Sensitivity Analysis of Italian Lolium spp. to Glyphosate in Agricultural Environments



Abstract
1. Introduction
2. Results
2.1. Dose-Response Experiments
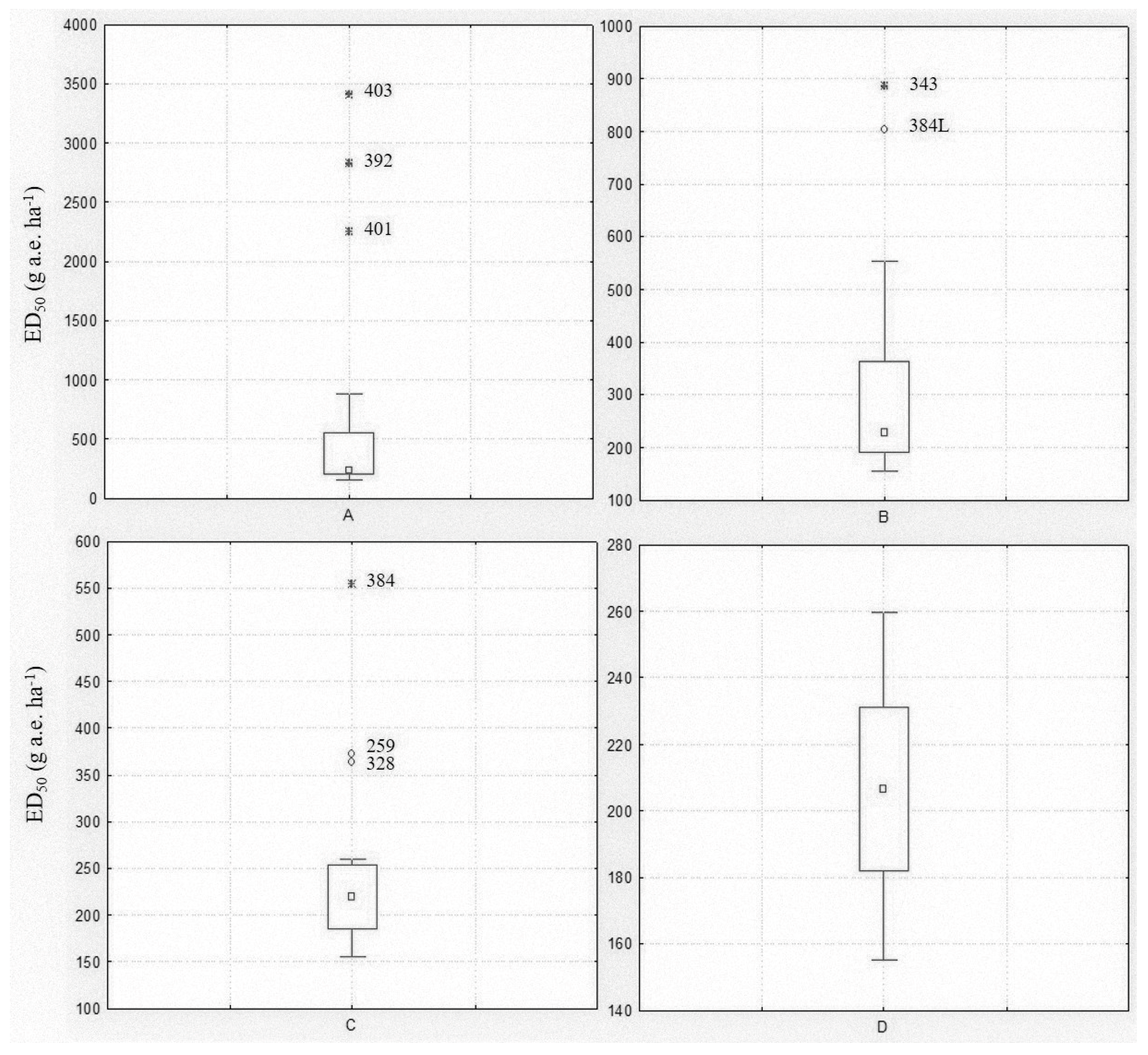
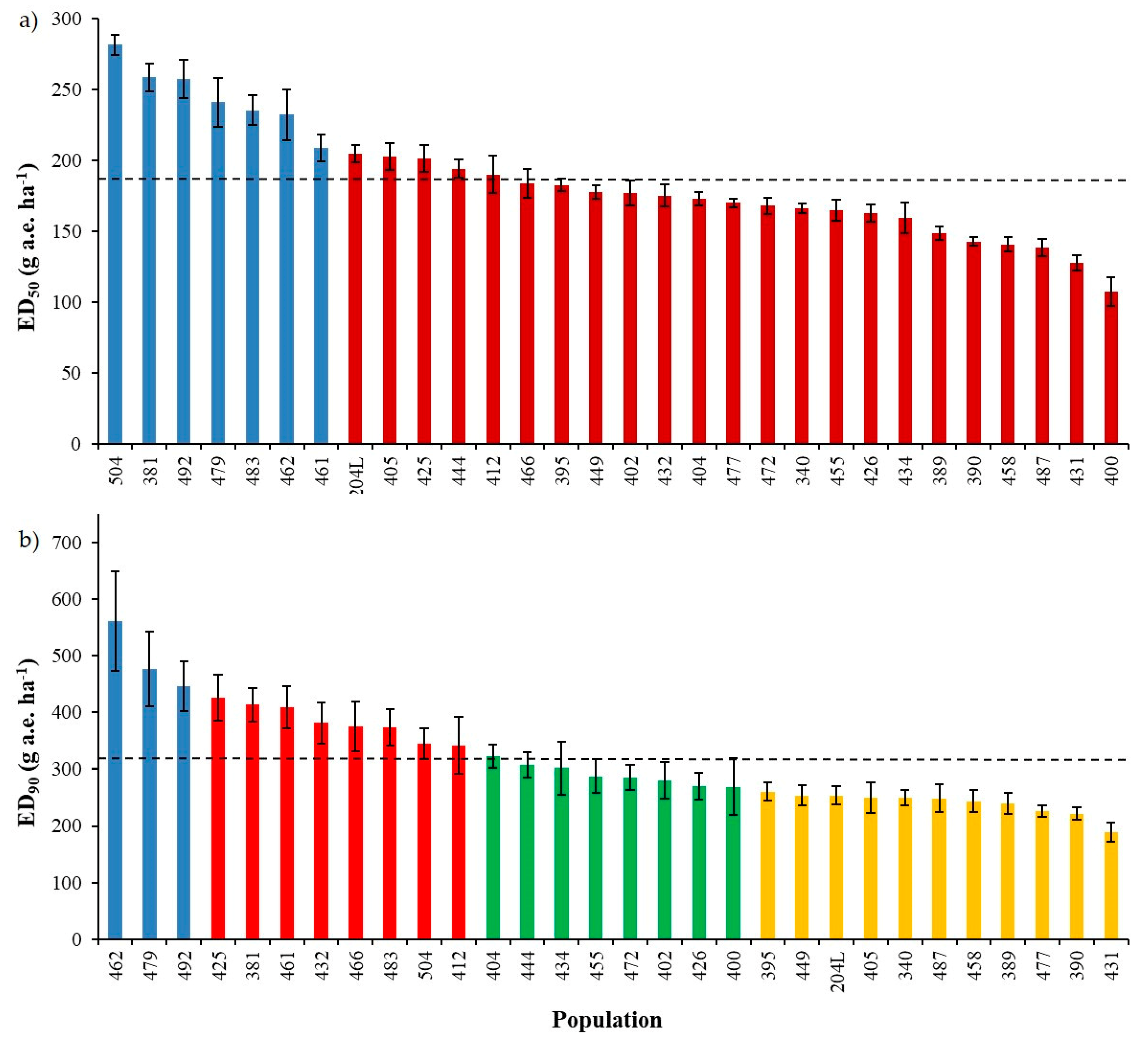
2.1.1. Spring Dose-Response Experiment
2.1.2. Autumn Dose-Response Experiment
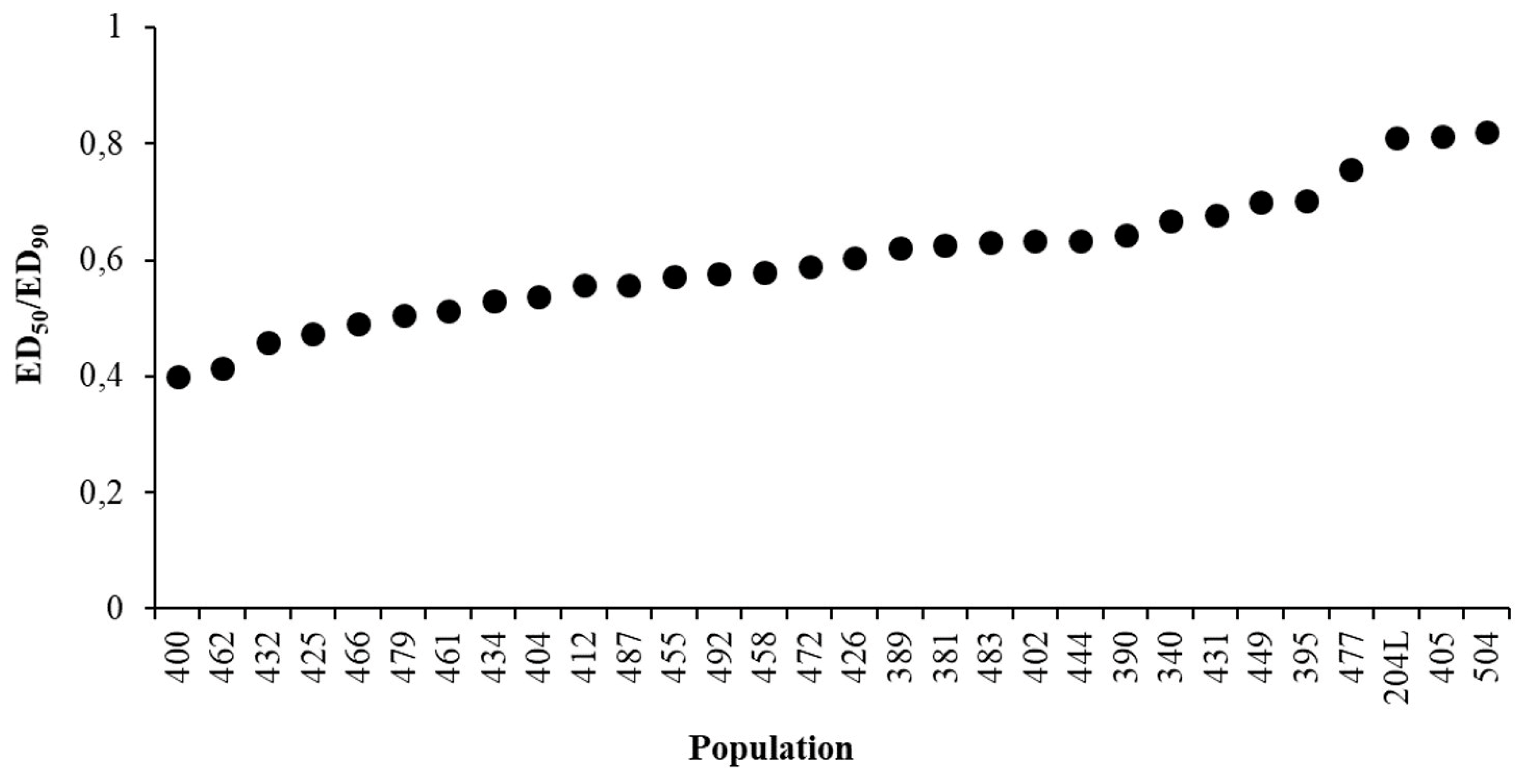
2.2. Sensitivity Line Calculation
