Two Self-Incompatibility Sites Occur Simultaneously in the Same Acianthera Species (Orchidaceae, Pleurothallidinae)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
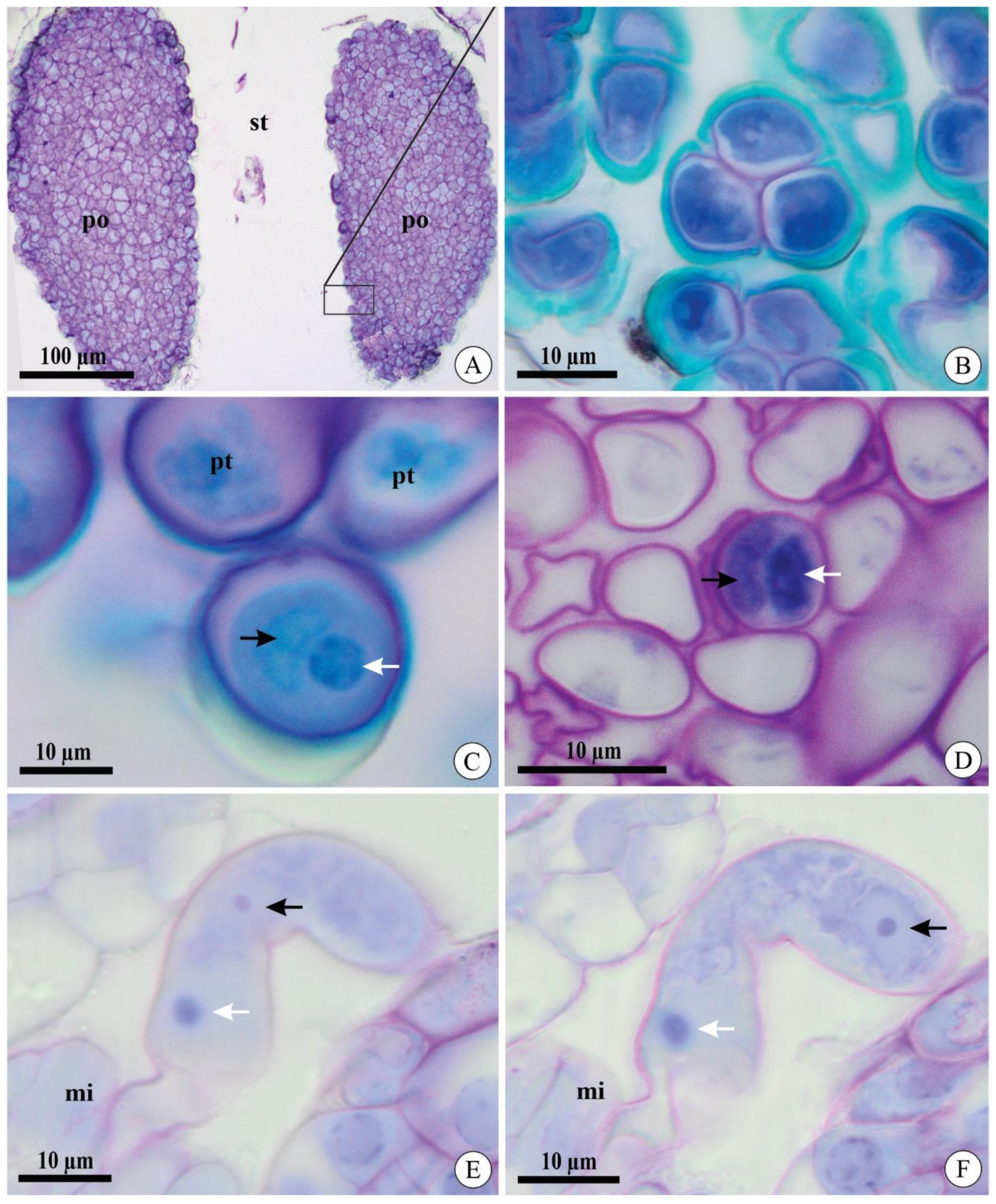
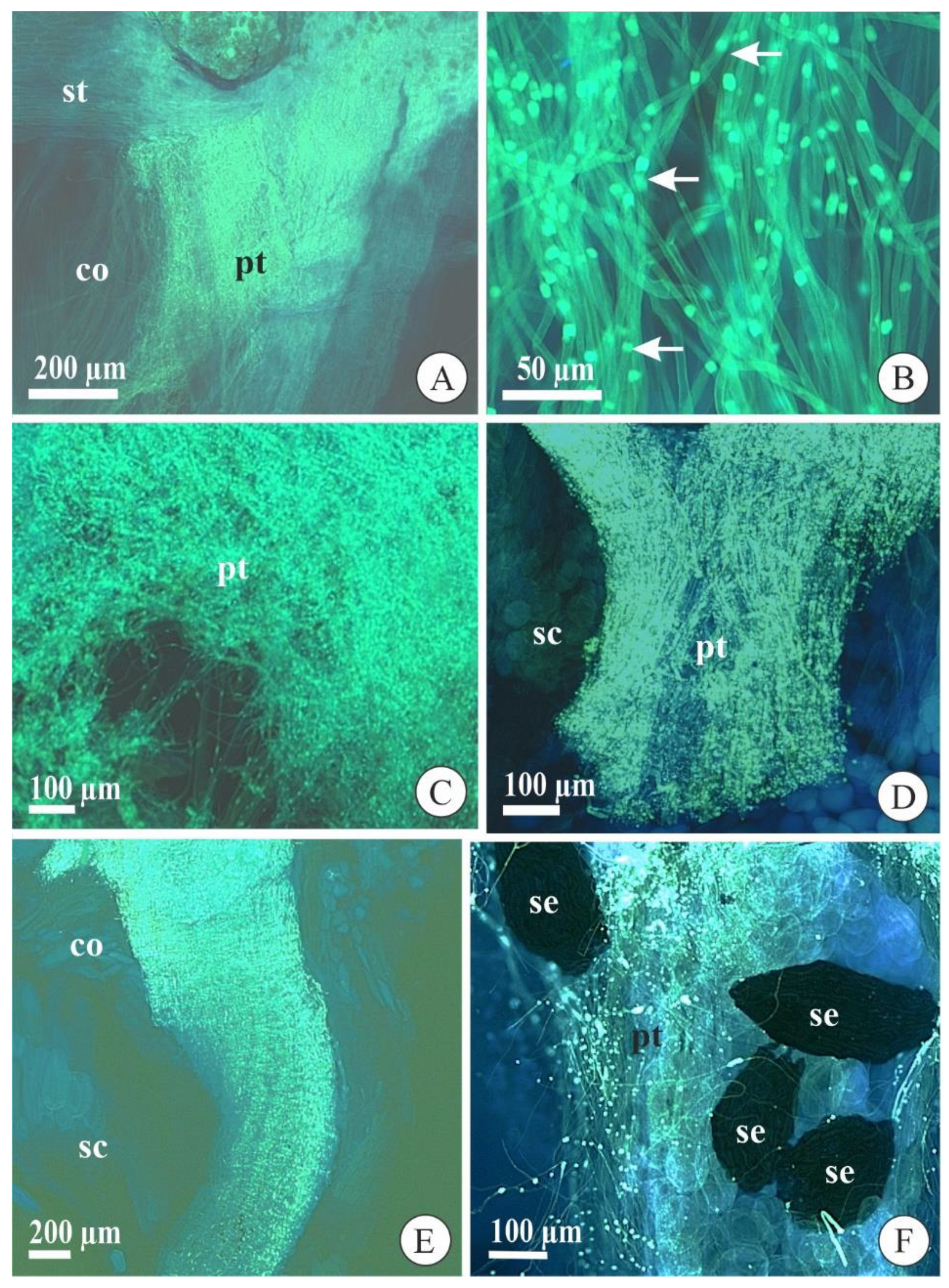
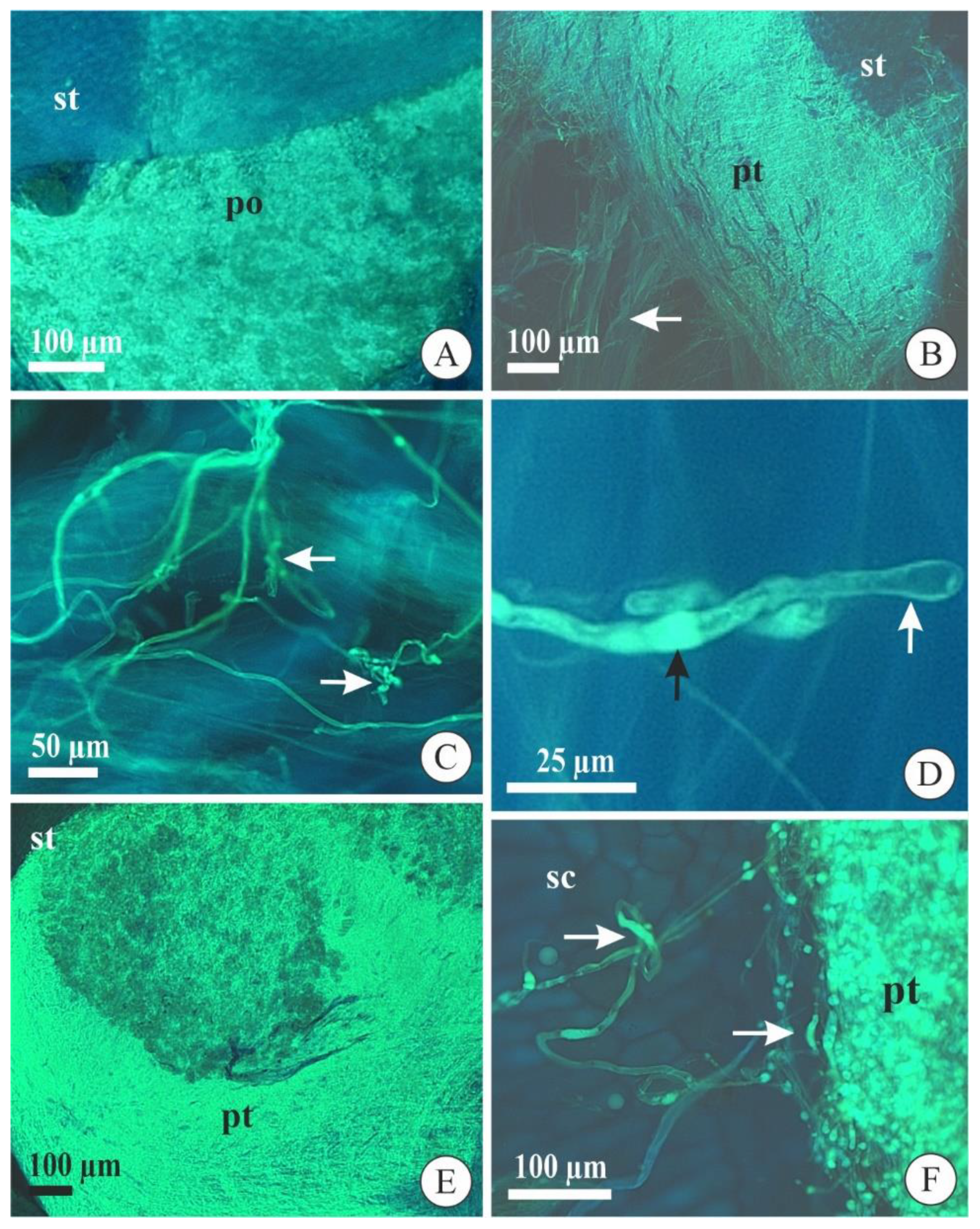
2.1. Pollen Tube Development after Cross-Pollination
2.2. Pollen Tube Development after Self-Pollination
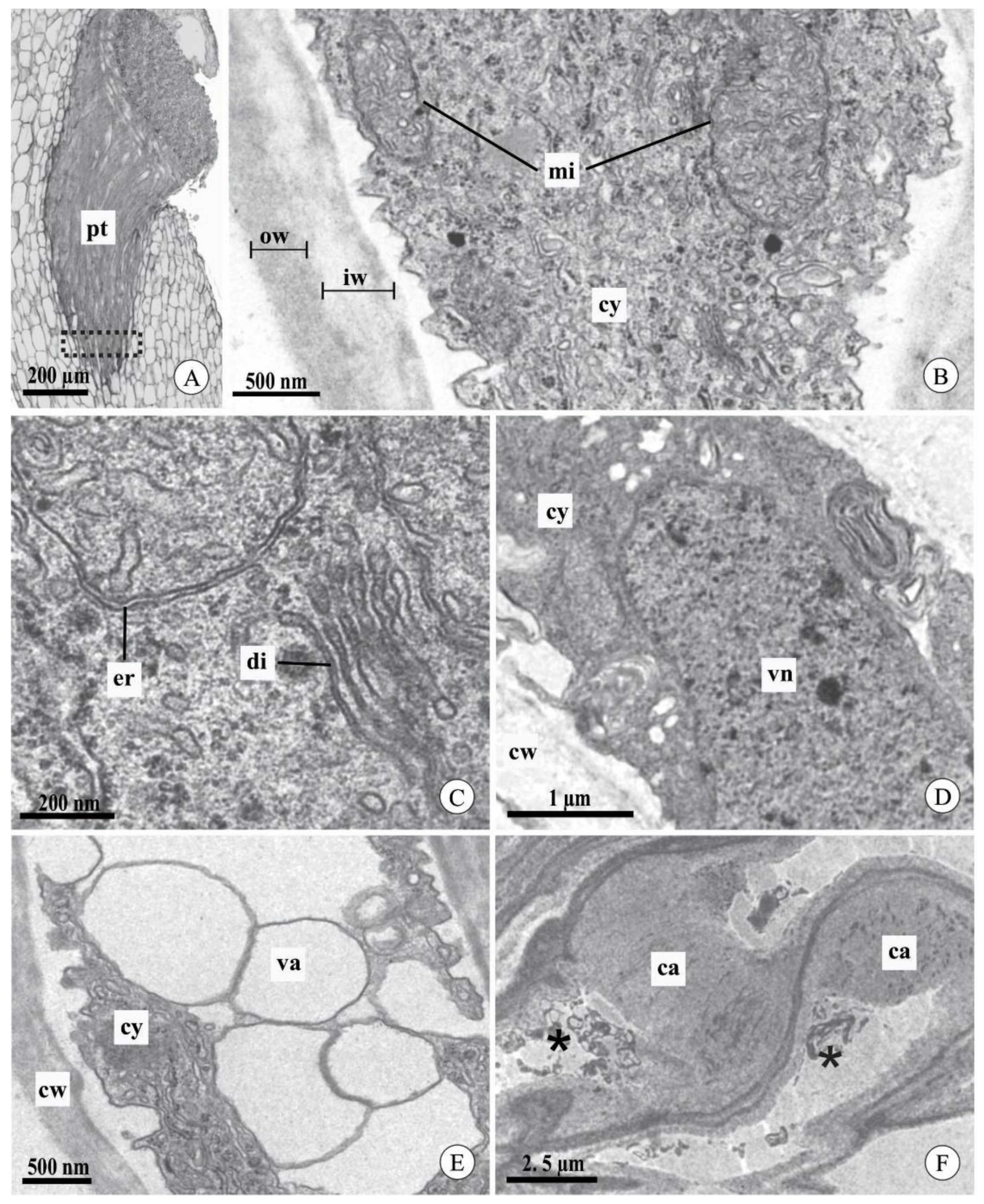
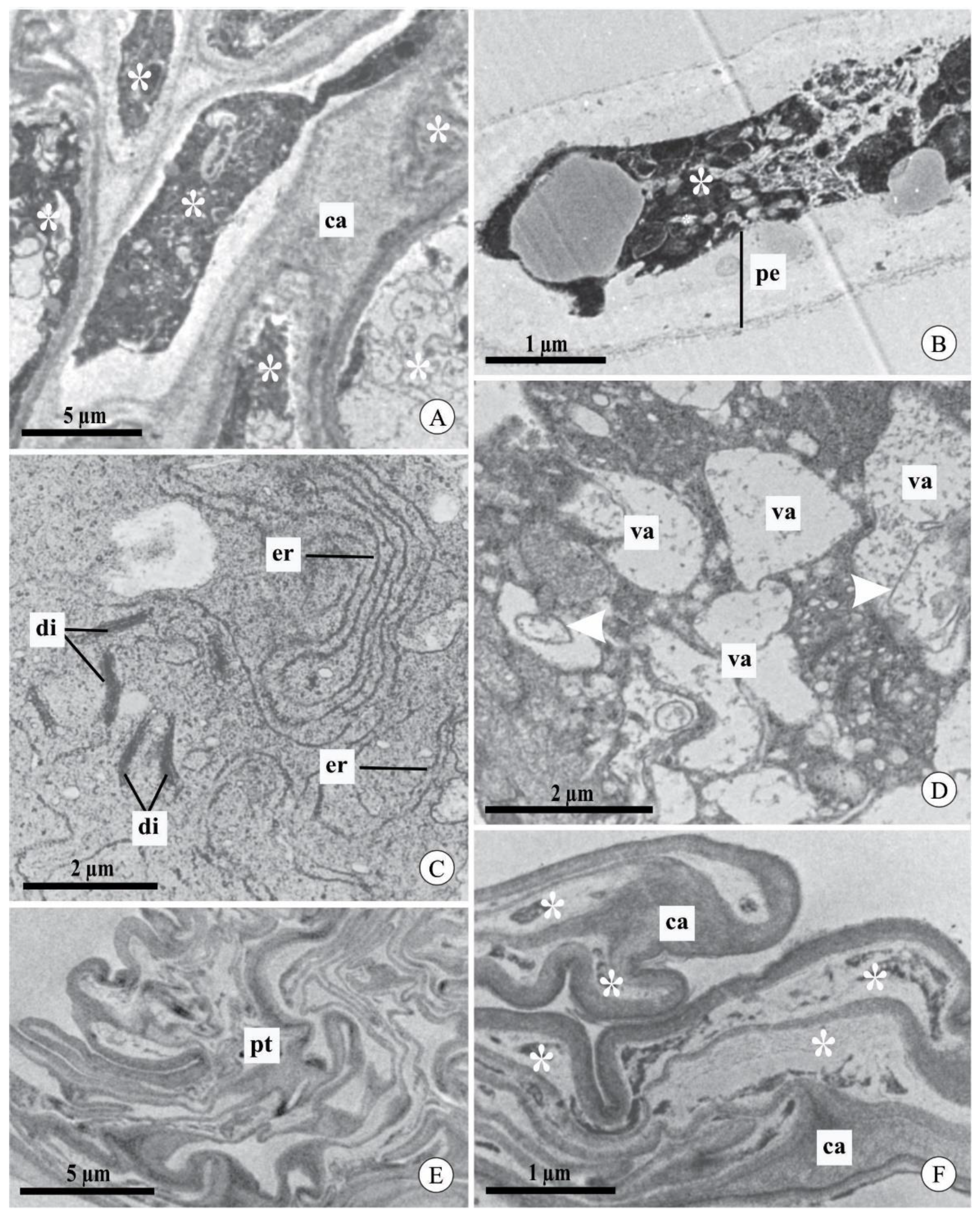
2.3. Pollen Tube Ultrastructure after Experimental Pollination
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Nettancourt, D. Incompatibility in angiosperms. Sex. Plant. Reprod. 1997, 10, 185–199. [Google Scholar] [CrossRef]
- Richards, A.J. Plant Breeding Systems; Chapman & Hall: London, UK, 1997. [Google Scholar]
- Allen, A.M.; Hiscock, S.J. Evolution and phylogeny of self-incompatibility systems in angiosperms. In Self-Incompatibility in Flowering Plants: Evolution, Diversity, and Mechanisms; Franklin-Tong, V.E., Ed.; Springer: Berlin, Germany, 2008; pp. 73–101. [Google Scholar]
- Seavey, S.F.; Bawa, K.S. Late-acting self-incompatibility in angiosperms. Bot. Rev. 1986, 52, 195–218. [Google Scholar] [CrossRef]
- Gibbs, P.E. Late-acting self-incompatibility—The pariah breeding system in flowering plants. New Phytol. 2014, 203, 717–734. [Google Scholar] [CrossRef]
- Johansen, B. Incompatibility in Dendrobium (Orchidaceae). Bot. J. Linn. Soc. 1990, 103, 165–196. [Google Scholar] [CrossRef]
