plant Nutr. 1997, 20, 549–558. [Google Scholar] [CrossRef]
- Li, N.; Yang, Y.; Wang, L.; Zhou, C.; Jing, J.; Sun, X.; Tian, X. Combined effects of nitrogen and sulfur fertilization on maize growth, physiological traits, N and S uptake, and their diagnosis. Field Crop. Res. 2019, 242, 107593. [Google Scholar] [CrossRef]
- Bar, Y.; Apelbaum, A.; Kafkafi, U.; Goren, R. Relationship between chloride and nitrate and its effect on growth and mineral composition of avocado and citrus plants. J. Plant Nutr. 1997, 20, 715–731. [Google Scholar] [CrossRef]
- Chen, W.; He, Z.L.; Yang, X.E.; Mishra, S.; Stoffella, P.J. Chlorine Nutrition of Higher Plants: Progress and Perspectives. J. Plant Nutr. 2010, 33, 943–952. [Google Scholar] [CrossRef]
- Nurzyńska-Wierdak, R.; Dzida, K. Influence of plant density and term of harvest on yield and chemical composition of sweet marjoram (Origanum majorana L.). Acta Sci Pol. Hort. Cult. 2009, 8, 51–61. [Google Scholar]
- Dzida, K. Yield and Biological Value of Thyme (Thymus vulgaris L.) and Savory (Satureia hortensis L.) Herbage Depending on Nitrogen and Potassium Fertilization; Wydawnictwo Uniwersytetu Przyrodniczego w Lublinie: Lublin, Poland, 2013. [Google Scholar]
- Esmaili, E.; Kapourchal, S.; Malakouti, M.; Homaee, M. Interactive effect of salinity and two nitrogen fertilizers on growth and composition of sorghum. Plant Soil Environ. 2008, 54, 537–546. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
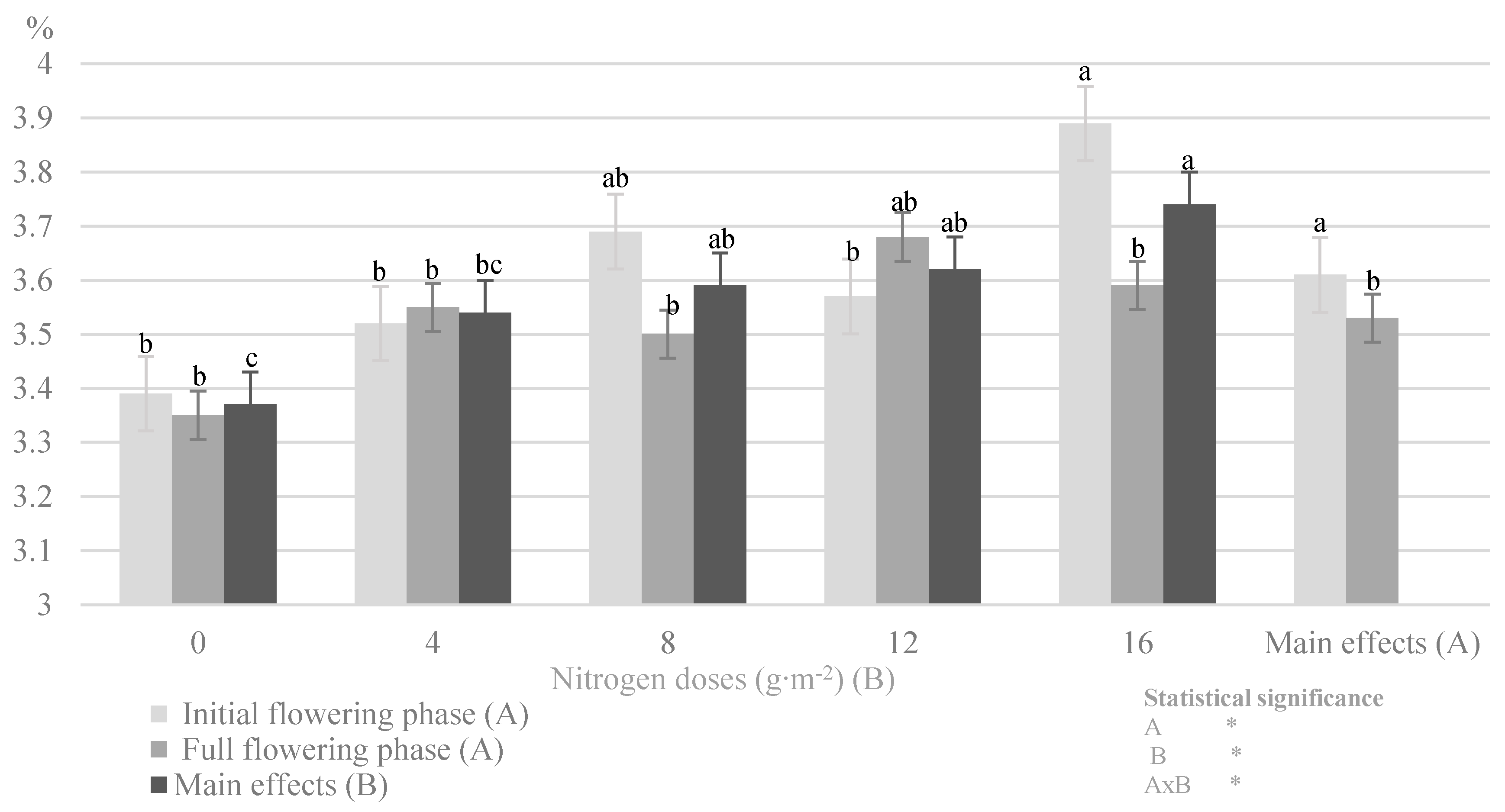
| Nitrogen Dose (g∙m−2) (B) | N-NH4 (% DM) | N-NO3 (% DM) | ||||
|---|---|---|---|---|---|---|
| Plant Development Phases (A) | ||||||
| Beginning of Flowering | Full Flowering | Main Effects (B) | Beginning of Flowering | Full Flowering | Main Effects (B) | |
| 0 | 0.054 ± 0.031 bc | 0.013 ± 0.001 cd | 0.028 ± 0.034 b | 0.073 ± 0.039 ab | 0.002 ± 0.001 c | 0.038 ± 0.045 bc |
