Co-Translational Protein Folding and Sorting in Chloroplasts


Abstract
1. Introduction
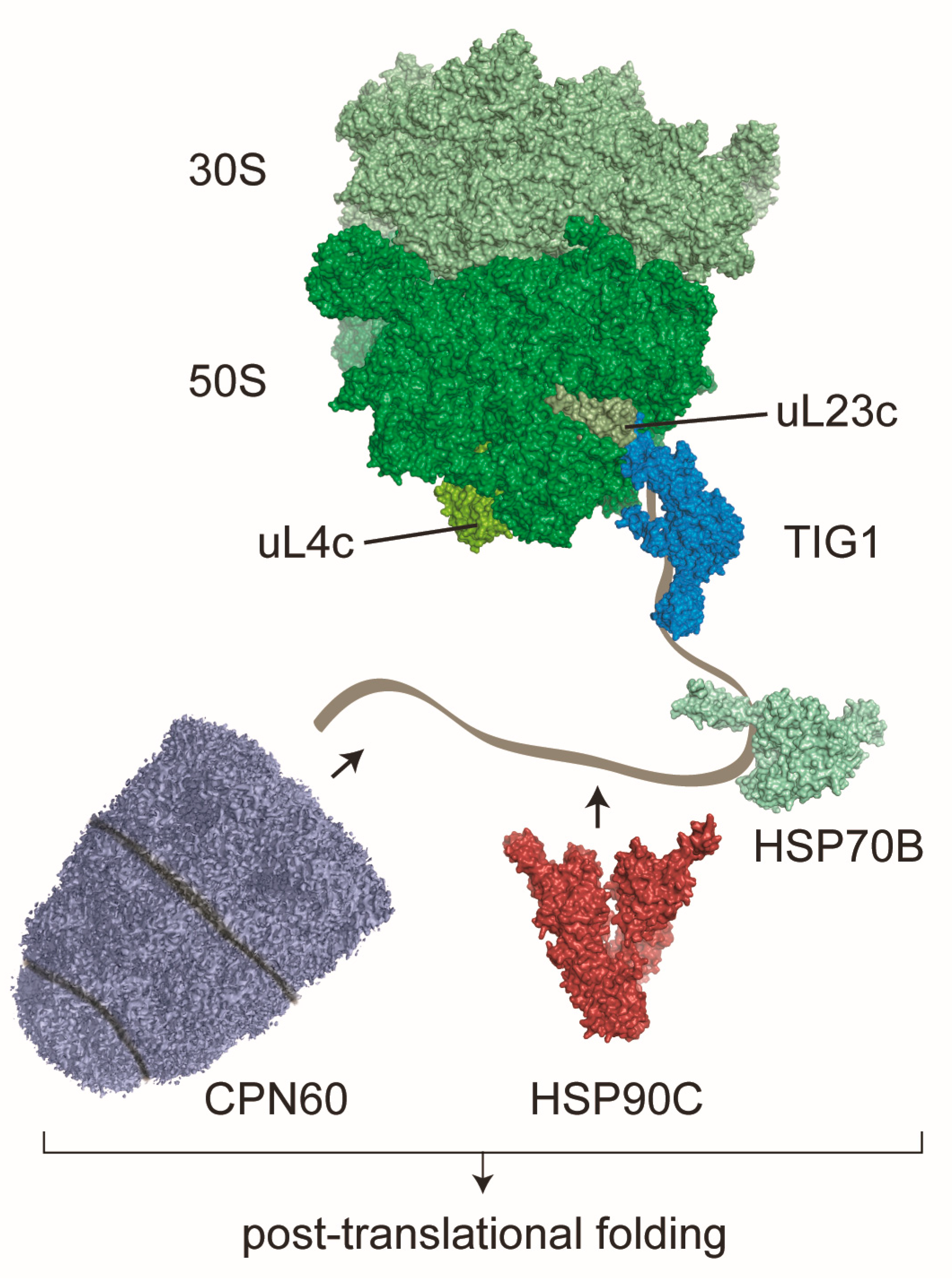
2. The Ribosome Associated Molecular Chaperone Trigger Factor

3. Other Co-Translationally Acting Chaperones in Chloroplasts
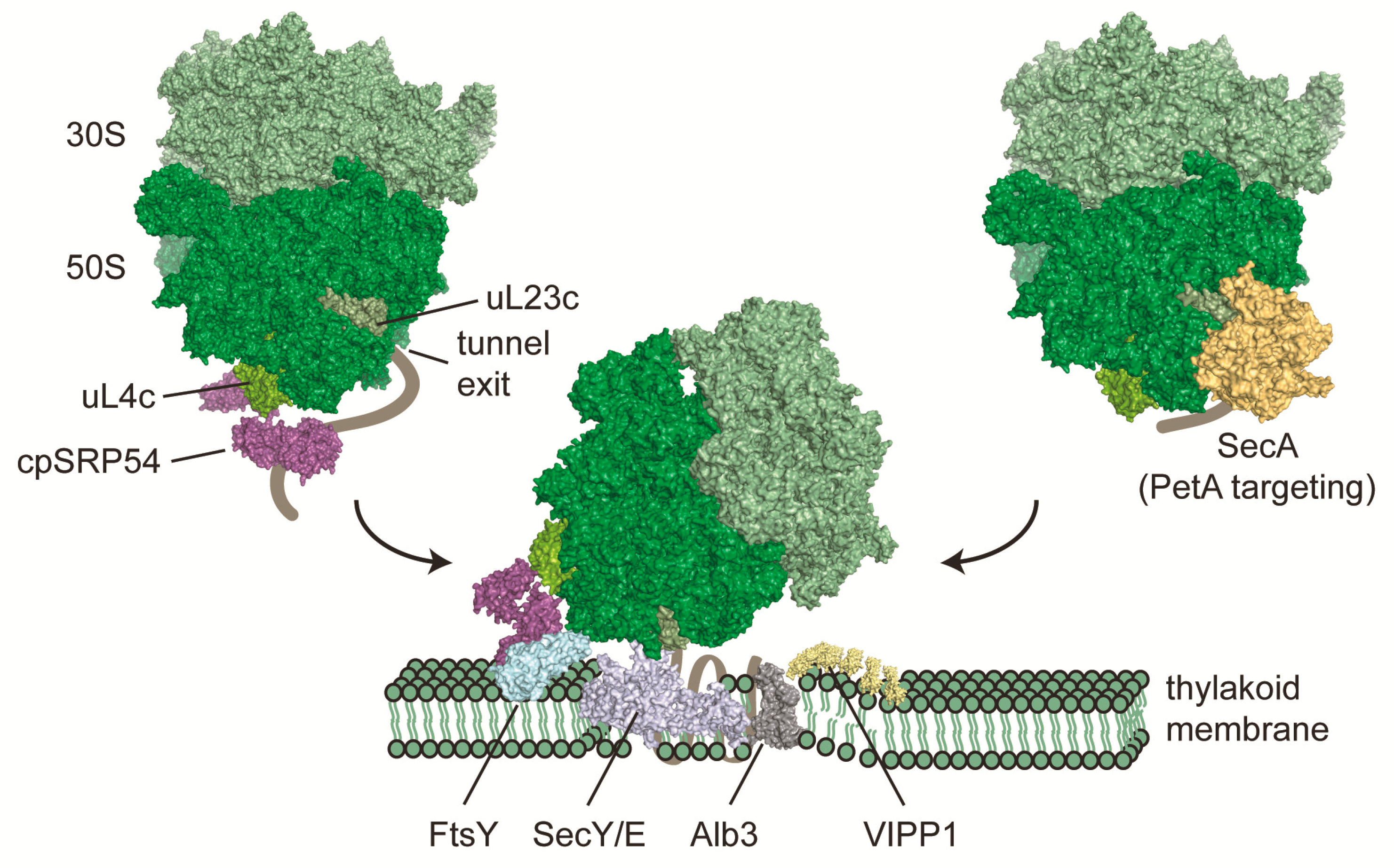
4. Co-Translational Targeting of Chloroplast-Encoded Proteins to Thylakoid Membranes

5. Ribosome Binding Sites of Co-Translationally Acting Factors
6. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. In vivo aspects of protein folding and quality control. Science 2016, 353. [Google Scholar] [CrossRef] [PubMed]
- Kramer, G.; Shiber, A.; Bukau, B. Mechanisms of Cotranslational Maturation of Newly Synthesized Proteins. Annu. Rev. Biochem. 2019, 88, 337–364. [Google Scholar] [CrossRef] [PubMed]
- Deuerling, E.; Gamerdinger, M.; Kreft, S.G. Chaperone Interactions at the Ribosome. Cold Spring Harb. Perspect. Biol. 2019, 11. [Google Scholar] [CrossRef]
- Zhang, G.; Ignatova, Z. Folding at the birth of the nascent chain: Coordinating translation with co-translational folding. Curr. Opin. Struct. Biol. 2011, 21, 25–31. [Google Scholar] [CrossRef]
- Woolhead, C.A.; McCormick, P.J.; Johnson, A.E. Nascent membrane and secretory proteins differ in FRET-detected folding far inside the ribosome and in their exposure to ribosomal proteins. Cell 2004, 116, 725–736. [Google Scholar] [CrossRef]
- Kosolapov, A.; Deutsch, C. Tertiary interactions within the ribosomal exit tunnel. Nat. Struct. Mol. Biol. 2009, 16, 405–411. [Google Scholar] [CrossRef]
- Nilsson, O.B.; Hedman, R.; Marino, J.; Wickles, S.; Bischoff, L.; Johansson, M.; Müller-Lucks, A.; Trovato, F.; Puglisi, J.D.; O’Brien, E.P.; et al. Cotranslational Protein Folding inside the Ribosome Exit Tunnel. Cell Rep. 2015, 12, 1533–1540. [Google Scholar] [CrossRef]
- Pechmann, S.; Willmund, F.; Frydman, J. The ribosome as a hub for protein quality control. Mol. Cell 2013, 49, 411–421. [Google Scholar] [CrossRef]
- Breiman, A.; Fieulaine, S.; Meinnel, T.; Giglione, C. The intriguing realm of protein biogenesis: Facing the green co-translational protein maturation networks. Biochim. Biophys. Acta 2016, 1864, 531–550. [Google Scholar] [CrossRef]
- Giglione, C.; Fieulaine, S.; Meinnel, T. N-terminal protein modifications: Bringing back into play the ribosome. Biochimie 2015, 114, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Aviram, N.; Schuldiner, M. Targeting and translocation of proteins to the endoplasmic reticulum at a glance. J. Cell Sci 2017, 130, 4079–4085. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.C.; Jan, C.H.; Weissman, J.S. Targeting and plasticity of mitochondrial proteins revealed by proximity-specific ribosome profiling. Science 2014, 346, 748–751. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F. Why chloroplasts and mitochondria retain their own genomes and genetic systems: Colocation for redox regulation of gene expression. Proc. Natl. Acad. Sci. USA 2015, 112, 10231–10238. [Google Scholar] [CrossRef]
- Maier, U.G.; Zauner, S.; Woehle, C.; Bolte, K.; Hempel, F.; Allen, J.F.; Martin, W.F. Massively convergent evolution for ribosomal protein gene content in plastid and mitochondrial genomes. Genome Biol. Evol. 2013, 5, 2318–2329. [Google Scholar] [CrossRef]
- Leister, D. Retrograde signaling in plants: From simple to complex scenarios. Front. Plant Sci. 2012, 3, 135. [Google Scholar] [CrossRef]
- Pfannschmidt, T.; Schütze, K.; Fey, V.; Sherameti, I.; Oelmüller, R. Chloroplast redox control of nuclear gene expression—A new class of plastid signals in interorganellar communication. Antioxid. Redox Signal. 2003, 5, 95–101. [Google Scholar] [CrossRef]