- Castro, J.B.; Singer, R.B. A literature review of the pollination strategies and breeding systems in Oncidiinae orchids. Acta Bot. Bras. 2019, 33, 618–643. [Google Scholar] [CrossRef]
- Borba, E.L.; Barbosa, A.R.; Melo, M.C.; Gontijo, S.L.; Oliveira, H.O. Mating systems in the Pleurothallidinae (Orchidaceae): Evolutionary and systematic implications. Lankesteriana 2011, 11, 207–221. [Google Scholar]
- Borba, E.L.; Semir, J.; Shepherd, G.J. Self-incompatibility, inbreeding depression, and crossing potential in five Brazilian Pleurothallis (Orchidaceae) species. Ann. Bot. 2001, 88, 89–99. [Google Scholar] [CrossRef]
- Barbosa, A.R.; Melo, M.C.; Borba, E.L. Self-incompatibility and myophily in Octomeria (Orchidaceae, Pleurothallidinae) species. Plant. Syst. Evol. 2009, 283, 1–8. [Google Scholar] [CrossRef]
- Gontijo, S.L.; Barbosa, A.R.; Melo, M.C.; Borba, E.L. Occurrence of different sites of self-incompatibility reaction in four Anathallis (Orchidaceae, Pleurothallidinae) species. Plant. Spec. Biol. 2010, 25, 129–135. [Google Scholar] [CrossRef]
- Melo, M.C.; Taucce, P.P.G.; Borba, E.L. Reproductive biology and isolation mechanisms in rupicolous species of the Acianthera prolifera complex (Orchidaceae) occurring in southeastern Brazil. Plant. Syst. Evol. 2011, 293, 161–176. [Google Scholar] [CrossRef]