| 4 | 0.045 ± 0.026 bc | 0.065 ± 0.036 ab | 0.055 ± 0.032 a | 0.080 ± 0.009 ab | 0.069 ± 0.010 ab | 0.074 ± 0.010 a |
| 8 | 0.021 ± 0.006 c | 0.043 ± 0.020 b | 0.032 ± 0.018 b | 0.091 ± 0.042 a | 0.052 ± 0.008 b | 0.072 ± 0.035 ab |
| 12 | 0.092 ± 0.002 a | 0.020 ± 0.001 c | 0.056 ± 0.037 a | 0.079 ± 0.017 ab | 0.022 ± 0.003 c | 0.051 ± 0.032 b |
| 16 | 0.015 ± 0.006 c | 0.039 ± 0.002 bc | 0.027 ± 0.013 b | 0.087 ± 0.021 a | 0.011 ± 0.002 c | 0.049 ± 0.042 b |
| Main effects (A) | 0.045 ± 0.033 a | 0.034 ± 0.028 b | 0.082 ± 0.027 a | 0.031 ± 0.026 b | ||
| Statistical significance | ||||||
| A | * | * | ||||
| B | * | * | ||||
| AxB | * | * | ||||
| Plant Development Phases (A) | Nitrogen Dose (g∙m−2) (B) | P | K | Ca | Mg | S | Cl |
|---|---|---|---|---|---|---|---|
| (% DM) | |||||||
| Beginning of flowering | 0 | 0.33 ± 0.029 bc | 2.15 ± 0.066 b | 1.21 ± 0.22 d | 0.46 ± 0.055 a | 0.29 ± 0.014 bc | 0.57 ± 0.116 |
| 4 | 0.36 ± 0.036 b | 2.19 ± 0.056 a | 1.55 ± 0.02 cd | 0.34 ± 0.013 c | 0.35 ± 0.012 ab | 0.34 ± 0.079 | |
| 8 | 0.39 ± 0.042 a | 2.07 ± 0.117 b | 1.67 ± 0.11 ab | 0.35 ± 0.013 c | 0.36 ± 0.017 ab | 0.45 ± 0.050 | |
| 12 | 0.37 ± 0.024 ab | 2.19 ± 0.061 a | 1.65 ± 0.19 b | 0.33 ± 0.036 c | 0.32 ± 0.026 b | 0.73 ± 0.218 | |
| 16 | 0.38 ± 0.030 ab | 2.30 ± 0.026 a | 1.57 ± 0.07 c | 0.32 ± 0.010 c | 0.38 ± 0.041 a | 0.85 ± 0.288 | |
| Full flowering | 0 | 0.32 ± 0.063 bc | 2.15 ± 0.039 b | 0.77 ± 0.07 e | 0.45 ± 0.030 a | 0.30 ± 0.022 bc | 0.58 ± 0.125 |
| 4 | 0.31 ± 0.041 c | 2.20 ± 0.086 a | 1.61 ± 0.29 bc | 0.40 ± 0.014 b | 0.33 ± 0.012 b | 0.35 ± 0.056 | |
| 8 | 0.39 ± 0.024 a | 1.69 ± 0.052 c | 1.70 ± 0.32 a | 0.44 ± 0.021 a | 0.32 ± 0.025 b | 0.57 ± 0.103 | |
| 12 | 0.34 ± 0.012 bc | 1.57 ± 0.106 c | 1.65 ± 0.38 b | 0.40 ± 0.014 b | 0.29 ± 0.007 c | 0.69 ± 0.284 | |