- Bock, R. Plastid biotechnology: Prospects for herbicide and insect resistance, metabolic engineering and molecular farming. Curr. Opin. Biotechnol. 2007, 18, 100–106. [Google Scholar] [CrossRef]
- Zoschke, R.; Bock, R. Chloroplast Translation: Structural and Functional Organization, Operational Control and Regulation. Plant Cell 2018, 30, 745–770. [Google Scholar] [CrossRef]
- Barkan, A. Expression of plastid genes: Organelle-specific elaborations on a prokaryotic scaffold. Plant Physiol. 2011, 155, 1520–1532. [Google Scholar] [CrossRef]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef] [PubMed]
- Nickelsen, J.; Bohne, A.-V.; Westhoff, P. Chloroplast gene expression-translation. In Plastid Biology; Springer: New York, NY, USA, 2014; Volume 5, pp. 49–78. [Google Scholar] [CrossRef]
- Trösch, R.; Mühlhaus, T.; Schroda, M.; Willmund, F. ATP-dependent molecular chaperones in plastids-More complex than expected. Biochim. Biophys. Acta 2015, 1847, 872–888. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.E. Two paths diverged in the stroma: Targeting to dual SEC translocase systems in chloroplasts. Photosynth. Res. 2018, 138, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Ziehe, D.; Dünschede, B.; Schünemann, D. From bacteria to chloroplasts: Evolution of the chloroplast SRP system. Biol. Chem. 2017, 398, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Rochaix, J.D.; Bassi, R. LHC-like proteins involved in stress responses and biogenesis/repair of the photosynthetic apparatus. Biochem. J. 2019, 476, 581–593. [Google Scholar] [CrossRef]
- Kobayashi, K. Role of membrane glycerolipids in photosynthesis, thylakoid biogenesis and chloroplast development. J. Plant Res. 2016, 129, 565–580. [Google Scholar] [CrossRef]
- Bohne, A.V.; Schwenkert, S.; Grimm, B.; Nickelsen, J. Roles of Tetratricopeptide Repeat Proteins in Biogenesis of the Photosynthetic Apparatus. Int. Rev. Cell Mol. Biol. 2016, 324, 187–227. [Google Scholar] [CrossRef]
- Plöchinger, M.; Schwenkert, S.; von Sydow, L.; Schröder, W.P.; Meurer, J. Functional Update of the Auxiliary Proteins PsbW, PsbY, HCF136, PsbN, TerC and ALB3 in Maintenance and Assembly of PSII. Front. Plant Sci. 2016, 7, 423. [Google Scholar] [CrossRef]
- Bastien, O.; Botella, C.; Chevalier, F.; Block, M.A.; Jouhet, J.; Breton, C.; Girard-Egrot, A.; Marechal, E. New Insights on Thylakoid Biogenesis in Plant Cells. Int. Rev. Cell Mol. Biol. 2016, 323, 1–30. [Google Scholar] [CrossRef]
- Rast, A.; Heinz, S.; Nickelsen, J. Biogenesis of thylakoid membranes. Biochim. Biophys. Acta 2015, 1847, 821–830. [Google Scholar] [CrossRef]
- Crooke, E.; Wickner, W. Trigger factor: A soluble protein that folds pro-OmpA into a membrane-assembly-competent form. Proc. Natl. Acad. Sci. USA 1987, 84, 5216–5220. [Google Scholar] [CrossRef] [PubMed]
- Lill, R.; Crooke, E.; Guthrie, B.; Wickner, W. The “trigger factor cycle” includes ribosomes, presecretory proteins, and the plasma membrane. Cell 1988, 54, 1013–1018. [Google Scholar] [CrossRef]
- Hoffmann, A.; Bukau, B.; Kramer, G. Structure and function of the molecular chaperone Trigger Factor. Biochim. Biophys. Acta 2010, 1803, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Ferbitz, L.; Maier, T.; Patzelt, H.; Bukau, B.; Deuerling, E.; Ban, N. Trigger factor in complex with the ribosome forms a molecular cradle for nascent proteins. Nature 2004, 431, 590–596. [Google Scholar] [CrossRef]
- Deeng, J.; Chan, K.Y.; van der Sluis, E.O.; Berninghausen, O.; Han, W.; Gumbart, J.; Schulten, K.; Beatrix, B.; Beckmann, R. Dynamic Behavior of Trigger Factor on the Ribosome. J. Mol. Biol. 2016, 428, 3588–3602. [Google Scholar] [CrossRef]