- Dressler, R.L. The Orchids: Natural History and Classification; Harvard University Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Luer, C.A. Icones Pleurothallidinarum, I. systematics of Pleurothallidinae. Monogr. Syst. Bot. Mo. Bot. Gard. 1986, 15, 1–81. [Google Scholar]
- Dressler, R.L. Phylogeny and Classification of the Orchid Family; Dioscorides Press: Portland, OR, USA, 1993. [Google Scholar]
- Christensen, D.E. Notes on the reproductive biology of Stelis argentata Lindl. (Orchidaceae: Pleurothallidinae) in eastern Ecuador. Lindleyana 1992, 7, 28–33. [Google Scholar]
- Millner, H.J.; McCrea, A.R.; Baldwin, T.C. An investigation of self-incompatibility within the genus Restrepia. Am. J. Bot. 2015, 102, 487–494. [Google Scholar] [CrossRef]
- Bogarín, D.; Fernández, M.; Borkent, A.; Heemskerk, A.; Pupulin, F.; Ramírez, S.; Smets, E.; Gravendeel, B. Pollination of Trichosalpinx (Orchidaceae: Pleurothallidinae) by biting midges (Diptera: Ceratopogonidae). Bot. J. Linn. Soc. 2018, 186, 510–543. [Google Scholar] [CrossRef]
- Huda, M.K.; Wilcock, C.C. Rapid floral senescence following male function and breeding systems of some tropical orchids. Plant Biol. 2011, 14, 278–284. [Google Scholar] [CrossRef]
- São Leão, L.C.; Sá-Haiad, B.; Rodarte, A.T.A.; de Araujo Pimentel, R.R.; Benevides, C.R.; Santiago-Fernandes, L.D.R.; Lima, H.A. Reproductive biology of two synchronopatric neotropical species of Epidendrum (Orchidaceae). Flora 2019, 251, 95–104. [Google Scholar] [CrossRef]