| 16 | 0.38 ± 0.026 ab | 1.50 ± 0.074 c | 1.37 ± 0.22 d | 0.30 ± 0.011 c | 0.28 ± 0.010 c | 0.80 ± 0.291 | |
| Main effects Beginning of flowering Full flowering | 0.37 ± 0.031 a | 2.18 ± 0.318 a | 1.53 ± 0.21 | 0.36 ± 0.061 b | 0.34 ± 0.039 a | 0.59 ± 0.251 | |
| 0.35 ± 0.040 b | 1.82 ± 0.309 b | 1.42 ± 0.43 | 0.40 ± 0.057 a | 0.30 ± 0.025 b | 0.60 ± 0.242 | ||
| Nitrogen dose | |||||||
| 0 | 0.32 ± 0.047 b | 2.15 ± 0.052 a | 0.99 ± 0.28 b | 0.45 ± 0.043 a | 0.30 ± 0.018 b | 0.57 ± 0.117 b | |
| 4 | 0.34 ± 0.045 ab | 2.20 ± 0.069a | 1.58 ± 0.20 a | 0.37 ± 0.033 b | 0.34 ± 0.016 a | 0.34 ± 0.067 c | |
| 8 | 0.39 ± 0.030 a | 1.89 ± 0.214 b | 1.68 ± 0.23 a | 0.39 ± 0.055 b | 0.35 ± 0.030 a | 0.51 ± 0.101 b | |
| 12 | 0.35 ± 0.021 ab | 1.88 ± 0.335 b | 1.65 ± 0.29 a | 0.36 ± 0.046 b | 0.30 ± 0.025 b | 0.71 ± 0.248 a | |
| 16 | 0.38 ± 0.026 a | 1.91 ± 0.424 b | 1.47 ± 0.18 a | 0.31 ± 0.014 c | 0.33 ± 0.061 a | 0.82 ± 0.283 a | |
| Statistical significance | |||||||
| A | * | * | n.s. | * | * | n.s. | |
| B | * | * | * | * | * | * | |
| AxB | * | * | * | * | * | n.s. | |
| Plant Development Phases (A) | Nitrogen Dose (g∙m−2) (B) | HA | SEC | TSC | EC | Corg | pHH2O |
|---|---|---|---|---|---|---|---|
| cmol(+)·kg−1 | (mS∙cm−1) | (%) | |||||
| Beginning of flowering | 0 | 3.16 ± 0.04 ab | 5.11 ± 0.04 b | 8.27 ± 0.02 c | 0.087 ± 0.008 | 0.772 ± 0.004 d | 5.85 |
| 4 | 3.15 ± 0.04 b | 5.33 ± 0.04 a | 8.48 ± 0.04 a | 0.115 ± 0.001 | 0.817 ± 0.002 c | 5.90 | |
| 8 | 2.78 ± 0.03 c | 3.91 ± 0.02 f | 6.69 ± 0.03 f | 0.126 ± 0.033 | 0.777 ± 0.002 d | 5.81 | |
| 12 | 3.21 ± 0.02 ab | 5.16 ± 0.04 b | 8.37 ± 0.04 b | 0.121 ± 0.029 | 0.842 ± 0.001 b | 5.97 | |
| 16 | 3.22 ± 0.02 a | 3.12 ± 0.03 g | 6.34 ± 0.03 g | 0.145 ± 0.034 | 0.893 ± 0.003 a | 5.89 | |
| Full flowering | 0 | 2.78 ± 0.05 c | 4.90 ± 0.03 c | 7.78 ± 0.03 d | 0.085 ± 0.014 | 0.603 ± 0.003 h | 6.03 |