- Oh, E.; Becker, A.H.; Sandikci, A.; Huber, D.; Chaba, R.; Gloge, F.; Nichols, R.J.; Typas, A.; Gross, C.A.; Kramer, G.; et al. Selective ribosome profiling reveals the cotranslational chaperone action of trigger factor in vivo. Cell 2011, 147, 1295–1308. [Google Scholar] [CrossRef]
- Saio, T.; Guan, X.; Rossi, P.; Economou, A.; Kalodimos, C.G. Structural basis for protein antiaggregation activity of the trigger factor chaperone. Science 2014, 344, 1250494. [Google Scholar] [CrossRef]
- Patzelt, H.; Rüdiger, S.; Brehmer, D.; Kramer, G.; Vorderwülbecke, S.; Schaffitzel, E.; Waitz, A.; Hesterkamp, T.; Dong, L.; Schneider-Mergener, J.; et al. Binding specificity of Escherichia coli trigger factor. Proc. Natl. Acad. Sci. USA 2001, 98, 14244–14249. [Google Scholar] [CrossRef]
- Deuerling, E.; Schulze-Specking, A.; Tomoyasu, T.; Mogk, A.; Bukau, B. Trigger factor and DnaK cooperate in folding of newly synthesized proteins. Nature 1999, 400, 693–696. [Google Scholar] [CrossRef]
- Teter, S.A.; Houry, W.A.; Ang, D.; Tradler, T.; Rockabrand, D.; Fischer, G.; Blum, P.; Georgopoulos, C.; Hartl, F.U. Polypeptide flux through bacterial Hsp70: DnaK cooperates with trigger factor in chaperoning nascent chains. Cell 1999, 97, 755–765. [Google Scholar] [CrossRef]
- Mashaghi, A.; Kramer, G.; Bechtluft, P.; Zachmann-Brand, B.; Driessen, A.J.; Bukau, B.; Tans, S.J. Reshaping of the conformational search of a protein by the chaperone trigger factor. Nature 2013, 500, 98–101. [Google Scholar] [CrossRef]
- Ries, F.; Carius, Y.; Rohr, M.; Gries, K.; Keller, S.; Lancaster, C.R.D.; Willmund, F. Structural and molecular comparison of bacterial and eukaryotic trigger factors. Sci. Rep. 2017, 7, 10680. [Google Scholar] [CrossRef]
- Olinares, P.D.; Ponnala, L.; van Wijk, K.J. Megadalton complexes in the chloroplast stroma of Arabidopsis thaliana characterized by size exclusion chromatography, mass spectrometry, and hierarchical clustering. Mol. Cell. Proteom. 2010, 9, 1594–1615. [Google Scholar] [CrossRef]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef]
- Rohr, M.; Ries, F.; Herkt, C.; Gotsmann, V.L.; Westrich, L.D.; Gries, K.; Trösch, R.; Christmann, J.; Chaux, F.; Jung, M.; et al. The role of plastidic trigger factor serving protein biogenesis in green algae and land plants. Plant Physiol. 2019. [Google Scholar] [CrossRef]
- Bai, C.; Guo, P.; Zhao, Q.; Lv, Z.; Zhang, S.; Gao, F.; Gao, L.; Wang, Y.; Tian, Z.; Wang, J.; et al. Protomer Roles in Chloroplast Chaperonin Assembly and Function. Mol. Plant 2015, 8, 1478–1492. [Google Scholar] [CrossRef]
- Willmund, F.; Dorn, K.V.; Schulz-Raffelt, M.; Schroda, M. The chloroplast DnaJ homolog CDJ1 of Chlamydomonas reinhardtii is part of a multichaperone complex containing HSP70B, CGE1, and HSP90C. Plant Physiol. 2008, 148, 2070–2082. [Google Scholar] [CrossRef]
- Willmund, F.; Mühlhaus, T.; Wojciechowska, M.; Schroda, M. The NH2-terminal domain of the chloroplast GrpE homolog CGE1 is required for dimerization and cochaperone function in vivo. J. Biol. Chem. 2007, 282, 11317–11328. [Google Scholar] [CrossRef]
- Nilsson, O.B.; Müller-Lucks, A.; Kramer, G.; Bukau, B.; von Heijne, G. Trigger Factor Reduces the Force Exerted on the Nascent Chain by a Cotranslationally Folding Protein. J. Mol. Biol. 2016, 428, 1356–1364. [Google Scholar] [CrossRef]
- Martinez-Hackert, E.; Hendrickson, W.A. Promiscuous substrate recognition in folding and assembly activities of the trigger factor chaperone. Cell 2009, 138, 923–934. [Google Scholar] [CrossRef]