- Charlesworth, D.; Charlesworth, B. Inbreeding depression and its evolutionary consequences. Ann. Rev. Ecol. Syst. 1987, 18, 237–268. [Google Scholar] [CrossRef]
- Hufford, K.M.; Hamrick, L.L. Viability selection at three early life stages of the tropical tree, Platypodium elegans (Fabaceae, Papilionoideae). Evolution 2003, 57, 518–526. [Google Scholar] [CrossRef]
- Arditti, J. Aspects of the physiology of orchids. Adv. Bot. Res. 1979, 7, 421–655. [Google Scholar]
- Avadhani, P.N.; Nair, H.; Arditti, J.; Hew, C.S. Physiology of orchid flower. In Orchid Biology, Review and Perspectives, 6th ed.; Arditti, J., Ed.; Wiley-Interscience: New York, NY, USA, 1994; pp. 189–362. [Google Scholar]
- Sage, T.L.; Bertin, R.; Williams, E.G. Ovarian and other late-acting self-incompatibility. In Genetic Control of Self-Incompatibility and Reproductive Development in Flowering Plants; Williams, E.G., Clark, A.E., Knox, R.B., Eds.; Kluwer: Dordrecht, The Netherlands, 1994; Volume 2, pp. 116–140. [Google Scholar]
- Zhang, C.C.; Wang, L.Y.; Wei, K.; Wu, L.Y.; Li, H.L.; Zhang, F.; Cheng, H.; Ni, D.J. Transcriptome analysis reveals self-incompatibility in the tea plant (Camellia sinensis) might be under gametophytic control. BMC Genom. 2016, 17, 359. [Google Scholar] [CrossRef]
- Duarte, M.O. Sementes abortadas ou óvulos não fecundados? Investigação da possível ocorrência de dois sítios de autoincompatibilidade em Acianthera (Orchidaceae). Ph.D. Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil, February 2020. [Google Scholar]