| 4 | 2.80 ± 0.02 c | 4.32 ± 0.02 e | 7.12 ± 0.03 e | 0.101 ± 0.011 | 0.617 ± 0.003 g | 6.20 | |
| 8 | 2.43 ± 0.04 d | 4.31 ± 0.02 e | 6.74 ± 0.03 f | 0.108 ± 0.020 | 0.847 ± 0.002 b | 6.33 | |
| 12 | 2.42 ± 0.03 d | 4.72 ± 0.03 d | 7.14 ± 0.04 e | 0.130 ± 0.046 | 0.662 ± 0.002 f | 6.33 | |
| 16 | 2.81 ± 0.02 c | 3.91 ± 0.02 f | 6.72 ± 0.04 f | 0.121 ± 0.029 | 0.763 ± 0.002 e | 6.19 | |
| Main effects Beginning of flowering Full flowering | 3.10 ± 0.17 a | 4.53 ± 0.87 a | 7.63 ± 0.93 a | 0.119 ± 0.034 | 0.820 ± 0.045 a | 5.81–5.97 | |
| 2.65 ± 0.18 b | 4.44 ± 0.35 b | 7.08 ± 0.37 b | 0.109 ± 0.029 | 0.699 ± 0.094 b | 6.03–6.33 | ||
| Nitrogen dose | |||||||
| 0 | 2.97 ± 0.20 a | 5.01 ± 0.11 a | 7.98 ± 0.28 a | 0.086 ± 0.011 b | 0.689 ± 0.089 e | 5.85–6.03 | |
| 4 | 2.98 ± 0.18 a | 4.83 ± 0.52 c | 7.80 ± 0.71 b | 0.108 ± 0.025 ab | 0.716 ± 0.107 d | 5.90–6.20 | |
| 8 | 2.60 ± 0.17 c | 4.11 ± 0.20 d | 6.72 ± 0.39 d | 0.117 ± 0.028 a | 0.811 ± 0.046 b | 5.81–6.33 | |
| 12 | 2.81 ± 0.41 b | 4.94 ± 0.22 b | 7.75 ± 0.61 c | 0.126 ± 0.032 a | 0.752 ± 0.096 c | 5.97–6.33 | |
| 16 | 3.01 ± 0.21 a | 3.52 ± 0.41 e | 6.53 ± 0.20 e | 0.133 ± 0.036 a | 0.828 ± 0.069 a | 5.89–6.19 | |
| Statistical significance | - | ||||||
| A | * | * | * | n.s | * | ||
| B | * | * | * | * | * | ||
| AxB | * | * | * | n.s | * | ||
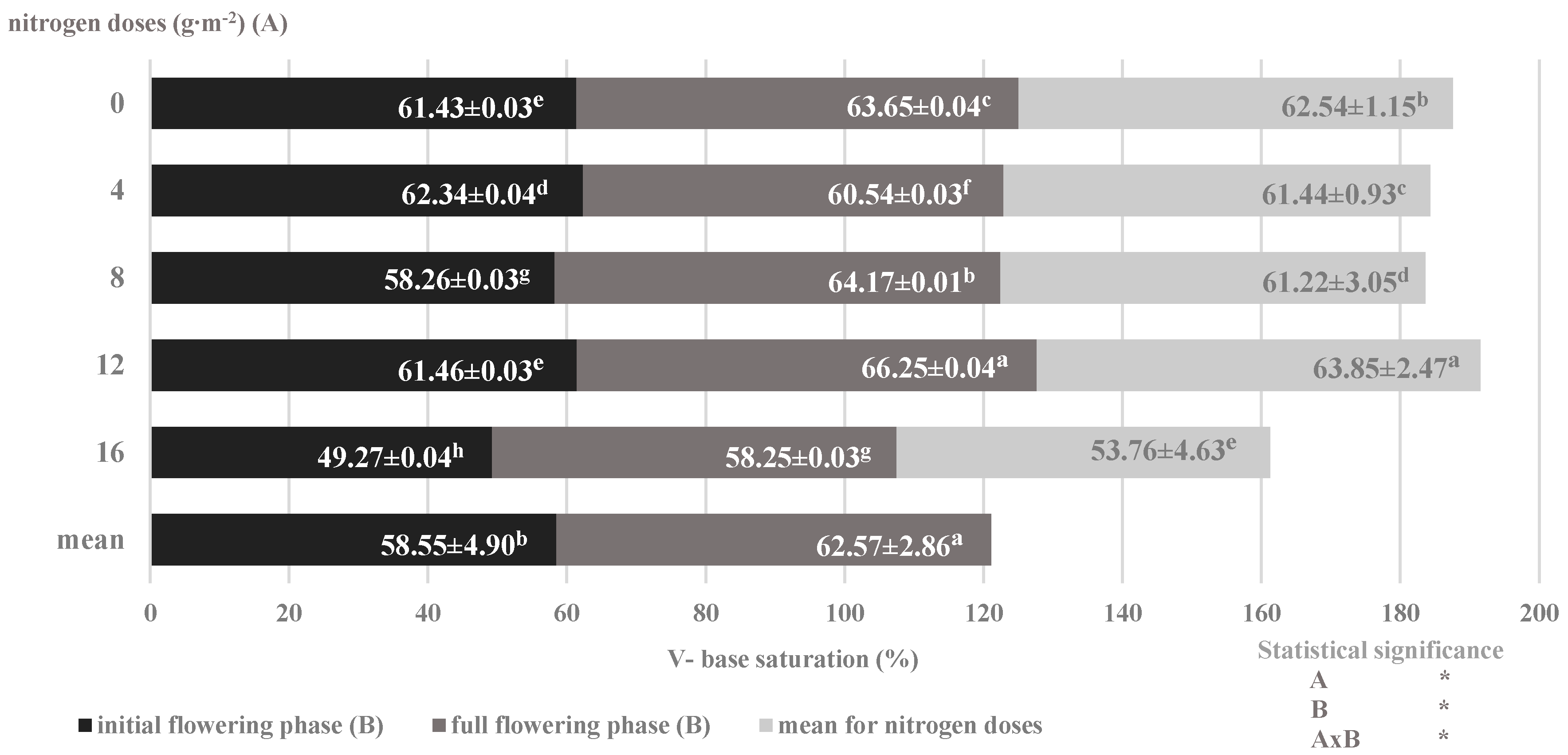
| Plant Development Phases (A) | Nitrogen Dose (g∙m−2) (B) | N-NH4 | N-NO3 | P-PO4 | K | Ca | Mg | S-SO4 | Cl |
|---|---|---|---|---|---|---|---|---|---|
| (mg∙dm−3) | |||||||||
| Beginning of flowering | 0 | 10.75 ± 0.54 cd | 36.57 ± 0.62 ab | 22.64 ± 1.08 d | 50.65 ± 0.51 | 331.83 ± 1.53 | 35.85 ± 0.44 | 19.02 ± 0.65 c | 46.96 ± 0.93 b |
| 4 | 15.95 ± 0.38 bc | 27.65 ± 0.39 bc | 31.08 ± 0.89 bc | 65.75 ± 0.69 | 330.50 ± 2.09 | 50.57 ± 0.62 | 22.12 ± 0.74 c | 26.46 ± 0.86 c | |