3. Discussion
4. Materials and Methods
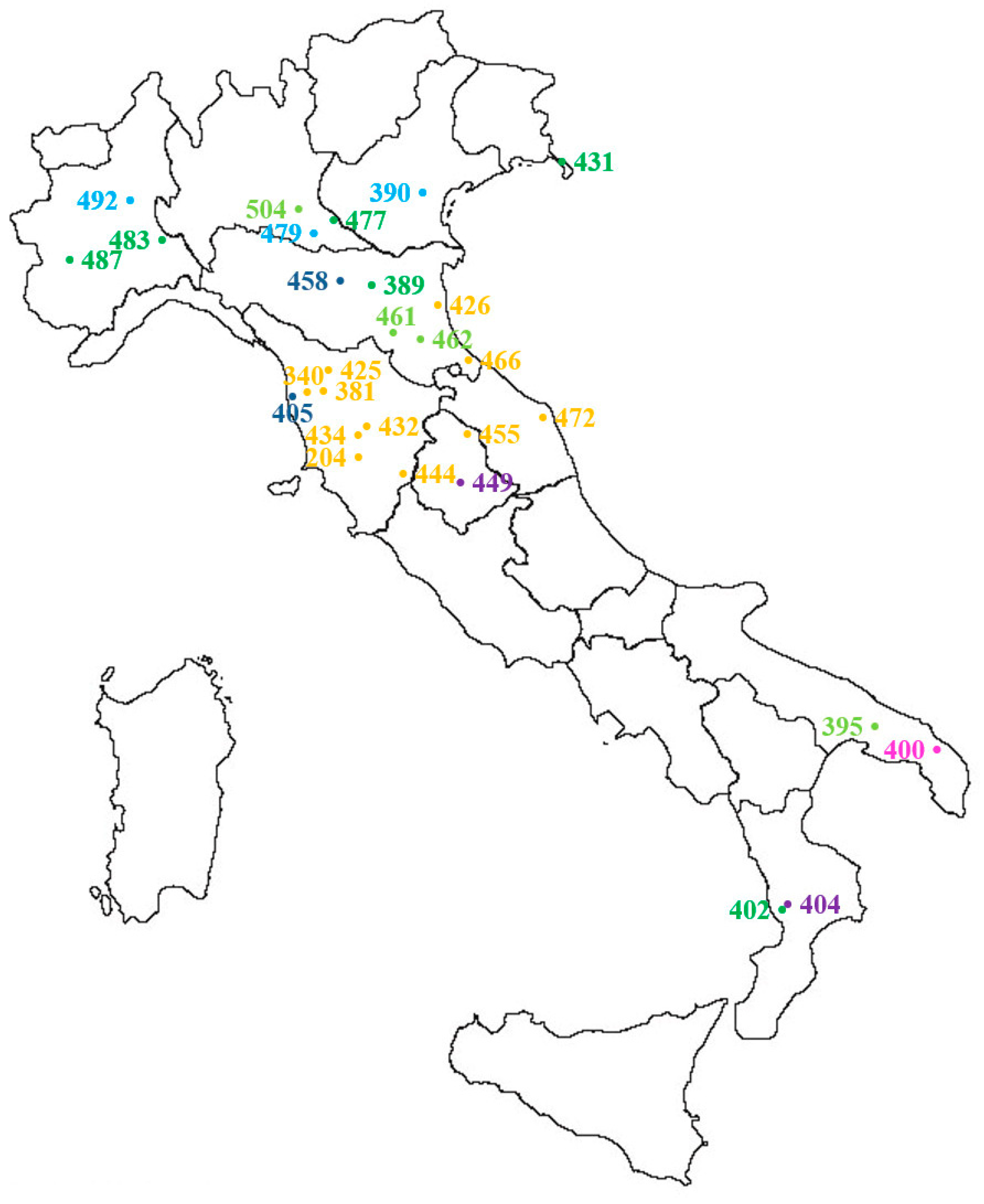
4.1. Plant Material
4.2. Dose-Response Experiments
4.2.1. Spring Dose-Response Experiment
4.2.2. Autumn Dose-Response Experiment
4.3. Statistical Analyses
4.4. Sensitivity Line Calculation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Preston, C.; Wakelin, A.M.; Dolman, F.C.; Bostamam, Y.; Boutsalis, P. A Decade of Glyphosate-Resistant Lolium around the world: Mechanisms, genes, fitness, and agronomic management. Weed Sci. 2009, 57, 435–441. [Google Scholar] [CrossRef]
- Heap, I.; Duke, S.O. Overview of glyphosate-resistant weeds worldwide. Pest Manag. Sci. 2018, 74, 1040–1049. [Google Scholar] [CrossRef]
- Fernández-Moreno, P.T.; Bastida, F.; De Prado, R. Evidence, mechanism and alternative chemical seedbank-level control of glyphosate resistance of a rigid ryegrass (Lolium rigidum) biotype from Southern Spain. Front. Plant Sci. 2017, 8, 1–16. [Google Scholar] [CrossRef]
- Heap, I.M. The International Survey of Herbicide Resistant Weeds. Available online: http://weedscience.org/ (accessed on 24 January 2020).