- Bieri, P.; Leibundgut, M.; Saurer, M.; Boehringer, D.; Ban, N. The complete structure of the chloroplast 70S ribosome in complex with translation factor pY. EMBO J. 2017, 36, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Kityk, R.; Kopp, J.; Sinning, I.; Mayer, M.P. Structure and dynamics of the ATP-bound open conformation of Hsp70 chaperones. Mol. Cell 2012, 48, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhang, X.; Sommer, F.; Ta, N.; Wang, N.; Schroda, M.; Cong, Y.; Liu, C. Hetero-oligomeric CPN60 resembles highly symmetric group-I chaperonin structure revealed by Cryo-EM. Plant J. 2019, 98, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Vorderwülbecke, S.; Kramer, G.; Merz, F.; Kurz, T.A.; Rauch, T.; Zachmann-Brand, B.; Bukau, B.; Deuerling, E. Low temperature or GroEL/ES overproduction permits growth of Escherichia coli cells lacking trigger factor and DnaK. FEBS Lett. 2004, 559, 181–187. [Google Scholar] [CrossRef]
- Calloni, G.; Chen, T.; Schermann, S.M.; Chang, H.C.; Genevaux, P.; Agostini, F.; Tartaglia, G.G.; Hayer-Hartl, M.; Hartl, F.U. DnaK functions as a central hub in the E. coli chaperone network. Cell Rep. 2012, 1, 251–264. [Google Scholar] [CrossRef]
- Hayer-Hartl, M.; Bracher, A.; Hartl, F.U. The GroEL-GroES Chaperonin Machine: A Nano-Cage for Protein Folding. Trends Biochem. Sci. 2016, 41, 62–76. [Google Scholar] [CrossRef]
- Ying, B.W.; Taguchi, H.; Kondo, M.; Ueda, T. Co-translational involvement of the chaperonin GroEL in the folding of newly translated polypeptides. J. Biol. Chem. 2005, 280, 12035–12040. [Google Scholar] [CrossRef]
- Albanese, V.; Yam, A.Y.; Baughman, J.; Parnot, C.; Frydman, J. Systems analyses reveal two chaperone networks with distinct functions in eukaryotic cells. Cell 2006, 124, 75–88. [Google Scholar] [CrossRef]
- Westrich, L.D.; Gotsmann, V.L.; Herkt, C.; Ries, F.; Kazek, T.; Trösch, R.; Ramundo, S.; Nickelsen, J.; Armbruster, L.; Wirtz, M.; et al. The proteomic inventory reveals the chloroplast ribosome as nexus within a diverse protein network. bioRxiv 2019. [Google Scholar] [CrossRef]
- Vitlin Gruber, A.; Feiz, L. Rubisco Assembly in the Chloroplast. Front. Mol. Biosci. 2018, 5, 24. [Google Scholar] [CrossRef]
- Wilson, R.H.; Hayer-Hartl, M. Complex Chaperone Dependence of Rubisco Biogenesis. Biochemistry 2018, 57, 3210–3216. [Google Scholar] [CrossRef]
- Meyer, D.I.; Dobberstein, B. Identification and characterization of a membrane component essential for the translocation of nascent proteins across the membrane of the endoplasmic reticulum. J. Cell Biol. 1980, 87, 503–508. [Google Scholar] [CrossRef]
- Walter, P.; Blobel, G. Purification of a membrane-associated protein complex required for protein translocation across the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 1980, 77, 7112–7116. [Google Scholar] [CrossRef]
- Walter, P.; Blobel, G. Signal recognition particle contains a 7S RNA essential for protein translocation across the endoplasmic reticulum. Nature 1982, 299, 691–698. [Google Scholar] [CrossRef]
- Poritz, M.A.; Bernstein, H.D.; Strub, K.; Zopf, D.; Wilhelm, H.; Walter, P. An E. coli ribonucleoprotein containing 4.5S RNA resembles mammalian signal recognition particle. Science 1990, 250, 1111–1117. [Google Scholar] [CrossRef]
- Grudnik, P.; Bange, G.; Sinning, I. Protein targeting by the signal recognition particle. Biol. Chem. 2009, 390, 775–782. [Google Scholar] [CrossRef]
- Walter, P.; Johnson, A.E. Signal sequence recognition and protein targeting to the endoplasmic reticulum membrane. Annu. Rev. Cell Biol. 1994, 10, 87–119. [Google Scholar] [CrossRef]
- Pool, M.R.; Stumm, J.; Fulga, T.A.; Sinning, I.; Dobberstein, B. Distinct modes of signal recognition particle interaction with the ribosome. Science 2002, 297, 1345–1348. [Google Scholar] [CrossRef]
- von Heijne, G. Transcending the impenetrable: How proteins come to terms with membranes. Biochim. Biophys. Acta 1988, 947, 307–333. [Google Scholar] [CrossRef]
- Izard, J.W.; Kendall, D.A. Signal peptides: Exquisitely designed transport promoters. Mol. Microbiol. 1994, 13, 765–773. [Google Scholar] [CrossRef]