- Hao, Y.Q.; Zhao, X.F.; She, D.Y.; Xu, B.; Zhang, D.Y.; Liao, W.J. The role of late-acting self-incompatibility and early-acting inbreeding depression in governing female fertility in monkshood, Aconitum kusnezoffii. PLoS ONE 2012, 7, e4703. [Google Scholar] [CrossRef]
- Xiong, H.; Zou, F.; Guo, S.; Yuan, D.; Niu, G. Self-sterility may be due to prezygotic late-acting self-incompatibility and early-acting inbreeding depression in Chinese chestnut. J. Am. Soc. Hort. Sci. 2019, 144, 172–181. [Google Scholar] [CrossRef]
- Swanson, R.; Edlund, A.F.; Preuss, D. Species specificity in pollen-pistil interactions. Annu. Rev. Genet. 2004, 38, 793–818. [Google Scholar] [CrossRef]
- Chebli, Y.; Geitmann, A. Mechanical principles governing pollen tube growth. Funct. Plant. Sci. Biotechnol. 2007, 2, 232–245. [Google Scholar]
- Dardelle, F.; Lehner, A.; Ramdani, Y.; Bardor, M.; Lerouge, P.; Driouich, A.; Mollet, J.C. Biochemical and immunocytological characterizations of Arabidopsis pollen tube cell wall. Plant. Physiol. 2010, 153, 1563–1576. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K.; Kunkel, J.G.; Rounds, C.M.; Winship, L.J. Calcium entry into pollen tubes. Trends Plant. Sci. 2012, 17, 32–38. [Google Scholar] [CrossRef]