- Barroso, A.A.; de S Costa, M.G.; Neto, N.J.; dos Santos, J.I.; Balbuena, T.S.; Carbonari, C.A.; Alves, P.L. Protein identification before and after glyphosate exposure in Lolium multiflorum genotypes. Pest Manag. Sci. 2018, 74, 1125–1133. [Google Scholar] [CrossRef]
- Duke, S.O. Why have no new herbicide modes of action appeared in recent years? Pest Manag. Sci. 2012, 68, 505–512. [Google Scholar] [CrossRef]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef]
- Duke, S.; Powles, S. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- Mallory-Smith, C.; Retzinger, J. Revised classification of herbicides by site of action for weed resistance. Weed Technol. 2003, 17, 605–619. [Google Scholar] [CrossRef]
- Solomon, K.R. Estimated exposure to glyphosate in humans via environmental, occupational, and dietary pathways: An updated review of the scientific literature. Pest Manag. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Schönbrunn, E.; Eschenburg, S.; Shuttleworth, W.A.; Schloss, J.V.; Amrhein, N.; Evans, J.N.; Kabsch, W. Interaction of the herbicide glyphosate with its target enzyme 5-enolpyruvylshikimate 3-phosphate synthase in atomic detail. PNAS 2001, 98, 1376–1380. [Google Scholar] [CrossRef] [PubMed]
- Baylis, A.D. Why glyphosate is a global herbicide: Strengths, weaknesses and prospects. Pest Manag. Sci. 2000, 56, 299–308. [Google Scholar] [CrossRef]
- Neve, P.; Diggle, A.J.; Smith, F.P.; Powles, S.B. Simulating evolution of glyphosate resistance in Lolium rigidum I: Population biology of a rare resistance trait. Weed Res. 2003, 43, 404–417. [Google Scholar] [CrossRef]
- Powles, S.B. Evolved glyphosate-resistant weeds around the world: Lessons to be learnt. Pest Manag. Sci. 2008, 64, 360–365. [Google Scholar] [CrossRef]
- Owen, M.D.K. Weed species shifts in glyphosate-resistant crops. Pest Manag. Sci. 2008, 64, 377–387. [Google Scholar] [CrossRef]
- Gonzalez-Torralva, F.; Gil-Humanes, J.; Barro, F.; Dominguez-Valenzuela, J.; De Prado, R. First evidence for a target site mutation in the EPSPS2 gene in glyphosate-resistant Sumatran fleabane from citrus orchards. Agron. Sustain. Dev. 2014, 34, 553–560. [Google Scholar] [CrossRef]
- Urbano, J.M.; Borrego, A.; Torres, V.; Leon, J.M.; Jimenez, C.; Dinelli, G.; Barnes, J. Glyphosate-resistant Hairy Fleabane (Conyza bonariensis) in Spain. Weed Technol. 2007, 21, 396–401. [Google Scholar] [CrossRef]
- Powles, S.B.; Lorraine-colwill, D.F.; Dellow, J.J.; Preston, C. Evolved resistance to glyphosate in rigid ryegrass (Lolium rigidum) in Australia. Weed Sci. 1998, 46, 604–607. [Google Scholar] [CrossRef]
- Collavo, A.; Sattin, M. Resistance to glyphosate in Lolium rigidum selected in Italian perennial crops: Bioevaluation, management and molecular bases of target-site resistance. Weed Res. 2012, 52, 16–24. [Google Scholar] [CrossRef]
- Busi, R.; Powles, S.B. Evolution of glyphosate resistance in a Lolium rigidum population by glyphosate selection at sublethal doses. Heredity 2009, 103, 318–325. [Google Scholar] [CrossRef]
- GIRE—Gruppo Italiano Resistenza Erbicidi. Available online: www.resistenzaerbicidi.it (accessed on 24 January 2020).