- Valent, Q.A.; Kendall, D.A.; High, S.; Kusters, R.; Oudega, B.; Luirink, J. Early events in preprotein recognition in E. coli: Interaction of SRP and trigger factor with nascent polypeptides. EMBO J. 1995, 14, 5494–5505. [Google Scholar] [CrossRef] [PubMed]
- Paetzel, M.; Dalbey, R.E.; Strynadka, N.C. Crystal structure of a bacterial signal peptidase in complex with a beta-lactam inhibitor. Nature 1998, 396, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Schibich, D.; Gloge, F.; Pöhner, I.; Björkholm, P.; Wade, R.C.; von Heijne, G.; Bukau, B.; Kramer, G. Global profiling of SRP interaction with nascent polypeptides. Nature 2016, 536, 219–223. [Google Scholar] [CrossRef]
- Mary, C.; Scherrer, A.; Huck, L.; Lakkaraju, A.K.; Thomas, Y.; Johnson, A.E.; Strub, K. Residues in SRP9/14 essential for elongation arrest activity of the signal recognition particle define a positively charged functional domain on one side of the protein. RNA 2010, 16, 969–979. [Google Scholar] [CrossRef]
- Beckert, B.; Kedrov, A.; Sohmen, D.; Kempf, G.; Wild, K.; Sinning, I.; Stahlberg, H.; Wilson, D.N.; Beckmann, R. Translational arrest by a prokaryotic signal recognition particle is mediated by RNA interactions. Nat. Struct. Mol. Biol. 2015, 22, 767–773. [Google Scholar] [CrossRef]
- Halic, M.; Becker, T.; Pool, M.R.; Spahn, C.M.; Grassucci, R.A.; Frank, J.; Beckmann, R. Structure of the signal recognition particle interacting with the elongation-arrested ribosome. Nature 2004, 427, 808–814. [Google Scholar] [CrossRef]
- Voorhees, R.M.; Hegde, R.S. Structures of the scanning and engaged states of the mammalian SRP-ribosome complex. Elife 2015, 4. [Google Scholar] [CrossRef]
- Wild, K.; Becker, M.M.M.; Kempf, G.; Sinning, I. Structure, dynamics and interactions of large SRP variants. Biol. Chem. 2019, 401, 63–80. [Google Scholar] [CrossRef]
- Pechmann, S.; Chartron, J.W.; Frydman, J. Local slowdown of translation by nonoptimal codons promotes nascent-chain recognition by SRP in vivo. Nat. Struct. Mol. Biol. 2014, 21, 1100–1105. [Google Scholar] [CrossRef]
- Bornemann, T.; Jöckel, J.; Rodnina, M.V.; Wintermeyer, W. Signal sequence-independent membrane targeting of ribosomes containing short nascent peptides within the exit tunnel. Nat. Struct. Mol. Biol. 2008, 15, 494–499. [Google Scholar] [CrossRef]
- Chartron, J.W.; Hunt, K.C.; Frydman, J. Cotranslational signal-independent SRP preloading during membrane targeting. Nature 2016, 536, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Berndt, U.; Oellerer, S.; Zhang, Y.; Johnson, A.E.; Rospert, S. A signal-anchor sequence stimulates signal recognition particle binding to ribosomes from inside the exit tunnel. Proc. Natl. Acad. Sci. USA 2009, 106, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Noriega, T.R.; Chen, J.; Walter, P.; Puglisi, J.D. Real-time observation of signal recognition particle binding to actively translating ribosomes. Elife 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Noriega, T.R.; Tsai, A.; Elvekrog, M.M.; Petrov, A.; Neher, S.B.; Chen, J.; Bradshaw, N.; Puglisi, J.D.; Walter, P. Signal recognition particle-ribosome binding is sensitive to nascent chain length. J. Biol. Chem. 2014, 289, 19294–19305. [Google Scholar] [CrossRef]
- Luirink, J.; ten Hagen-Jongman, C.M.; van der Weijden, C.C.; Oudega, B.; High, S.; Dobberstein, B.; Kusters, R. An alternative protein targeting pathway in Escherichia coli: Studies on the role of FtsY. EMBO J. 1994, 13, 2289–2296. [Google Scholar] [CrossRef]
- de Leeuw, E.; Poland, D.; Mol, O.; Sinning, I.; ten Hagen-Jongman, C.M.; Oudega, B.; Luirink, J. Membrane association of FtsY, the E. coli SRP receptor. FEBS Lett. 1997, 416, 225–229. [Google Scholar] [CrossRef]