- Herrero, M.; Dickinson, H.G. Pollen tube development in Petunia hybrida following compatible and incompatible intraspecific matings. J. Cell Sci. 1981, 47, 365–383. [Google Scholar] [PubMed]
- Geitmann, A.; Franklin-Tong, V.E.; Emons, A.C. The self-incompatibility response in Papaver rhoeas pollen causes early and striking alterations to organelles. Cell Death Differ. 2004, 11, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, L.V.; Zakharova, E.V.; Timofeeva, G.V.; Andreev, I.M.; Golivanov, Y.Y.; Bogoutdinova, L.R.; Baranova, E.N.; Khaliluev, M.R. Aminooxyacetic acid (AOA), inhibitor of 1-aminocyclopropane-1-carboxilic acid (ACC) synthesis, suppresses self-incompatibility-induced programmed cell death in self-incompatible Petunia hybrida L. pollen tubes. Protoplasma 2020, 257, 213–227. [Google Scholar] [CrossRef] [PubMed]
- van Doorn, W.G.; Beers, E.P.; Dangl, J.L.; Franklin-Tong, V.E.; Gallois, P.; Hara-Nishimura, I.; Jones, A.M.; Kawai-Yamada, M.; Lam, E.; Mundy, J.; et al. Morphological classification of plant cell deaths. Cell Death Differ. 2011, 18, 1241–1246. [Google Scholar] [CrossRef]
- Franklin-Tong, V.; Franklin, F.C.H. The different mechanisms of gametophytic self-incompatibility. Philos. Trans. R. Soc. B 2003, 358, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Sijacic, P.; Wang, X.; Skirpan, A.L.; Wang, Y.; Dowd, P.E.; McCubbin, A.G.; Huang, S.; Kao, T.H. Identification of the pollen determinant of S-RNase-mediated self-incompatibility. Nature 