| 8 | 11.90 ± 0.98 cd | 54.07 ± 0.67 a | 31.57 ± 0.94 bc | 67.95 ± 1.03 | 306.00 ± 1.49 | 48.77 ± 0.37 | 21.60 ± 0.62 c | 46.10 ± 0.43 b | |
| 12 | 14.52 ± 0.75 bc | 24.50 ± 0.48 bc | 35.63 ± 0.84 ab | 65.45 ± 0.79 | 317.00 ± 1.98 | 42.30 ± 0.26 | 36.80 ± 0.88 ab | 63.63 ± 0.56 a | |
| 16 | 16.45 ± 0.40 b | 25.20 ± 0.51 bc | 30.13 ± 1.05 bc | 58.80 ± 0.80 | 297.16 ± 0.92 | 53.35 ± 0.83 | 34.45 ± 0.64 ab | 42.24 ± 0.37 b | |
| Full flowering | 0 | 013.47 ± 0.94 bcd | 10.75 ± 0.54 c | 33.24 ± 0.52 ab | 63.82 ± 0.52 | 365.66 ± 1.48 | 47.00 ± 0.19 | 23.50 ± 0.82 bc | 21.55 ± 0.33 d |
| 4 | 48.75 ± 0.66 d | 14.35 ± 0.48 c | 34.77 ± 0.72 ab | 83.45 ± 072 | 373.16 ± 1.86 | 60.90 ± 0.10 | 33.05 ± 0.53 ab | 31.87 ± 0.28 c | |
| 8 | 815.72 ± 0.41 bc | 13.65 ± 0.36 c | 33.88 ± 0.35 ab | 70.90 ± 0.88 | 431.66 ± 2.12 | 59.87 ± 0.49 | 48.90 ± 0.73 a | 64.35 ± 0.42 a | |
| 12 | 1217.85 ± 1.03 ab | 22.05 ± 0.45 bc | 37.34 ± 0.83 a | 83.25 ± 0.90 | 335.16 ± 1.70 | 55.00 ± 0.34 | 32.95 ± 0.43 bc | 69.31 ± 1.18 a | |
| 16 | 1621.87 ± 0.76 a | 24.15 ± 0.50 bc | 29.67 ± 0.89 c | 70.87 ± 0.35 | 367.16 ± 1.62 | 60.62 ± 0.39 | 31.87 ± 0.59 bc | 51.06 ± 0.80 b | |
| Main effects Beginning of flowering Full flowering Nitrogen dose 0 4 8 12 16 | 13.91 ± 0.61b | 33.60 ± 1.02 a | 30.21 ± 0.95 b | 61.72 ± 0.84 b | 316.50 ± 1.64 b | 46.17 ± 0.96 b | 26.80 ± 0.98 a | 45.08 ± 0.62 | |
| 15.53 ± 1.13 a | 16.99 ± 0.66 b | 33.78 ± 0.89 a | 74.46 ± 0.97 a | 374.56 ± 1.97 a | 56.68 ± 1.16 a | 34.05 ± 1.02 a | 47.63 ± 0.73 | ||
| 12.11 ± 0.94 b | 23.66 ± 0.41 ab | 27.94 ± 1.06 c | 57.23 ± 0.78 | 348.75 ± 1.40 | 41.42 ± 0.67 c | 21.26 ± 0.35 b | 34.25 ± 0.78 d | ||
| 12.33 ± 0.28 b | 21.00 ± 0.30 b | 32.92 ± 0.72 b | 74.60 ± 0.59 | 351.83 ± 1.03 | 55.73 ± 0.69 a | 27.58 ± 0.31 ab | 29.16 ± 0.38 d | ||