- Ladefoged, S.A.; Christiansen, G. A GTP-binding protein of Mycoplasma hominis: A small sized homolog to the signal recognition particle receptor FtsY. Gene 1997, 201, 37–44. [Google Scholar] [CrossRef]
- Powers, T.; Walter, P. Reciprocal stimulation of GTP hydrolysis by two directly interacting GTPases. Science 1995, 269, 1422–1424. [Google Scholar] [CrossRef]
- Jagath, J.R.; Rodnina, M.V.; Wintermeyer, W. Conformational changes in the bacterial SRP receptor FtsY upon binding of guanine nucleotides and SRP. J. Mol. Biol. 2000, 295, 745–753. [Google Scholar] [CrossRef]
- Shan, S.O.; Walter, P. Induced nucleotide specificity in a GTPase. Proc. Natl. Acad. Sci. USA 2003, 100, 4480–4485. [Google Scholar] [CrossRef]
- Akopian, D.; Shen, K.; Zhang, X.; Shan, S.O. Signal recognition particle: An essential protein-targeting machine. Annu. Rev. Biochem. 2013, 82, 693–721. [Google Scholar] [CrossRef]
- Jarvis, P. Targeting of nucleus-encoded proteins to chloroplasts in plants. New Phytol. 2008, 179, 257–285. [Google Scholar] [CrossRef]
- Schwenkert, S.; Dittmer, S.; Soll, J. Structural components involved in plastid protein import. Essays Biochem. 2018, 62, 65–75. [Google Scholar] [CrossRef]
- Bock, R. Cell and Molecular Biology of Plastids; Springer: Berlin/Heidelberg, Germany, 2007; Volume 1. [Google Scholar]
- Adam, Z.; Charuvi, D.; Tsabari, O.; Knopf, R.R.; Reich, Z. Biogenesis of thylakoid networks in angiosperms: Knowns and unknowns. Plant Mol. Biol. 2011, 76, 221–234. [Google Scholar] [CrossRef]
- Franklin, A.E.; Hoffman, N.E. Characterization of a chloroplast homologue of the 54-kDa subunit of the signal recognition particle. J. Biol. Chem. 1993, 268, 22175–22180. [Google Scholar]
- Rosenblad, M.A.; Zwieb, C.; Samuelsson, T. Identification and comparative analysis of components from the signal recognition particle in protozoa and fungi. BMC Genom. 2004, 5, 5. [Google Scholar] [CrossRef]
- Rosenblad, M.A.; Trager, C.; Schünemann, D. Structural diversity of signal recognition particle RNAs in plastids. Plant Signal. Behav. 2013, 8. [Google Scholar] [CrossRef]
- Richter, C.V.; Träger, C.; Schünemann, D. Evolutionary substitution of two amino acids in chloroplast SRP54 of higher plants cause its inability to bind SRP RNA. FEBS Lett. 2008, 582, 3223–3229. [Google Scholar] [CrossRef]
- Li, X.; Henry, R.; Yuan, J.; Cline, K.; Hoffman, N.E. A chloroplast homologue of the signal recognition particle subunit SRP54 is involved in the posttranslational integration of a protein into thylakoid membranes. Proc. Natl. Acad. Sci. USA 1995, 92, 3789–3793. [Google Scholar] [CrossRef]
- Schünemann, D.; Gupta, S.; Persello-Cartieaux, F.; Klimyuk, V.I.; Jones, J.D.; Nussaume, L.; Hoffman, N.E. A novel signal recognition particle targets light-harvesting proteins to the thylakoid membranes. Proc. Natl. Acad. Sci. USA 1998, 95, 10312–10316. [Google Scholar] [CrossRef]
- Dünschede, B.; Träger, C.; Schröder, C.V.; Ziehe, D.; Walter, B.; Funke, S.; Hofmann, E.; Schünemann, D. Chloroplast SRP54 Was Recruited for Posttranslational Protein Transport via Complex Formation with Chloroplast SRP43 during Land Plant Evolution. J. Biol. Chem. 2015, 290, 13104–13114. [Google Scholar] [CrossRef]
- Funke, S.; Knechten, T.; Ollesch, J.; Schünemann, D. A unique sequence motif in the 54-kDa subunit of the chloroplast signal recognition particle mediates binding to the 43-kDa subunit. J. Biol. Chem. 2005, 280, 8912–8917. [Google Scholar] [CrossRef]
- Hermkes, R.; Funke, S.; Richter, C.; Kuhlmann, J.; Schünemann, D. The alpha-helix of the second chromodomain of the 43 kDa subunit of the chloroplast signal recognition particle facilitates binding to the 54 kDa subunit. FEBS Lett. 2006, 580, 3107–3111. [Google Scholar] [CrossRef]