- Panozzo, S.; Scarabel, L.; Collavo, A.; Sattin, M. Protocols for robust herbicide resistance testing in different weed species. J. Vis. Exp. 2015, 101, 1–10. [Google Scholar] [CrossRef]
- Vidotto, F.; Tesio, F.; Tabacchi, M.; Ferrero, A. Herbicide sensitivity of Echinochloa spp. accessions in Italian rice fields. Crop Prot. 2007, 26, 285–293. [Google Scholar] [CrossRef]
- Loddo, D.; Kudsk, P.; Costa, B.; Dalla Valle, N.; Sattin, M. Sensitivity analysis of Alisma plantago-aquatica L., Cyperus difformis L. and Schoenoplectus mucronatus (L.) Palla to Penoxsulam. Agronomy 2018, 8, 220. [Google Scholar] [CrossRef]
- Davies, L.R.; Hull, R.; Moss, S.; Neve, P. The first cases of evolving glyphosate resistance in UK poverty brome (Bromus sterilis) populations. Weed Sci. 2018, 67, 41–47. [Google Scholar] [CrossRef]
- Paterson, E.A.; Shenton, Z.L.; Straszewski, A.E. Establishment of the baseline sensitivity and monitoring response of Papaver rhoeas populations to florasulam. Pest Manag. Sci. 2002, 58, 964–966. [Google Scholar] [CrossRef] [PubMed]
- Russell, P.E. Sensitivity Baseline in Fungicide Resistance Research And Management; Crop Life International: Brussels, Belgium, 2002; pp. 1–56. [Google Scholar]
- Russell, P. Resistance management and the registration of pesticide products in Europe. Pestic. Outlook 2001, 12, 56–59. [Google Scholar] [CrossRef]
- Barroso, J.; Loureiro, I.; Escorial, M.C.; Chueca, M.C. The response of Bromus diandrus and Lolium rigidum to dalapon and glyphosate I: Baseline sensitivity. Weed Res. 2010, 50, 312–319. [Google Scholar]
- Moss, S.R. Baseline sensitivity to herbicides: A guideline to methodologies. In Proceedings of the British Crop Protection Conference—Weeds, Brighton, UK, 2001; pp. 769–774. [Google Scholar]
- Collavo, A.; Sattin, M. First glyphosate-resistant Lolium spp. biotypes found in a European annual arable cropping system also affected by ACCase and ALS resistance. Weed Res. 2014, 54, 325–334. [Google Scholar] [CrossRef]
- Dayan, F.E. Current status and future prospects in herbicide discovery. Plants 2019, 8, 341. [Google Scholar] [CrossRef]
- Sammons, R.D.; Gaines, T.A. Glyphosate resistance: State of knowledge. Pest Manag. Sci. 2014, 70, 1367–1377. [Google Scholar] [CrossRef]
- Pedersen, B.P.; Neve, P.; Andreasen, C.; Powles, S.B. Ecological fitness of a glyphosate-resistant Lolium rigidum population: Growth and seed production along a competition gradient. Basic Appl. Ecol. 2007, 8, 258–268. [Google Scholar] [CrossRef]
- Hess, M.; Barralis, G.; Bleiholder, H.; Buhr, L.; Eggers, T.H.; Hack, H.; Stauss, R. Use of the extended BBCH scale-general for the descriptions of the growth stages of mono and dicotyledonous weed species. Weed Res. 1997, 37, 433–441. [Google Scholar] [CrossRef]
- Beckie, H.J.; Ashworth, M.B.; Flower, K.C. Herbicide resistance management: Recent developments and trends. Plants 2019, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Onofri, A. Bioassay97: A new Excel VBA macro to perform statistical analyses on herbicide dose-response data. Riv. Ital. di Agrometeorol. 2005, 3, 40–45. [Google Scholar]