2004, 429, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Bosch, M.; Franklin-Tong, V.E. Self-incompatibility in Papaver: Signaling to trigger PCD in incompatible pollen. J. Exp. Bot. 2008, 59, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.C.; Huang, J.; Zhang, Y.Q.; Li, P.X.; Zhang, G.Q.; Xu, Q.; Chen, L.J.; Wang, J.Y.; Luo, Y.B.; Liu, Z.J. Lack of S-RNase-based gametophytic self-incompatibility in orchids suggests that this system evolved after the monocot-eudicot split. Front. Plant. Sci. 2017, 8, 1106. [Google Scholar] [CrossRef]
- Niu, S.C.; Huang, J.; Xu, Q.; Li, P.X.; Yang, H.J.; Zhang, Y.Q.; Zhang, G.Q.; Chen, L.J.; Niu, Y.X.; Luo, Y.B. Morphological type identification of self-incompatibility in Dendrobium and its phylogenetic evolution pattern. Int. J. Mol. Sci. 2018, 19, 2595. [Google Scholar] [CrossRef]
- Borba, E.L.; Semir, J. Pollinator specificity and convergence in fly-pollinated Pleurothallis (Orchidaceae) species: A multiple population approach. Ann. Bot. 2001, 88, 75–88. [Google Scholar] [CrossRef]
- Duarte, M.O.; Oliveira, D.M.T.; Borba, E.L. Ontogenesis of ovary and fruit of Acianthera johannensis (Pleurothallidinae, Orchidaceae) reveals a particular female embryology. Flora 2019, 259, 151462. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Co.: New York, NY, USA, 1940. [Google Scholar]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Martin, F.W. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technol. 1959, 34, 125–128. [Google Scholar] [CrossRef]