- Ziehe, D.; Dünschede, B.; Schünemann, D. Molecular mechanism of SRP-dependent light-harvesting protein transport to the thylakoid membrane in plants. Photosynth. Res. 2018, 138, 303–313. [Google Scholar] [CrossRef]
- Hristou, A.; Gerlach, I.; Stolle, D.S.; Neumann, J.; Bischoff, A.; Dünschede, B.; Nowaczyk, M.M.; Zoschke, R.; Schünemann, D. Ribosome-associated chloroplast SRP54 enables efficient co-translational membrane insertion of key photosynthetic proteins. Plant Cell 2019. [Google Scholar] [CrossRef]
- Nilsson, R.; van Wijk, K.J. Transient interaction of cpSRP54 with elongating nascent chains of the chloroplast-encoded D1 protein; ‘cpSRP54 caught in the act’. FEBS Lett. 2002, 524, 127–133. [Google Scholar] [CrossRef]
- Nilsson, R.; Brunner, J.; Hoffman, N.E.; van Wijk, K.J. Interactions of ribosome nascent chain complexes of the chloroplast-encoded D1 thylakoid membrane protein with cpSRP54. EMBO J. 1999, 18, 733–742. [Google Scholar] [CrossRef]
- Piskozub, M.; Kroliczewska, B.; Kroliczewski, J. Ribosome nascent chain complexes of the chloroplast-encoded cytochrome b6 thylakoid membrane protein interact with cpSRP54 but not with cpSecY. J. Bioenerg. Biomembr. 2015, 47, 265–278. [Google Scholar] [CrossRef]
- Zoschke, R.; Barkan, A. Genome-wide analysis of thylakoid-bound ribosomes in maize reveals principles of cotranslational targeting to the thylakoid membrane. Proc. Natl. Acad. Sci. USA 2015, 112, E1678–E1687. [Google Scholar] [CrossRef]
- Luirink, J.; Samuelsson, T.; de Gier, J.W. YidC/Oxa1p/Alb3: Evolutionarily conserved mediators of membrane protein assembly. FEBS Lett. 2001, 501, 1–5. [Google Scholar] [CrossRef]
- Hennon, S.W.; Soman, R.; Zhu, L.; Dalbey, R.E. YidC/Alb3/Oxa1 Family of Insertases. J. Biol. Chem. 2015, 290, 14866–14874. [Google Scholar] [CrossRef] [PubMed]
- Driessen, A.J.; Nouwen, N. Protein translocation across the bacterial cytoplasmic membrane. Annu. Rev. Biochem. 2008, 77, 643–667. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.L.; Hardy, S.J. SecB, one small chaperone in the complex milieu of the cell. Cell. Mol. Life Sci. 2002, 59, 1617–1623. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Bordes, P.; Genevaux, P. Multitasking SecB chaperones in bacteria. Front. Microbiol. 2014, 5, 666. [Google Scholar] [CrossRef]
- Zoschke, R.; Watkins, K.P.; Miranda, R.G.; Barkan, A. The PPR-SMR protein PPR53 enhances the stability and translation of specific chloroplast RNAs in maize. Plant J. 2016, 85, 594–606. [Google Scholar] [CrossRef]
- Röhl, T.; van Wijk, K.J. In vitro reconstitution of insertion and processing of cytochrome f in a homologous chloroplast translation system. J. Biol. Chem. 2001, 276, 35465–35472. [Google Scholar] [CrossRef]
- Voelker, R.; Barkan, A. Two nuclear mutations disrupt distinct pathways for targeting proteins to the chloroplast thylakoid. EMBO J. 1995, 14, 3905–3914. [Google Scholar] [CrossRef]
- Voelker, R.; Mendel-Hartvig, J.; Barkan, A. Transposon-disruption of a maize nuclear gene, tha1, encoding a chloroplast SecA homologue: In vivo role of cp-SecA in thylakoid protein targeting. Genetics 1997, 145, 467–478. [Google Scholar]
- Rütgers, M.; Schroda, M. A role of VIPP1 as a dynamic structure within thylakoid centers as sites of photosystem biogenesis? Plant Signal. Behav. 2013, 8, e27037. [Google Scholar] [CrossRef]
- Wilson, D.N.; Beckmann, R. The ribosomal tunnel as a functional environment for nascent polypeptide folding and translational stalling. Curr. Opin. Struct. Biol. 2011, 21, 274–282. [Google Scholar] [CrossRef]