- Tukey, J.W. Exploratory data analysis. In The Future of Data Analysis; Addison-Wesley: New York, NY, USA, 1977; p. 688. [Google Scholar]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R package for determining the relevant number of clusters in a data set. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Population Code | Origin (Municipality) | Crop or Collection Site | Notes | ||
|---|---|---|---|---|---|---|
| (a) | ||||||
| LOL | RI | 07 | 328 | Santo Stefano Belbo | vineyard | Partially R [19] |
| LOL | RI | 08 | 204L | Legnaro | wheat | S check used by IPSP-CNR |
| LOL | SS | 08 | 259 | Cortona | wheat | Partially R [31] |
| LOL | SS | 08 | 340 | Collesalvetti | wheat | |
| LOL | SS | 08 | 343 | Pomarance | wheat | R pop. [31] |
| LOL | SS | 10 | 381 | Pontedera | wheat | |
| LOL | SS | 10 | 384 | Cascina | wheat | R pop. [31] |
| LOL | SS | 11 | 384L | Cascina | Reproduced from glyphosate-resistant plants of pop. 10–384 | |
| LOL | SS | 10 | 389 | Castenaso | meadow | |
| LOL | MU | 11 | 390 | Legnaro | field margin | |
| LOL | SS | 11 | 392 | Palo del Colle | olive grove | R pop. (unpublished data) |
| LOL | RI | 11 | 395 | Acquaviva delle Fonti | roadside | |
| LOL | RI | 11 | 400 | Torchiarolo | set aside | |
| LOL | SS | 11 | 401 | Lamezia Terme | olive grove | R pop. (unpublished data) |
| LOL | RI | 11 | 402 | Lamezia Terme | meadow | |
| LOL | SS | 11 | 403 | Cascina | sunflower | R pop. [31] |
| LOL | RI | 11 | 404 | Lamezia Terme | olive grove | |
| LOL | SS | 11 | 405 | Livorno | lucerne | |
| LOL | MU | 11 | 412 | Commercial turf seed | ||
| LOL | SS | 11 | 425 | Cascina | wheat | |
| (b) | ||||||
| LOL | MU | 12 | 426 | Ravenna | wheat | |
| LOL | SS | 12 | 431 | Duino Aurisina | meadow | |
| LOL | SS | 12 | 432 | Siena | wheat | |
| LOL | SS | 12 | 434 | Sovicille | wheat | |
| LOL | SS | 12 | 444 | San Casciano dei Bagni | wheat | |
| LOL | MU | 12 | 449 | Marsciano | vineyard | |
| LOL | SS | 12 | 455 | Gubbio | wheat | |
| LOL | MU | 12 | 458 | Montecchio Emilia | lucerne | |
| LOL | SS | 12 | 461 | Brisighella | roadside | |
| LOL | MU | 12 | 462 | Forlì | roadside | |
| LOL | MU | 12 | 466 | Coriano | wheat | |
| LOL | SS | 12 | 472 | Osimo | wheat | |
| LOL | SS | 12 | 477 | Pozzolengo | meadow | |
| LOL | SS | 12 | 479 | Cremona | field margin (maize) | |
| LOL | MU | 12 | 483 | Alessandria | meadow | |
| LOL | SS | 12 | 487 | Saluzzo | meadow | |
| LOL | SS | 12 | 492 | Cigliano | field margin | |
| LOL | SS | 12 | 504 | Pontoglio | roadside | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panozzo, S.; Collavo, A.; Sattin, M. Sensitivity Analysis of Italian Lolium spp. to Glyphosate in Agricultural Environments. Plants 2020, 9, 165. https://doi.org/10.3390/plants9020165
Panozzo S, Collavo A, Sattin M. Sensitivity Analysis of Italian Lolium spp. to Glyphosate in Agricultural Environments. Plants. 2020; 9(2):165. https://doi.org/10.3390/plants9020165
Chicago/Turabian StylePanozzo, Silvia, Alberto Collavo, and Maurizio Sattin. 2020. "Sensitivity Analysis of Italian Lolium spp. to Glyphosate in Agricultural Environments" Plants 9, no. 2: 165. https://doi.org/10.3390/plants9020165
APA StylePanozzo, S., Collavo, A., & Sattin, M. (2020). Sensitivity Analysis of Italian Lolium spp. to Glyphosate in Agricultural Environments. Plants, 9(2), 165. https://doi.org/10.3390/plants9020165