- Karnovsky, M.J. A formaldehyde-glutaraldehyde fixative of light osmolality for use in electron microscopy. J. Cell Biol. 1965, 27, 137A–138A. [Google Scholar]
- Roland, J.C. General preparations and staining of thin sections. In Electron Microscopy of Plant Cells; Hall, J.L., Ed.; Elsevier: New York, NY, USA, 1978; pp. 1–62. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte, M.O.; Oliveira, D.M.T.; Borba, E.L. Two Self-Incompatibility Sites Occur Simultaneously in the Same Acianthera Species (Orchidaceae, Pleurothallidinae). Plants 2020, 9, 1758. https://doi.org/10.3390/plants9121758
Duarte MO, Oliveira DMT, Borba EL. Two Self-Incompatibility Sites Occur Simultaneously in the Same Acianthera Species (Orchidaceae, Pleurothallidinae). Plants. 2020; 9(12):1758. https://doi.org/10.3390/plants9121758
Chicago/Turabian StyleDuarte, Mariana Oliveira, Denise Maria Trombert Oliveira, and Eduardo Leite Borba. 2020. "Two Self-Incompatibility Sites Occur Simultaneously in the Same Acianthera Species (Orchidaceae, Pleurothallidinae)" Plants 9, no. 12: 1758. https://doi.org/10.3390/plants9121758
APA StyleDuarte, M. O., Oliveira, D. M. T., & Borba, E. L. (2020). Two Self-Incompatibility Sites Occur Simultaneously in the Same Acianthera Species (Orchidaceae, Pleurothallidinae). Plants, 9(12), 1758. https://doi.org/10.3390/plants9121758