- Voss, N.R.; Gerstein, M.; Steitz, T.A.; Moore, P.B. The geometry of the ribosomal polypeptide exit tunnel. J. Mol. Biol. 2006, 360, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Kramer, G.; Boehringer, D.; Ban, N.; Bukau, B. The ribosome as a platform for co-translational processing, folding and targeting of newly synthesized proteins. Nat. Struct. Mol. Biol. 2009, 16, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.J.; Soltis, P.S.; Bell, C.D.; Burleigh, J.G.; Soltis, D.E. Phylogenetic analysis of 83 plastid genes further resolves the early diversification of eudicots. Proc. Natl. Acad. Sci. USA 2010, 107, 4623–4628. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.L.; Ruhlman, T.A.; Jansen, R.K. Plastid-Nuclear Interaction and Accelerated Coevolution in Plastid Ribosomal Genes in Geraniaceae. Genome Biol. Evol. 2016, 8, 1824–1838. [Google Scholar] [CrossRef]
- Graf, M.; Arenz, S.; Huter, P.; Dönhöfer, A.; Novácek, J.; Wilson, D.N. Cryo-EM structure of the spinach chloroplast ribosome reveals the location of plastid-specific ribosomal proteins and extensions. Nucleic Acids Res. 2017, 45, 2887–2896. [Google Scholar] [CrossRef]
- Boerema, A.P.; Aibara, S.; Paul, B.; Tobiasson, V.; Kimanius, D.; Forsberg, B.O.; Wallden, K.; Lindahl, E.; Amunts, A. Structure of the chloroplast ribosome with chl-RRF and hibernation-promoting factor. Nat. Plants 2018, 4, 212–217. [Google Scholar] [CrossRef]
- Gabashvili, I.S.; Gregory, S.T.; Valle, M.; Grassucci, R.; Worbs, M.; Wahl, M.C.; Dahlberg, A.E.; Frank, J. The polypeptide tunnel system in the ribosome and its gating in erythromycin resistance mutants of L4 and L22. Mol. Cell 2001, 8, 181–188. [Google Scholar] [CrossRef]
- Trifa, Y.; Privat, I.; Gagnon, J.; Baeza, L.; Lerbs-Mache, S. The nuclear RPL4 gene encodes a chloroplast protein that co-purifies with the T7-like transcription complex as well as plastid ribosomes. J. Biol. Chem. 1998, 273, 3980–3985. [Google Scholar] [CrossRef]
- Sun, Y.; Valente-Paterno, M.I.; Bakhtiari, S.; Law, C.; Zhan, Y.; Zerges, W. Photosystem Biogenesis Is Localized to the Translation Zone in the Chloroplast of Chlamydomonas. Plant Cell 2019. [Google Scholar] [CrossRef]
- Uniacke, J.; Zerges, W. Photosystem II assembly and repair are differentially localized in Chlamydomonas. Plant Cell 2007, 19, 3640–3654. [Google Scholar] [CrossRef]
- Uniacke, J.; Zerges, W. Chloroplast protein targeting involves localized translation in Chlamydomonas. Proc. Natl. Acad. Sci. USA 2009, 106, 1439–1444. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Prokaryotes | Eukaryotes | |||
|---|---|---|---|---|
| Category | Cytosol | Chloroplasts | Mitochondria | |
| Sorting factors | SRP54 (protein & 4.5S RNA) SecA/SecB | SRP54 (protein & 7S RNA) SND1 1 | cpSRP54 (protein only) SecA | unknown |
| Ribosome-associated chaperones | Trigger factor | Hsp70 RAC 2 (Hsp40 & Hsp70) NAC 3 (complex of α & β subunits) | Trigger factor | No trigger factor, others unknown |
| Nascent polypeptide binding chaperones | Hsp70 (DnaK) Chaperonin (GroEL/ES) | Hsp70 Prefoldin Chaperonin | HSP70B HSP90C Chaperonin | unknown |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ries, F.; Herkt, C.; Willmund, F. Co-Translational Protein Folding and Sorting in Chloroplasts. Plants 2020, 9, 214. https://doi.org/10.3390/plants9020214
Ries F, Herkt C, Willmund F. Co-Translational Protein Folding and Sorting in Chloroplasts. Plants. 2020; 9(2):214. https://doi.org/10.3390/plants9020214
Chicago/Turabian StyleRies, Fabian, Claudia Herkt, and Felix Willmund. 2020. "Co-Translational Protein Folding and Sorting in Chloroplasts" Plants 9, no. 2: 214. https://doi.org/10.3390/plants9020214
APA StyleRies, F., Herkt, C., & Willmund, F. (2020). Co-Translational Protein Folding and Sorting in Chloroplasts. Plants, 9(2), 214. https://doi.org/10.3390/plants9020214