| 13.81 ± 0.59 b | 33.86 ± 0.84 a | 32.73 ± 0.69 b | 69.42 ± 0.43 | 368.83 ± 1.66 | 54.32 ± 0.72 ab | 35.25 ± 0.73 a | 55.23 ± 0.20 b | ||
| 16.18 ± 0.60 ab | 23.27 ± 0.45 ab | 36.49 ± 0.95 a | 74.35 ± 0.47 | 326.08 ± 1.25 | 48.65 ± 0.73 b | 34.87 ± 0.39 a | 66.47 ± 0.54 a | ||
| 19.16 ± 0.81 a | 24.67 ± 0.21 ab | 29.90 ± 0.99 bc | 64.83 ± 0.34 | 332.17 ± 0.92 | 56.98 ± 0.81 a | 33.16 ± 0.55 a | 46.65 ± 0.59 c | ||
| Statistical significance | |||||||||
| A | * | * | * | * | * | * | * | n.s. | |
| B | * | * | * | n.s. | n.s. | * | * | * | |
| AxB | * | * | * | n.s. | n.s. | n.s. | * | * | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skubij, N.; Dzida, K.; Jarosz, Z.; Pitura, K.; Jaroszuk-Sierocińska, M. Nutritional Value of Savory Herb (Satureja hortensis L.) and Plant Response to Variable Mineral Nutrition Conditions in Various Phases of Development. Plants 2020, 9, 706. https://doi.org/10.3390/plants9060706
Skubij N, Dzida K, Jarosz Z, Pitura K, Jaroszuk-Sierocińska M. Nutritional Value of Savory Herb (Satureja hortensis L.) and Plant Response to Variable Mineral Nutrition Conditions in Various Phases of Development. Plants. 2020; 9(6):706. https://doi.org/10.3390/plants9060706
Chicago/Turabian StyleSkubij, Natalia, Katarzyna Dzida, Zbigniew Jarosz, Karolina Pitura, and Monika Jaroszuk-Sierocińska. 2020. "Nutritional Value of Savory Herb (Satureja hortensis L.) and Plant Response to Variable Mineral Nutrition Conditions in Various Phases of Development" Plants 9, no. 6: 706. https://doi.org/10.3390/plants9060706
APA StyleSkubij, N., Dzida, K., Jarosz, Z., Pitura, K., & Jaroszuk-Sierocińska, M. (2020). Nutritional Value of Savory Herb (Satureja hortensis L.) and Plant Response to Variable Mineral Nutrition Conditions in Various Phases of Development. Plants, 9(6), 706. https://doi.org/10.3390/plants9060706